
Organic Acids Improve Growth Performance with Potential Regulation of Redox Homeostasis, Immunity, and Microflora in Intestines of Weaned Piglets

Abstract
:1. Introduction
2. Materials and Methods
2.1. Piglets, Dietary Treatments, and Sample Collections
2.2. Redox Status, Digestive Enzyme Activity, and Histologic Analysis
2.3. Real-Time q-PCR Analyses
2.4. Pyrosequencing of Bacterial 16S rRNA Gene Amplification
2.5. Statistical Analyses
3. Results
3.1. Growth Performance
3.2. Serum Redox Status
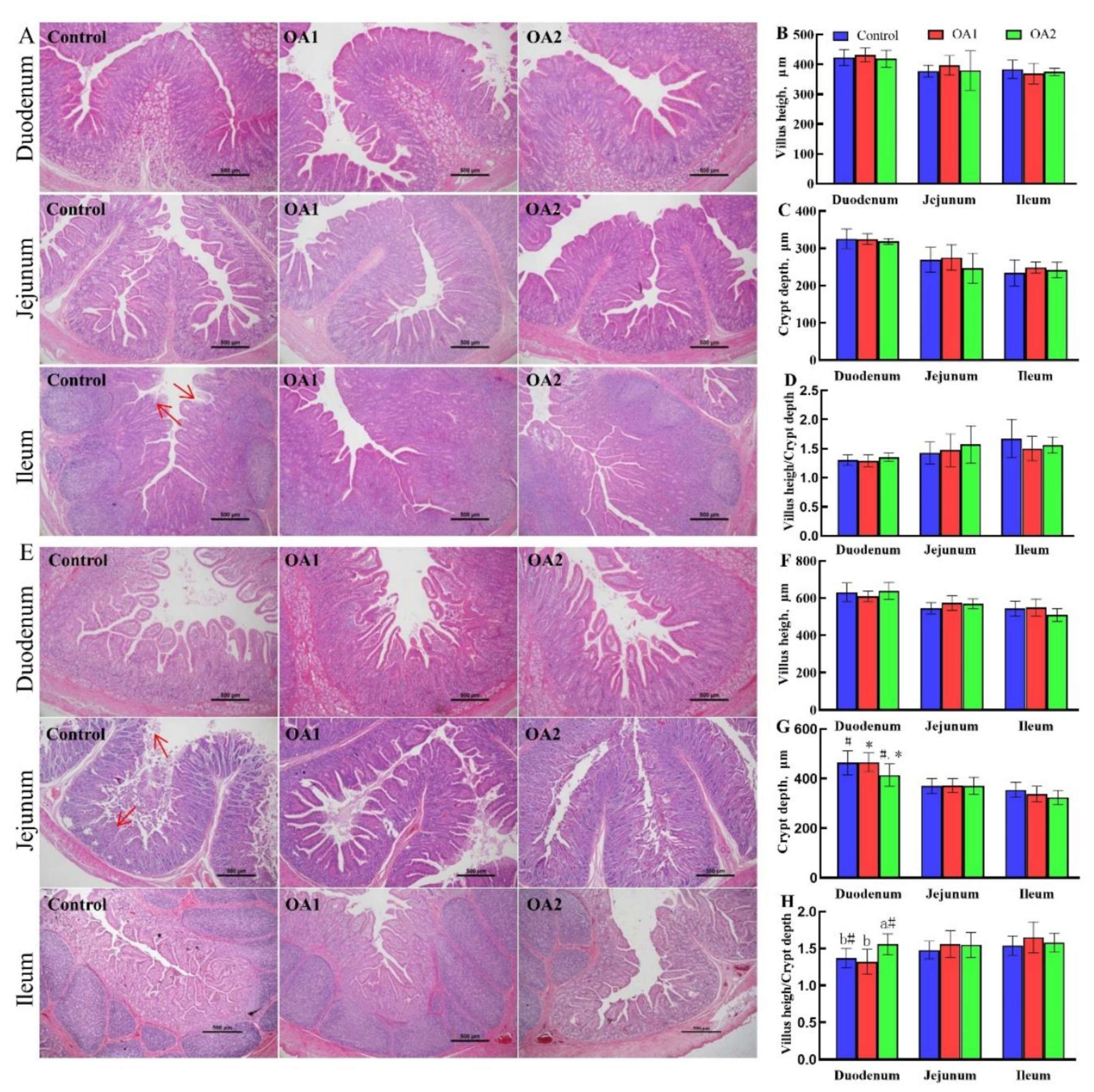
3.3. Small Intestinal Histology and Chyme Digestive Enzyme Activity
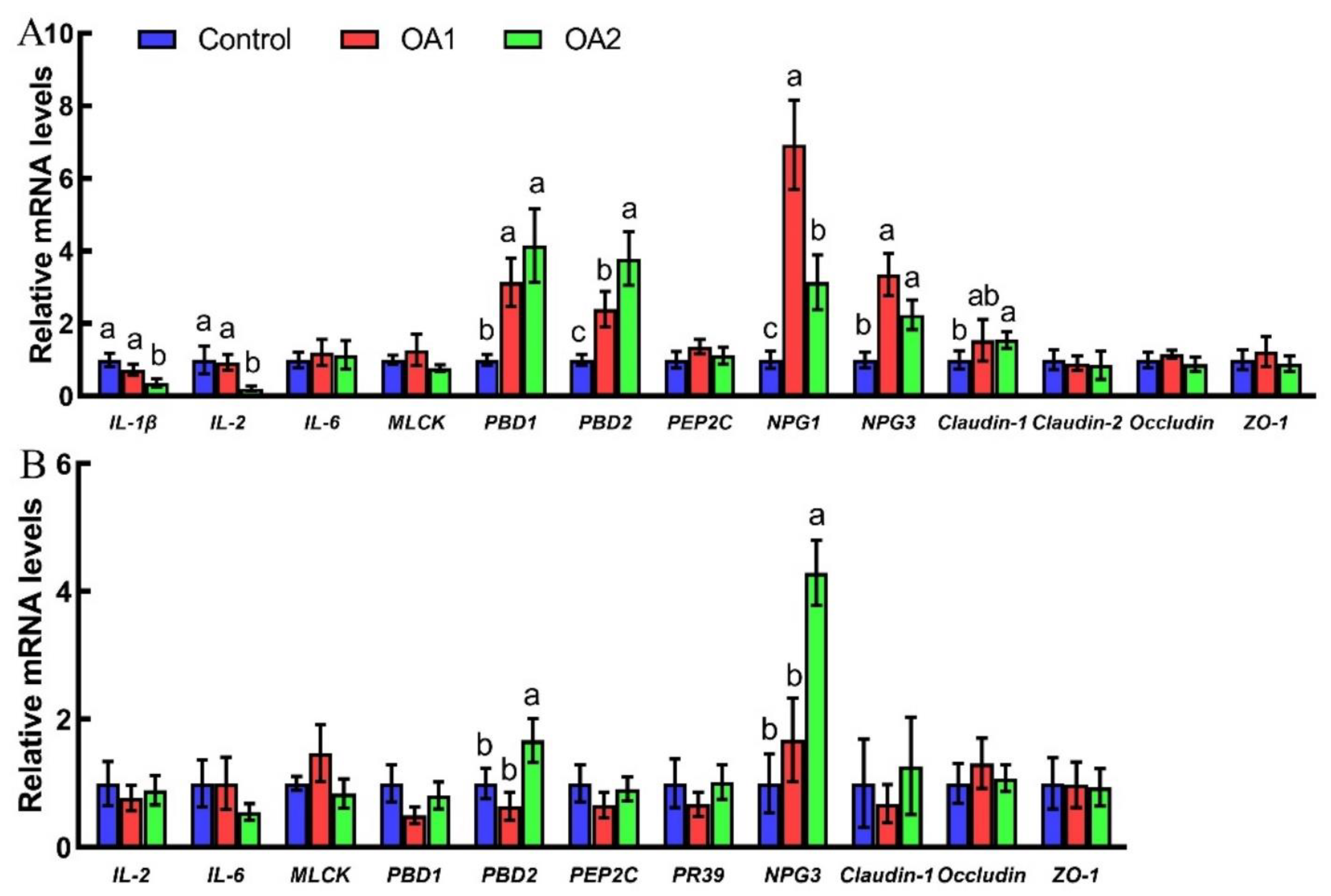
3.4. Expression of Cytokine-, Tight Junction-, and Host Defense Peptide-Related Genes
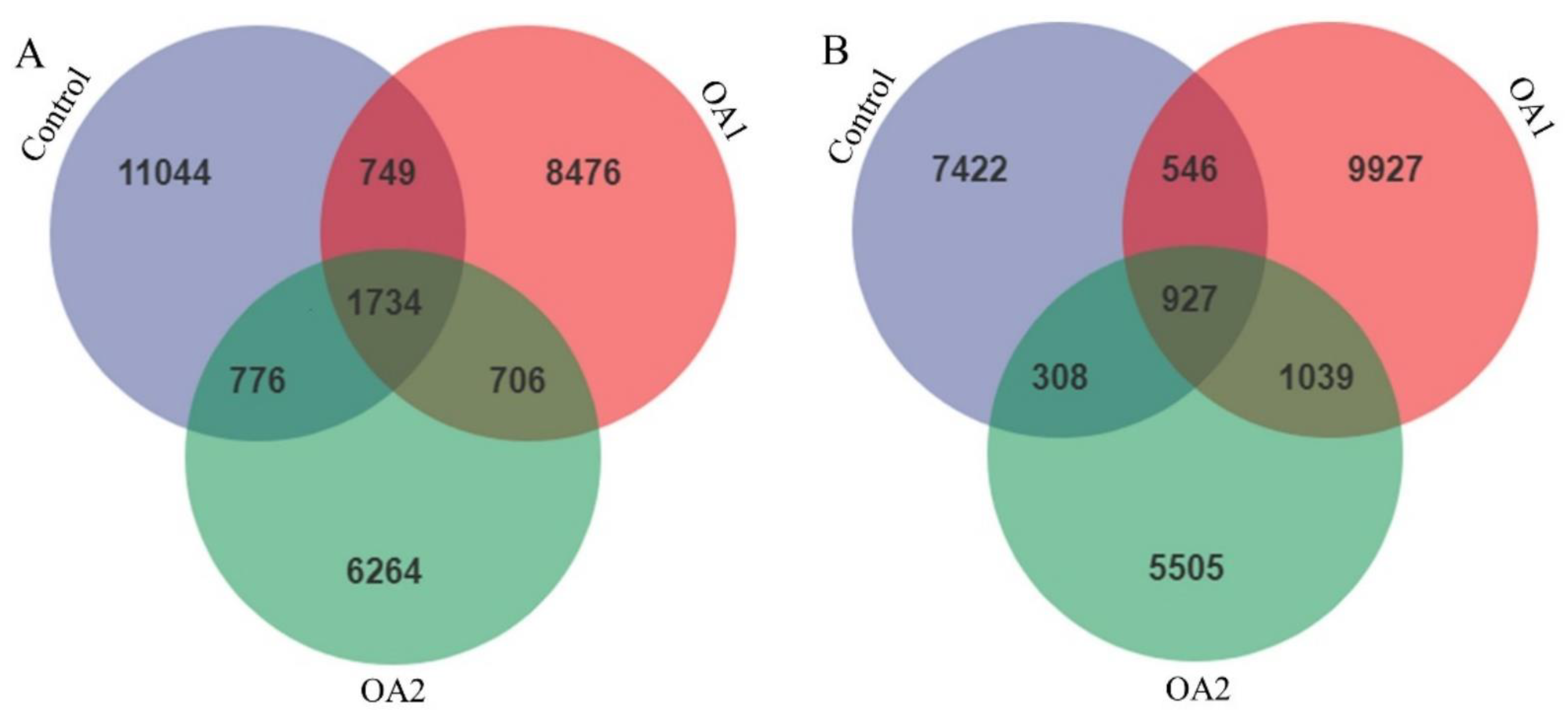
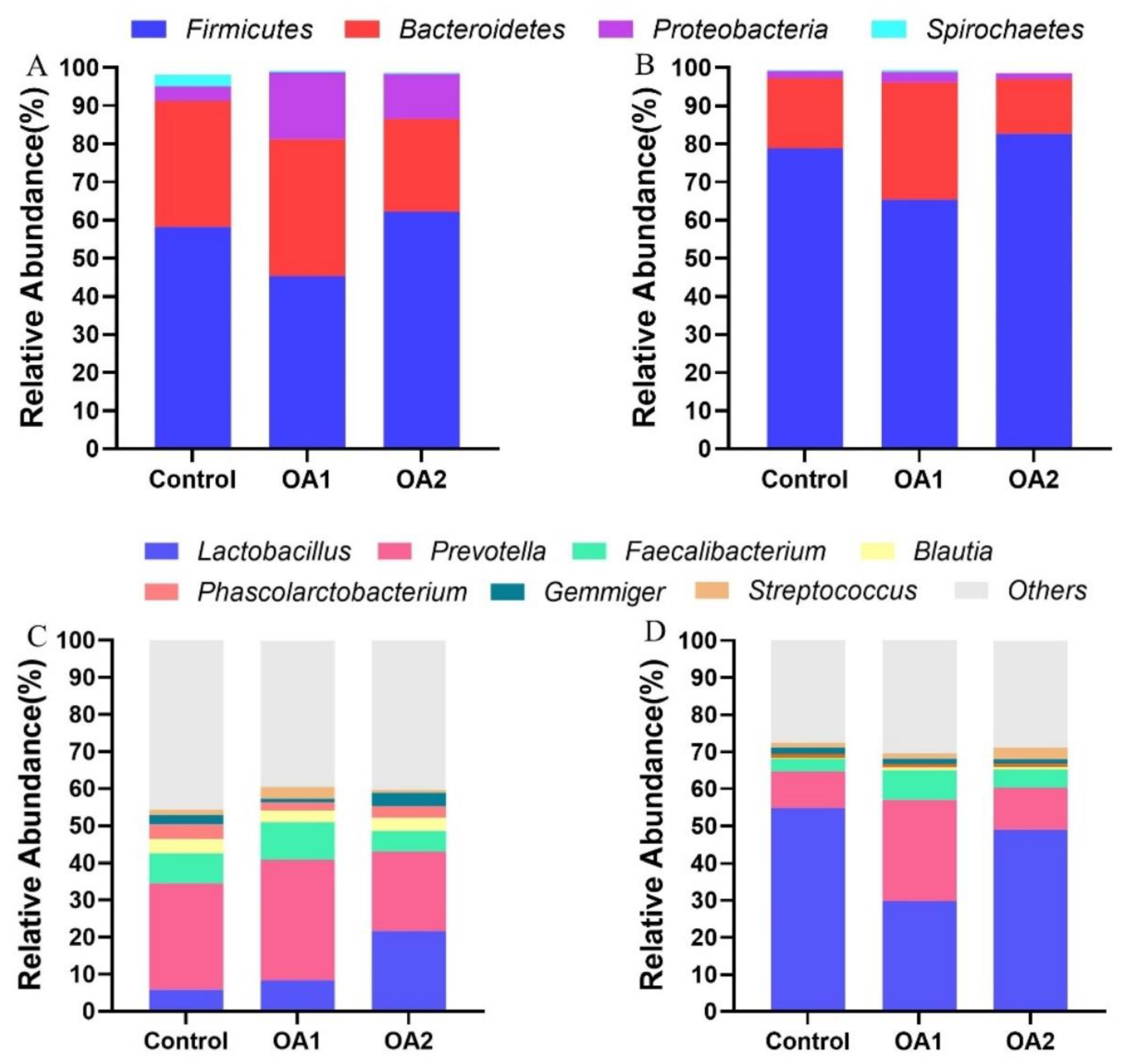
3.5. Gut Microbiota
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Laine, T.M.; Lyytikainen, T.; Yliaho, M.; Anttila, M. Risk factors for post-weaning diarrhoea on piglet producing farms in Finland. Acta Vet. Scand. 2008, 50, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Lee, K.Y.; Kim, I.H. Effects of dietary protected organic acids on growth performance, nutrient digestibility, fecal microflora, diarrhea score, and fecal gas emission in weanling pigs. Can J. Anim. Sci. 2019, 99, 514–520. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.X.; Yao, W.L.; Li, J.; Shao, Y.F.; He, Q.Y.; Xia, J.; Huang, F.R. Dietary garcinol supplementation improves diarrhea and intestinal barrier function associated with its modulation of gut microbiota in weaned piglets. J. Anim. Sci. Biotechnol. 2020, 11, 12. [Google Scholar] [CrossRef] [Green Version]
- Luise, D.; Bertocchi, M.; Motta, V.; Salvarani, C.; Bosi, P.; Luppi, A.; Fanelli, F.; Mazzoni, M.; Archetti, I.; Maiorano, G.; et al. Bacillus sp. probiotic supplementation diminish the Escherichia coli F4ac infection in susceptible weaned pigs by influencing the intestinal immune response, intestinal microbiota and blood metabolomics. J. Anim. Sci. Biotechnol. 2019, 10, 74. [Google Scholar] [CrossRef] [PubMed]
- Suiryanrayna, M.V.A.N.; Ramana, J.V. A review of the effects of dietary organic acids fed to swine. J. Anim. Sci. Biotechnol. 2015, 6, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pluske, J.R. Feed- and feed additives-related aspects of gut health and development in weanling pigs. J. Anim. Sci. Biotechnol. 2013, 4, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.S.; Tang, C.H.; Zhao, Q.Y.; Zhan, T.F.; Zhang, K.; Han, Y.M.; Zhang, J.M. Effects of dietary supplementation with combinations of organic and medium chain fatty acids as replacements for chlortetracycline on growth performance, serum immunity, and fecal microbiota of weaned piglets. Livest Sci. 2018, 216, 210–218. [Google Scholar] [CrossRef]
- Degroote, J.; Vergauwen, H.; Van Noten, N.; Wang, W.; De Smet, S.; Van Ginneken, C.; Michiels, J. The Effect of Dietary Quercetin on the Glutathione Redox System and Small Intestinal Functionality of Weaned Piglets. Antioxidants 2019, 8, 312. [Google Scholar] [CrossRef] [Green Version]
- Grilli, E.; Tugnoli, B.; Passey, J.L.; Stahl, C.H.; Piva, A.; Moeser, A.J. Impact of dietary organic acids and botanicals on intestinal integrity and inflammation in weaned pigs. BMC Vet. Res. 2015, 11, 96. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Song, P.X.; Fan, P.X.; Hou, C.L.; Thacker, P.; Ma, X. Dietary Sodium Butyrate Decreases Postweaning Diarrhea by Modulating Intestinal Permeability and Changing the Bacterial Communities in Weaned Piglets. J. Nutr. 2015, 145, 2774–2780. [Google Scholar] [CrossRef]
- Long, S.F.; Xu, Y.T.; Pan, L.; Wang, Q.Q.; Wang, C.L.; Wu, J.Y.; Wu, Y.Y.; Han, Y.M.; Yun, C.H.; Piao, X.S. Mixed organic acids as antibiotic substitutes improve performance, serum immunity, intestinal morphology and microbiota for weaned piglets. Anim. Feed Sci. Technol. 2018, 235, 23–32. [Google Scholar] [CrossRef]
- Luise, D.; Correa, F.; Bosi, P.; Trevisi, P. A Review of the Effect of Formic Acid and Its Salts on the Gastrointestinal Microbiota and Performance of Pigs. Animals 2020, 10, 887. [Google Scholar] [CrossRef]
- Partanen, K.H.; Mroz, Z. Organic acids for performance enhancement in pig diets. Nutr. Res. Rev. 1999, 12, 117–145. [Google Scholar] [CrossRef] [Green Version]
- Oleskin, A.V.; Shenderov, B.A. Microbial Communication and Microbiota-Host Interactions: Biomedical, Biotechnological, and Biopolitical Implications; Nova Science Publishers: Hauppauge, NY, USA, 2020. [Google Scholar]
- Oleskin, A.V.; Shenderov, B.A.; Rogovsky, V.S. Role of Neurochemicals in the Interaction between the Microbiota and the Immune and the Nervous System of the Host Organism. Probiotics Antimicrob. Proteins 2017, 9, 215–234. [Google Scholar] [CrossRef]
- Oleskin, A.V.; Shenderov, B.A. Neuromodulatory effects and targets of the SCFAs and gasotransmitters produced by the human symbiotic microbiota. Microb. Ecol. Health Dis. 2016, 27, 30971. [Google Scholar] [CrossRef] [PubMed]
- Sofos, J.N.; Pierson, M.D.; Blocher, J.C.; Busta, F.F. Mode of Action of Sorbic Acid on Bacterial-Cells and Spores. Int. J. Food Microbiol. 1986, 3, 1–17. [Google Scholar] [CrossRef]
- Zhao, L.; Deng, J.; Xu, Z.J.; Zhang, W.P.; Khalil, M.M.; Karrow, N.A.; Sun, L.H. Mitigation of Aflatoxin B1 Hepatoxicity by Dietary Hedyotis diffusa Is Associated with Activation of NRF2/ARE Signaling in Chicks. Antioxidants 2021, 10, 878. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.H.; Zhang, N.Y.; Zhu, M.K.; Zhao, L.; Zhou, J.C.; Qi, D.S. Prevention of Aflatoxin B1 Hepatoxicity by Dietary Selenium Is Associated with Inhibition of Cytochrome P450 Isozymes and Up-Regulation of 6 Selenoprotein Genes in Chick Liver. J. Nutr. 2015, 146, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.C.; Chen, P.; Zhang, C.; Khalil, M.M.; Zhang, N.Y.; Qi, D.S.; Wang, Y.W.; Sun, L.H. Yeast culture promotes the production of aged laying hens by improving intestinal digestive enzyme activities and the intestinal health status. Poult. Sci. 2020, 99, 2026–2032. [Google Scholar] [CrossRef]
- Kim, K.; He, Y.; Xiong, X.; Ehrlich, A.; Li, X.; Raybould, H.; Atwill, E.R.; Maga, E.A.; Jørgensen, J.; Liu, Y. Dietary supplementation of Bacillus subtilis influenced intestinal health of weaned pigs experimentally infected with a pathogenic E. coli. J. Anim. Sci. Biotechnol. 2019, 10, 52. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Domig, K.J.; Ettle, T.; Windisch, W.; Mair, C.; Schedle, K. Evaluation of potential reference genes for relative quantification by RT-qPCR in different porcine tissues derived from feeding studies. Int. J. Mol. Sci. 2011, 12, 1727–1734. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Zhang, L.; Chu, X.H.; Ma, R.; Wang, Y.W.; Liu, Q.; Zhang, N.Y.; Karrow, N.A.; Sun, L.H. Effects of deoxynivalenol on the porcine growth performance and intestinal microbiota and potential remediation by a modified HSCAS binder. Food Chem. Toxicol. 2020, 141, 111373. [Google Scholar] [CrossRef]
- Li, Z.; Yi, G.; Yin, J.; Sun, P.; Li, D.; Knight, C. Effects of organic acids on growth performance, gastrointestinal pH, intestinal microbial populations and immune responses of weaned pigs. Asian-Australas J. Anim. Sci. 2008, 21, 252–261. [Google Scholar] [CrossRef]
- Overland, M.; Kjos, N.P.; Borg, M.; Skjerve, E.; Sorum, H. Organic acids in diets for entire male pigs: Effect on skatole level, microbiota in digesta, and growth performance. Livest Sci. 2008, 115, 169–178. [Google Scholar] [CrossRef]
- Nguyen, D.H.; Kim, I.H. Protected Organic Acids Improved Growth Performance, Nutrient Digestibility, and Decreased Gas Emission in Broilers. Animals 2020, 10, 416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.; Toghyani, M.; Kheravii, S.K.; Pineda, L.; Han, Y.; Swick, R.A.; Wu, S.B. Potential of blended organic acids to improve performance and health of broilers infected with necrotic enteritis. Anim. Nutr. 2021, 7, 440–449. [Google Scholar] [CrossRef]
- Dai, D.; Qiu, K.; Zhang, H.J.; Wu, S.G.; Han, Y.M.; Wu, Y.Y.; Qi, G.H.; Wang, J. Organic Acids as Alternatives for Antibiotic Growth Promoters Alter the Intestinal Structure and Microbiota and Improve the Growth Performance in Broilers. Front. Microbiol. 2021, 11, 618144. [Google Scholar] [CrossRef]
- Zhu, K.H.; Xu, X.R.; Sun, D.F.; Tang, J.L.; Zhang, Y.K. Effects of drinking water acidification by organic acidifier on growth performance, digestive enzyme activity and caecalbacteria in growing rabbits. Anim. Feed Sci. Technol. 2014, 190, 87–94. [Google Scholar] [CrossRef]
- Van Bunnik, B.A.D.; Hagenaars, T.J.; Bolder, N.M.; Nodelijk, G.; de Jong, M.C.M. Interaction effects between sender and receiver processes in indirect transmission of Campylobacter jejuni between broilers. BMC Vet. Res. 2012, 8, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, C.C.; Yang, S.F.; Zhu, L.H.; Cai, X.; Sheng, Y.S.; Zhu, S.W.; Xu, J.X. Regulation of N-acetyl cysteine on gut redox status and major microbiota in weaned piglets. J. Anim. Sci. 2014, 92, 1504–1511. [Google Scholar] [CrossRef] [Green Version]
- Vergauwen, H.; Degroote, J.; Prims, S.; Wang, W.; Fransen, E.; De Smet, S.; Casteleyn, C.; Van Cruchten, S.; Michiels, J.; Van Ginneken, C. Artificial rearing influences the morphology, permeability and redox state of the gastrointestinal tract of low and normal birth weight piglets. J. Anim. Sci. Biotechnol. 2017, 8, 30. [Google Scholar] [CrossRef] [Green Version]
- Wen, C.Y.; Li, F.N.; Guo, Q.P.; Zhang, L.Y.; Duan, Y.H.; Wang, W.L.; Li, J.Z.; He, S.P.; Chen, W.; Yin, Y.L. Protective effects of taurine against muscle damage induced by diquat in 35 days weaned piglets. J. Anim. Sci. Biotechnol. 2020, 11, 56. [Google Scholar] [CrossRef]
- Corino, C.; Prost, M.; Pizzi, B.; Rossi, R. Dietary Plant Extracts Improve the Antioxidant Reserves in Weaned Piglets. Antioxidants 2021, 10, 702. [Google Scholar] [CrossRef] [PubMed]
- Milton, J.J.; Roger, B.; Sabine, S.; Miriam, H.; Waldiceu, V.; Ana, P. Ingestion of organic acids and cinnamaldehyde improves tissue homeostasis of piglets exposed to enterotoxic Escherichia coli (ETEC). J. Anim. Sci. 2020, 98, skaa012. [Google Scholar]
- Chen, J.; Guo, J.; Guan, W.T.; Song, J.J.; Deng, Z.X.; Cheng, L.; Deng, Y.L.; Chen, F.; Zhang, S.H.; Zhang, Y.Z.; et al. Effect of pad-fan cooling and dietary organic acid supplementation during late gestation and lactation on reproductive performance and antioxidant status of multiparous sows in hot weather. Trop. Anim. Health Prod. 2018, 50, 973–982. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Dai, D.; Zhang, H.J.; Wu, S.G.; Han, Y.M.; Wu, Y.Y.; Qi, G.H. Organic acids modulate systemic metabolic perturbation caused by salmonella pullorum challenge in early-stage broilers. Front. Physiol. 2019, 10, 1418. [Google Scholar] [CrossRef]
- Sun, X.; Cui, Y.; Su, Y.; Gao, Z.; Diao, X.; Li, J.; Zhu, X.; Li, D.; Li, Z.; Wang, C.; et al. Dietary Fiber Ameliorates Lipopolysaccharide-Induced Intestinal Barrier Function Damage in Piglets by Modulation of Intestinal Microbiome. Msystems 2021, 6, e01374-20. [Google Scholar] [CrossRef] [PubMed]
- Moeser, A.J.; Pohl, C.S.; Rajput, M. Weaning stress and gastrointestinal barrier development: Implications for lifelong gut health in pigs. Anim. Nutr. 2017, 3, 313–321. [Google Scholar] [CrossRef]
- Zhang, Z.; Jiang, Z.; Lin, Y.; Zheng, L.; Yu, D. Effect of Stress From Early Weaning Pig on Its Digestive Enzymes Activity. Chin. J. Anim. Husb. 1999, 35, 6–8. [Google Scholar]
- Xu, Y.T.; Lahaye, L.; He, Z.X.; Zhang, J.X.; Yang, C.B.; Piao, X.S. Micro-encapsulated essential oils and organic acids combination improves intestinal barrier function, inflflammatory responses and microbiota of weaned piglets challenged with enterotoxigenic Escherichia coli F4(K88+). Anim. Nutr. 2020, 6, 269–277. [Google Scholar] [CrossRef]
- Jackman, J.A.; Boyd, R.D.; Elrod, C.C. Medium-chain fatty acids and monoglycerides as feed additives for pig production: Towards gut health improvement and feed pathogen mitigation. J. Anim. Sci. Biotechnol. 2020, 11, 44. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Huang, N.N.; Xiong, J.; Wei, H.K.; Jiang, S.W.; Peng, J. Caprylic acid and nonanoic acid upregulate endogenous host defense peptides to enhance intestinal epithelial immunological barrier function via histone deacetylase inhibition. Int. Immunopharmacol. 2018, 65, 303–311. [Google Scholar] [CrossRef]
- Zhou, Z.Z.; Huang, J.; Hao, H.H.; Wei, H.K.; Zhou, Y.F.; Peng, J. Applications of new functions for inducing host defense peptides and synergy sterilization of medium chain fatty acids in substituting in-feed antibiotics. J. Funct. Foods 2019, 52, 348–359. [Google Scholar] [CrossRef]
- Xu, Y.T.; Liu, L.; Long, S.F.; Pan, L.; Piao, X.S. Effect of organic acids and essential oils on performance, intestinal health and digestive enzyme activities of weaned pigs. Anim. Feed Sci. Technol. 2018, 235, 110–119. [Google Scholar] [CrossRef]
- Qiu, Y.Q.; Liu, S.L.; Hou, L.; Li, K.B.; Wang, L.; Gao, K.G.; Yang, X.F.; Jiang, Z.Y. Supplemental Choline Modulates Growth Performance and Gut Inflammation by Altering the Gut Microbiota and Lipid Metabolism in Weaned Piglets. J. Nutr. 2021, 151, 20–29. [Google Scholar] [CrossRef]
- Wang, J.; Han, Y.; Meng, F.; Zhao, J.; Zhou, Z.; Fan, H. Fecal microbiota succession of piglets from birth to post-weaning by 454 pyrosequencing analysis. Trans. Tianjin Univ. 2017, 23, 211–220. [Google Scholar] [CrossRef]
- Guevarra, R.B.; Hong, S.H.; Cho, J.H.; Kim, B.R.; Shin, J.; Lee, J.H.; Kang, B.N.; Kim, Y.H.; Wattanaphansak, S.; Isaacson, R.E.; et al. The dynamics of the piglet gut microbiome during the weaning transition in association with health and nutrition. J. Anim. Sci. Biotechnol. 2018, 9, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.M.; Mao, B.Y.; Gu, J.Y.; Wu, J.Y.; Cui, S.M.; Wang, G.; Zhao, J.X.; Zhang, H.; Chen, X. Blautia—a new functional genus with potential probiotic properties? Gut Microbes 2021, 13, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Shahi, S.K.; Freedman, S.N.; Mangalam, A.K. Gut microbiome in multiple sclerosis: The players involved and the roles they play. Gut Microbes 2017, 8, 607–615. [Google Scholar] [CrossRef] [PubMed]
- Polansky, O.; Sekelova, Z.; Faldynova, M.; Sebkova, A.; Sisak, F.; Rychlik, I. Important Metabolic Pathways and Biological Processes Expressed by Chicken Cecal Microbiota. J. Appl. Environ. Microbiol. 2016, 82, 1569–1576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, A.; Lordan, C.; Ross, R.P.; Cotter, P.C. Gut microbes from the phylogenetically diverse genus Eubacterium and their various contributions to gut health. Gut Microbes 2020, 12, 28. [Google Scholar] [CrossRef] [PubMed]
- Tugnoli, B.; Giovagnoni, G.; Piva, A.; Grilli, E. From Acidifiers to Intestinal Health Enhancers: How Organic Acids Can Improve Growth Efficiency of Pigs. Animals 2020, 10, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.R.; Xu, Y.S.; Ji, M.M.; Zhang, L.; Li, D.; Lang, Q.; Zhang, L.; Ji, G.; Liu, B.C. Association of the oral microbiome with the progression of impaired fasting glucose in a Chinese elderly population. J. Oral Microbiol. 2019, 11, 1605789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patra, A.K.; Park, T.; Braun, H.S.; Geiger, S.; Pieper, R.; Yu, Z.T.; Aschenbach, J.R. Dietary Bioactive Lipid Compounds Rich in Menthol Alter Interactions Among Members of Ruminal Microbiota in Sheep. Front. Microbiol. 2019, 10, 2038. [Google Scholar] [CrossRef] [PubMed]
- Riviere, A.; Selak, M.; Lantin, D.; Leroy, F.; De Vuyst, L. Bifidobacteria and Butyrate-Producing Colon Bacteria: Importance and Strategies for Their Stimulation in the Human Gut. Front. Microbiol. 2016, 7, 979. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
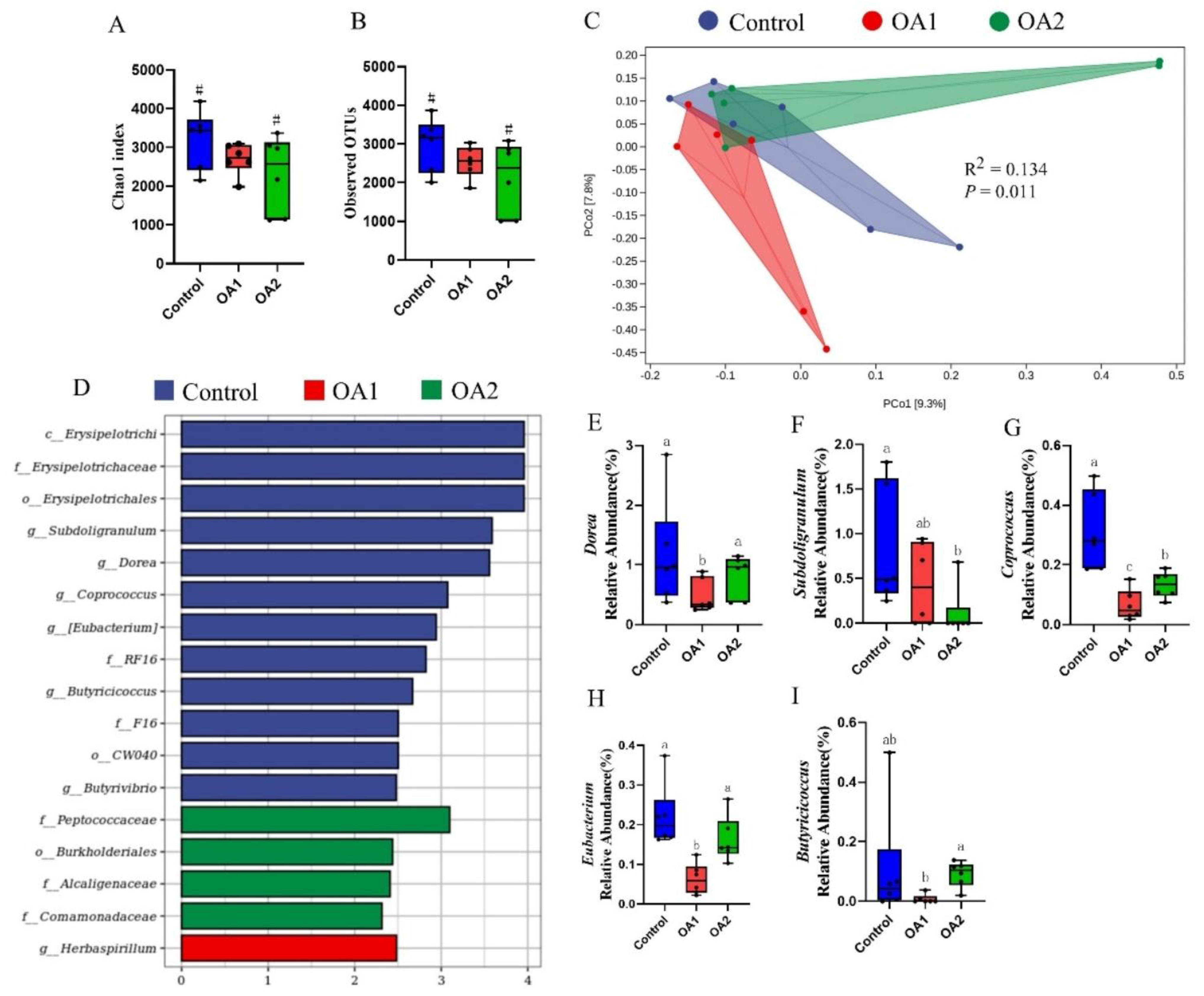{kind=link}
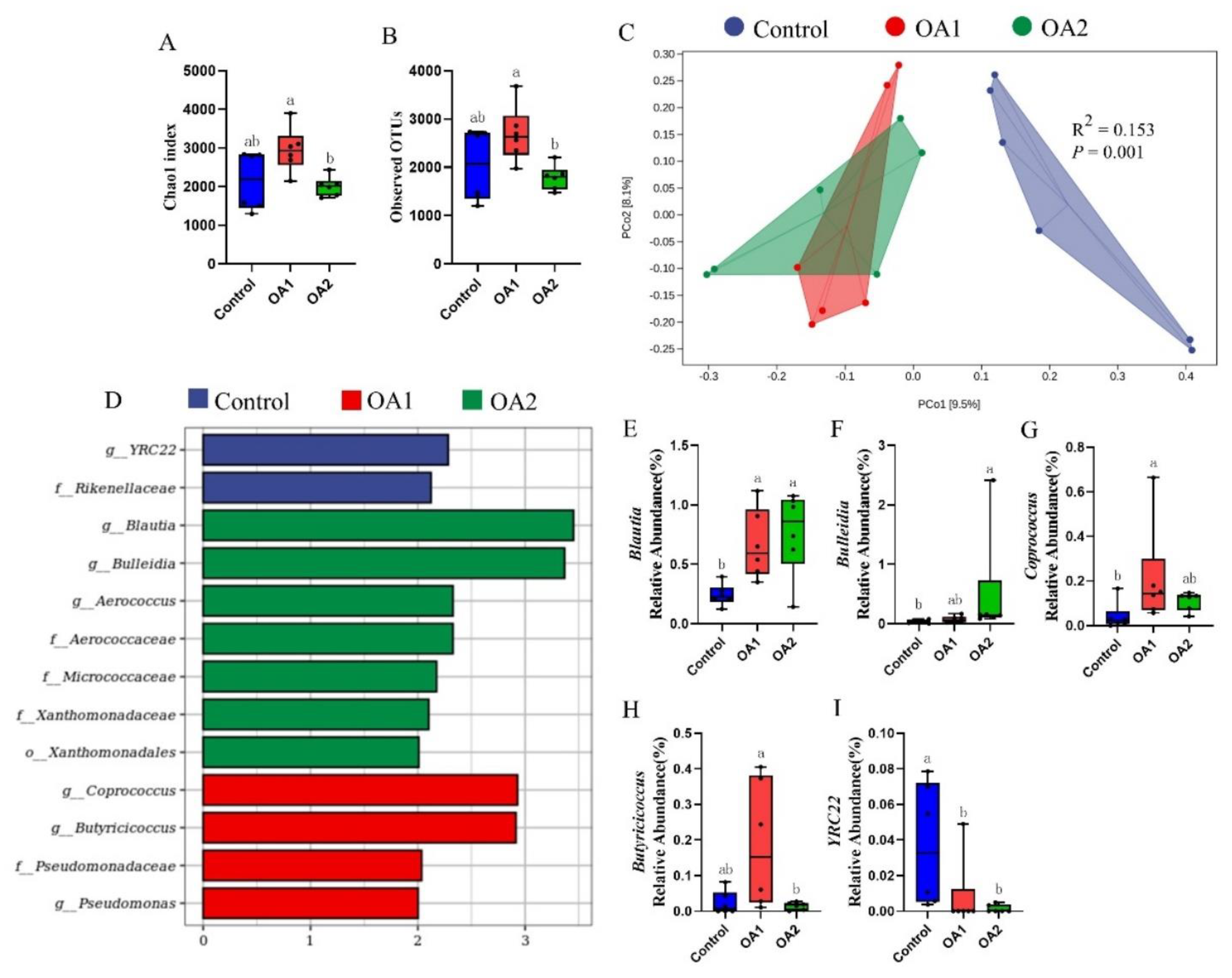{kind=link}
| Control | OA1 | OA2 | p-Value | |
|---|---|---|---|---|
| Days 1–14 | ||||
| Feed intake, kg/piglet | 2.92 ± 0.25 b | 3.24 ± 0.45 a | 2.89 ± 0.24 b | 0.02 |
| Body weight gain, kg/piglet | 1.68 ± 0.17 b | 1.90 ± 0.24 a | 1.69 ± 0.16 b | 0.02 |
| Feed/gain ratio, kg/kg | 1.75 ± 0.16 | 1.71 ± 0.18 | 1.72 ± 0.24 | 0.88 |
| Diarrhea rate, % | 10.2 ± 3.1 a | 9.6 ± 2.9 a | 7.0 ± 2.7 b | 0.03 |
| Mortality rate, % | 0.83 ± 2.89 | 0.00 ± 0.00 | 1.67 ± 3.89 | 0.36 |
| Days 15–49 | ||||
| Feed intake, kg/piglet | 24.0 ± 1.3 a,b | 24.4 ± 1.7 a | 23.1 ± 1.3 b | 0.08 |
| Body weight gain, kg/piglet | 16.7 ± 0.8 | 17.3 ± 1.2 # | 16.4 ± 1.0 # | 0.13 |
| Feed/gain ratio, kg/kg | 1.43 ± 0.05 # | 1.41 ± 0.05 | 1.40 ± 0.03 # | 0.23 |
| Diarrhea rate, % | 1.27 ± 0.73 a | 1.16 ± 0.81 a | 0.59 ± 0.54 b | 0.05 |
| Mortality rate, % | 0.83 ± 2.89 | 3.33 ± 4.92 | 2.50 ± 4.52 | 0.35 |
| Days 1–49 | ||||
| Feed intake, kg/piglet | 26.9 ± 1.5 a,b | 27.7 ± 1.9 a | 26.0 ± 1.4 b | 0.05 |
| Body weight gain, kg/piglet | 18.4 ± 0.9 b,# | 19.2 ± 1.3 a,# | 18.1 ± 1.1 b | 0.06 |
| Feed/gain ratio, kg/kg | 1.46 ± 0.05 | 1.44 ± 0.05 | 1.43 ± 0.04 | 0.26 |
| Diarrhea rate, % | 4.01 ± 1.03 a | 3.80 ±1.32 a | 2.58 ± 0.98 b | 0.01 |
| Mortality rate, % | 1.67 ± 3.89 | 3.33 ± 4.92 | 4.17 ± 6.69 | 0.51 |
| Control | OA1 | OA2 | p value | |
|---|---|---|---|---|
| Day 14 | ||||
| T-AOC, U/L | 125 ± 24 b | 150 ± 36 a,b | 163 ± 30.4 a | 0.13 |
| GPX, U/mL | 635 ± 57 | 594 ± 49 | 664 ± 123 | 0.36 |
| SOD, U/mL | 37.6 ± 4.0 | 39.7 ± 4.0 | 40.7 ± 4.7 | 0.45 |
| CAT, U/mL | 3.9 ± 1.2 # | 4.9 ± 0.3 # | 5.2 ± 2.4 | 0.35 |
| MDA, nmol/mL | 2.3 ± 0.9 | 2.7 ± 0.7 | 4.1 ± 2.1 | 0.13 |
| PC, nmol/mL | 1.3 ± 0.8 | 0.9 ± 0.4 | 1.2 ± 0.4 | 0.67 |
| Day 49 | ||||
| T-AOC, U/L | 189 ± 22 # | 197 ± 26 | 211 ± 19 # | 0.27 |
| GPX, U/mL | 889 ± 89 | 908 ± 45 | 855 ± 57 | 0.40 |
| SOD, U/mL | 41.4 ± 1.0 b | 47.0 ± 1.6 a | 47.2 ± 2.1 a | 0.00 |
| CAT, U/mL | 2.5 ± 0.6 | 2.7 ± 1.0 | 2.6 ± 0.9 | 0.93 |
| MDA, nmol/mL | 7.0 ± 1.1 a | 3.8 ± 1.8 b | 4.5 ± 1.5 b | 0.01 |
| PC, nmol/mL | 0.5 ± 0.2 | 0.5 ± 0.2 | 0.4 ± 0.2 | 0.80 |
| Control | OA1 | OA2 | p-Value | |
|---|---|---|---|---|
| Day 14 | ||||
| α-amylase, U/g | 69.2 ± 27.4 | 88.7 ± 22.2 | 81.4 ± 26.7 | 0.47 |
| Lipase, U/g | 57.9 ± 23.8 | 66.7 ± 23.0 | 43.7 ± 22.8 | 0.26 |
| Neutral protease, U/g | 61.8 ± 16.7 | 73.5 ± 13.8 | 62.0 ± 18.2 | 0.39 |
| Day 49 | ||||
| α-amylase, U/g | 27.0 ± 12.4 | 22.7 ± 8.1 | 25.5 ± 15.0 | 0.82 |
| Lipase, U/g | 46.3 ± 19.6 | 31.7 ± 12.0 | 30.8 ± 5.8 | 0.15 |
| Neutral protease, U/g | 46.2 ± 15.0 | 48.8 ± 15.3 | 40.3 ± 18.0 | 0.87 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiang, X.-D.; Deng, Z.-C.; Wang, Y.-W.; Sun, H.; Wang, L.; Han, Y.-M.; Wu, Y.-Y.; Liu, J.-G.; Sun, L.-H. Organic Acids Improve Growth Performance with Potential Regulation of Redox Homeostasis, Immunity, and Microflora in Intestines of Weaned Piglets. Antioxidants 2021, 10, 1665. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10111665
Xiang X-D, Deng Z-C, Wang Y-W, Sun H, Wang L, Han Y-M, Wu Y-Y, Liu J-G, Sun L-H. Organic Acids Improve Growth Performance with Potential Regulation of Redox Homeostasis, Immunity, and Microflora in Intestines of Weaned Piglets. Antioxidants. 2021; 10(11):1665. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10111665
Chicago/Turabian StyleXiang, Xin-Dong, Zhang-Chao Deng, You-Wei Wang, Hua Sun, Li Wang, Yan-Ming Han, Yuan-Yuan Wu, Jian-Gao Liu, and Lv-Hui Sun. 2021. "Organic Acids Improve Growth Performance with Potential Regulation of Redox Homeostasis, Immunity, and Microflora in Intestines of Weaned Piglets" Antioxidants 10, no. 11: 1665. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10111665