
Hydroxy-Selenomethionine Improves the Selenium Status and Helps to Maintain Broiler Performances under a High Stocking Density and Heat Stress Conditions through a Better Redox and Immune Response

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chickens, Treaztments, and Sample Collection
2.2. Selenium Concentration and Antioxidant Parameter Analysis
2.3. Serum Biochemical, Immunoglobulin and Histological Analyses
2.4. Western-Blot Analyses
2.5. Statistical Analysis
3. Results
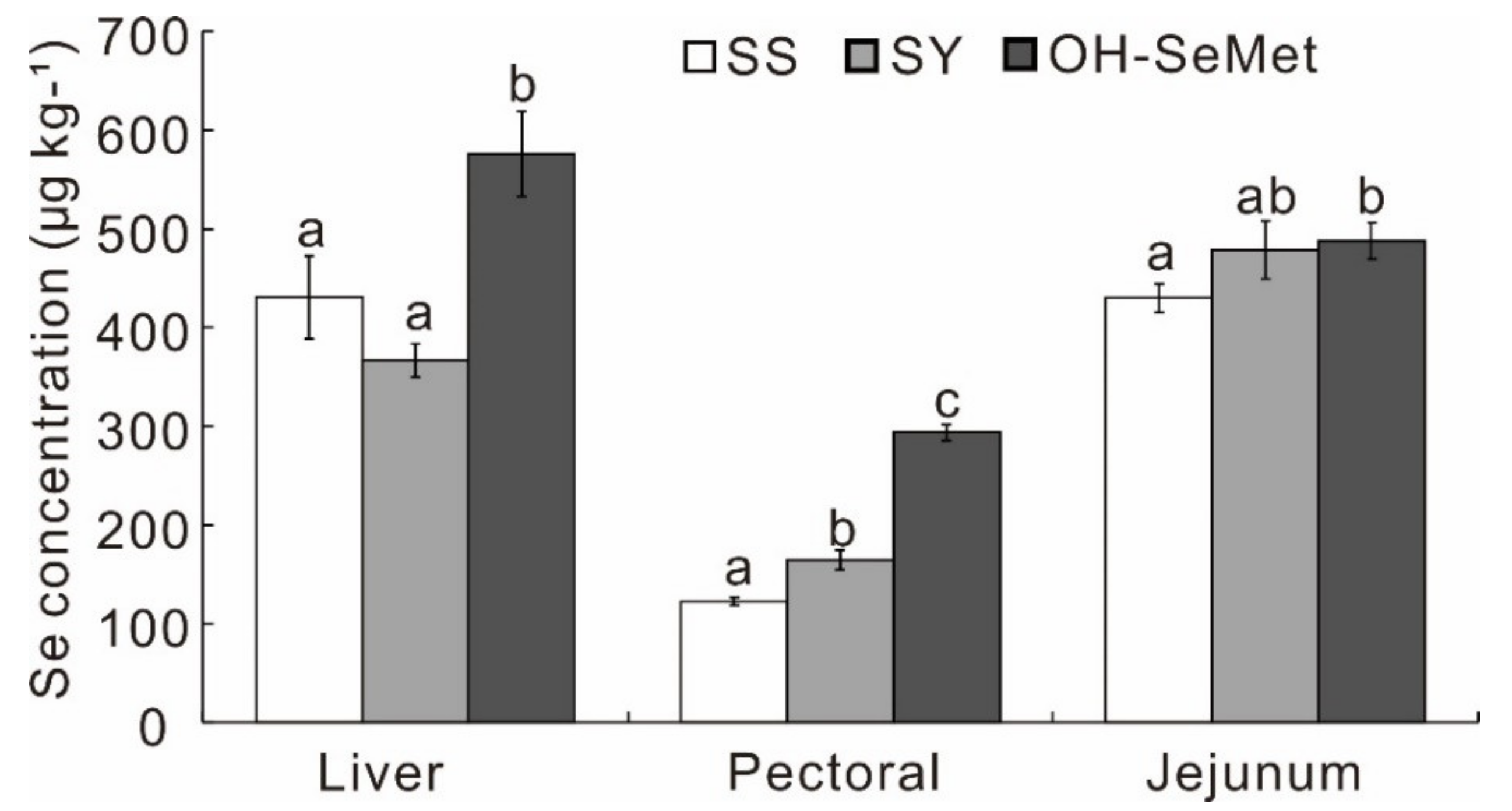
3.1. Growth Performance and Deposition of Selenium
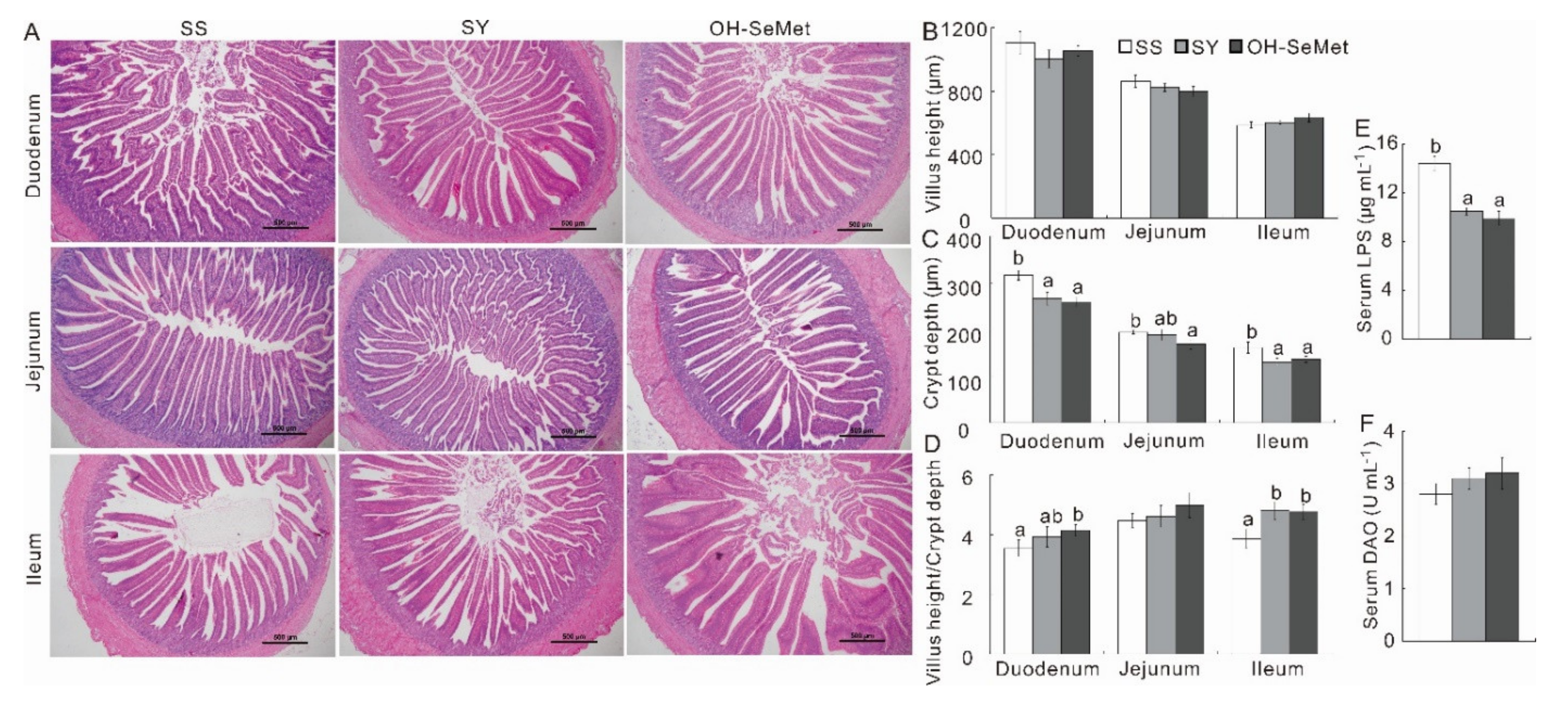
3.2. Serum Biochemistry and Small Intestinal Histology
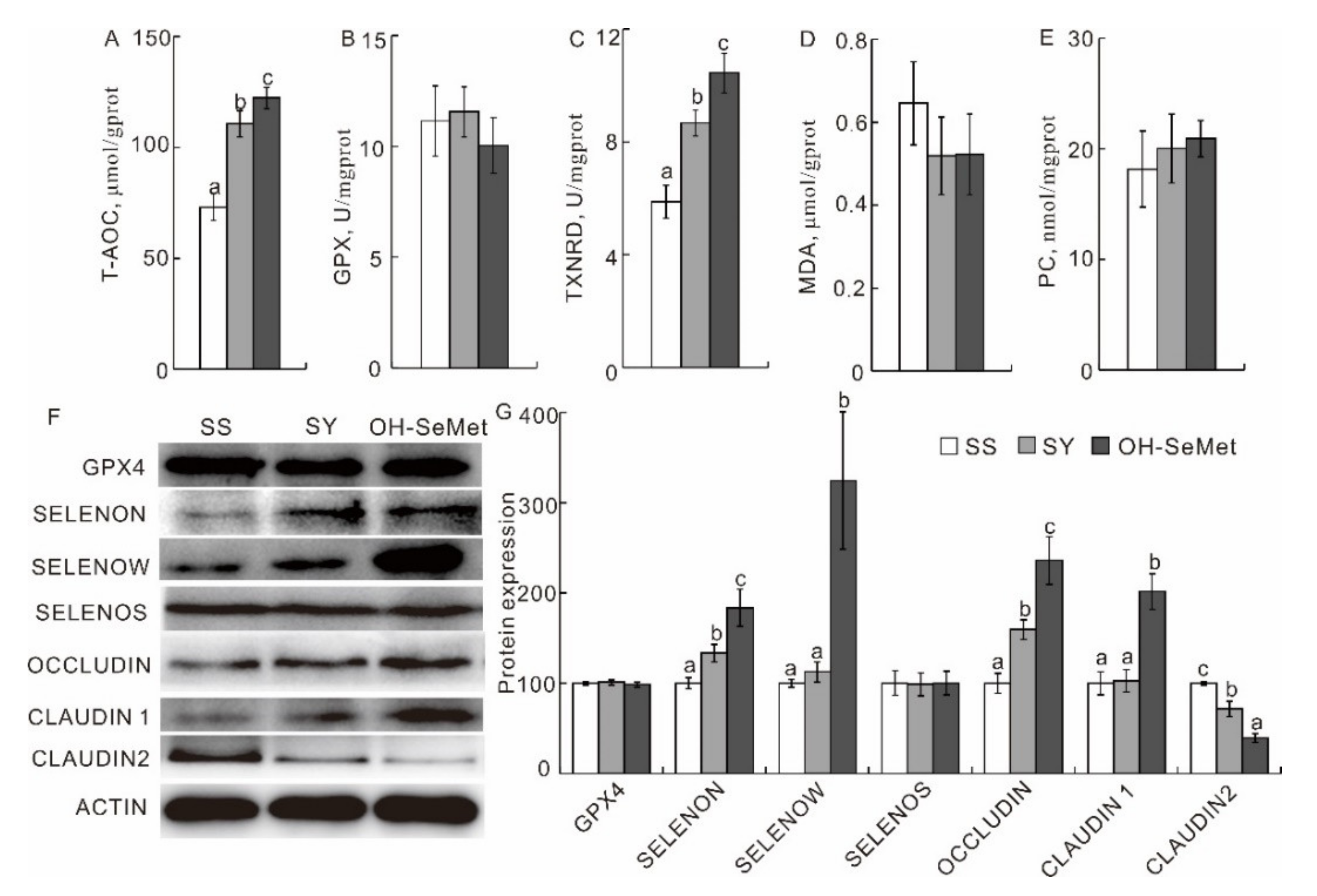
3.3. Redox Status and Production of Selenoprotein and Tight Junction-Related Proteins in the Jejunum
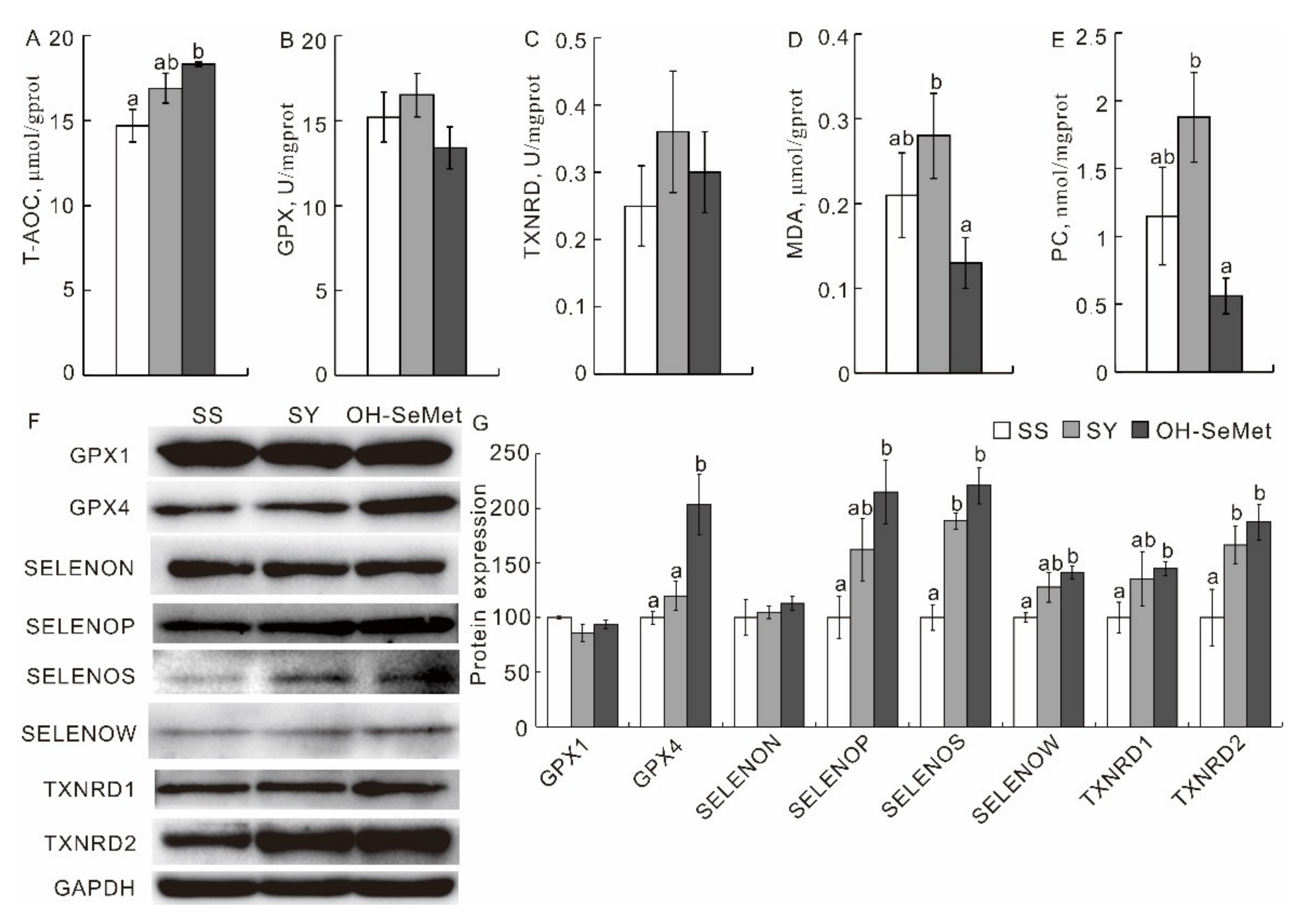
3.4. Redox Status and Production of Selenoproteins in the Pectoral Muscle
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, H.Y.; Piao, X.S.; Zhang, Q.; Li, P.; Yi, J.Q.; Liu, J.D.; Li, Q.Y.; Wang, G.Q. The effects of Forsythia suspensa extract and berberine on growth performance, immunity, antioxidant activities, and intestinal microbiota in broilers under high stocking density. Poult. Sci. 2013, 92, 1981–1988. [Google Scholar] [CrossRef] [PubMed]
- Goo, D.; Kim, J.H.; Park, G.H.; Delos-Reyes, J.B.; Kil, D.Y. Effect of Heat Stress and Stocking Density on Growth Performance, Breast Meat Quality and Intestinal Barrier Function in Broiler Chickens. Animals 2019, 9, 107. [Google Scholar] [CrossRef] [Green Version]
- Lara, L.J.; Rostagno, M.H. Impact of Heat Stress on Poultry Production. Animals 2013, 3, 356–369. [Google Scholar] [CrossRef]
- Padgett, D.A.; Glaser, R. How stress influences the immune response. Trends Immunol. 2003, 24, 444–448. [Google Scholar] [CrossRef]
- Niu, Z.Y.; Liu, F.Z.; Yan, Q.L.; Li, W.C. Effects of different levels of vitamin E on growth performance and immune responses of broilers under heat stress. Poult. Sci. 2009, 88, 2101–2107. [Google Scholar] [CrossRef] [PubMed]
- Rayman, M.P.; Stranges, S. Epidemiology of selenium and type 2 diabetes: Can we make sense of it? Free Radic. Biol. Med. 2013, 65, 1557–1564. [Google Scholar] [CrossRef]
- Labunskyy, V.M.; Hatfield, D.L.; Gladyshev, V.N. Selenoproteins: Molecular pathways and physiological roles. Physiol Rev. 2014, 94, 739–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, X.D.; Lu, L.; Li, S.F. Effects of selenium source and level on growth performance, tissue selenium concentrations, antioxidation, and immune functions of heat-stressed broilers. Biol. Trace Elem. Res. 2012, 150, 158–165. [Google Scholar] [CrossRef]
- Habibian, M.; Sadeghi, G.; Ghazi, S.; Moeini, M.M. Selenium as a feed supplement for heat-stressed poultry: A review. Biol. Trace Elem. Res. 2015, 165, 183–193. [Google Scholar] [CrossRef]
- Shakeri, M.; Oskoueian, E.; Le, H.H.; Shakeri, M. Strategies to Combat Heat Stress in Broiler Chickens: Unveiling the Roles of Selenium, Vitamin E and Vitamin C. Vet. Sci. 2020, 7, 71. [Google Scholar] [CrossRef]
- Safdari-Rostamabad, M.; Hosseini-Vashan, S.J.; Perai, A.H.; Sarir, H. Nanoselenium Supplementation of Heat-Stressed Broilers: Effects on Performance, Carcass Characteristics, Blood Metabolites, Immune Response, Antioxidant Status, and Jejunal Morphology. Biol. Trace Elem. Res. 2017, 178, 105–116. [Google Scholar] [CrossRef]
- Surai, P.F.; Kochish, I.I.; Fisinin, V.I.; Velichko, O.A. Selenium in Poultry Nutrition: From Sodium Selenite to Organic Selenium Sources. J. Poult. Sci. 2018, 55, 79–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EFSA. Scientific Opinion of the Panel on Additives and Products or Substances used in Animal Feed (FEEDAP) on a request from the European Commission on the safety and efficacy of SELSAF (Selenium enriched yeast from Saccharomyces cerevisiae CNCM I-3399) as feed additive for all species. EFSA J. 2009, 9, 2279. [Google Scholar]
- EFSA. Scientific opinion on safety and efficacy of hydroxyanalogue of selenomethionine as feed additive for all species. EFSA J. 2013, 11, 3046. [Google Scholar] [CrossRef] [Green Version]
- EFSA. Assessment of the application for renewal of authorisation of selenomethionine produced by Saccharomyces cerevisiae NCYC R397 for all animal species. EFSA J. 2019, 17, e05539. [Google Scholar]
- EFSA. Assessment of the application for renewal of authorisation of selenomethionine produced by Saccharomyces cerevisiae CNCM I-3060 (selenised yeast inactivated) for all animal species. EFSA J. 2018, 16, e05386. [Google Scholar]
- Surai, P.F.; Kochish, I.I. Nutritional modulation of the antioxidant capacities in poultry: The case of selenium. Poult. Sci. 2019, 98, 4231–4239. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Wang, J.; Liu, W.; Bu, D.P.; Liu, S.J.; Zhang, K.Z. Hydroxy-selenomethionine: A novel organic selenium source that improves antioxidant status and selenium concentrations in milk and plasma of mid-lactation dairy cows. J. Dairy Sci. 2017, 100, 9602–9610. [Google Scholar] [CrossRef] [Green Version]
- Juniper, D.T.; Rymer, C.; Briens, M. Bioefficacy of hydroxy-selenomethionine as a selenium supplement in pregnant dairy heifers and on the selenium status of their calves. J. Dairy Sci. 2019, 102, 7000–7010. [Google Scholar] [CrossRef] [PubMed]
- Jlali, M.; Briens, M.; Rouffineau, F.; Geraert, P.A.; Mercier, Y. Evaluation of the efficacy of 2-hydroxy-4-methylselenobutanoic acid on growth performance and tissue selenium retention in growing pigs. J. Anim. Sci. 2014, 92, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Li, N.Y.; Sun, Z.J.; Ansari, A.R.; Cui, L.; Hu, Y.F.; Li, Z.W.; Briens, M.; Kai, L.; Sun, L.H.; Karrow, N.A.; et al. Impact of Maternal Selenium Supplementation from Late Gestation and Lactation on Piglet Immune Function. Biol. Trace Elem. Res. 2020, 194, 159–167. [Google Scholar] [CrossRef]
- Jlali, M.; Briens, M.; Rouffineau, F.; Mercerand, F.; Geraert, P.A.; Mercier, Y. Effect of 2-hydroxy-4-methylselenobutanoic acid as a dietary selenium supplement to improve the selenium concentration of table eggs. J. Anim. Sci. 2013, 91, 1745–1752. [Google Scholar] [CrossRef] [PubMed]
- Tufarelli, V.; Ceci, E.; Laudadio, V. 2-Hydroxy-4-Methylselenobutanoic Acid as New Organic Selenium Dietary Supplement to Produce Selenium-Enriched Eggs. Biol. Trace Elem. Res. 2016, 171, 453–458. [Google Scholar] [CrossRef]
- Briens, M.; Mercier, Y.; Rouffineau, F.; Mercerand, F.; Geraert, P.A. 2-Hydroxy-4-methylselenobutanoic acid induces additional tissue selenium enrichment in broiler chickens compared with other selenium sources. Poult. Sci. 2014, 93, 85–93. [Google Scholar] [CrossRef] [PubMed]
- De Marco, M.; Conjat, A.S.; Briens, M.; Hachemi, M.A.; Geraert, P.A. Bio-efficacy of organic selenium compounds in broiler chickens. Ital. J. Anim. Sci. 2021, 20, 514–525. [Google Scholar] [CrossRef]
- Zhao, L.; Sun, L.H.; Huang, J.Q.; Briens, M.; Qi, D.S.; Xu, S.W.; Lei, X.G. A Novel Organic Selenium Compound Exerts Unique Regulation of Selenium Speciation, Selenogenome, and Selenoproteins in Broiler Chicks. J. Nutr. 2017, 147, 789–797. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.H.; Huang, J.Q.; Deng, J.; Lei, X.G. Avian selenogenome: Response to dietary Se and vitamin E deficiency and supplementation. Poult. Sci. 2019, 98, 4247–4254. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.H.; Zhang, N.Y.; Zhu, M.K.; Zhao, L.; Zhou, J.C.; Qi, D.S. Prevention of Aflatoxin B1 Hepatoxicity by Dietary Selenium Is Associated with Inhibition of Cytochrome P450 Isozymes and Up-Regulation of 6 Selenoprotein Genes in Chick Liver. J. Nutr. 2015, 146, 655–661. [Google Scholar] [CrossRef]
- McClung, J.P.; Roneker, C.A.; Mu, W.; Lisk, D.J.; Langlais, P.; Liu, F.; Lei, X.G. Development of insulin resistance and obesity in mice overexpressing cellular glutathione peroxidase. Proc. Natl. Acad. Sci. USA 2004, 101, 8852–8857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Rivas, P.A.; Chauhan, S.S.; Ha, M.; Fegan, N.; Dunshea, F.R.; Warner, R.D. Effects of heat stress on animal physiology, metabolism, and meat quality: A review. Meat Sci. 2020, 162, 108025. [Google Scholar] [CrossRef]
- Melaku, W.; Mengistu, U. Performance of Cobb 500 and Hubbard Broilers on Different Treatment; LAP LAMBERT Academic Publishing: Saarbrücken, Germany, 2015. [Google Scholar]
- Angeles-Hernandez, J.C. Effect of selenium supplementation on productive performance and antioxidant status of broilers under heat stress: A meta-analysis and a meta-regression. Anim. Prod. Sci. 2021, 61, 208–214. [Google Scholar]
- Michiels, J.; Majdeddin, M.; Pincemail, J.; De-Marco, M.; Liu, Y.G.; Briens, M. Dietary hydroxy-selenomethionine helps finishing broilers to cope with heat stress. In Proceedings of the 30th Annual Australian Poultry Science Symposium 2019, Sydney, Australia, 17–20 February 2019. [Google Scholar]
- Juzaitis-Boelter, C.P.; Davis, A.J.; De Marco, M.; Brackenridge, J.K.; Shirley, R.B. Hydroxy-selenomethionine sustains egg production and improves egg selenium deposition in aged laying hens under heat stress. In Proceedings of the 30th Annual Australian Poultry Science Symposium 2019, Sydney, Australia, 17–20 February 2019. [Google Scholar]
- Sun, L.L.; Gao, S.T.; Wang, K.; Xu, J.C.; Sanz-Fernandez, M.V.; Baumgard, L.H.; Bu, D.P. Effects of source on bioavailability of selenium, antioxidant status, and performance in lactating dairy cows during oxidative stress-inducing conditions. J. Dairy Sci. 2019, 102, 311–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sejian, V.; Bhatta, R.; Gaughan, J.B.; Dunshea, F.R.; Lacetera, N. Review: Adaptation of animals to heat stress. Animals 2018, 12, s431–s444. [Google Scholar] [CrossRef] [Green Version]
- Geraert, P.A.; Padilha, J.C.; Guillaumin, S. Metabolic and endocrine changes induced by chronic heat exposure in broiler chickens: Growth performance, body composition and energy retention. Br. J. Nutr. 1996, 75, 195–204. [Google Scholar] [CrossRef]
- Ebrahimzadeh, S.K.; Farhoomand, P.; Noori, K. Immune response of broiler chickens fed diets supplemented with different level of chromium methionine under heat stress conditions. Asian-Austral. J. Anim. 2012, 25, 256. [Google Scholar] [CrossRef] [Green Version]
- Ndubuisi, D.I.; Daudu, O.M.; Abdulrashid, M. The role of selenium in ameliorating the impact of heat stress on growth, hormones, minerals and tibia measurements of broiler chickens. Nigerian J. Anim. Prod. 2021, 48, 176–184. [Google Scholar] [CrossRef]
- Sohail, M.U.; Ijaz, A.; Yousaf, M.S.; Ashraf, K.; Zaneb, H.; Aleem, M.; Rehman, H. Alleviation of cyclic heat stress in broilers by dietary supplementation of mannan-oligosaccharide and Lactobacillus-based probiotic: Dynamics of cortisol, thyroid hormones, cholesterol, C-reactive protein, and humoral immunity. Poult. Sci. 2010, 89, 1934–1938. [Google Scholar] [CrossRef]
- Bakhshalinejad, R.; Kakhki, R.A.M.; Zoidis, E. Effects of different dietary sources and levels of selenium supplements on growth performance, antioxidant status and immune parameters in Ross 308 broiler chickens. Br. Poult. Sci. 2018, 59, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Saunders, P.R.; Santos, J.; Hanssen, N.P.M.; Yates, D.J.A.; Perdue, M.H. Physical and psychological stress in rats enhances colonic epithelial permeability via peripheral CRH. Digest. Dis. Sci. 2002, 47, 208–215. [Google Scholar] [CrossRef]
- Alhenaky, A.; Abdelqader, A.; Abuajamieh, M.; Al-Fataftah, A.R. The effect of heat stress on intestinal integrity and Salmonella invasion in broiler birds. J. Therm. Biol. 2017, 70, 9–14. [Google Scholar] [CrossRef]
- Weber, C.R.; Nalle, S.C.; Tretiakova, M.; Rubin, D.T.; Turner, J.R. Claudin-1 and claudin-2 expression is elevated in inflammatory bowel disease and may contribute to early neoplastic transformation. Lab. Investig. 2008, 88, 1110–1120. [Google Scholar] [CrossRef] [Green Version]
- Tang, J.Y.; Cao, L.; Jia, G.; Liu, G.M.; Chen, X.L.; Tian, G.; Cai, J.Y.; Shang, H.Y.; Zhao, H. The protective effect of selenium from heat stress-induced porcine small intestinal epithelial cell line (IPEC-J2) injury is associated with regulation expression of selenoproteins. Br. J. Ntr. 2019, 122, 1081–1090. [Google Scholar] [CrossRef]
- Gilani, S.; Howarth, G.S.; Kitessa, S.M.; Forder, R.E.A.; Tran, C.D.; Hughes, R.J. New biomarkers for intestinal permeability induced by lipopolysaccharide in chickens. Anim. Prod. Sci. 2016, 56, 1984–1997. [Google Scholar] [CrossRef]
- Cerutti, A.; Chen, K.; Chorny, A. Immunoglobulin Responses at the Mucosal Interface. Annu. Rev. Immunol. 2011, 29, 273–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Handlogten, M.W.; Zhu, M.; Ahuja, S. Intracellular response of CHO cells to oxidative stress and its influence on metabolism and antibody production. Biochem. Eng. J. 2018, 133, 12–20. [Google Scholar] [CrossRef]
- Yu, K.; Lieber, M.R. Current insights into the mechanism of mammalian immunoglobulin class switch recombination. Crit. Rev. Biochem. Mol. Biol. 2019, 54, 333–351. [Google Scholar] [CrossRef]
- Eisen, H.N. Affinity enhancement of antibodies: How low-affinity antibodies produced early in immune responses are followed by high-affinity antibodies later and in memory B-cell responses. Cancer Immunol. Res. 2014, 2, 381–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlander, D.; Stålberg, J.; Larsson, A. Chicken Antibodies. Upsala J. Med. Sci. 1999, 104, 179–189. [Google Scholar] [CrossRef]
- Bassiouni, A.A.; Zaki, M.M.; Hady, M.M. Effect of vitamin E and selenium on the immune response of chickens against living Newcastle disease vaccine. Vet. Med. J. 1990, 38, 145–155. [Google Scholar]
- Niu, Z.Y.; Liu, F.Z.; Yan, Q.L.; Li, L. Effects of different levels of selenium on growth performance and immunocompetence of broilers under heat stress. Arch. Anim. Nutr. 2009, 63, 56–65. [Google Scholar] [CrossRef]
- Kovalenko, M.V.; Stepchenko, L.M.; Shevtsova, A.I.; Brazaluk, O.Z.; Surai, P.F. Effect of selenium-containing supplements on the indices of specific immunity and nonspecific resistance in chicken. Fiziol. Zh. 2008, 54, 69–73. [Google Scholar] [PubMed]
- Caroprese, M.; Bradford, B.; Rhoads, R.P. Impact of Climate Change on Immune Responses in Agricultural Animals. Front. Vet. Sci. 2021, 8, 732203. [Google Scholar] [CrossRef]
- Matsushita, T. Regulatory and effector B cells: Friends or foes? J. Dermatol. Sci. 2019, 93, 2–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nettford, S.K.; Prabhu, K.S. Selenium and Selenoproteins in Gut Inflammation—A Review. Antioxidants 2018, 7, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, R.M.; Sunde, R.A. Selenoprotein Transcript Level and Enzyme Activity as Biomarkers for Selenium Status and Selenium Requirements of Chickens (Gallus gallus). PLoS ONE 2016, 11, e0151665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seyedali, A.; Berry, M.J. Nonsense-mediated decay factors are involved in the regulation of selenoprotein mRNA levels during selenium deficiency. RNA 2014, 20, 1248–1256. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SS | SY | OH-SeMet | |
|---|---|---|---|
| d 1 to 21 | |||
| BWG, g/bird | 579 ± 30 a | 578 ± 27 a | 607 ± 39 b |
| FI, g/bird | 810 ± 34 | 807 ± 32 | 830 ± 36 |
| FCR, g/g | 1.40 ± 0.03 a | 1.40 ± 0.03 a | 1.37 ± 0.03 b |
| Mortality, % | 0 ± 0 | 0.9 ± 3.2 | 0 ± 0 |
| d 22 to 42 | |||
| BWG, g/bird | 958 ± 96 | 953 ± 142 | 987 ± 113 |
| FI, g/bird | 1855 ± 144 | 1823 ± 193 | 1894 ±194 |
| FCR, g/g | 1.94 ± 0.07 | 1.93 ± 0.10 | 1.92 ± 0.06 |
| Mortality, % | 3.7 ± 5.5 | 3.7 ± 5.5 | 3.7 ± 7.2 |
| d 1 to 42 | |||
| BWG, g/bird | 1537 ± 120 | 1535 ± 160 | 1594 ± 139 |
| FI, g/bird | 2665 ± 174 | 2633 ± 216 | 2724 ± 223 |
| FCR, g/g | 1.74 ± 0.04 a | 1.72 ± 0.06 ab | 1.71 ± 0.03 b |
| Mortality, % | 3.7 ± 5.5 | 4.6 ± 5.5 | 3.7 ± 7.2 |
| SS | SY | OH-SeMet | |
|---|---|---|---|
| Hormones | |||
| Cortisol, ng/mL | 22.8 ± 2.6 a | 20.3 ± 2.5 b | 18.9 ± 1.6 b |
| T3, nmol/L | 1.08 ± 0.07 a | 0.91 ± 0.13 b | 0.84 ± 0.09 b |
| T4, nmol/L | 31.6 ± 2.7 | 31.7 ± 2.6 | 31.6 ± 2.6 |
| Cytokines | |||
| IL-6, pg/mL | 5.4 ± 0.6 a | 4.9 ± 0.4 b | 4.3 ± 0.7 b |
| IL-10, pg/mL | 8.7 ± 1.9 b | 9.2 ± 1.4 b | 12.4 ± 1.5 a |
| TNF-a, pg/mL | 12.9 ± 3.1 | 11.4 ± 1.4 | 12.5 ± 2.2 |
| Immunoglobulin | |||
| IgA, ug/mL | 53.7 ± 4.4 a | 44.5 ± 4.1 b | 39.3 ± 4.5 c |
| IgG, ug/mL | 268 ± 55 b | 256 ± 29 b | 363 ± 28 a |
| IgM, ug/mL | 100.5 ± 10.7 a | 85.4 ± 9.7 b | 82.0 ± 11.9 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, H.; Zhao, L.; Xu, Z.-J.; De Marco, M.; Briens, M.; Yan, X.-H.; Sun, L.-H. Hydroxy-Selenomethionine Improves the Selenium Status and Helps to Maintain Broiler Performances under a High Stocking Density and Heat Stress Conditions through a Better Redox and Immune Response. Antioxidants 2021, 10, 1542. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10101542
Sun H, Zhao L, Xu Z-J, De Marco M, Briens M, Yan X-H, Sun L-H. Hydroxy-Selenomethionine Improves the Selenium Status and Helps to Maintain Broiler Performances under a High Stocking Density and Heat Stress Conditions through a Better Redox and Immune Response. Antioxidants. 2021; 10(10):1542. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10101542
Chicago/Turabian StyleSun, Hua, Ling Zhao, Zi-Jian Xu, Michele De Marco, Mickael Briens, Xiang-Hua Yan, and Lv-Hui Sun. 2021. "Hydroxy-Selenomethionine Improves the Selenium Status and Helps to Maintain Broiler Performances under a High Stocking Density and Heat Stress Conditions through a Better Redox and Immune Response" Antioxidants 10, no. 10: 1542. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10101542