
Impact of Germination Time on Resveratrol, Phenolic Acids, and Antioxidant Capacities of Different Varieties of Peanut (Arachis hypogaea Linn.) from China




Abstract
:1. Introduction
2. Materials and Methods
2.1. Peanut Samples
2.2. Chemical Reagents
2.3. Germination and Sample Preparation
2.4. Determination of Moisture Content
2.5. Determination of Total Phenolic Content (TPC)
2.6. Determination of Total Flavonoid Content (TFC)
2.7. Determination of Monomeric Anthocyanin Content (MAC)
2.8. Determination of DPPH Free Radical Scavenging Activity (DPPH)
2.9. Determination of ABTS Free Radical Scavenging Activity (ABTS)
2.10. Determination of Ferric Reducing Antioxidant Capacity (FRAP)
2.11. Determination of Vitamin C Content
2.12. Determination of Trans-Resveratrol
2.13. Determination of Phenolic Acid
2.14. Statistical Analysis
3. Results
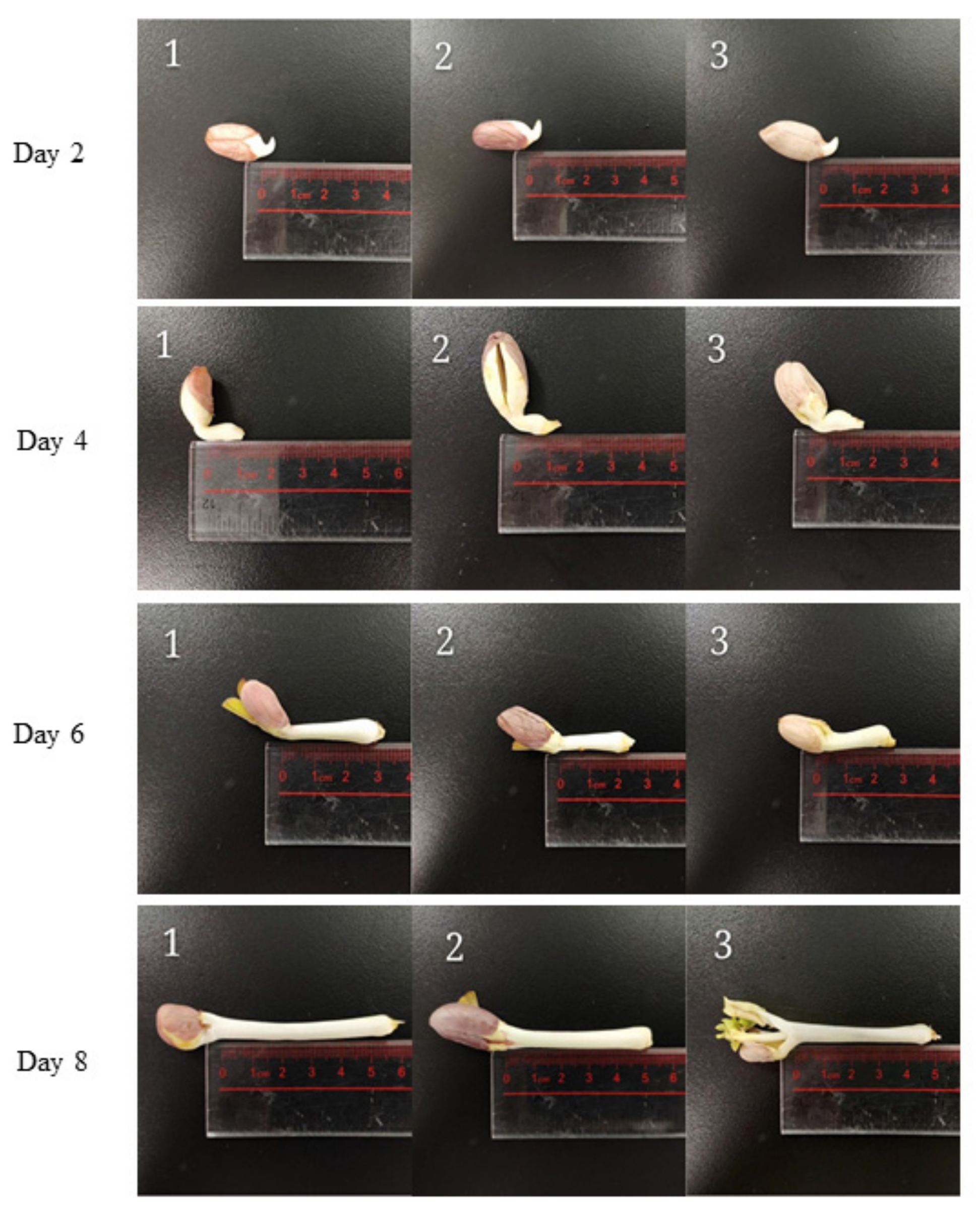
3.1. Radicle Length of Different Varieties of Peanuts during Germination
3.2. Moisture Content of Different Varieties of Peanuts during Germination
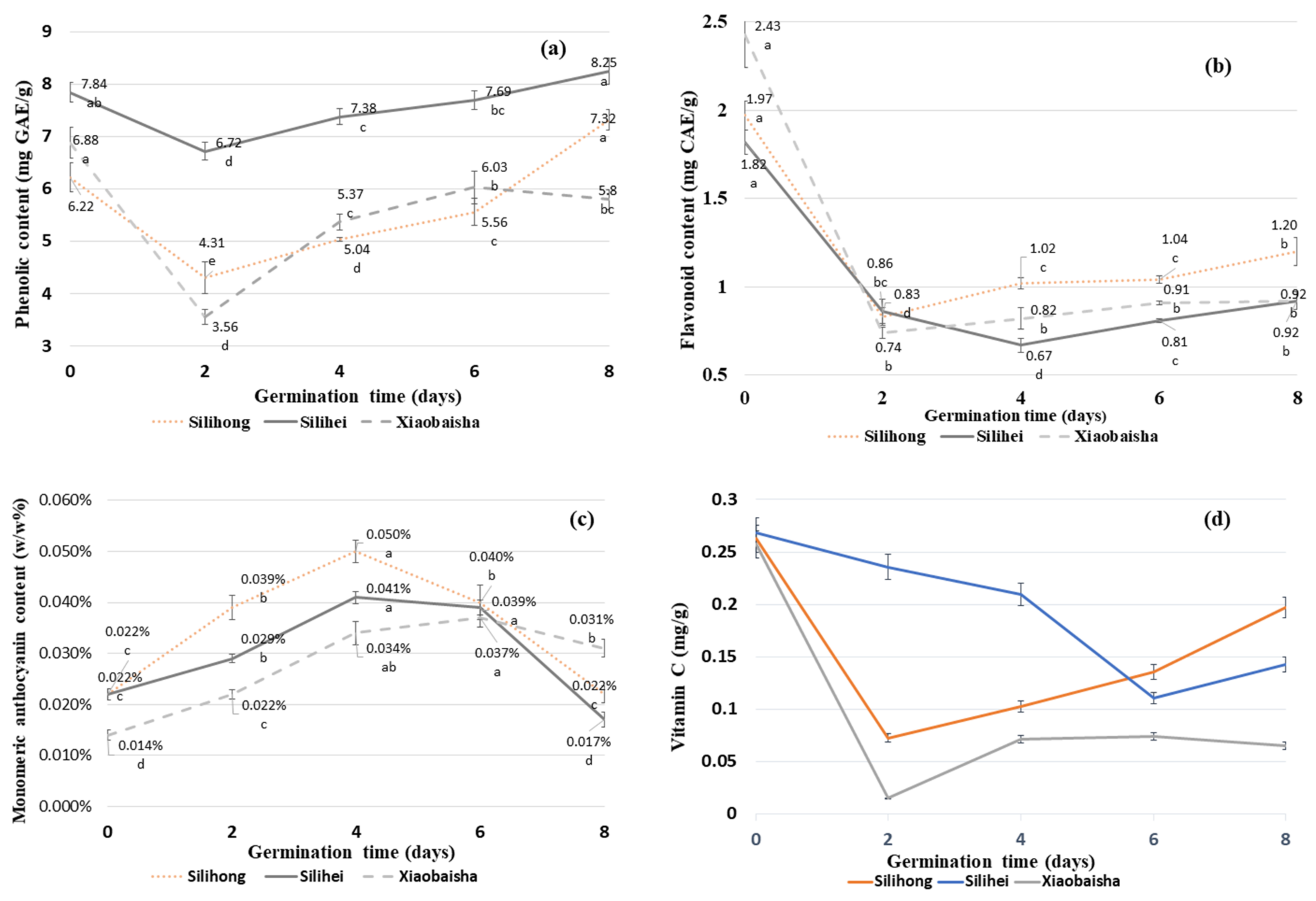
3.3. Total Phenolic Content (TPC) of Different Varieties of Peanuts during Germination
3.4. Total Flavonoid Content (TFC) of Different Varieties of Peanuts during Germination
3.5. Monomeric Anthocyanin Content (MAC) of Different Varieties of Peanuts during Germination
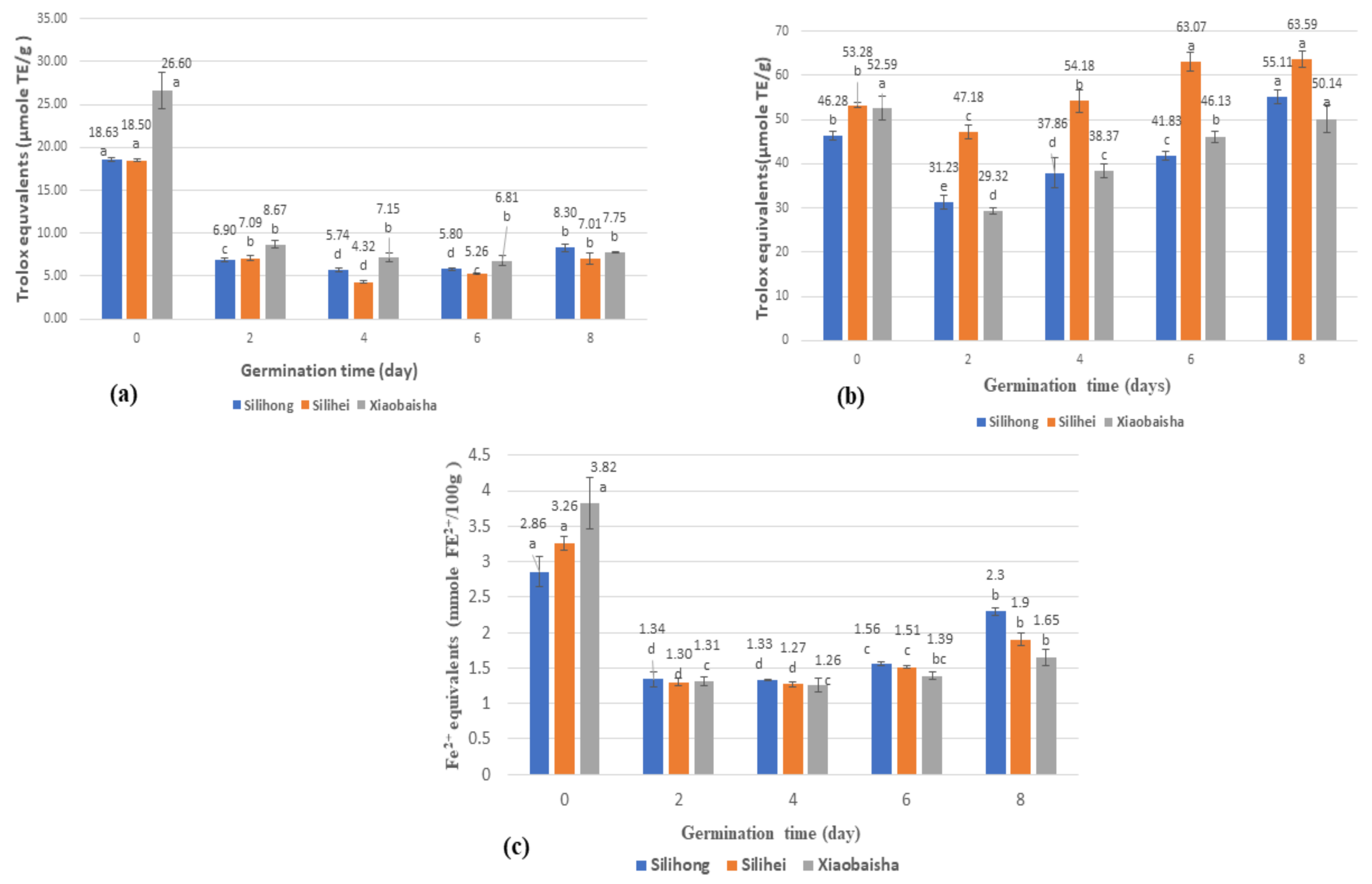
3.6. DPPH Free Radical Scavenging Activity (DPPH) of Different Varieties of Peanuts during Germination
3.7. ABTS Free Radical Scavenging Activity (ABTS) of Different Varieties of Peanuts during Germination
3.8. Ferric Reducing Antioxidant Capacity (FRAP) of Different Varieties of Peanuts during Germination
3.9. Vitamin C Content of Different Varieties of Peanuts during Germination
3.10. Trans-Resveratrol of Different Varieties of Peanuts during Germination
3.11. Phenolic acid Profile of Different Varieties of Peanuts during Germination
4. Discussion
4.1. Effects of Germination Time on Radicles Length of Different Varieties of Peanuts
4.2. Effects of Germination on the Moisture Content of Different Varieties of Peanuts
4.3. Effects of Germination on Total Phenolic Content (TPC) of Different Varieties of Peanuts
4.4. Effects of Germination on Total Flavonoid Content (TFC) of Different Varieties of Peanuts
4.5. Effects of Germination on Monomeric Anthocyanin Content (MAC) of Different Varieties of Peanuts
4.6. Effects of Germination on Antioxidant Capacities of Different Varieties of Peanuts
4.7. Correlation Analyses between Phenolic Content and Antioxidant Activities of Peanut during Germination
4.8. Effects of Germination on Vitamin C Content of Different Varieties of Peanuts
4.9. Effects of Germination Time on Trans-Resveratrol Content of Different Varieties of Peanuts
4.10. Effects of Germination on Phenolic Acid Profile of Different Varieties of Peanuts
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Abbreviation | Full Form |
| TPC | Total phenolic content |
| TFC | Total flavonoid content |
| MAC | Monomeric anthocyanin content |
| DPPH | 2,2-diphenyl-1-picrylhydrazyl |
| FRAP | Ferric Reducing Antioxidant Power |
| SK-N-SH | Human Neuroblastoma Cell Line (ATCC HTB-11) |
| mmol/L | Millimoles per Litre |
| μg | Microgram |
| ng | Nanogram |
| DW | Dry Weight basis |
| ABTS | 2,2′-azino-bis (3-ethylbenzothiazoline-6-sulfonic acid) |
| TPTZ | 2,4,6-tri(2-pyridyl)-s-triazine |
| TFA | Trifluoroacetic acid |
| nm | Nanometer |
| μL | Microliter |
| GAE | Gallic acid equivalents |
| CAE | Catechin equivalents |
| TE | Trolex equivalents |
| FE2+ | Fe2+ equivalent |
| HPLC | High-Performance Liquid Chromatography |
| RP | Reverse Phase |
| BHT | Butylated hydroxytoluene |
| PVDF | Polyvinylidene difluoride |
| LC | Liquid Chromatography |
| UV | Ultra Violet |
| IBM | International Business Machines Corporation |
| SPSS | Statistical Package for the Social Sciences |
References
- Limmongkon, A.; Janhom, P.; Amthong, A.; Kawpanuk, M.; Nopprang, P.; Poohadsuan, J.; Somboon, T.; Saijeen, S.; Surangkul, D.; Srikummool, M.; et al. Antioxidant activity, total phenolic, and resveratrol content in five cultivars of peanut sprouts. Asian Pac. J. Trop. Biomed. 2017, 7, 332–338. [Google Scholar] [CrossRef]
- Guo, C.; Xie, Y.J.; Zhu, M.T.; Xiong, Q.; Chen, Y.; Yu, Q.; Xie, J.H. Influence of different cooking methods on the nutritional and potentially harmful components of peanuts. Food Chem. 2020, 316, 126269. [Google Scholar] [CrossRef] [PubMed]
- FAO Food and Agriculture Organization of the United Nations (FAO). Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 25 March 2021).
- Attree, R.; Du, B.; Xu, B. Distribution of phenolic compounds in seed coat and cotyledon, and their contribution to antioxidant capacities of red and black seed coat peanuts (Arachis hypogaea L.). Ind. Crops Prod. 2015, 67, 448–456. [Google Scholar] [CrossRef]
- Bonku, R.; Yu, J. Health aspects of peanuts as an outcome of its chemical composition. Food Sci. Hum. Wellness 2020, 9, 21–30. [Google Scholar] [CrossRef]
- Meenu, M.; Sharma, A.; Guha, P.; Mishra, S. A rapid high-performance liquid chromatography photodiode array detection method to determine phenolic compounds in mung bean (Vigna radiata L.). Int. J. Food Prop. 2016, 19, 2223–2237. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Meenu, M.; Yu, H.; Xu, B. An investigation on phenolic and antioxidant capacity of under-utilized food legumes consumed in China. Foods 2020, 9, 438. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.C.; Qian, H.; Sun, X.L.; Cui, Y.; Wang, H.Y.; Du, C.; Xia, X.H. The effects of germination on chemical composition of peanut seed. Food Sci. Technol. Res. 2014, 20, 883–889. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.D.; Wen, Y.Y.; Chiou, J.M.; Wang, K.H.; Chiou, R.Y.Y. Comparative characterization of peanuts grown by aquatic floating cultivation and field cultivation for seed and resveratrol production. J. Agric. Food Chem. 2003, 51, 1582–1585. [Google Scholar] [CrossRef]
- Wang, C.; Wan, X.; Xie, Y.; Lu, X.; Deng, Y.; Zheng, Y. Effects of different methods on the yield and growth of peanut sprouts. Guangdong Agric. Sci. 2018, 6, 7–13. [Google Scholar]
- Xie, M.; Chen, H.; Zhang, G.; Lin, Y.; Su, W. Effects of temperature, soil moisture and sowing depth on the seed germination and seedling growth of peanut. J. Peanut Sci. 2017, 46, 52–59. [Google Scholar]
- Wang, K.H.; Lai, Y.H.; Chang, J.C.; Ko, T.F.; Shyu, S.L.; Chiou, R.Y.Y. Germination of peanut kernels to enhance resveratrol biosynthesis and prepare sprouts as a functional vegetable. J. Agric. Food Chem. 2005, 53, 242–246. [Google Scholar] [CrossRef]
- Ha, A.W.; Kim, W.K.; Kim, J.H.; Kang, N.E. The supplementation effects of peanut sprout on reduction of abdominal fat and health indices in overweight and obese women. Nutr. Res. Pract. 2015, 9, 249–255. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Meenu, M.; Xu, B.; Yu, H. Impact of processing technologies on isoflavones, phenolic acids, and antioxidant capacities of soymilk prepared from 15 soybean varieties. Food Chem. 2021, 345, 128612. [Google Scholar] [CrossRef]
- Xu, Z.; Meenu, M.; Xu, B. Effects of UV-C treatment and ultrafine-grinding on the biotransformation of ergosterol to vitamin D2, physiochemical properties, and antioxidant properties of shiitake and Jew’s ear. Food Chem. 2020, 309, 125738. [Google Scholar] [CrossRef]
- Xu, Z.; Meenu, M.; Chen, P.; Xu, B. Comparative study on phytochemical profiles and antioxidant capacities of chestnuts produced in different geographic area in China. Antioxidants 2020, 9, 190. [Google Scholar] [CrossRef] [Green Version]
- Limmongkon, A.; Nopprang, P.; Chaikeandee, P.; Somboon, T.; Wongshaya, P.; Pilaisangsuree, V. LC-MS/MS profiles and interrelationships between the anti-inflammatory activity, total phenolic content and antioxidant potential of Kalasin 2 cultivar peanut sprout crude extract. Food Chem. 2018, 239, 569–578. [Google Scholar] [CrossRef]
- Bo, S.; Ciccone, G.; Castiglione, A.; Gambino, R.; De Michieli, F.; Villois, P.; Durazzo, M.; Cavallo-Perin, P.; Cassader, M. Anti-inflammatory and antioxidant effects of resveratrol in healthy smokers a randomized, double-blind, placebo-controlled, cross-over trial. Curr. Med. Chem. 2013, 20, 1323–1331. [Google Scholar] [CrossRef]
- Saleh, M.C.; Connell, B.J.; Saleh, T.M. Resveratrol induced neuroprotection is mediated via both estrogen receptor subtypes, ERα and ERβ. Neurosci. Lett. 2013, 548, 217–221. [Google Scholar] [CrossRef]
- Lertkaeo, P.; Limmongkon, A.; Srikummool, M.; Boonsong, T.; Supanpaiboon, W.; Surangkul, D. Antioxidative and neuroprotective activities of peanut sprout extracts against oxidative stress in SK-N-SH cells. Asian Pac. J. Trop. Biomed. 2017, 7, 64–69. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.J.; Luo, Q.; Lei, Q.; Xia, W.; Zhang, Y.; Fu, X. Study on the changes of nutritional and functional components during peanut germination. Hubei Agric. Sci. 2018, 57, 89–96. [Google Scholar]
- Xu, B.; Chang, S.K.C. Characterization of phenolic substances and antioxidant properties of food soybeans grown in the North Dakota-Minnesota region. J. Agric. Food Chem. 2008, 56, 9102–9113. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.J.; Chang, S.K.C. A comparative study on phenolic profiles and antioxidant activities of legumes as affected by extraction solvents. J. Food Sci. 2007, 72, S159–S166. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Durst, R.W.; Wrolstad, R.E.; Eisele, T.; Giusti, M.M.; Hach, J.; Hofsommer, H.; Koswig, S.; Krueger, D.A.; Kupina, S.; et al. Determination of Total Monomeric Anthocyanin Pigment Content of Fruit Juices, Beverages, Natural Colorants, and Wines by the pH Differential Method: Collaborative Study. J. AOAC Int. 2005, 88, 1269–1278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, N.J.; Rice-Evans, C.; Davies, M.J.; Gopinathan, V.; Milner, A. A novel method for measuring antioxidant capacity and its application to monitoring the antioxidant status in premature neonates. Clin. Sci. 1993, 84, 407–412. [Google Scholar] [CrossRef] [Green Version]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Xue, Z.; Wang, C.; Zhai, L.; Yu, W.; Chang, H.; Kou, X.; Zhou, F. Bioactive compounds and antioxidant activity of mung bean (Vigna radiata L.), soybean (Glycine max L.) and black bean (Phaseolus vulgaris L.) during the germination process. Czech J. Food Sci. 2016, 34, 68–78. [Google Scholar] [CrossRef]
- Xu, S.; Luo, H.; Chen, H.; Guo, J.; Yu, B.; Zhang, H.; Li, W.; Chen, W.; Zhou, X.; Huang, L.; et al. Optimization of extraction of total trans-resveratrol from peanut seeds and its determination by HPLC. J. Sep. Sci. 2020, 43, 1024–1031. [Google Scholar] [CrossRef]
- Zhou, X.; Zhang, Q. The influence of different temperatures on the growth of soybean sprouts. Gansu Sci. Technol. 2012, 28, 142–143. [Google Scholar]
- Yu, L.; Zhang, C.; Bi, J.; Wang, M.; Lin, R.; Yang, Z. Research of peanut sprout production by different varieties of peanut and its nutrition analysis. Food Ind. Technol. 2017, 38, 314–319. [Google Scholar]
- Wang, X.; Wang, C. Changes of physiological indicators, components and their correlation analysis in peanut during germination. Food Sci. Technol. 2018, 319, 182–187. [Google Scholar]
- Bove, J.; Jullien, M.; Grappin, P. Functional genomics in the study of seed germination. Genome Biol. 2002, 3, 1–5. [Google Scholar]
- Ji, H.; Ren, Y.; Zhang, M.; Zhao, K.; Shi, Y.; Xie, W. Nutrient and metabolism of peanut sprouts during the growth. J. Beijing Agric. Coll. 2013, 28, 17–19. [Google Scholar]
- Rao, P.U.; Deosthale, Y.G. Polyphenoloxidase activity in germinated legume seeds. J. Food Sci. 1987, 52, 1549–1551. [Google Scholar] [CrossRef]
- Win, M.M.; Abdul-Hamid, A.; Baharin, B.S.; Anwar, F.; Sabu, M.C.; Pak-Dek, M.S. Phenolic compounds and antioxidant activity of peanut’s skin, hull, raw kernel and roasted kernel flour. Pak. J. Bot 2011, 43, 1635–1642. [Google Scholar]
- Saleh, H.M.; Hassan, A.A.; Mansour, E.H.; Fahmy, H.A.; El-Bedawey, A.E.F.A. Melatonin, phenolics content and antioxidant activity of germinated selected legumes and their fractions. J. Saudi Soc. Agric. Sci. 2019, 18, 294–301. [Google Scholar] [CrossRef]
- Heimler, D.; Vignolini, P.; Dini, M.G.; Romani, A. Rapid tests to assess the antioxidant activity of Phaseolus vulgaris L. dry beans. J. Agric. Food Chem. 2005, 53, 3053–3056. [Google Scholar] [CrossRef]
- Yu, J.; Ahmedna, M.; Goktepe, I. Effects of processing methods and extraction solvents on concentration and antioxidant activity of peanut skin phenolics. Food Chem. 2005, 90, 199–206. [Google Scholar] [CrossRef]
- Yodpitak, S.; Mahatheeranont, S.; Boonyawan, D.; Sookwong, P.; Roytrakul, S.; Norkaew, O. Cold plasma treatment to improve germination and enhance the bioactive phytochemical content of germinated brown rice. Food Chem. 2019, 289, 328–339. [Google Scholar] [CrossRef]
- James, S.; Nwabueze, T.U.; Ndife, J.; Onwuka, G.I.; Usman, M.A.A. Influence of fermentation and germination on some bioactive components of selected lesser legumes indigenous to Nigeria. J. Agric. Food Res. 2020, 2, 100086. [Google Scholar] [CrossRef]
- Türker, N.; Erdoğdu, F. Effects of pH and temperature of extraction medium on effective diffusion coefficient of anthocynanin pigments of black carrot (Daucus carota var. L.). J. Food Eng. 2006, 76, 579–583. [Google Scholar] [CrossRef]
- Lin, P.Y.; Lai, H.M. Bioactive compounds in legumes and their germinated products. J. Agric. Food Chem. 2006, 54, 3807–3814. [Google Scholar] [CrossRef] [PubMed]
- Thaipong, K.; Boonprakob, U.; Crosby, K.; Cisneros-Zevallos, L.; Byrne, D.H. Comparison of ABTS, DPPH, FRAP, and ORAC assays for estimating antioxidant activity from guava fruit extracts. J. Food Compos. Anal. 2006, 19, 669–675. [Google Scholar] [CrossRef]
- Zhang, Z.; Wan, S.; Dai, L.; Chen, J.; Miao, H. Study on antioxidant capacity and functional components of seed kernels of different types of peanut varieties. J. Food Biotechnol. 2009, 28, 741–747. [Google Scholar]
- Guo, X.; Li, T.; Tang, K.; Liu, R.H. Effect of germination on phytochemical profiles and antioxidant activity of mung bean sprouts (Vigna radiata). J. Agric. Food Chem. 2012, 60, 11050–11055. [Google Scholar] [CrossRef] [PubMed]
- Frias, J.; Miranda, M.L.; Doblado, R.; Vidal-Valverde, C. Effect of germination and fermentation on the antioxidant vitamin content and antioxidant capacity of Lupinus albus L. var. Multolupa. Food Chem. 2005, 92, 211–220. [Google Scholar] [CrossRef]
- Ingham, J.L. 3,5,4′-trihydroxystilbene as a phytoalexin from groundnuts (Arachis hypogaea). Phytochemistry 1976, 15, 1791–1793. [Google Scholar] [CrossRef]
- Sanders, T.H.; McMichael, R.W.; Hendrix, K.W. Occurrence of resveratrol in edible peanuts. J. Agric. Food Chem. 2000, 48, 1243–1246. [Google Scholar] [CrossRef]
- Huang, X.; Cai, W.; Xu, B. Kinetic changes of nutrients and antioxidant capacities of germinated soybean (glycine max l.) and mung bean (Vigna radiata L.) with germination time. Food Chem. 2014, 143, 268–276. [Google Scholar] [CrossRef]
- López-Amorós, M.L.; Hernández, T.; Estrella, I. Effect of germination on legume phenolic compounds and their antioxidant activity. J. Food Compos. Anal. 2006, 19, 277–283. [Google Scholar] [CrossRef]
- Kole, R.K.; Karmakar, P.R.; Poi, R.; Mazumdar, D.; Chandra, B.; Viswavidyalaya, K. Allelopathic inhibition of teak leaf extract: A potential pre-emergent herbicide. J. Crop Weed. 2011, 7, 101–109. [Google Scholar]
- Yan, J.; Bi, H.-H.; Liu, Y.-Z.; Zhang, M.; Zhou, Z.-Y.; Tan, J.-W. Phenolic compounds from Merremia umbellata subsp. orientalis and their allelopathic effects on Arabidopsis seed germination. Molecules 2010, 15, 8241–8250. [Google Scholar] [CrossRef] [Green Version]
- Zanardo, D.I.L.; Lima, R.B.; Ferrarese, M.D.L.L.; Bubna, G.A.; Ferrarese-Filho, O. Soybean root growth inhibition and lignification induced by p-coumaric acid. Environ. Exp. Bot. 2009, 66, 25–30. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Varieties | Germination Time (Days) | Trans-Resveratrol (ng/g) |
|---|---|---|
| 0 | 26.52 ± 3.19 c | |
| 2 | 117.36 ± 11.94 c | |
| Silihong | 4 | 468.10 ± 72.17 b |
| 6 | 638.51 ± 113.35 a | |
| 8 | 415.93 ± 27.00 b | |
| 0 | 19.91 ± 5.57 b | |
| 2 | 42.64 ± 1.57 b | |
| Silihei | 4 | 59.34 ± 5.35 b |
| 6 | 487.23 ± 49.29 a | |
| 8 | 631.14 ± 185.92 a | |
| 0 | 40.08 ± 2.44 d | |
| 2 | 52.18 ± 1.85 cd | |
| Xiaobaisha | 4 | 119.67 ± 13.82 c |
| 6 | 281.74 ± 77.82 a | |
| 8 | 194.90 ± 43.26 b |
| Varieties | Phenolic Acid (μg/g) | Germination Time (Days) | ||||
|---|---|---|---|---|---|---|
| 0 | 2 | 4 | 6 | 8 | ||
| Silihong | Gallic acid | 4.58 ± 0.38 c | 32.15 ± 2.17 ab | 25.54 ± 2.49 b | 28.99 ± 2.75 b | 38.44 ± 3.54 a |
| Protocatechuic acid | 0.93 ± 0.13 cd | 0.53 ± 0.11 d | 1.41 ± 0.22 bc | 4.23 ± 0.58 a | 2.02 ± 0.18 b | |
| Protocatechualdehyde | 3.72 ± 0.23 b | 4.50 ± 0.00 a | 1.82 ± 0.05 c | 0.99 ± 0.15 d | 4.01 ± 0.38 ab | |
| p-hydroxybenzoic acid | 1.52 ± 0.08 b | 2.48 ± 0.23 b | 0.74 ± 0.07 b | 9.06 ± 2.22 a | 9.29 ± 0.02 a | |
| Gentisic acid | 23.95 ± 1.10 a | 15.07 ± 1.40 b | 13.88 ± 1.19 bc | 10.56 ± 1.95 c | 11.01 ± 0.68 c | |
| Chlorogenic acid | 19.46 ± 1.69 c | 30.80 ± 2.41 c | 59.04 ± 3.78 c | 105.24 ± 19.93 b | 144.43 ± 8.80 a | |
| Syringic acid | 5.87 ± 0.01 a | 1.16 ± 0.21 c | 0.61 ± 0.21 c | 2.46 ± 0.68 b | 3.09 ± 0.02 b | |
| p-Coumaric acid + Syringaldehyde | 6.82 ± 0.20 d | 14.99 ± 1.51 c | 19.68 ± 2.23 b | 21.02 ± 0.14 b | 38.73 ± 2.09 a | |
| Ferulic acid | 1.36 ± 0.21 e | 3.51 ± 0.26 d | 7.95 ± 0.55 b | 6.21 ± 0.34 c | 9.47 ± 0.19 a | |
| Sinapic acid | 4.77 ± 0.47 b | 6.80 ± 0.48 a | 4.96 ± 0.26 b | 4.55 ± 0.70 b | 6.90 ± 0.12 a | |
| Silihei | Gallic acid | 5.47 ± 1.60 a | 8.13 ± 1.42 b | 8.53 ± 2.25 b | 13.95 ± 1.31 b | 30.87 ± 9.79 b |
| Protocatechuic acid | 0.66 ± 0.22 b | 0.51 ± 0.08 b | 0.40 ± 0.16 b | 3.71 ± 0.19 a | 6.36 ± 0.59 a | |
| Protocatechualdehyde | 4.05 ± 1.00 a | 3.76 ± 0.01 a | 0.76 ± 0.11 b | 0.68 ± 0.21 b | 0.71 ± 0.13 b | |
| p-hydroxybenzoic acid | 0.36 ± 0.09 d | 1.10 ± 0.08 c | 1.11 ± 0.26 c | 2.09 ± 0.17 b | 5.13 ± 0.46 a | |
| Gentisic acid | 26.11 ± 7.21 bc | 35.23 ± 2.71 ab | 20.30 ± 6.07 c | 35.99 ± 4.59 ab | 42.48 ± 6.23 a | |
| Chlorogenic acid | 10.52 ± 1.89 c | 26.61 ± 10.40 c | 19.32 ± 3.80 c | 61.45 ± 8.39 b | 111.05 ± 11.75 a | |
| Syringic acid | 2.26 ± 0.45 a | 0.74 ± 0.10 c | 0.63 ± 0.12 c | 1.14 ± 0.01 bc | 1.71 ± 0.16 ab | |
| p-Coumaric acid + Syringaldehyde | 3.84 ± 0.90 c | 16.52 ± 1.36 bc | 26.21 ± 9.02 b | 74.14 ± 8.28 a | 60.26 ± 8.17 a | |
| Ferulic acid | 1.67 ± 0.18 d | 3.45 ± 0.23 cd | 5.92 ± 2.22 c | 14.47 ± 1.56 a | 10.30 ± 1.20 b | |
| Sinapic acid | 5.55 ± 1.26 c | 10.51 ± 2.19 ab | 6.46 ± 2.42 bc | 12.07 ± 1.55 a | 9.82 ± 1.25 ab c | |
| Xiaobaisha | Gallic acid | 0.66 ± 0.08 d | 29.24 ± 0.86 b | 24.68 ± 3.15 bc | 21.14 ± 1.00 c | 39.78 ± 4.44 a |
| Protocatechuic acid | 1.45 ± 0.44 c | 0.69 ± 0.12 c | 0.61 ± 0.01 c | 5.70 ± 0.51 b | 8.20 ± 0.93 a | |
| Protocatechualdehyde | 0.91 ± 0.12 c | 4.16 ± 1.03 a | 2.99 ± 0.43 ab | 2.42 ± 0.32 bc | 1.95 ± 0.54 bc | |
| p-hydroxybenzoic acid | 2.51 ± 1.02 b | 1.26 ± 0.14 b | 0.41 ± 0.03 b | 5.63 ± 2.16 a | 8.31 ± 0.09 a | |
| Gentisic acid | 35.40 ± 9.80 a | 39.30 ± 5.03 a | 52.96 ± 5.11 a | 40.52 ± 4.68 a | 39.87 ± 9.24 a | |
| Chlorogenic acid | 19.63 ± 4.97 c | 30.38 ± 8.18 c | 84.05 ± 6.33 b | 106.60 ± 11.84 b | 186.65 ± 38.04 a | |
| Syringic acid | 5.31 ± 0.87 a | 1.46 ± 0.39 b | 0.13 ± 0.03 c | 0.83 ± 0.15 bc | 1.90 ± 0.26 b | |
| p-Coumaric acid + Syringaldehyde | 15.00 ± 0.45 c | 9.07 ± 0.94 c | 14.28 ± 0.49 c | 31.18 ± 3.71 b | 53.36 ± 8.63 a | |
| Ferulic acid | 1.16 ± 0.17 c | 3.07 ± 0.27 b | 5.04 ± 0.28 a | 6.15 ± 0.77 a | 6.07 ± 0.59 a | |
| Sinapic acid | 15.46 ± 1.67 b | 18.50 ± 5.16 b | 38.92 ± 5.71 a | 28.35 ± 2.15 ab | 32.58 ± 8.35 a | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Z.; Fan, Z.; Meenu, M.; Xu, B. Impact of Germination Time on Resveratrol, Phenolic Acids, and Antioxidant Capacities of Different Varieties of Peanut (Arachis hypogaea Linn.) from China. Antioxidants 2021, 10, 1714. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10111714
Zhou Z, Fan Z, Meenu M, Xu B. Impact of Germination Time on Resveratrol, Phenolic Acids, and Antioxidant Capacities of Different Varieties of Peanut (Arachis hypogaea Linn.) from China. Antioxidants. 2021; 10(11):1714. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10111714
Chicago/Turabian StyleZhou, Ziying, Zhili Fan, Maninder Meenu, and Baojun Xu. 2021. "Impact of Germination Time on Resveratrol, Phenolic Acids, and Antioxidant Capacities of Different Varieties of Peanut (Arachis hypogaea Linn.) from China" Antioxidants 10, no. 11: 1714. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10111714