
Hericium erinaceus and Coriolus versicolor Modulate Molecular and Biochemical Changes after Traumatic Brain Injury

 , , , , , , , , , and
, , , , , , , , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Controlled Cortical Impact (CCI) Experimental TBI
Experimental Design
2.3. Chemical Reagents
2.4. Sample Pretreatment
2.5. Extraction Method
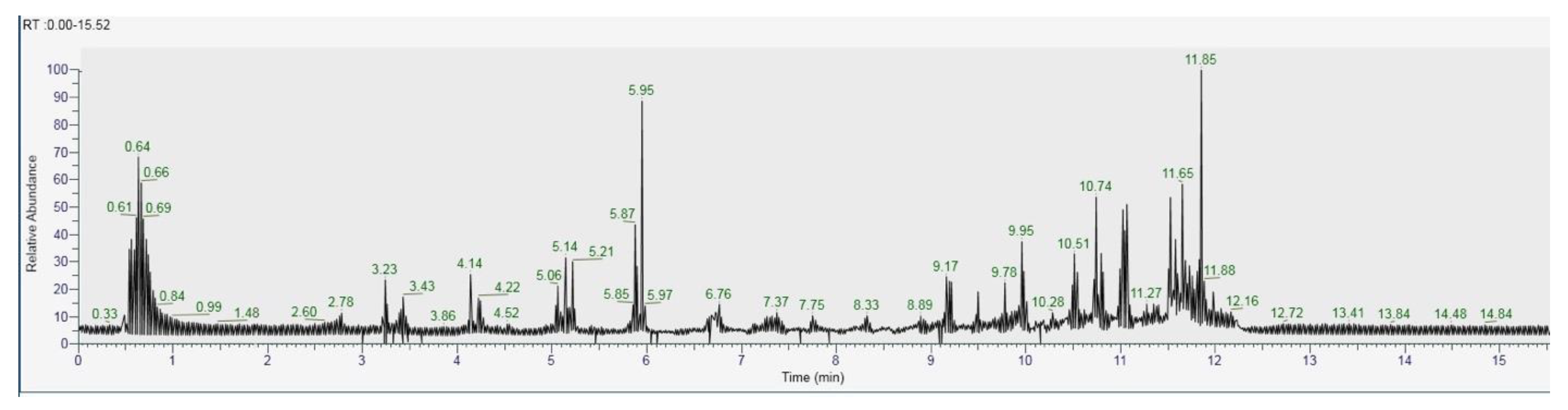
2.6. GC-MS/MS
2.7. LC–Orbitrap-MS
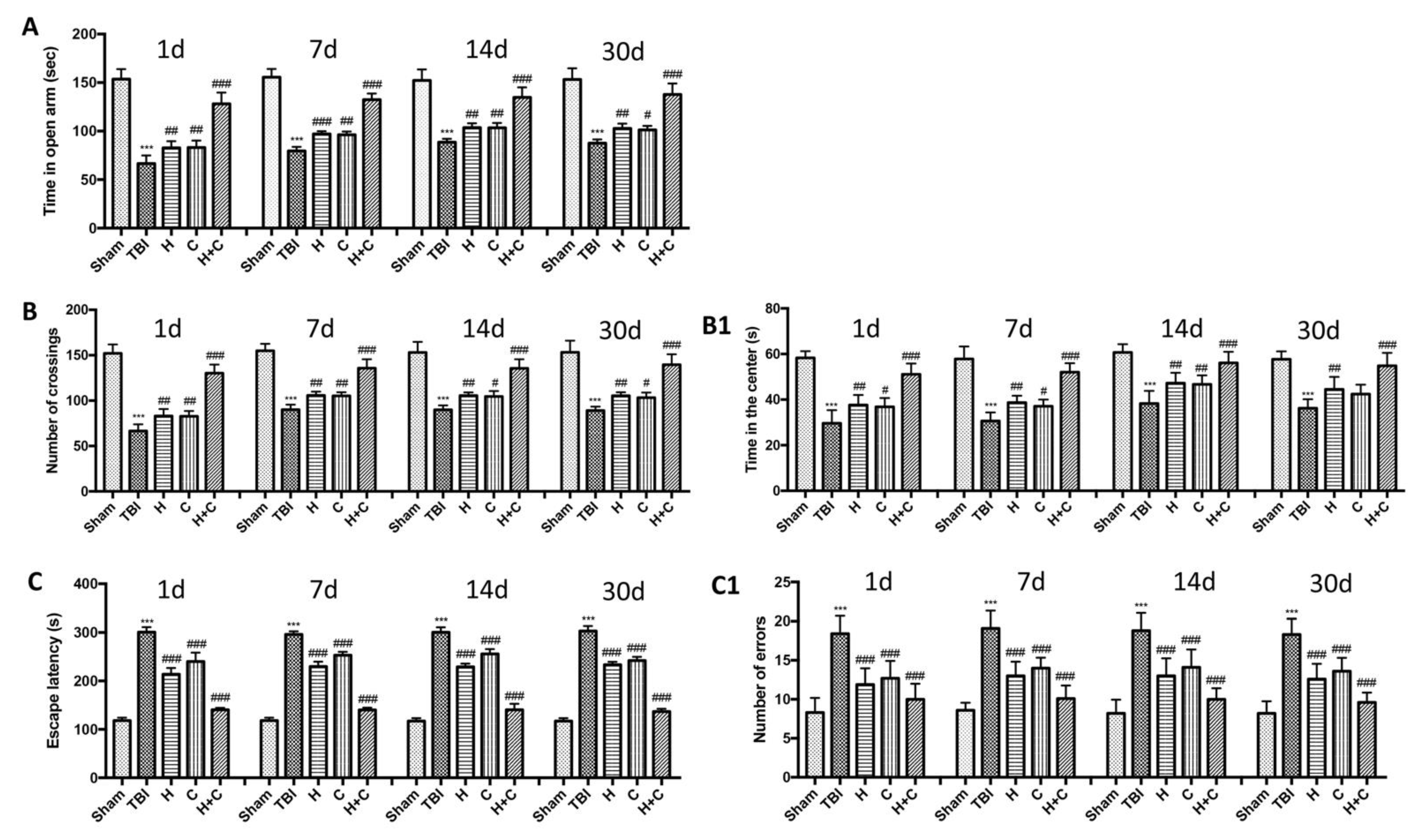
2.8. Behavioral Testing
2.8.1. Elevated Pluz-Maze (EPM)
2.8.2. Open Field Test (OF)
2.8.3. Barnes Maze
2.9. Histology
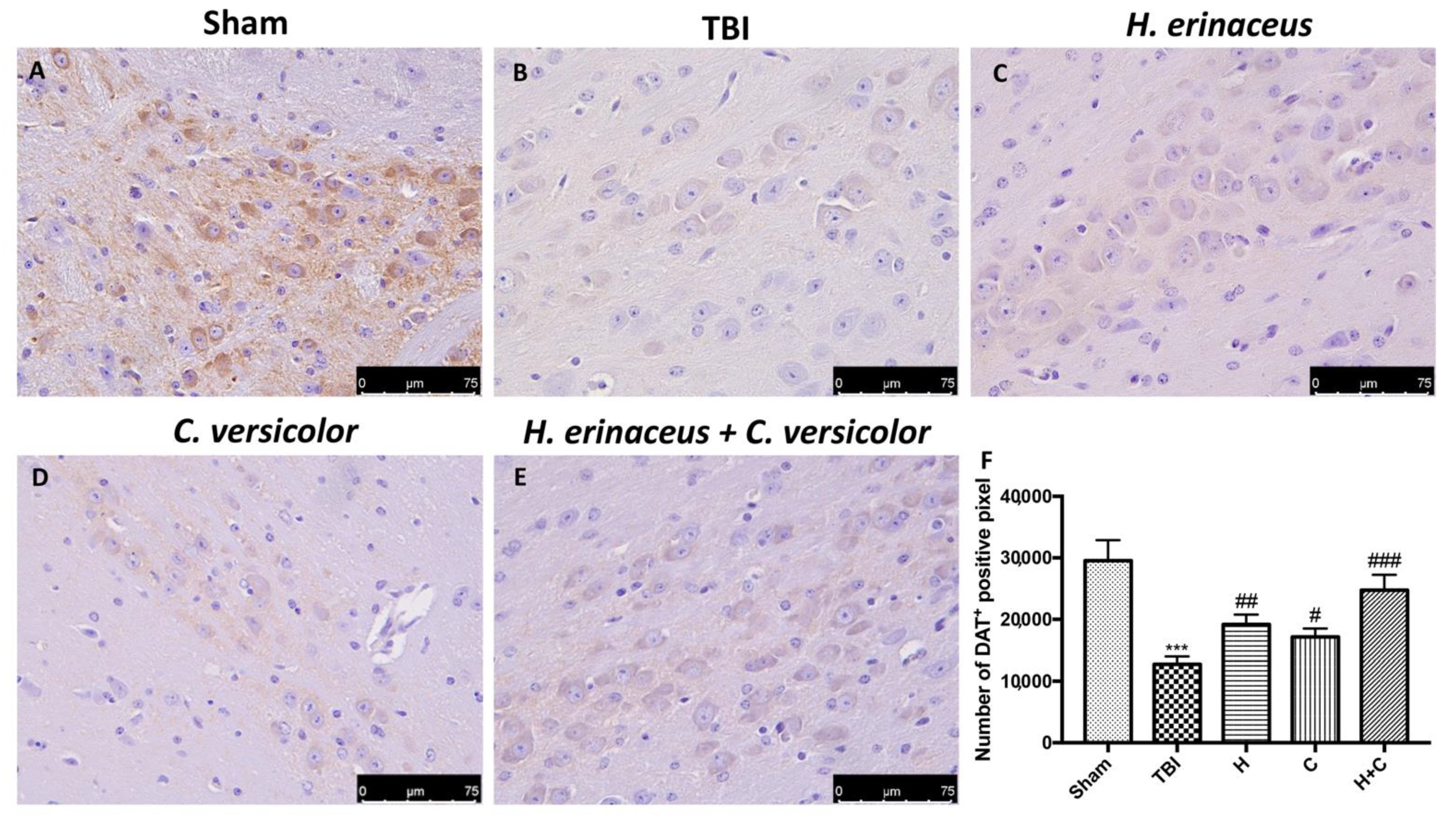
2.10. Immunohistochemical Analysis of Tyrosine Hydroxylase (TH), Dopamine Transporter (DAT)
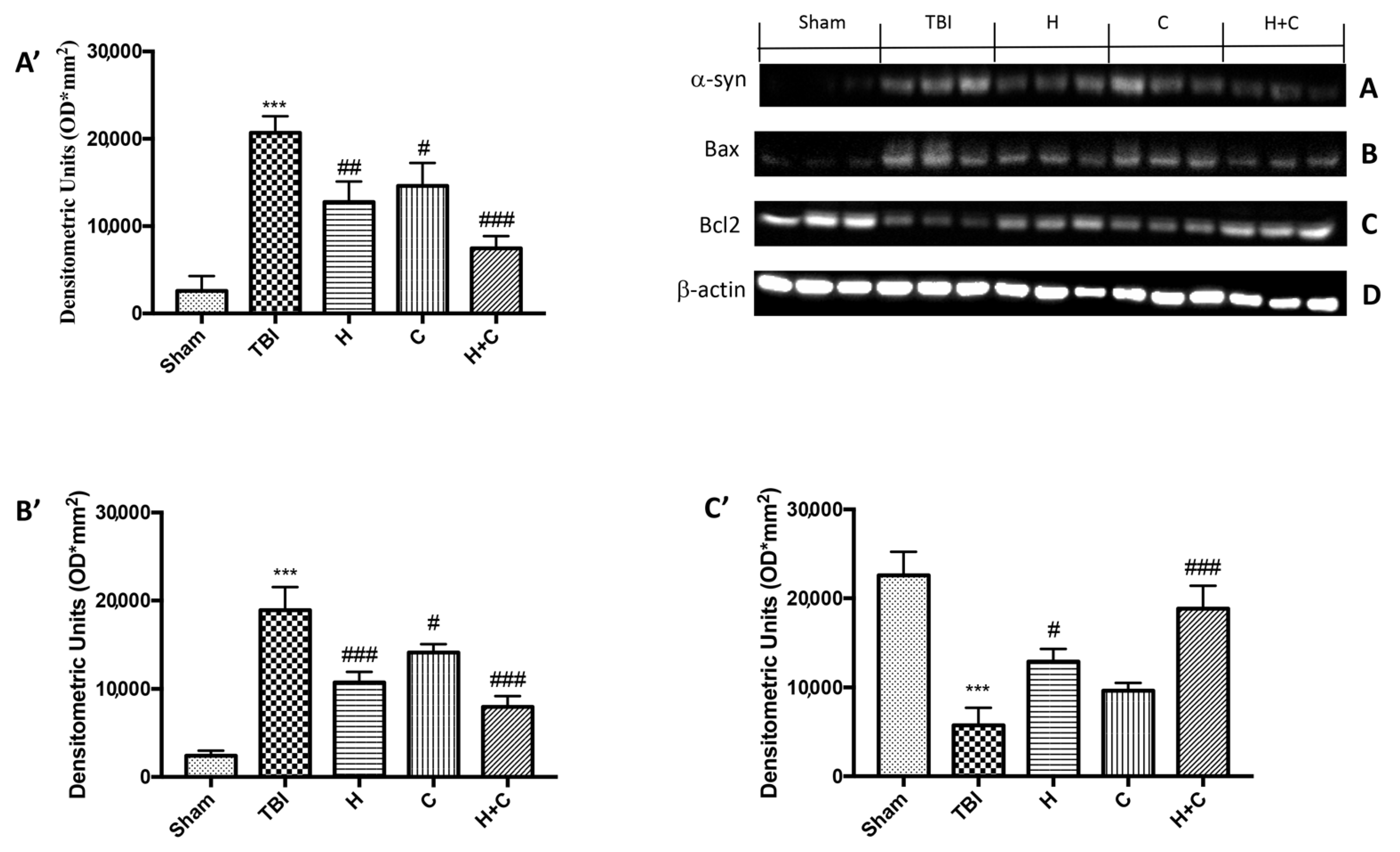
2.11. Western Blot Analysis for GFAP, Iba-1, IkB-α, NF-kB, Nrf2, HO-1, Hsp70, γ-GCs, Trx, α-syn, Bax, Bcl-2
2.12. Statistical Evaluation
3. Results
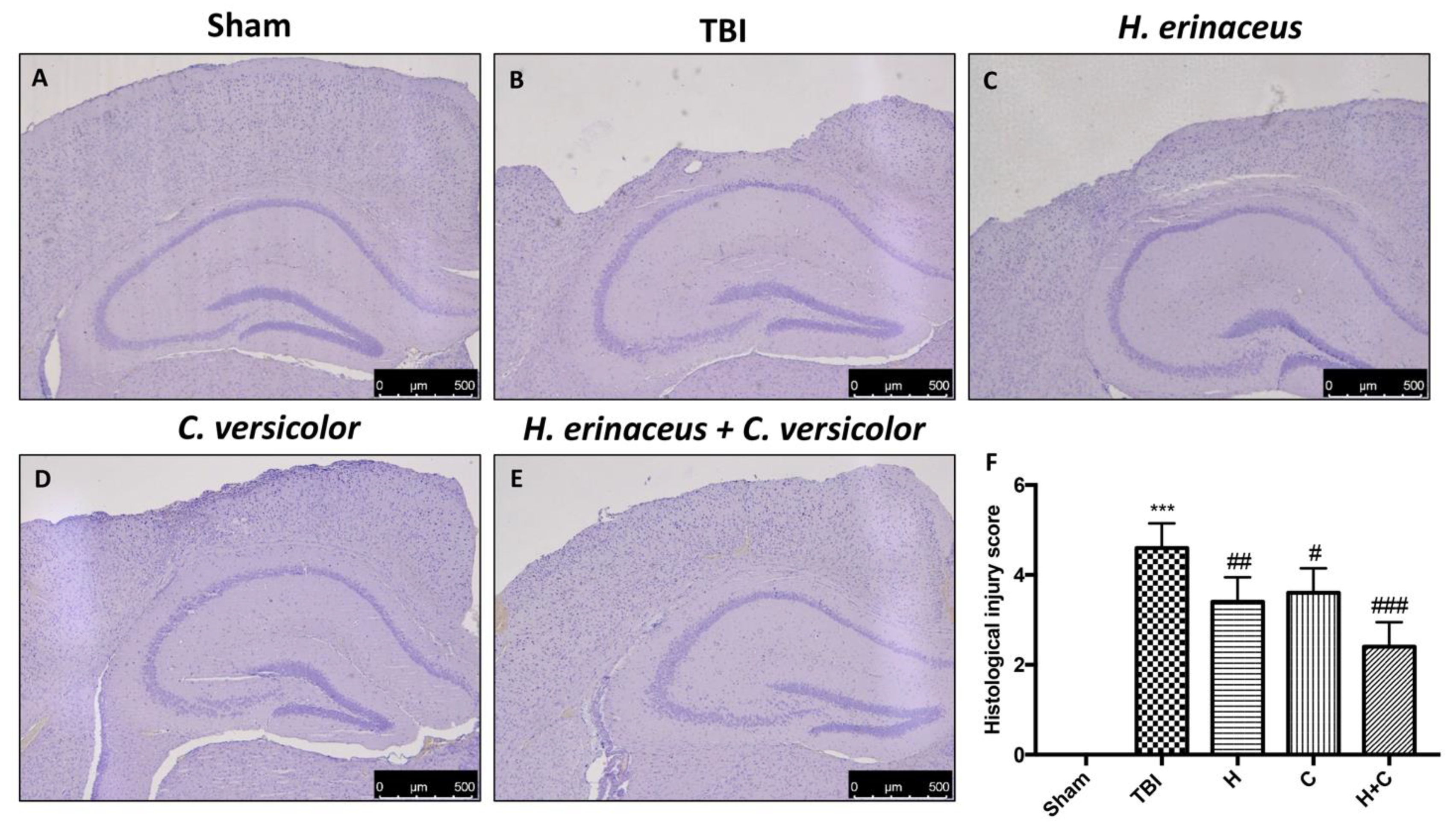
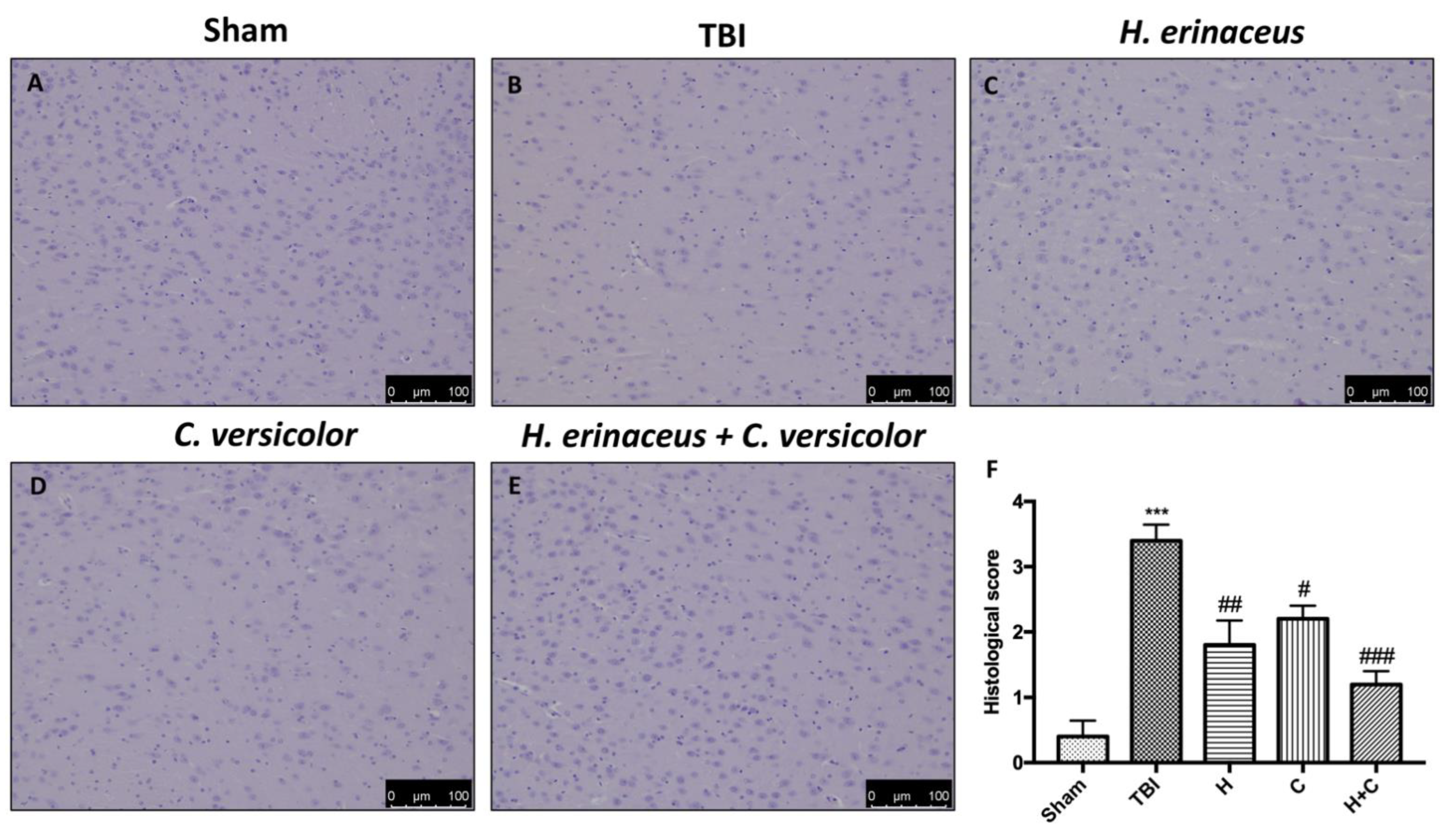
3.1. Effect of H. erinaceus or C. versicolor, or H. erinaceus Plus C. versicolor Treatment on Histological Changes TBI-Induced
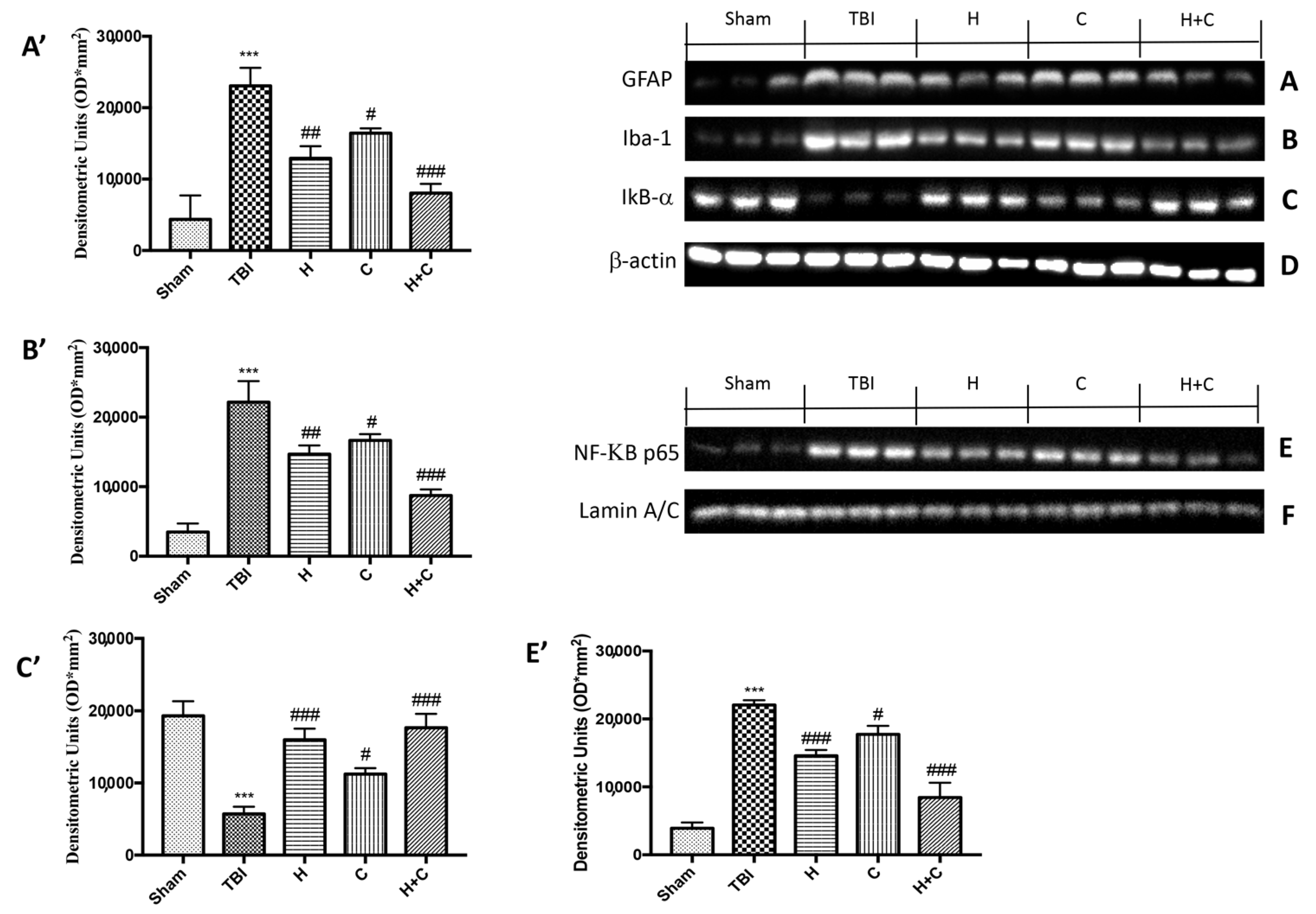
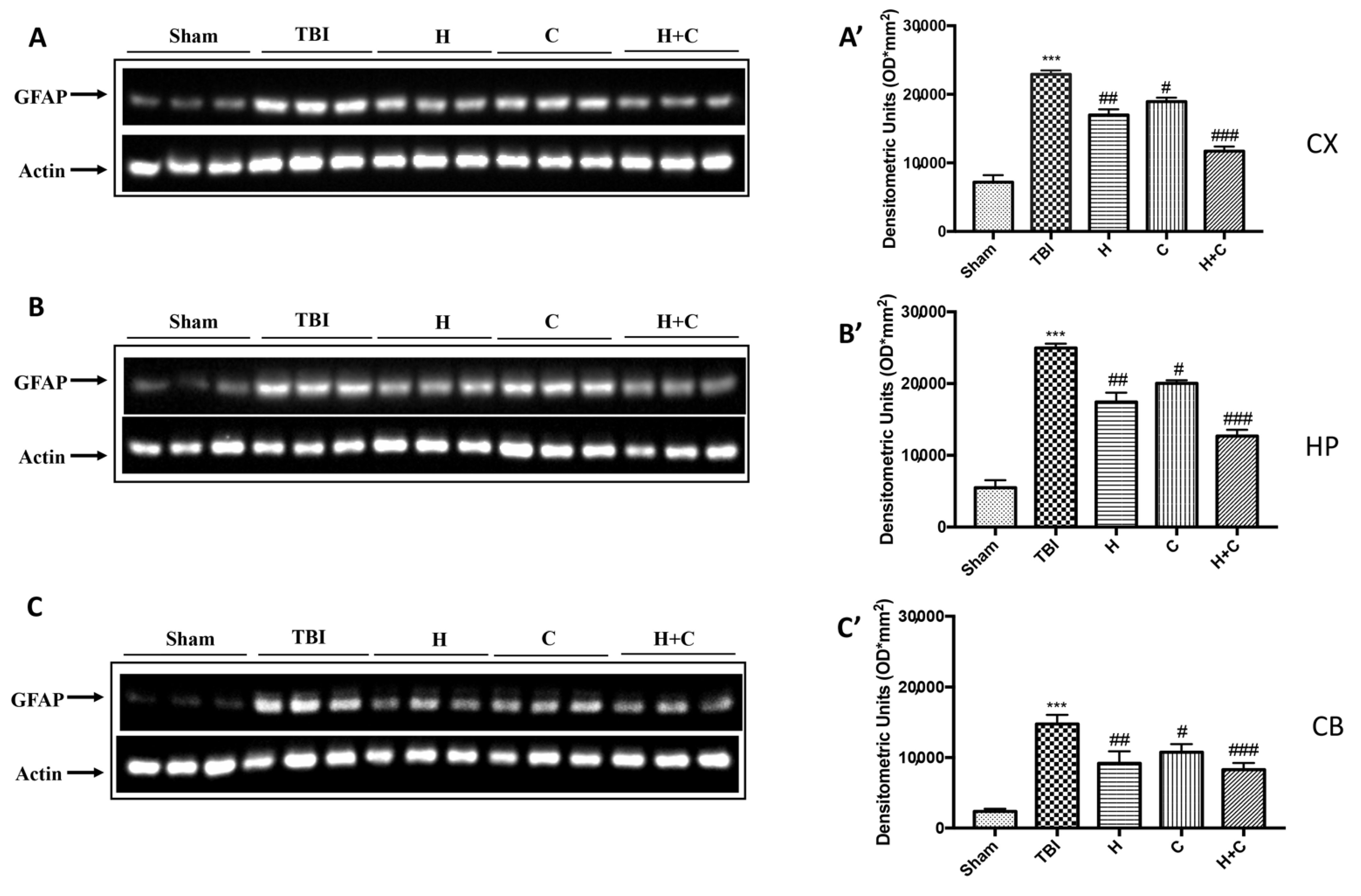
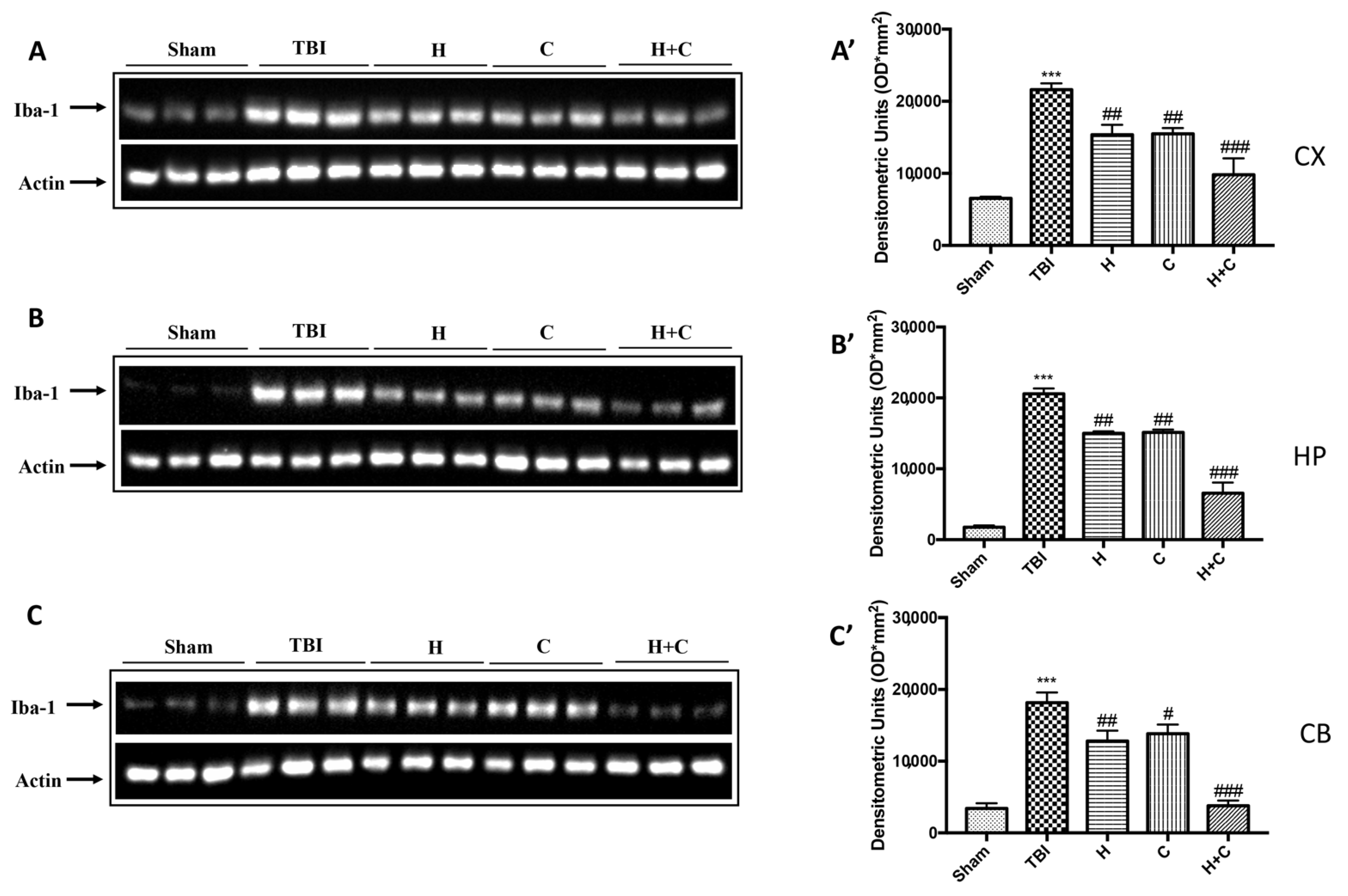
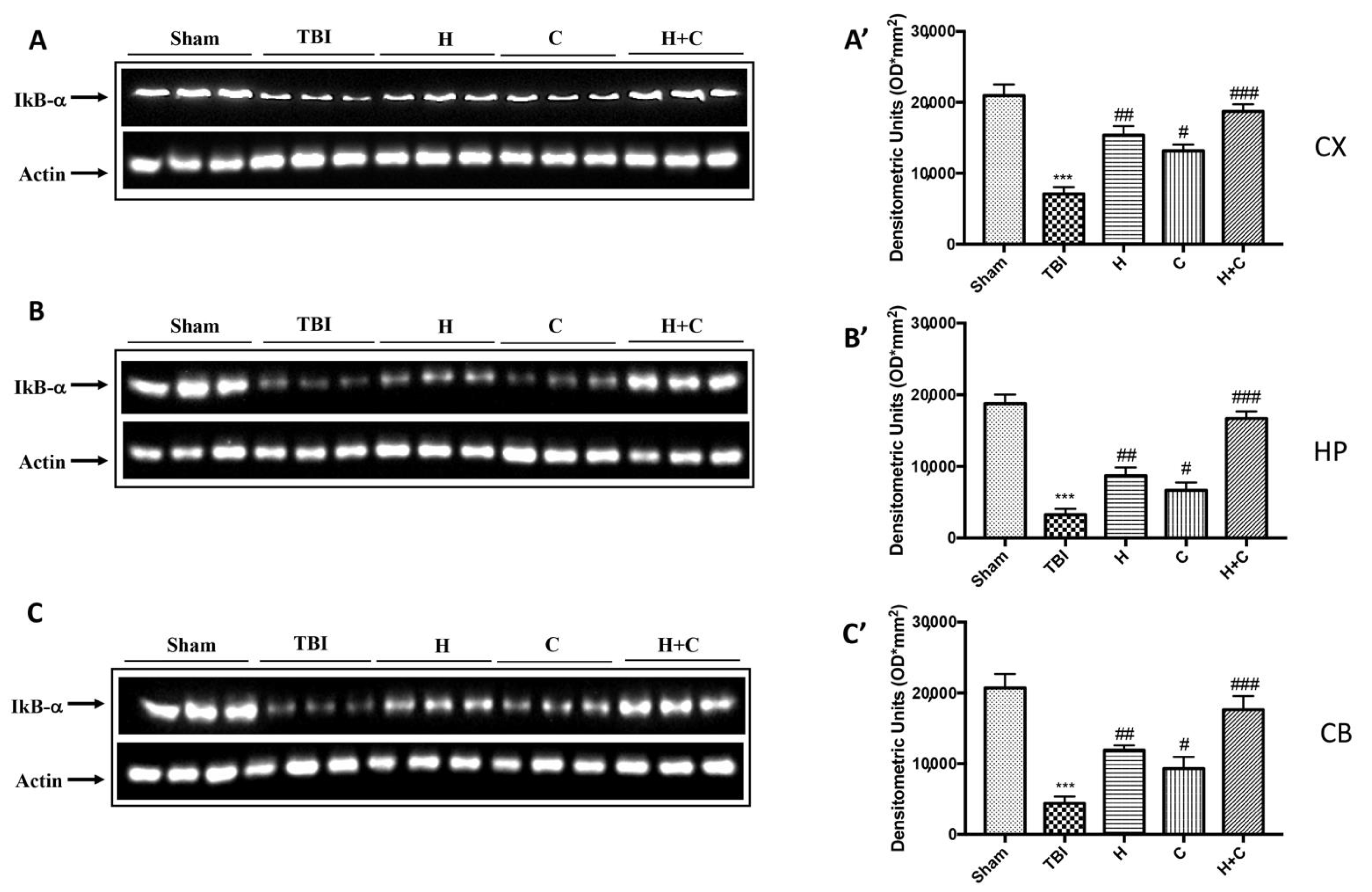
3.2. Effect of H. erinaceus or C. versicolor, or H. erinaceus Plus C. versicolor Treatment on Inflammatory Proteins Expression in the Chronic TBI
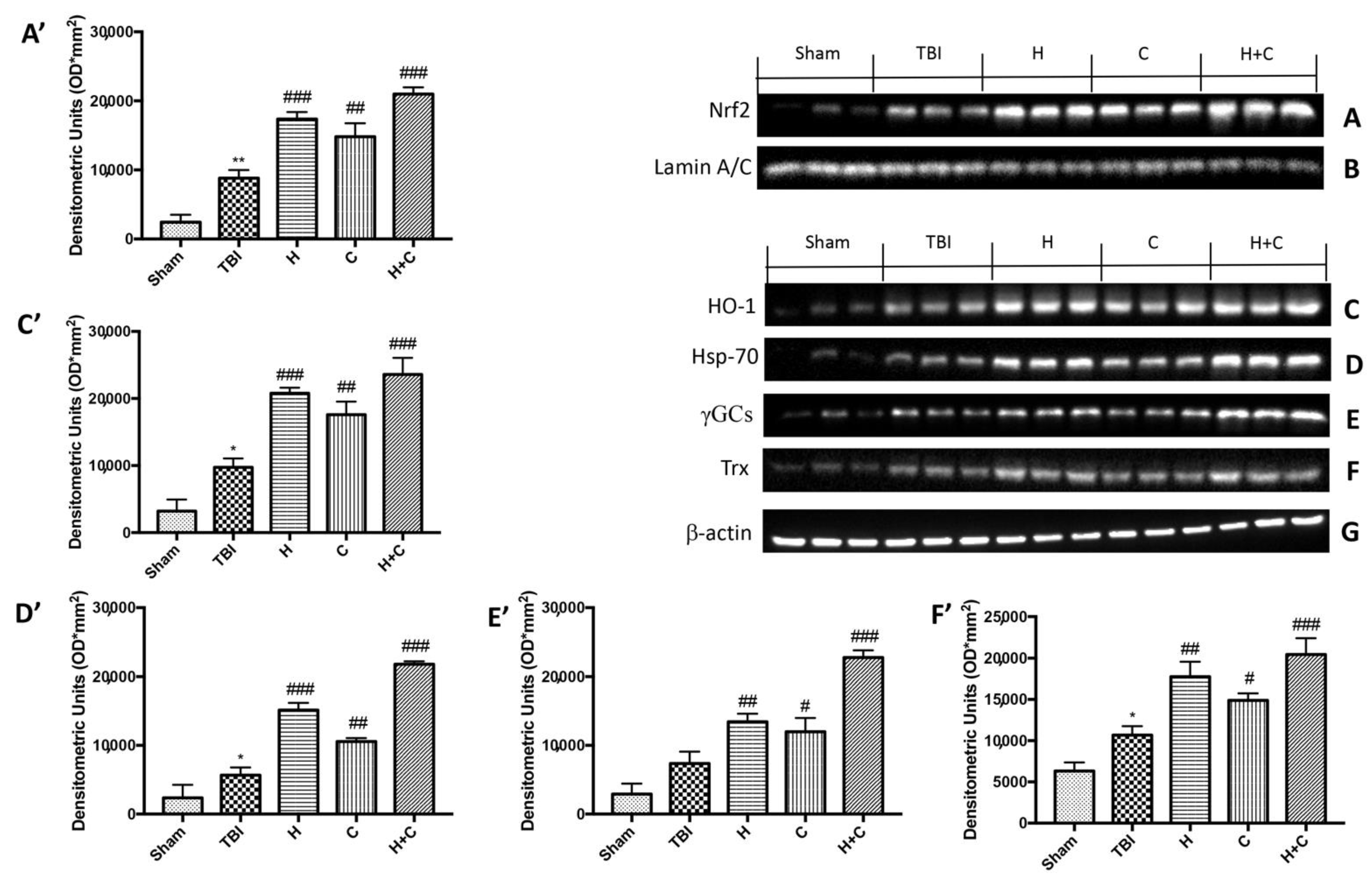
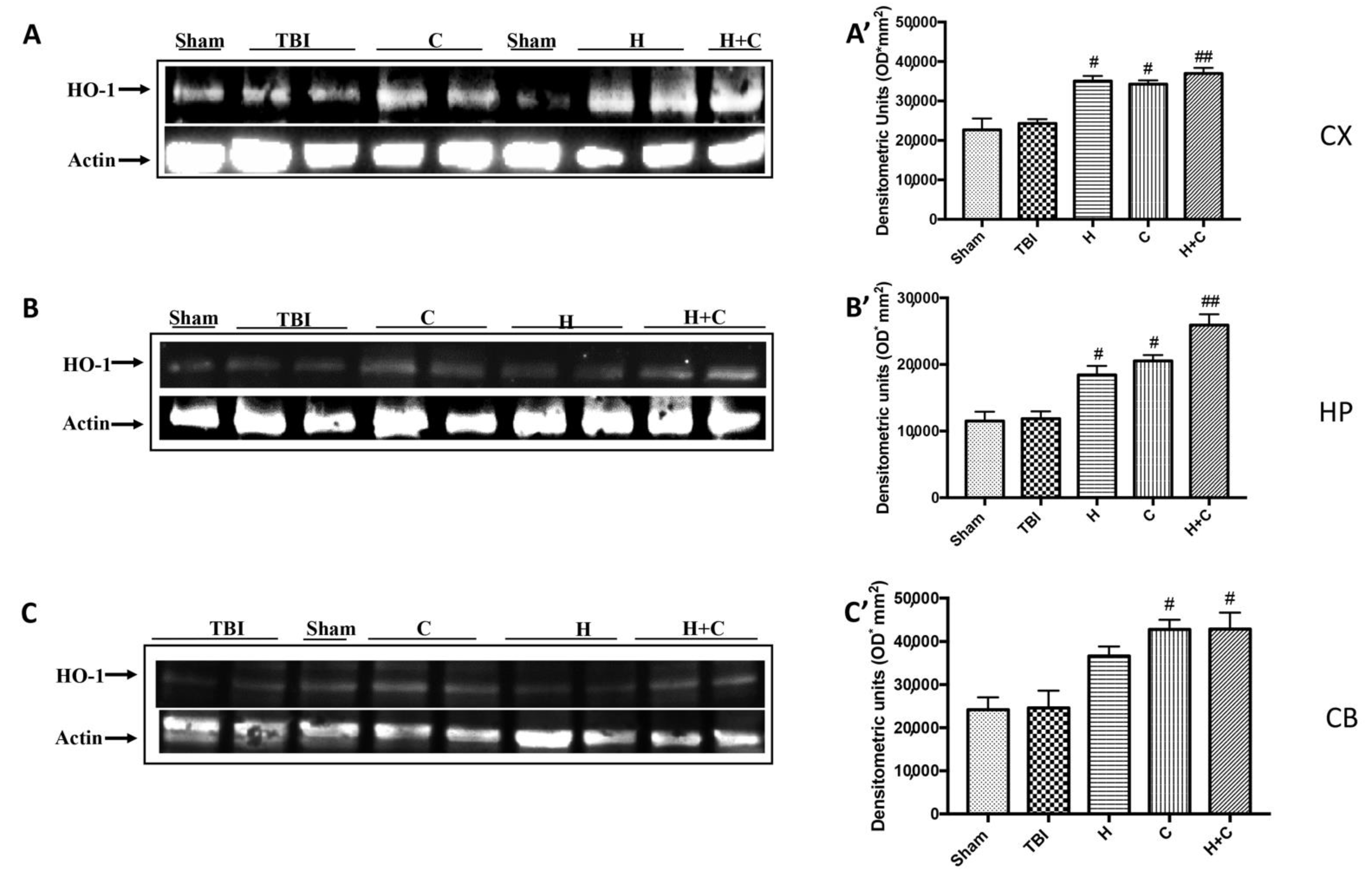
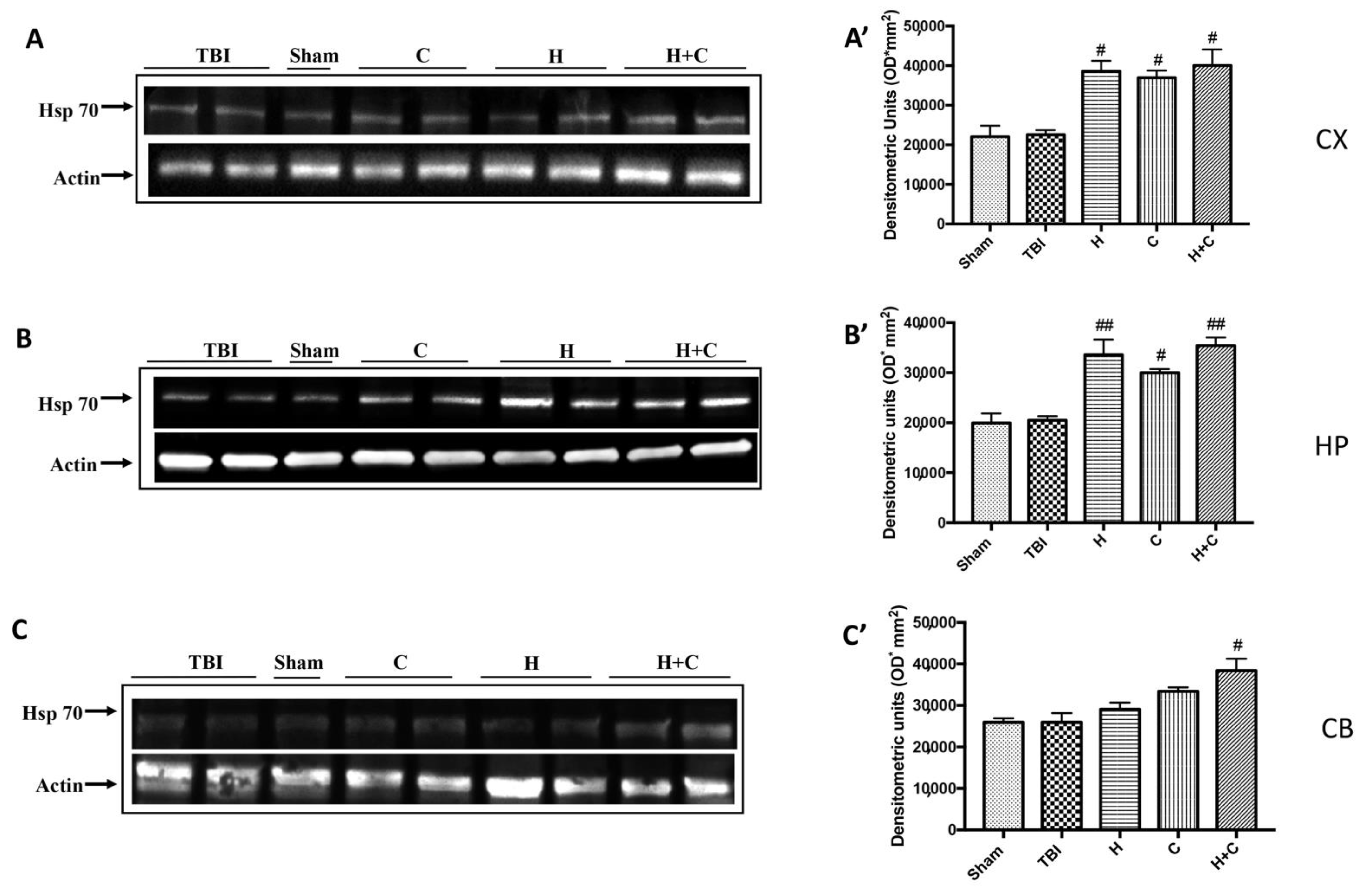
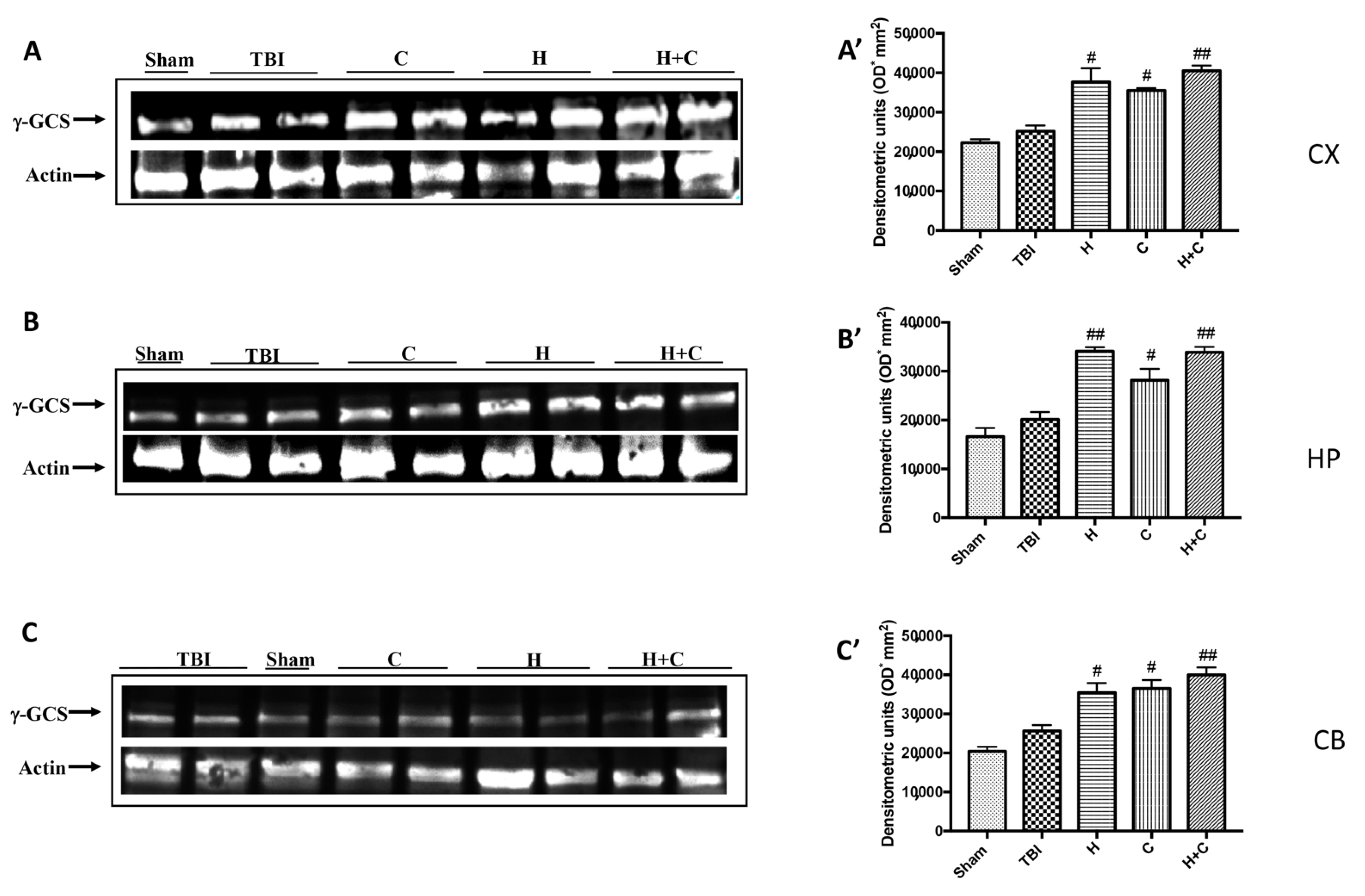
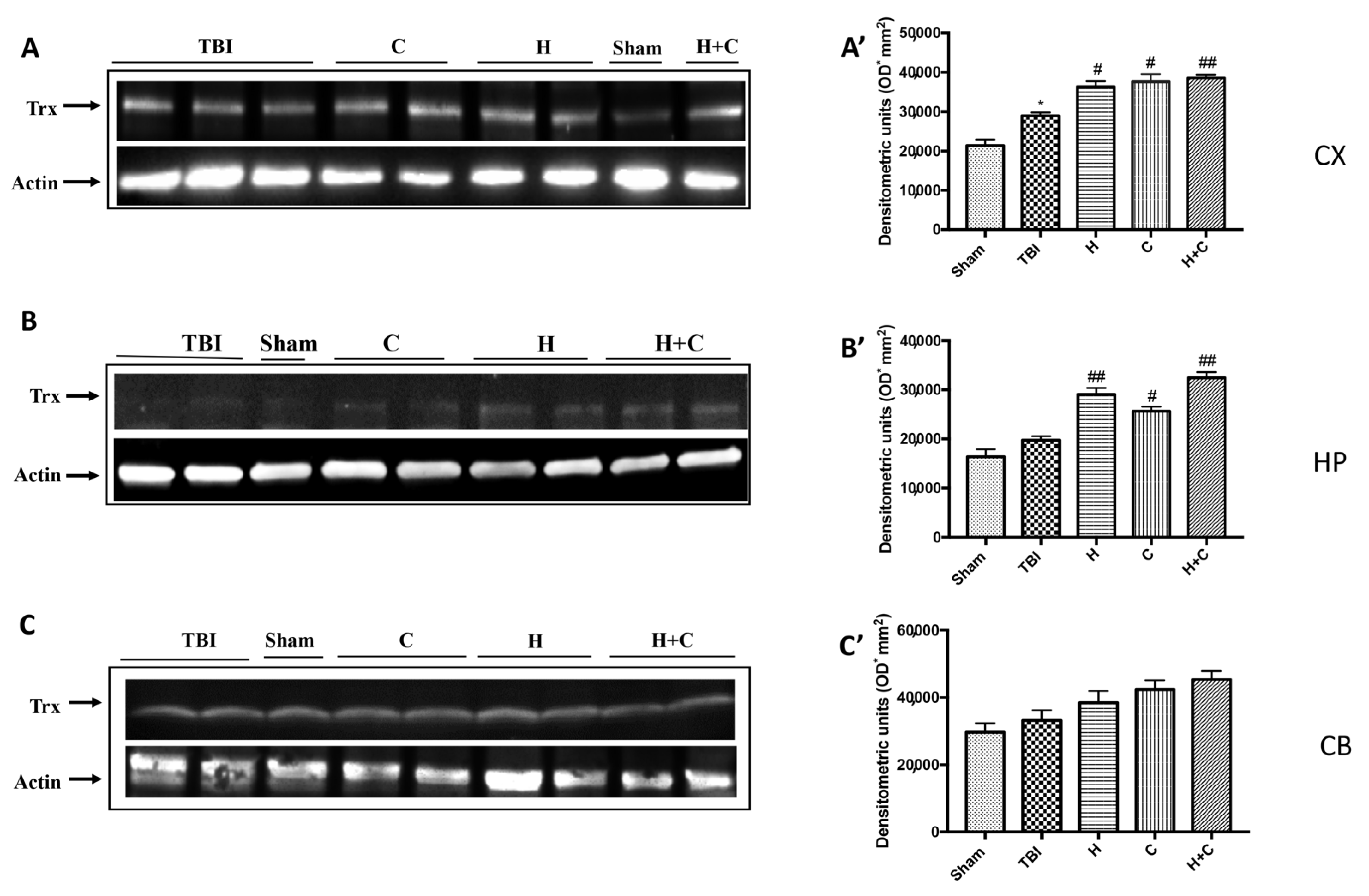
3.3. Effect of H. erinaceus or C. versicolor, or H. erinaceus Plus C. versicolor Treatment on Cellular Stress Response after Chronic TBI
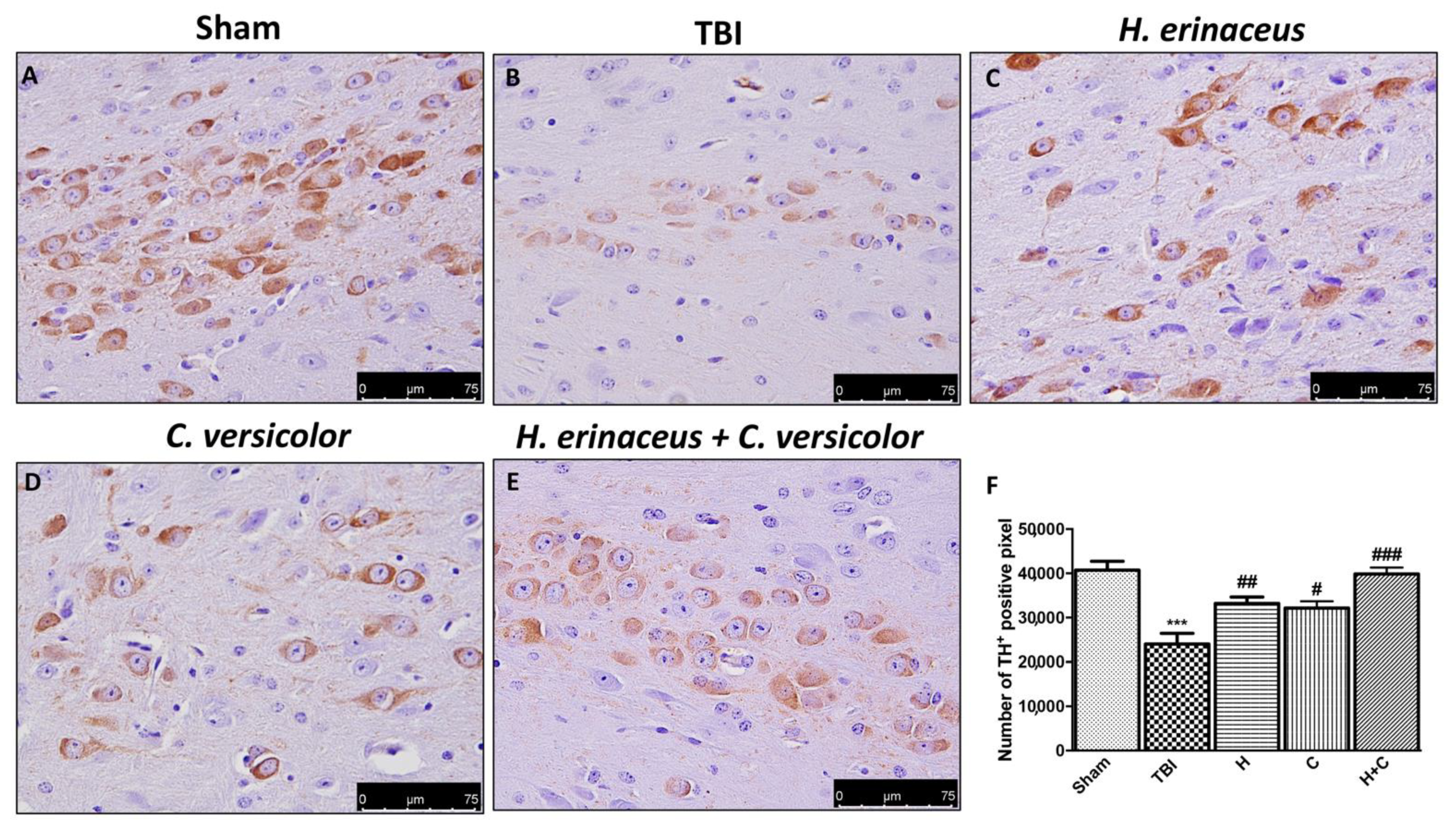
3.4. Effect of H. erinaceus or C. versicolor, or H. erinaceus Plus C. versicolor Treatment on Changes of PD Markers and on the Apoptotic Process after Chronic TBI
3.5. Effect of H. erinaceus or C. versicolor, or H. erinaceus Plus C. versicolor Treatment on Depression- and Anxiety-Like Behaviors in the Mouse after Chronic TBI
3.6. Characterization of H. erinaceus and C. versicolor
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lizhnyak, P.N.; Ottens, A.K. Proteomics: In pursuit of effective traumatic brain injury therapeutics. Expert Rev. Proteom. 2015, 12, 75–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sundman, M.H.; Hall, E.E.; Chen, N.K. Examining the relationship between head trauma and neurodegenerative disease: A review of epidemiology, pathology and neuroimaging techniques. J. Alzheimers Dis. Park. 2014, 4. [Google Scholar] [CrossRef] [Green Version]
- Jafari, S.; Etminan, M.; Aminzadeh, F.; Samii, A. Head injury and risk of Parkinson disease: A systematic review and meta-analysis. Mov. Disord 2013, 28, 1222–1229. [Google Scholar] [CrossRef] [PubMed]
- Crane, P.K.; Gibbons, L.E.; Dams-O’Connor, K.; Trittschuh, E.; Leverenz, J.B.; Keene, C.D.; Sonnen, J.; Montine, T.J.; Bennett, D.A.; Leurgans, S.; et al. Association of Traumatic Brain Injury With Late-Life Neurodegenerative Conditions and Neuropathologic Findings. JAMA Neurol. 2016, 73, 1062–1069. [Google Scholar] [CrossRef]
- Delic, V.; Beck, K.D.; Pang, K.C.H.; Citron, B.A. Biological links between traumatic brain injury and Parkinson’s disease. Acta Neuropathol. Commun. 2020, 8, 45. [Google Scholar] [CrossRef] [PubMed]
- Impellizzeri, D.; Campolo, M.; Bruschetta, G.; Crupi, R.; Cordaro, M.; Paterniti, I.; Cuzzocrea, S.; Esposito, E. Traumatic Brain Injury Leads to Development of Parkinson’s Disease Related Pathology in Mice. Front. Neurosci. 2016, 10, 458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardner, R.C.; Byers, A.L.; Barnes, D.E.; Li, Y.; Boscardin, J.; Yaffe, K. Mild TBI and risk of Parkinson disease: A Chronic Effects of Neurotrauma Consortium Study. Neurology 2018, 90, e1771–e1779. [Google Scholar] [CrossRef]
- Gaetz, M. The neurophysiology of brain injury. Clin. Neurophysiol. 2004, 115, 4–18. [Google Scholar] [CrossRef] [Green Version]
- Jullienne, A.; Badaut, J. Molecular contributions to neurovascular unit dysfunctions after brain injuries: Lessons for target-specific drug development. Future Neurol. 2013, 8, 677–689. [Google Scholar] [CrossRef] [Green Version]
- Impellizzeri, D.; Cordaro, M.; Bruschetta, G.; Siracusa, R.; Crupi, R.; Esposito, E.; Cuzzocrea, S. N-Palmitoylethanolamine-Oxazoline as a New Therapeutic Strategy to Control Neuroinflammation: Neuroprotective Effects in Experimental Models of Spinal Cord and Brain Injury. J. Neurotrauma 2017, 34, 2609–2623. [Google Scholar] [CrossRef]
- Fusco, R.; Gugliandolo, E.; Siracusa, R.; Scuto, M.; Cordaro, M.; D’Amico, R.; Evangelista, M.; Peli, A.; Peritore, A.F.; Impellizzeri, D.; et al. Formyl Peptide Receptor 1 Signaling in Acute Inflammation and Neural Differentiation Induced by Traumatic Brain Injury. Biology 2020, 9, 238. [Google Scholar] [CrossRef] [PubMed]
- Gentleman, S.M.; Leclercq, P.D.; Moyes, L.; Graham, D.I.; Smith, C.; Griffin, W.S.; Nicoll, J.A. Long-term intracerebral inflammatory response after traumatic brain injury. Forensic Sci. Int. 2004, 146, 97–104. [Google Scholar] [CrossRef]
- Lenzlinger, P.M.; Morganti-Kossmann, M.C.; Laurer, H.L.; McIntosh, T.K. The duality of the inflammatory response to traumatic brain injury. Mol. Neurobiol. 2001, 24, 169–181. [Google Scholar] [CrossRef]
- Schmidt, O.I.; Heyde, C.E.; Ertel, W.; Stahel, P.F. Closed head injury—An inflammatory disease? Brain Res. Brain Res. Rev. 2005, 48, 388–399. [Google Scholar] [CrossRef] [PubMed]
- Crupi, R.; Impellizzeri, D.; Cordaro, M.; Siracusa, R.; Casili, G.; Evangelista, M.; Cuzzocrea, S. N-palmitoylethanolamide Prevents Parkinsonian Phenotypes in Aged Mice. Mol. Neurobiol. 2018, 55, 8455–8472. [Google Scholar] [CrossRef] [PubMed]
- Friedman, M. Chemistry, Nutrition, and Health-Promoting Properties of Hericium erinaceus (Lion’s Mane) Mushroom Fruiting Bodies and Mycelia and Their Bioactive Compounds. J. Agric. Food Chem. 2015, 63, 7108–7123. [Google Scholar] [CrossRef] [PubMed]
- Tsai, Y.C.; Lin, Y.C.; Huang, C.C.; Villaflores, O.B.; Wu, T.Y.; Huang, S.M.; Chin, T.Y. Hericium erinaceus Mycelium and Its Isolated Compound, Erinacine A, Ameliorate High-Fat High-Sucrose Diet-Induced Metabolic Dysfunction and Spatial Learning Deficits in Aging Mice. J. Med. Food 2019, 22, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Tsai-Teng, T.; Chin-Chu, C.; Li-Ya, L.; Wan-Ping, C.; Chung-Kuang, L.; Chien-Chang, S.; Chi-Ying, H.F.; Chien-Chih, C.; Shiao, Y.J. Erinacine A-enriched Hericium erinaceus mycelium ameliorates Alzheimer’s disease-related pathologies in APPswe/PS1dE9 transgenic mice. J. Biomed. Sci. 2016, 23, 49. [Google Scholar] [CrossRef] [Green Version]
- Jang, H.J.; Kim, J.E.; Jeong, K.H.; Lim, S.C.; Kim, S.Y.; Cho, K.O. The Neuroprotective Effect of Hericium erinaceus Extracts in Mouse Hippocampus after Pilocarpine-Induced Status Epilepticus. Int. J. Mol. Sci. 2019, 20, 589. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.Y.; Huang, C.S.; Chen, Y.H.; Chen, C.C.; Chen, C.C.; Chuang, C.H. Anti-Inflammatory Effect of Erinacine C on NO Production Through Down-Regulation of NF-kappaB and Activation of Nrf2-Mediated HO-1 in BV2 Microglial Cells Treated with LPS. Molecules 2019, 24, 3317. [Google Scholar] [CrossRef] [Green Version]
- Trovato, A.; Siracusa, R.; Di Paola, R.; Scuto, M.; Ontario, M.L.; Bua, O.; Di Mauro, P.; Toscano, M.A.; Petralia, C.C.T.; Maiolino, L.; et al. Redox modulation of cellular stress response and lipoxin A4 expression by Hericium Erinaceus in rat brain: Relevance to Alzheimer’s disease pathogenesis. Immun. Ageing 2016, 13, 23. [Google Scholar] [CrossRef] [Green Version]
- Saleh, M.H.; Rashedi, I.; Keating, A. Immunomodulatory Properties of Coriolus versicolor: The Role of Polysaccharopeptide. Front. Immunol. 2017, 8, 1087. [Google Scholar] [CrossRef] [Green Version]
- Shi, S.H.; Yang, W.T.; Huang, K.Y.; Jiang, Y.L.; Yang, G.L.; Wang, C.F.; Li, Y. beta-glucans from Coriolus versicolor protect mice against S. typhimurium challenge by activation of macrophages. Int. J. Biol. Macromol. 2016, 86, 352–361. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Sun, Y.; Zhang, Q.; Zhang, H.; Yang, B.; Wang, Z.; Zhu, W.; Li, B.; Wang, Q.; Kuang, H. Screening and comparison of antioxidant activities of polysaccharides from Coriolus versicolor. Int. J. Biol. Macromol. 2014, 69, 12–19. [Google Scholar] [CrossRef]
- Fang, X.; Jiang, Y.; Ji, H.; Zhao, L.; Xiao, W.; Wang, Z.; Ding, G. The Synergistic Beneficial Effects of Ginkgo Flavonoid and Coriolus versicolor Polysaccharide for Memory Improvements in a Mouse Model of Dementia. Evid Based Complement. Altern. Med. 2015, 2015, 128394. [Google Scholar] [CrossRef] [PubMed]
- Trovato, A.; Siracusa, R.; Di Paola, R.; Scuto, M.; Fronte, V.; Koverech, G.; Luca, M.; Serra, A.; Toscano, M.A.; Petralia, A.; et al. Redox modulation of cellular stress response and lipoxin A4 expression by Coriolus versicolor in rat brain: Relevance to Alzheimer’s disease pathogenesis. Neurotoxicology 2016, 53, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Campolo, M.; Esposito, E.; Ahmad, A.; Di Paola, R.; Paterniti, I.; Cordaro, M.; Bruschetta, G.; Wallace, J.L.; Cuzzocrea, S. Hydrogen sulfide-releasing cyclooxygenase inhibitor ATB-346 enhances motor function and reduces cortical lesion volume following traumatic brain injury in mice. J. Neuroinflammation 2014, 11, 196. [Google Scholar] [CrossRef] [Green Version]
- Gugliandolo, E.; D’Amico, R.; Cordaro, M.; Fusco, R.; Siracusa, R.; Crupi, R.; Impellizzeri, D.; Cuzzocrea, S.; Di Paola, R. Neuroprotective Effect of Artesunate in Experimental Model of Traumatic Brain Injury. Front. Neurol. 2018, 9, 590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monro, J.A. Treatment of cancer with mushroom products. Arch. Environ. Health 2003, 58, 533–537. [Google Scholar] [CrossRef]
- Pellow, S.; Chopin, P.; File, S.E.; Briley, M. Validation of open:closed arm entries in an elevated plus-maze as a measure of anxiety in the rat. J. Neurosci. Methods 1985, 14, 149–167. [Google Scholar] [CrossRef]
- Prut, L.; Belzung, C. The open field as a paradigm to measure the effects of drugs on anxiety-like behaviors: A review. Eur. J. Pharmacol. 2003, 463, 3–33. [Google Scholar] [CrossRef]
- Barnes, C.A. Memory deficits associated with senescence: A neurophysiological and behavioral study in the rat. J. Comp. Physiol Psychol. 1979, 93, 74–104. [Google Scholar] [CrossRef] [PubMed]
- Siracusa, R.; Scuto, M.; Fusco, R.; Trovato, A.; Ontario, M.L.; Crea, R.; Di Paola, R.; Cuzzocrea, S.; Calabrese, V. Anti-inflammatory and Anti-oxidant Activity of Hidrox((R)) in Rotenone-Induced Parkinson’s Disease in Mice. Antioxidants 2020, 9, 824. [Google Scholar] [CrossRef] [PubMed]
- Campolo, M.; Paterniti, I.; Siracusa, R.; Filippone, A.; Esposito, E.; Cuzzocrea, S. TLR4 absence reduces neuroinflammation and inflammasome activation in Parkinson’s diseases in vivo model. Brain Behav. Immun. 2019, 76, 236–247. [Google Scholar] [CrossRef]
- Sawant, S.; Gokulan, R.; Dongre, H.; Vaidya, M.; Chaukar, D.; Prabhash, K.; Ingle, A.; Joshi, S.; Dange, P.; Joshi, S.; et al. Prognostic role of Oct4, CD44 and c-Myc in radio-chemo-resistant oral cancer patients and their tumourigenic potential in immunodeficient mice. Clin. Oral Investig. 2016, 20, 43–56. [Google Scholar] [CrossRef]
- Varghese, F.; Bukhari, A.B.; Malhotra, R.; De, A. IHC Profiler: An open source plugin for the quantitative evaluation and automated scoring of immunohistochemistry images of human tissue samples. PLoS ONE 2014, 9, e96801. [Google Scholar] [CrossRef] [Green Version]
- Siracusa, R.; Paterniti, I.; Bruschetta, G.; Cordaro, M.; Impellizzeri, D.; Crupi, R.; Cuzzocrea, S.; Esposito, E. The Association of Palmitoylethanolamide with Luteolin Decreases Autophagy in Spinal Cord Injury. Mol. Neurobiol. 2016, 53, 3783–3792. [Google Scholar] [CrossRef] [Green Version]
- Nonaka, M.; Chen, X.H.; Pierce, J.E.; Leoni, M.J.; McIntosh, T.K.; Wolf, J.A.; Smith, D.H. Prolonged activation of NF-kappaB following traumatic brain injury in rats. J. Neurotrauma 1999, 16, 1023–1034. [Google Scholar] [CrossRef]
- Cornelius, C.; Crupi, R.; Calabrese, V.; Graziano, A.; Milone, P.; Pennisi, G.; Radak, Z.; Calabrese, E.J.; Cuzzocrea, S. Traumatic brain injury: Oxidative stress and neuroprotection. Antioxid Redox Signal. 2013, 19, 836–853. [Google Scholar] [CrossRef]
- Trovato Salinaro, A.; Pennisi, M.; Di Paola, R.; Scuto, M.; Crupi, R.; Cambria, M.T.; Ontario, M.L.; Tomasello, M.; Uva, M.; Maiolino, L.; et al. Neuroinflammation and neurohormesis in the pathogenesis of Alzheimer’s disease and Alzheimer-linked pathologies: Modulation by nutritional mushrooms. Immun. Ageing 2018, 15, 8. [Google Scholar] [CrossRef] [Green Version]
- Uryu, K.; Giasson, B.I.; Longhi, L.; Martinez, D.; Murray, I.; Conte, V.; Nakamura, M.; Saatman, K.; Talbot, K.; Horiguchi, T.; et al. Age-dependent synuclein pathology following traumatic brain injury in mice. Exp. Neurol. 2003, 184, 214–224. [Google Scholar] [CrossRef]
- Xiong, Y.; Mahmood, A.; Chopp, M. Animal models of traumatic brain injury. Nat. Rev. Neurosci. 2013, 14, 128–142. [Google Scholar] [CrossRef] [Green Version]
- Rappold, P.M.; Tieu, K. Astrocytes and therapeutics for Parkinson’s disease. Neurotherapeutics 2010, 7, 413–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brough, D.; Tyrrell, P.J.; Allan, S.M. Regulation of interleukin-1 in acute brain injury. Trends Pharm. Sci. 2011, 32, 617–622. [Google Scholar] [CrossRef]
- Woodcock, T.; Morganti-Kossmann, M.C. The role of markers of inflammation in traumatic brain injury. Front. Neurol. 2013, 4, 18. [Google Scholar] [CrossRef] [Green Version]
- Acosta, S.A.; Tajiri, N.; de la Pena, I.; Bastawrous, M.; Sanberg, P.R.; Kaneko, Y.; Borlongan, C.V. Alpha-synuclein as a pathological link between chronic traumatic brain injury and Parkinson’s disease. J. Cell Physiol. 2015, 230, 1024–1032. [Google Scholar] [CrossRef] [PubMed]
- Phani, S.; Loike, J.D.; Przedborski, S. Neurodegeneration and inflammation in Parkinson’s disease. Park. Relat. Disord. 2012, 18 (Suppl. S1), S207–S209. [Google Scholar] [CrossRef]
- Bonito-Oliva, A.; Masini, D.; Fisone, G. A mouse model of non-motor symptoms in Parkinson’s disease: Focus on pharmacological interventions targeting affective dysfunctions. Front. Behav. Neurosci. 2014, 8, 290. [Google Scholar] [CrossRef] [Green Version]




















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Retention Time (min) | Area (%) |
|---|---|---|
| Hexadecanoic acid | 0.64 | 15.14 |
| Glycerol | 2.78 | 0.76 |
| Myristic acid | 3.23 | 2.79 |
| Palmitic acid | 3.43 | 1.01 |
| Stearic acid | 4.14 | 2.84 |
| Tetradecane | 4.22 | 0.97 |
| 3-Methoxy-4-benzaldehyde | 4.52 | 0.4 |
| Sphinganine | 5.06 | 1.02 |
| 8-Sphingenine | 5.14 | 2.89 |
| 4-Hydroxysphinganine | 5.21 | 2.76 |
| α-Cadinene | 5.87 | 5.05 |
| Hexadecane | 5.95 | 13.59 |
| Oxybenzaldehyde | 6.76 | 0.97 |
| Homovanillyl alcohol | 7.37 | 0.61 |
| Vanillic acid | 7.75 | 0.42 |
| Octadecane | 9.17 | 3.38 |
| Oleic Acid | 9.50 | 1.84 |
| Linoleic acid | 9.78 | 2.51 |
| 3,5-Dimethoxy-4-benzoate | 9.95 | 3.61 |
| (1-Methyldodecyl)-benzene | 10.51 | 2.91 |
| 9-Dodecenoic methyl ester | 11.03 | 4.78 |
| Kaempferol | 11.54 | 5.32 |
| Coumaric acid | 11.57 | 3.46 |
| Eicosane | 11.65 | 6.93 |
| Glucose | 11.85 | 14.04 |
| Compound | Retention Time (min) | Area (%) | Exact Mass |
|---|---|---|---|
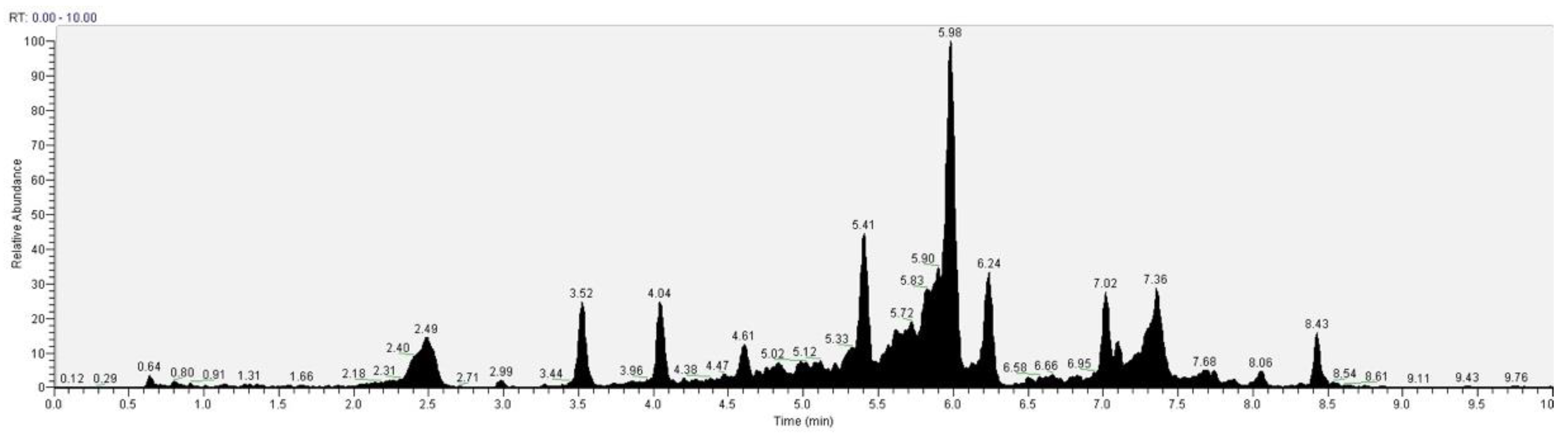
| Gallic acid | 2.49 | 3.61 | 169.014 |
| Caffeic acid | 2.99 | 0.79 | 179.034 |
| Catechin | 3.52 | 7.73 | 289.071 |
| Epicatechin | 4.04 | 7.56 | 289.071 |
| Vanillic acid | 5.41 | 11.93 | 167.041 |
| Syringic acid | 5.72 | 3.75 | 197.045 |
| Hydroxybenzoic acid | 5.98 | 29.84 | 138.03 |
| Ferulic acid | 6.24 | 10.03 | 193.057 |
| Naringenin | 6.5 | 0.88 | 272.067 |
| Rutin | 6.66 | 1.35 | 610.012 |
| Quercetin | 7.02 | 6.95 | 447.093 |
| Apigenin | 7.36 | 9.34 | 271.060 |
| Luteolin | 8.06 | 2.08 | 286.04 |
| Kaempferol | 8.43 | 6.24 | 285.040 |
| Compound | Retention Time (min) | Area (%) |
|---|---|---|
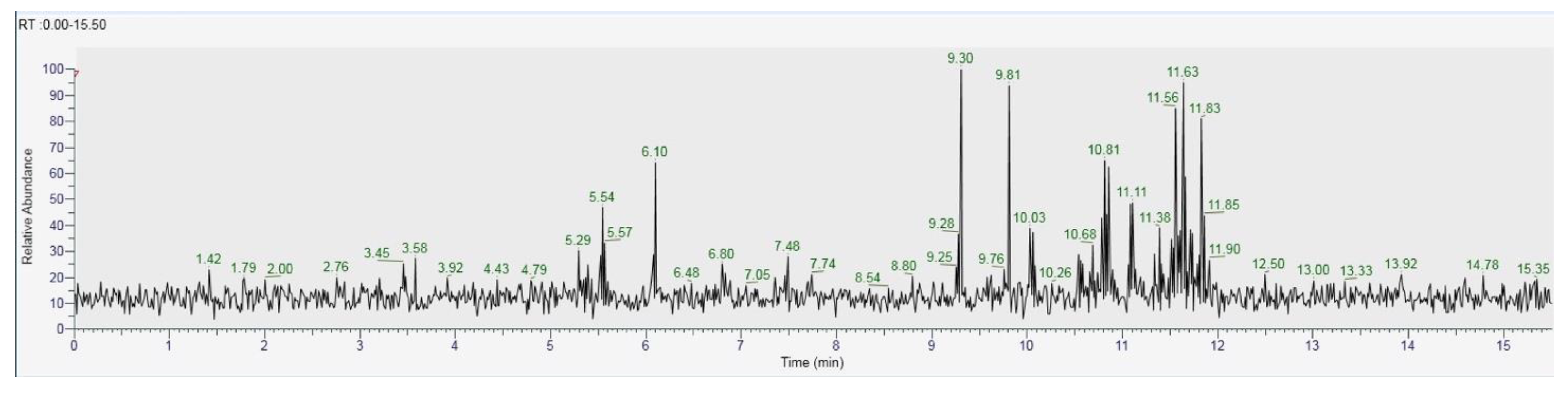
| Hexadecenoic acid | 3.45 | 0.74 |
| Palmitic acid | 5.29 | 2.05 |
| Stearic acid | 5.54 | 5.97 |
| Octadecane | 6.10 | 7.83 |
| Ergosterol | 9.30 | 15.34 |
| Ergothioneine | 9.81 | 12.72 |
| Vanillic acid | 10.03 | 4.22 |
| Oleic Acid | 10.81 | 8.73 |
| Linoleic acid | 11.11 | 5.01 |
| Kaempferol | 11.38 | 1.86 |
| Coumaric acid | 11.56 | 11.03 |
| Glucose | 11.63 | 14.05 |
| Tartaric acid | 11.83 | 10.45 |
| Compound | Retention Time (min) | Area (%) |
|---|---|---|
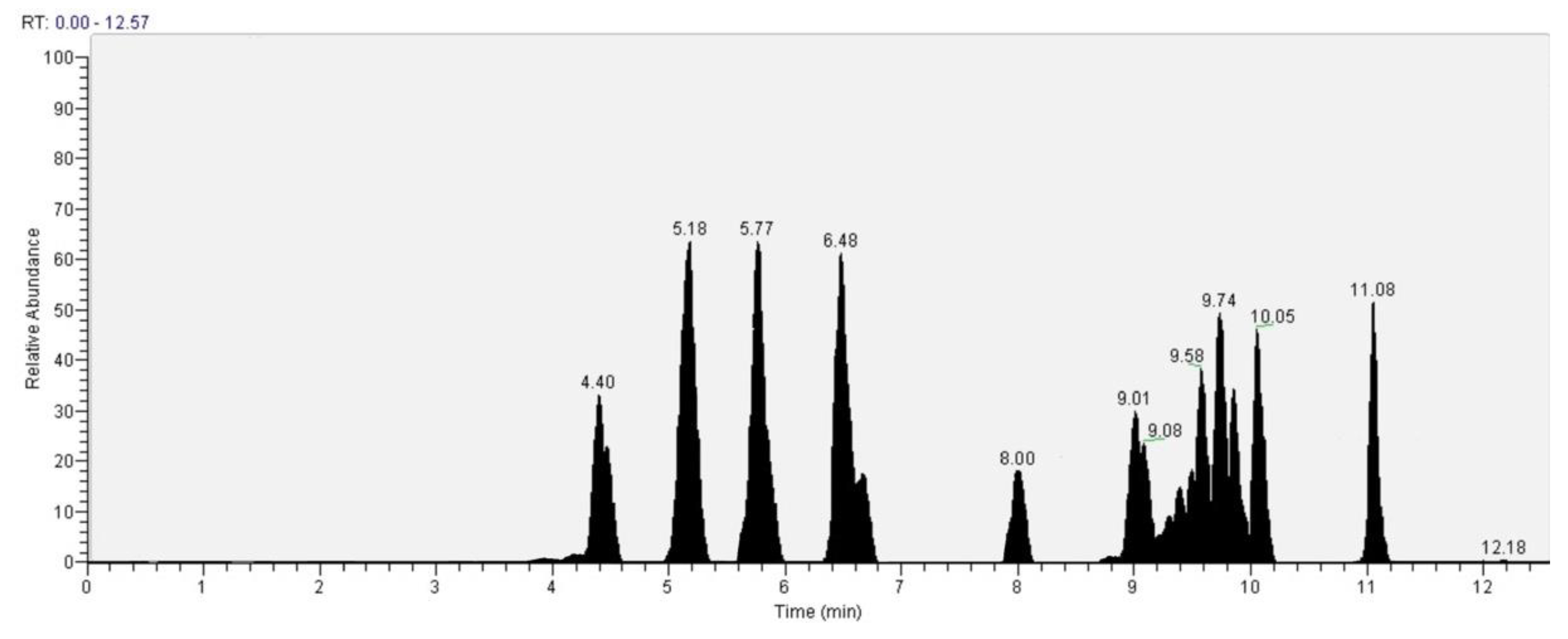
| Ascorbic acid | 4.4 | 4.75 |
| Caffeic acid | 4.47 | 3.22 |
| Catechin | 5.18 | 14.01 |
| Epicatechin | 5.77 | 13.46 |
| Vanillic acid | 6.48 | 11.07 |
| Chlorogenic acid | 6.53 | 2.98 |
| Hydroxybenzoic acid | 8.00 | 3.76 |
| Ferulic acid | 9.01 | 5.03 |
| p-Coumaric acid | 9.08 | 3.38 |
| Sinaptic acid | 9.58 | 6.28 |
| Cinnamic acid | 9.74 | 8.05 |
| Rutin | 9.83 | 5.97 |
| Quercetin | 10.05 | 8.31 |
| Kaempferol | 11.08 | 9.73 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Amico, R.; Trovato Salinaro, A.; Fusco, R.; Cordaro, M.; Impellizzeri, D.; Scuto, M.; Ontario, M.L.; Lo Dico, G.; Cuzzocrea, S.; Di Paola, R.; et al. Hericium erinaceus and Coriolus versicolor Modulate Molecular and Biochemical Changes after Traumatic Brain Injury. Antioxidants 2021, 10, 898. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10060898
D’Amico R, Trovato Salinaro A, Fusco R, Cordaro M, Impellizzeri D, Scuto M, Ontario ML, Lo Dico G, Cuzzocrea S, Di Paola R, et al. Hericium erinaceus and Coriolus versicolor Modulate Molecular and Biochemical Changes after Traumatic Brain Injury. Antioxidants. 2021; 10(6):898. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10060898
Chicago/Turabian StyleD’Amico, Ramona, Angela Trovato Salinaro, Roberta Fusco, Marika Cordaro, Daniela Impellizzeri, Maria Scuto, Maria Laura Ontario, Gianluigi Lo Dico, Salvatore Cuzzocrea, Rosanna Di Paola, and et al. 2021. "Hericium erinaceus and Coriolus versicolor Modulate Molecular and Biochemical Changes after Traumatic Brain Injury" Antioxidants 10, no. 6: 898. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10060898