
Fresh Phyllanthus emblica (Amla) Fruit Supplementation Enhances Milk Fatty Acid Profiles and the Antioxidant Capacities of Milk and Blood in Dairy Cows

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fresh Amla Fruit Extraction, Characterization, and Quantification of Metabolites
2.1.1. Amla Fruit Sample Preparation and Extraction
2.1.2. ESI-Q TRAP-MS/MS
2.1.3. Characterization and Quantification of Metabolites
2.2. Animals, Diets, and Experimental Design
2.3. Blood Sampling, Measurement, and Analysis
2.4. Milk Fatty Acid Composition
2.5. Determination of Total Flavonoids
2.6. Statistical Analysis
3. Results
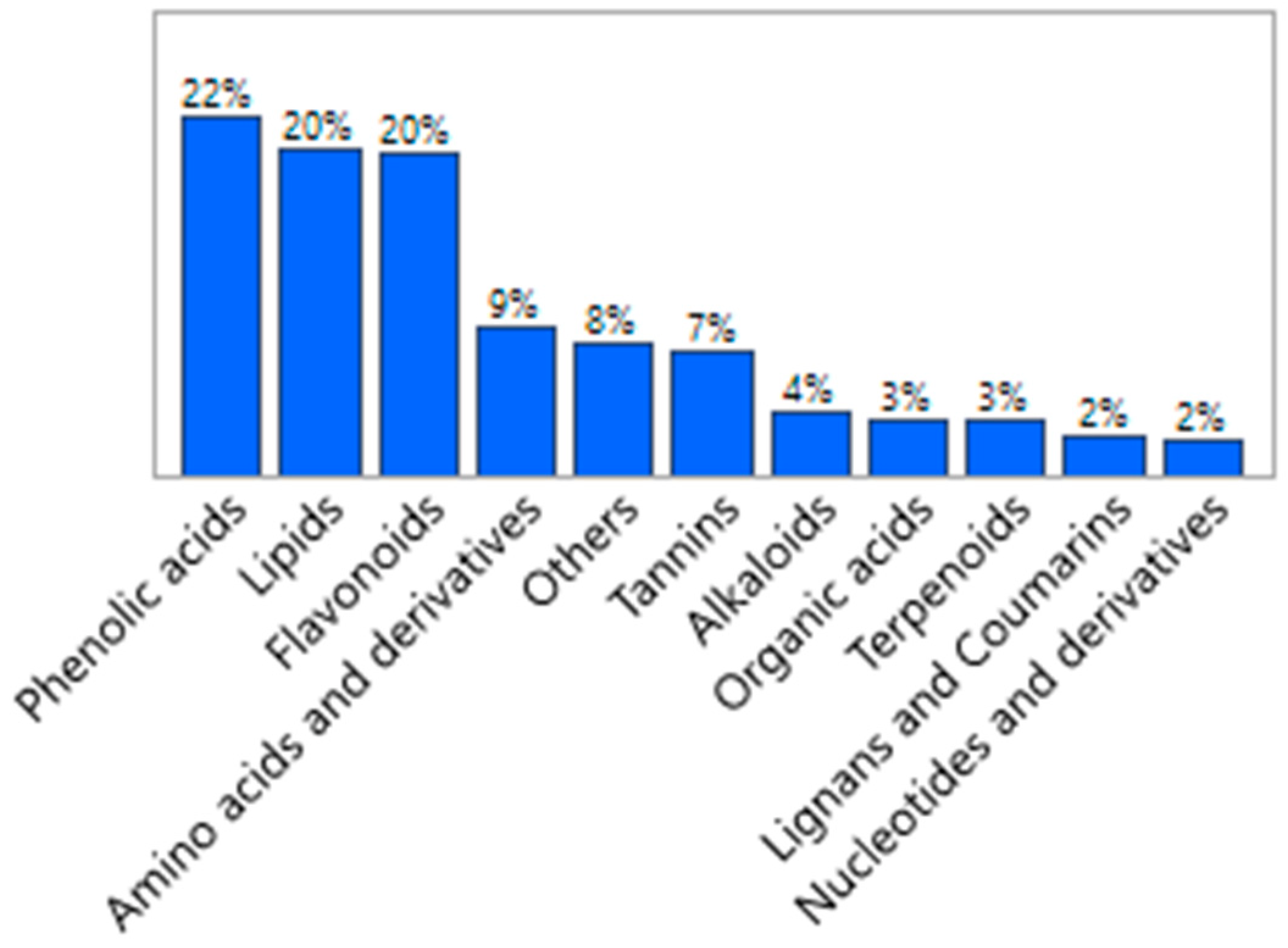
3.1. Characteristics of Fresh Amla Fruit
3.2. Milk Fatty Acid Profiles
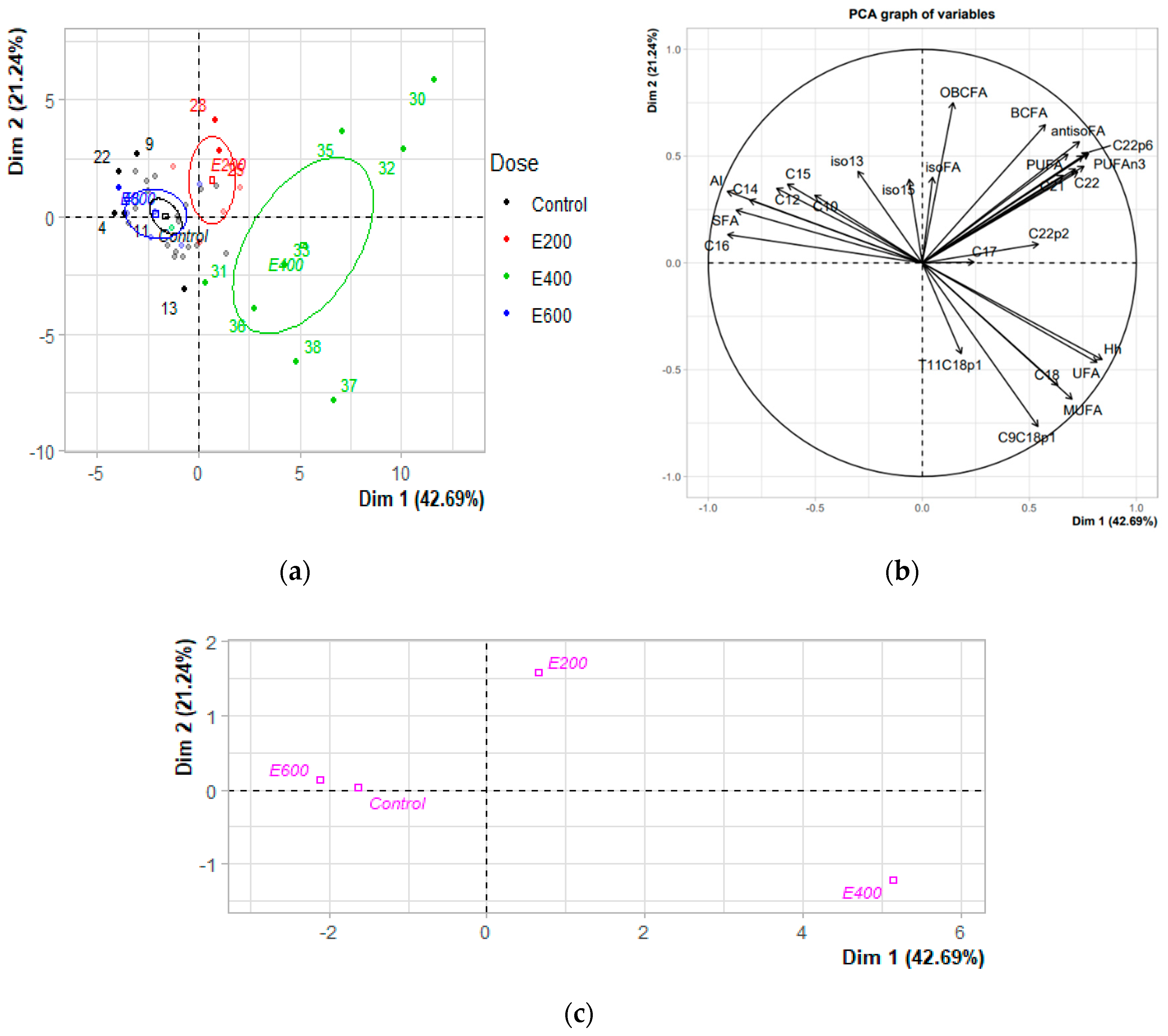
3.3. Exploring Milk Fatty Acid Data with PCA
3.4. Blood Biochemical Parameters
3.5. Antioxidant Capacity in Milk and Plasma
4. Discussion
4.1. Fresh Amla Fruit Characteristics
4.2. Milk Fatty Acid Profile
4.3. Blood Biochemical Parameters
4.4. Liver and Antioxidant Enzymes
4.5. Antioxidant Capacity and Status in the Milk
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, W.-C.; Chen, C.-Y.; Wu, S.-J. Almond skin polyphenol extract inhibits inflammation and promotes lipolysis in differentiated 3T3-L1 adipocytes. J. Med. Food 2017, 20, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Drong, C.; Meyer, U.; Von Soosten, D.; Frahm, J.; Rehage, J.; Breves, G.; Dänicke, S. Effect of monensin and essential oils on performance and energy metabolism of transition dairy cows. J. Anim. Physiol. Anim. Nutr. 2016, 100, 537–551. [Google Scholar] [CrossRef] [PubMed]
- Rochfort, S.; Parker, A.J.; Dunshea, F.R. Plant bioactives for ruminant health and productivity. Phytochemistry 2008, 69, 299–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khiaosa-Ard, R.; Bryner, S.F.; Scheeder, M.R.; Wettstein, H.R.; Leiber, F.; Kreuzer, M.; Soliva, C.R. Evidence for the inhibition of the terminal step of ruminal alpha-linolenic acid biohydrogenation by condensed tannins. J. Dairy Sci. 2009, 92, 177–188. [Google Scholar] [CrossRef] [Green Version]
- Vasta, V.; Makkar, H.P.; Mele, M.; Priolo, A. Ruminal biohydrogenation as affected by tannins in vitro. Br. J. Nutr. 2009, 102, 82–92. [Google Scholar] [CrossRef] [Green Version]
- Buccioni, A.; Minieri, S.; Rapaccini, S.; Antongiovanni, M.; Mele, M. Effect of chestnut and quebracho tannins on fatty acid profile in rumen liquid-and solid-associated bacteria: An in vitro study. Animal 2011, 5, 1521–1530. [Google Scholar] [CrossRef]
- Hassan, F.-U.; Ebeid, H.M.; Tang, Z.; Li, M.; Peng, L.; Peng, K.; Liang, X.; Yang, C. A Mixed Phytogenic Modulates the Rumen Bacteria Composition and Milk Fatty Acid Profile of Water Buffaloes. Front. Vet. Sci. 2020, 7. [Google Scholar] [CrossRef]
- Cosentino, C.; Colonna, M.; Musto, M.; Dimotta, A.; Freschi, P.; Tarricone, S.; Ragni, M.; Paolino, R. Effects of dietary supplementation with extruded linseed and oregano in autochthonous goat breeds on the fatty acid profile of milk and quality of Padraccio cheese. J. Dairy Sci. 2021, 104, 1445–1453. [Google Scholar] [CrossRef]
- Cabiddu, A.; Molle, G.; Decandia, M.; Spada, S.; Fiori, M.; Piredda, G.; Addis, M. Responses to condensed tannins of flowering sulla (Hedysarum coronarium L.) grazed by dairy sheep. Livest. Sci. 2009, 123, 230–240. [Google Scholar] [CrossRef]
- Abdoul-Aziz, S.K.A.; Zhang, Y.; Wang, J. Milk Odd and Branched Chain Fatty Acids in Dairy Cows: A Review on Dietary Factors and Its Consequences on Human Health. Animals 2021, 11, 3210. [Google Scholar] [CrossRef]
- Rafiee-Yarandi, H.; Ghorbani, G.R.; Alikhani, M.; Sadeghi-Sefidmazgi, A.; Drackley, J.K. A comparison of the effect of soybeans roasted at different temperatures versus calcium salts of fatty acids on performance and milk fatty acid composition of mid-lactation Holstein cows. J. Dairy Sci. 2016, 99, 5422–5435. [Google Scholar] [CrossRef] [Green Version]
- Addis, M.; Cabiddu, A.; Pinna, G.; Decandia, M.; Piredda, G.; Pirisi, A.; Molle, G. Milk and cheese fatty acid composition in sheep fed Mediterranean forages with reference to conjugated linoleic acid cis-9, trans-11. J. Dairy Sci. 2005, 88, 3443–3454. [Google Scholar] [CrossRef] [Green Version]
- Santos, N.W.; Yoshimura, E.H.; Machado, E.; Matumoto-Pintro, P.T.; Montanher, P.F.; Visentainer, J.V.; dos Santos, G.T.; Zeoula, L.M. Antioxidant effects of a propolis extract and vitamin E in blood and milk of dairy cows fed diet containing flaxseed oil. Livest. Sci. 2016, 191, 132–138. [Google Scholar] [CrossRef]
- Todaro, M.; Alabiso, M.; Scatassa, M.; Di Grigoli, A.; Mazza, F.; Maniaci, G.; Bonanno, A. Effect of the inclusion of fresh lemon pulp in the diet of lactating ewes on the properties of milk and cheese. Anim. Feed Sci. Technol. 2017, 225, 213–223. [Google Scholar] [CrossRef] [Green Version]
- Di Trana, A.; Bonanno, A.; Cecchini, S.; Giorgio, D.; Di Grigoli, A.; Claps, S. Effects of Sulla forage (Sulla coronarium L.) on the oxidative status and milk polyphenol content in goats. J. Dairy Sci. 2015, 98, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Huxley, R.R.; Neil, H. The relation between dietary flavonol intake and coronary heart disease mortality: A meta-analysis of prospective cohort studies. Eur. J. Clin. Nutr. 2003, 57, 904–908. [Google Scholar] [CrossRef] [Green Version]
- Whitley, A.C.; Stoner, G.D.; Darby, M.V.; Walle, T. Intestinal epithelial cell accumulation of the cancer preventive polyphenol ellagic acid—Extensive binding to protein and DNA. Biochem. Pharmacol. 2003, 66, 907–915. [Google Scholar] [CrossRef]
- Shingfield, K.J.; Bonnet, M.; Scollan, N.D. Recent developments in altering the fatty acid composition of ruminant-derived foods. Animal 2013, 7 (Suppl. S1), 132–162. [Google Scholar] [CrossRef]
- Gantait, S.; Mahanta, M.; Bera, S.; Verma, S.K. Advances in biotechnology of Emblica officinalis Gaertn. syn. Phyllanthus emblica L.: A nutraceuticals-rich fruit tree with multifaceted ethnomedicinal uses. 3 Biotech 2021, 11, 62. [Google Scholar] [CrossRef]
- Thilaga, S.; Largia, M.J.V.; Parameswari, A.; Nair, R.R.; Ganesh, D. High frequency somatic embryogenesis from leaf tissue of ‘Emblica officinalis’ Gaertn.—A high valued tree for non-timber forest products. Aust. J. Crop Sci. 2013, 7, 1480–1487. [Google Scholar]
- Wadhwa, M.; Bakshi, M.P.; Makkar, H.P. Waste to worth: Fruit wastes and by-products as animal feed. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2015, 10, 1–26. [Google Scholar] [CrossRef]
- Kunchana, K.; Jarisarapurin, W.; Chularojmontri, L.; Wattanapitayakul, S.K. Potential Use of Amla (Phyllanthus emblica L.) Fruit Extract to Protect Skin Keratinocytes from Inflammation and Apoptosis after UVB Irradiation. Antioxidants 2021, 10, 703. [Google Scholar] [CrossRef]
- Jacob, A.; Pandey, M.; Kapoor, S.; Saroja, R. Effect of the Indian gooseberry (amla) on serum cholesterol levels in men aged 35–55 years. Eur. J. Clin. Nutr. 1988, 42, 939–944. [Google Scholar]
- Wu, S.; Zhou, L. An edible value research of Phyllenthus emblica L. Acad J. Kunming Med. Coll. 1996, 17, 22–23. [Google Scholar]
- Yang, B.; Kortesniemi, M.; Liu, P.; Karonen, M.; Salminen, J.P. Analysis of hydrolyzable tannins and other phenolic compounds in emblic leafflower (Phyllanthus emblica L.) fruits by high performance liquid chromatography-electrospray ionization mass spectrometry. J. Agric. Food Chem. 2012, 60, 8672–8683. [Google Scholar] [CrossRef]
- Yang, B.; Liu, P. Composition and biological activities of hydrolyzable tannins of fruits of Phyllanthus emblica. J. Agric. Food Chem. 2014, 62, 529–541. [Google Scholar] [CrossRef] [PubMed]
- Ghosal, S. Active constituents of Emblica officinalis: Part I. The chemistry and antioxidative effects of two new hydrolysable tannins, Emblicanin A and B. Indian J. Chem. 1996, 35, 941–948. [Google Scholar]
- Tilahun, M.; Zhao, L.; Guo, Z.; Shen, Y.; Ma, L.; Callaway, T.R.; Xu, J.; Bu, D. Amla (Phyllanthus emblica) fresh fruit as new feed source to enhance ruminal fermentation and milk production in lactating dairy cows. Anim. Feed Sci. Technol. 2022, 283, 115160. [Google Scholar] [CrossRef]
- Lakhani, N.; Kamra, D.N.; Lakhani, P.; Alhussien, M.N. Immune status and haemato-biochemical profile of buffalo calves supplemented with phytogenic feed additives rich in tannins, saponins and essential oils. Trop. Anim. Health Prod. 2019, 51, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Lakhani, N.; Kamra, D.; Kala, A.; Agarwal, N.; Chaudhary, L.; Chaturvedi, V. Effects of dietary supplementation with rumen modifier and sodium sulphate on methane production and performance of buffalo calves. Anim. Nutr. Feed Technol. 2019, 19, 169–180. [Google Scholar] [CrossRef]
- Lozano-Sánchez, M.; Mendoza-Martínez, G.D.; Martínez-García, J.A.; la Torre-Hernández, M.E.D.; Chamorro-Ramírez, F.H.; Martínez-Aispuro, J.A.; Cordero-Mora, J.L.; Sánchez-Torres, M.T.; Hernández-García, P.A.; Jones, R. Evaluation of polyherbal with vitamin C activity on lamb performance and meat characteristics. Rev. Bras. Zootec. 2021, 50. [Google Scholar] [CrossRef]
- Sharma, A.; Prasad, S.; Singh, Y.; Bishisth, R. Effect of polyherbal preparation supplementation on immunity and udder health of periparturient Karan-Fries crossbred dairy cows. J. Appl. Anim. Res. 2014, 42, 217–221. [Google Scholar] [CrossRef] [Green Version]
- Rizwan, M.; Durrani, A.Z.; Ahmad, T.; Ahmad, S.S.; Chaudhry, M. Comparative therapeutic efficacy of procaine penicillin, Phyllanthus emblica fruit extract and Cocos nucifera oil against subclinical mastitis. Livest. Sci. 2021, 251, 104655. [Google Scholar] [CrossRef]
- Almatroodi, S.A.; Alsahli, M.A.; Almatroudi, A.; Dev, K.; Rafat, S.; Verma, A.K.; Rahmani, A.H. Amla (Emblica officinalis): Role in health management via controlling various biological activities. Gene Rep. 2020, 21, 100820. [Google Scholar] [CrossRef]
- Wang, F.; Chen, L.; Chen, S.; Chen, H.; Liu, Y. Characterization of two closely related citrus cultivars using UPLC-ESI-MS/MS-based widely targeted metabolomics. PLoS ONE 2021, 16, e0254759. [Google Scholar] [CrossRef]
- Wang, K.; Nan, X.; Zhao, P.; Liu, W.; Drackley, J.K.; Liu, S.; Zhang, K.; Bu, D. Effects of low dietary cation-anion difference induced by ruminal ammonium chloride infusion on performance, serum, and urine metabolites of lactating dairy cows. Asian-Australas. J. Anim. Sci. 2018, 31, 677–685. [Google Scholar] [CrossRef]
- Drackley, J.K.; Overton, T.R.; Ortiz-Gonzalez, G.; Beaulieu, A.D.; Barbano, D.M.; Lynch, J.M.; Perkins, E.G. Responses to increasing amounts of high-oleic sunflower fatty acids infused into the abomasum of lactating dairy cows. J. Dairy Sci. 2007, 90, 5165–5175. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Bu, D.P.; Wang, J.Q.; Dhiman, T.R.; Liu, S.J. Effectiveness of Oils Rich in Linoleic and Linolenic Acids to Enhance Conjugated Linoleic Acid in Milk from Dairy Cows. J. Dairy Sci. 2007, 90, 998–1007. [Google Scholar] [CrossRef]
- Bizuayehu, D.; Atlabachew, M.; Ali, M.T. Determination of some selected secondary metabolites and their invitro antioxidant activity in commercially available Ethiopian tea (Camellia sinensis). SpringerPlus 2016, 5, 412. [Google Scholar] [CrossRef] [Green Version]
- Littell, R.C.; Milliken, G.A.; Stroup, W.W.; Wolfinger, R.D.; Oliver, S. SAS for Mixed Models; SAS Institute Inc.: Cary, NC, USA, 2006. [Google Scholar]
- Mao, X.; Wu, L.-F.; Guo, H.-L.; Chen, W.-J.; Cui, Y.-P.; Qi, Q.; Li, S.; Liang, W.-Y.; Yang, G.-H.; Shao, Y.-Y. The genus Phyllanthus: An ethnopharmacological, phytochemical, and pharmacological review. Evid. Based Complementary Altern. Med. 2016, 2016, 7584952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.; Zhang, Q.; Liang, W.; Ma, Y.; Niu, L.; Zhang, L. Phytochemical Analysis Using UPLC-MSn Combined with Network Pharmacology Approaches to Explore the Biomarkers for the Quality Control of the Anticancer Tannin Fraction of Phyllanthus emblica L. Habitat in Nepal. Evid. Based Complement. Altern. Med. 2021, 2021, 6623791. [Google Scholar] [CrossRef]
- Khaled, S.E.; Hashem, F.A.-M.; Shabana, M.H.; Hammam, A.-M.M.; Madboli, A.N.A.; Al-Mahdy, D.A.; Farag, M.A. A biochemometric approach for the assessment of Phyllanthus emblica female fertility effects as determined via UPLC-ESI-qTOF-MS and GC-MS. Food Funct. 2019, 10, 4620–4635. [Google Scholar] [CrossRef] [PubMed]
- Chilliard, Y.; Glasser, F.; Ferlay, A.; Bernard, L.; Rouel, J.; Doreau, M. Diet, rumen biohydrogenation and nutritional quality of cow and goat milk fat. Eur. J. Lipid Sci. Technol. 2007, 109, 828–855. [Google Scholar] [CrossRef]
- Ulbricht, T.L.V.; Southgate, D.A.T. Coronary heart disease: Seven dietary factors. Lancet 1991, 338, 985–992. [Google Scholar] [CrossRef]
- Buccioni, A.; Serra, A.; Minieri, S.; Mannelli, F.; Cappucci, A.; Benvenuti, D.; Rapaccini, S.; Conte, G.; Mele, M. Milk production, composition, and milk fatty acid profile from grazing sheep fed diets supplemented with chestnut tannin extract and extruded linseed. Small Rumin. Res. 2015, 130, 200–207. [Google Scholar] [CrossRef]
- Valizadeh Yonjalli, R.; Mirzaei Aghjehgheshlagh, F.; Mahdavi, A.; Navidshad, B.; Staji, H. The effects of tannin extract and linseed oil on yield, physicochemical characteristics and fatty acid profile of ewe milk. Int. J. Dairy Technol. 2020, 73, 656–666. [Google Scholar] [CrossRef]
- Kraft, J.; Jetton, T.; Satish, B.; Gupta, D. Dairy-derived bioactive fatty acids improve pancreatic ß-cell function. FASEB J. 2015, 29, 608–625. [Google Scholar] [CrossRef]
- Wongtangtintharn, S.; Oku, H.; Iwasaki, H.; Toda, T. Effect of branched-chain fatty acids on fatty acid biosynthesis of human breast cancer cells. J. Nutr. Sci. Vitam. 2004, 50, 137–143. [Google Scholar] [CrossRef] [Green Version]
- Vlaeminck, B.; Fievez, V.; Cabrita, A.; Fonseca, A.; Dewhurst, R. Factors affecting odd-and branched-chain fatty acids in milk: A review. Anim. Feed Sci. Technol. 2006, 131, 389–417. [Google Scholar] [CrossRef]
- Fitria, P.D.; Amin, M.; Lokapirnasari, W.P.; Lamid, M. Supplementation of fermented coffee-peel flour to increase high-density lipoprotein (HDL) cholesterol, docosahexaenoic acids (DHA) and eicosapentaenoic acids (EPA) deposition in tilapia fillet. Biocatal. Agric. Biotechnol. 2020, 24, 101502. [Google Scholar] [CrossRef]
- Abarghuei, M.J.; Rouzbehan, Y.; Salem, A.Z.M.; Zamiri, M.J. Nitrogen balance, blood metabolites and milk fatty acid composition of dairy cows fed pomegranate-peel extract. Livest. Sci. 2014, 164, 72–80. [Google Scholar] [CrossRef]
- Szczechowiak, J.; Szkudelska, K.; Szumacher-Strabel, M.; Sadkowski, S.; Gwozdz, K.; El-Sherbiny, M.; Kozłowska, M.; Rodriguez, V.; Cieslak, A. Blood hormones, metabolic parameters and fatty acid proportion in dairy cows fed condensed tannins and oils blend. Ann. Anim. Sci. 2018, 18, 155–166. [Google Scholar] [CrossRef] [Green Version]
- Santos-Silva, J.; Francisco, A.; Alves, S.P.; Portugal, P.; Dentinho, T.; Almeida, J.; Soldado, D.; Jerónimo, E.; Bessa, R.J. Effect of dietary neutral detergent fibre source on lambs growth, meat quality and biohydrogenation intermediates. Meat Sci. 2019, 147, 28–36. [Google Scholar] [CrossRef]
- Ishlak, A.; Günal, M.; AbuGhazaleh, A.A. The effects of cinnamaldehyde, monensin and quebracho condensed tannin on rumen fermentation, biohydrogenation and bacteria in continuous culture system. Anim. Feed Sci. Technol. 2015, 207, 31–40. [Google Scholar] [CrossRef] [Green Version]
- Buccioni, A.; Pauselli, M.; Viti, C.; Minieri, S.; Pallara, G.; Roscini, V.; Rapaccini, S.; Marinucci, M.T.; Lupi, P.; Conte, G.; et al. Milk fatty acid composition, rumen microbial population, and animal performances in response to diets rich in linoleic acid supplemented with chestnut or quebracho tannins in dairy ewes. J. Dairy Sci. 2015, 98, 1145–1156. [Google Scholar] [CrossRef] [Green Version]
- Aprianita, A.; Donkor, O.N.; Moate, P.J.; Williams, S.R.; Auldist, M.J.; Greenwood, J.S.; Hannah, M.C.; Wales, W.J.; Vasiljevic, T. Effects of dietary cottonseed oil and tannin supplements on protein and fatty acid composition of bovine milk. J Dairy Res 2014, 81, 183–192. [Google Scholar] [CrossRef]
- Kennelly, J.J. The fatty acid composition of milk fat as influenced by feeding oilseeds. Anim. Feed Sci. Technol. 1996, 60, 137–152. [Google Scholar] [CrossRef]
- Bryszak, M.; Szumacher-Strabel, M.; El-Sherbiny, M.; Stochmal, A.; Oleszek, W.; Roj, E.; Patra, A.K.; Cieslak, A. Effects of berry seed residues on ruminal fermentation, methane concentration, milk production, and fatty acid proportions in the rumen and milk of dairy cows. J. Dairy Sci. 2019, 102, 1257–1273. [Google Scholar] [CrossRef] [Green Version]
- Carreño, D.; Hervás, G.; Toral, P.G.; Belenguer, A.; Frutos, P. Ability of different types and doses of tannin extracts to modulate in vitro ruminal biohydrogenation in sheep. Anim. Feed Sci. Technol. 2015, 202, 42–51. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.W.; Gai, F.; Gasco, L.; Brugiapaglia, A.; Lussiana, C.; Guo, K.J.; Tong, J.M.; Zoccarato, I. Effects of chestnut tannins on carcass characteristics, meat quality, lipid oxidation and fatty acid composition of rabbits. Meat Sci 2009, 83, 678–683. [Google Scholar] [CrossRef]
- Alipanahi, Z.; Fatahnia, F.; Jafari, H.; Taasoli, G.; Mirzaei-Alamouti, H.; Barrett, D.; Pormalekshahi, A. Effect of oak acorn with or without polyethylene glycol in diets containing extruded soybean on milk fatty acid profile, ruminal fermentation and plasma metabolites of lactating goats. Livest. Sci. 2019, 221, 57–62. [Google Scholar] [CrossRef]
- Merlot, A.M.; Kalinowski, D.S.; Richardson, D.R. Unraveling the mysteries of serum albumin—More than just a serum protein. Front. Physiol. 2014, 5, 299. [Google Scholar] [CrossRef] [Green Version]
- Evans, T. Albumin as a drug—Biological effects of albumin unrelated to oncotic pressure. Aliment. Pharmacol. Ther. 2002, 16, 6–11. [Google Scholar] [CrossRef]
- Frutos, P.; Raso, M.; Hervás, G.; Mantecón, Á.R.; Pérez, V.; Javier Giráldez, F. Is there any detrimental effect when a chestnut hydrolysable tannin extract is included in the diet of finishing lambs? Anim. Res. 2004, 53, 127–136. [Google Scholar] [CrossRef] [Green Version]
- Govind, P.; Pandey, S. Phytochemical and toxicity study of Emblica officinalis (Amla). Int. Res. J. Pharm. 2011, 2, 270–272. [Google Scholar]
- Jaijoy, K.; Soonthornchareonnon, N.; Panthong, A.; Sireeratawong, S. Anti-inflammatory activity and toxicity of the standardised water extract of Phyllanthus emblica L. Planta Med. 2010, 76, P127. [Google Scholar] [CrossRef]
- Morsy, A.S.; Soltan, Y.A.; El-Zaiat, H.M.; Alencar, S.M.; Abdalla, A.L. Bee propolis extract as a phytogenic feed additive to enhance diet digestibility, rumen microbial biosynthesis, mitigating methane formation and health status of late pregnant ewes. Anim. Feed Sci. Technol. 2021, 273, 114834. [Google Scholar] [CrossRef]
- Danfær, A. Nutrient metabolism and utilization in the liver. Livest. Prod. Sci. 1994, 39, 115–127. [Google Scholar] [CrossRef]
- Larsen, M.; Kristensen, N. Precursors for liver gluconeogenesis in periparturient dairy cows. Animal 2013, 7, 1640–1650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, R.; Xiao, W.; Ren, G.; Zhou, D.; Tan, C.; Tan, Z.; Han, X.; Tang, S.; Zhou, C.; Wang, M. Dietary tea catechin inclusion changes plasma biochemical parameters, hormone concentrations and glutathione redox status in goats. Asian-Australas. J. Anim. Sci. 2011, 24, 1681–1689. [Google Scholar] [CrossRef]
- Zhang, X.; Jiang, C.; Jin, Y.; Li, P.; Zhong, J. The effect of substitution of mixed grass hay with Urtica cannabina hay and/or Leymus chinensis hay on blood biochemical profile, carcass traits, and intramuscular fatty acid composition in finishing lambs. Anim. Feed Sci. Technol. 2021, 272, 114780. [Google Scholar] [CrossRef]
- Cozzi, G.; Ravarotto, L.; Gottardo, F.; Stefani, A.L.; Contiero, B.; Moro, L.; Brscic, M.; Dalvit, P. Short communication: Reference values for blood parameters in Holstein dairy cows: Effects of parity, stage of lactation, and season of production. J. Dairy Sci. 2011, 94, 3895–3901. [Google Scholar] [CrossRef]
- Singla, A.; Hundal, J.S.; Patra, A.K.; Wadhwa, M.; Nagarajappa, V.; Malhotra, P. Effect of dietary supplementation of Emblica officinalis fruit pomace on methane emission, ruminal fermentation, nutrient utilization, and milk production performance in buffaloes. Environ. Sci. Pollut. Res. 2021, 28, 18120–18133. [Google Scholar] [CrossRef]
- Costa, E.d.S.; Ribiero, C.; Silva, T.; Ribeiro, R.; Vieira, J.; Lima, A.d.O.; Barbosa, A.; da Silva Júnior, J.; Bezerra, L.; Oliveira, R. Intake, nutrient digestibility, nitrogen balance, serum metabolites and growth performance of lambs supplemented with Acacia mearnsii condensed tannin extract. Anim. Feed Sci. Technol. 2021, 272, 114744. [Google Scholar] [CrossRef]
- Dawson, T.M.; Dawson, V.L. REVIEW: Nitric Oxide: Actions and Pathological Roles. Neuroscientist 1995, 1, 7–18. [Google Scholar] [CrossRef]
- Chan, M.M.-Y.; Mattiacci, J.A.; Hwang, H.S.; Shah, A.; Fong, D. Synergy between ethanol and grape polyphenols, quercetin, and resveratrol, in the inhibition of the inducible nitric oxide synthase pathway. Biochem. Pharmacol. 2000, 60, 1539–1548. [Google Scholar] [CrossRef]
- Srivastava, R.; Husain, M.; Hasan, S.; Athar, M. Green tea polyphenols and tannic acid act as potent inhibitors of phorbol ester-induced nitric oxide generation in rat hepatocytes independent of their antioxidant properties. Cancer Lett. 2000, 153, 1–5. [Google Scholar] [CrossRef]
- Santos, N.; Santos, G.; Silva-Kazama, D.; Grande, P.; Pintro, P.; De Marchi, F.; Jobim, C.; Petit, H. Production, composition and antioxidants in milk of dairy cows fed diets containing soybean oil and grape residue silage. Livest. Sci. 2014, 159, 37–45. [Google Scholar] [CrossRef]
- Delgadillo-Puga, C.; Cuchillo-Hilario, M.; León-Ortiz, L.; Ramírez-Rodríguez, A.; Cabiddu, A.; Navarro-Ocaña, A.; Morales-Romero, A.M.; Medina-Campos, O.N.; Pedraza-Chaverri, J. Goats’ feeding supplementation with Acacia farnesiana pods and their relationship with milk composition: Fatty acids, polyphenols, and antioxidant activity. Animals 2019, 9, 515. [Google Scholar] [CrossRef] [Green Version]
- Emami, A.; Ganjkhanlou, M.; Nasri, M.F.; Zali, A.; Rashidi, L.; Sharifi, M. Antioxidant status of dairy goats fed diets containing pomegranate seed oil or linseed oil. Small Rumin. Res. 2017, 153, 175–179. [Google Scholar] [CrossRef]
- De Feo, V.; Quaranta, E.; Fedele, V.; Claps, S.; Rubino, R.; Pizza, C. Flavonoids and terpenoids in goat milk in relation to forage intake. Ital. J. Food. Sci. 2006, 18, 85–92. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Item | Value |
|---|---|
| TMR | |
| Ingredient, g/kg DM | |
| Corn silage | 347.6 ± 0.64 |
| Alfalfa hay | 152.5 ± 0.62 |
| Concentrate mix 1 | 499.9 ± 0.06 |
| Chemical composition (g/kg DM) | |
| Dry matter | 488 ± 4.1 |
| Ash | 86.7 ± 6.41 |
| Crude protein (CP) | 164 ± 1.5 |
| Neutral detergent fiber (aNDF) | 291 ± 4.7 |
| Acid detergent fiber | 173 ± 5.4 |
| Ether extract (EE) | 14.4 ± 0.96 |
| Non-fiber carbohydrates (NFC) * | 451 ± 8.8 |
| Acid insoluble ash | 43.1 ± 4.92 |
| Item | Alfalfa Hay | Corn Silage | Concentrate Mixture | Amla Fruit | SE * |
|---|---|---|---|---|---|
| Chemical composition (g/kg DM unless noted) | |||||
| Dry matter | 872 | 286 | 860 | 231 | 4.1 |
| Ash | 103 | 64 | 83 | 28 | 4.6 |
| Crude protein (CP) | 169 | 73.0 | 224 | 35.4 | 2.41 |
| Neutral detergent fiber (aNDF) | 387 | 413 | 172 | 347 | 5.2 |
| Acid detergent fiber | 275 | 257 | 83.6 | 237 | 7.63 |
| Ether extract (EE) | 8.13 | 7.07 | 30.4 | 6.44 | 1.422 |
| Non-fiber carbohydrates 1 | 332 | 453 | 514 | 588 | 8.8 |
| Total phenolic content (mg TA/g DM) 2 | - | - | - | 51.2 | 1.84 |
| Total flavonoid content (mg QT/g DM) 3 | - | - | - | 87.8 | 1.63 |
| Cow | Weeks | |||||||
|---|---|---|---|---|---|---|---|---|
| Adaptation | Period 1 | Washout | Period 2 | |||||
| −2 | 2 | 4 | 6 | 8 | 10 | 12 | 14 | |
| Fresh amla fruit (g/d) | ||||||||
| MY010 | 0 | 200 | 400 | 600 | 0 | 0 | 0 | 0 |
| MY001 | 0 | 200 | 400 | 600 | 0 | 0 | 0 | 0 |
| P71 | 0 | 200 | 400 | 600 | 0 | 0 | 0 | 0 |
| 090922 | 0 | 200 | 400 | 600 | 0 | 0 | 0 | 0 |
| 1108 | 0 | 0 | 0 | 0 | 0 | 200 | 400 | 600 |
| MY002 | 0 | 0 | 0 | 0 | 0 | 200 | 400 | 600 |
| P34 | 0 | 0 | 0 | 0 | 0 | 200 | 400 | 600 |
| P72 | 0 | 0 | 0 | 0 | 0 | 200 | 400 | 600 |
| Items | Fresh Amla Fruit Doses Supplemented g/day | Treatment by Dose, p | ||||||
|---|---|---|---|---|---|---|---|---|
| 0 | 200 | 400 | 600 | SE * | Linear | Quadratic | Cubic | |
| Saturated FA (SFA) | ||||||||
| C10:0 | 0.67 | 0.72 (0.67) 1 | 0.15 (0.69) | 0.98 (0.67) | 0.13 0.15 | 0.45 | <0.01 | <0.001 |
| C12:0 | 2.38 | 2.50 (2.70) | 0.90 (1.90) | 2.84 (2.43) | 0.22 0.20 | 0.79 | <0.001 | <0.001 |
| C14:0 | 9.67 | 8.99 (10.0) | 6.38 (8.98) | 10.0 (9.88) | 0.50 0.40 | 0.47 | <0.001 | <0.01 |
| C15:0 | 1.34 | 1.35 (1.33) | 1.00 (1.41) | 1.42 (1.27) | 0.01 0.15 | 0.83 | 0.05 | 0.03 |
| C16:0 (palmitic acid) | 43.2 | 39.6 (41.7) | 34.4 (44.9) | 43.0 (42.5) | 1.5 1.6 | 0.37 | <0.001 | 0.03 |
| C17:0 | 0.63 | 0.72 (0.66) | 0.75 (0.64) | 0.61 (0.57) | 0.04 0.04 | 0.86 | <0.01 | 0.53 |
| C18:0 (stearic acid) | 6.34 | 7.17 (6.34) | 11.2 (6.00) | 6.95 (6.63) | 0.73 0.87 | 0.06 | <0.001 | <0.01 |
| C20:0 | 0.20 | 0.43 (0.18) | 0.89 (0.17) | 0.39 (0.22) | 0.17 0.02 | 0.19 | 0.04 | 0.16 |
| C21:0 | 1.02 | 1.32 (0.87) | 1.94 (1.01) | 1.40 (1.16) | 0.25 0.10 | 0.12 | 0.10 | 0.23 |
| C22:0 | 0.29 | 0.57 (0.25) | 0.93 (0.31) | 0.44 (0.29) | 0.17 0.07 | 0.27 | 0.02 | 0.25 |
| C23:0 | 0.91 | 1.47 (0.94) | 1.25 (0.88) | 0.99 (0.91) | 0.26 0.27 | 0.99 | 0.13 | 0.57 |
| C24:0 | 0.15 | 0.24 (0.13) | 0.19 (0.12) | 0.16 (0.24) | 0.04 0.03 | 0.81 | 0.16 | 0.43 |
| Branched chain FA (BCFA) | ||||||||
| iso C13:0 | 0.10 | 0.13 (0.11) | 0.06 (0.08) | 0.10 (0.11) | 0.02 0.01 | 0.29 | 0.55 | <0.01 |
| iso C15:0 | 0.14 | 0.36 (0.19) | 0.05 (0.06) | 0.06 (0.15) | 0.05 0.04 | 0.03 | 0.07 | <0.01 |
| iso C17:0 | 0.51 | 0.50 (0.54) | 0.51 (0.48) | 0.46 (0.48) | 0.04 0.02 | 0.44 | 0.54 | 0.65 |
| anteiso C13:0 | 0.12 | 0.12 (0.13) | 0.11 (0.12) | 0.10 (0.11) | 0.02 0.02 | 0.32 | 0.84 | 0.98 |
| anteiso C15:0 | 0.22 | 0.24 (0.23) | 0.18 (0.21) | 0.22 (0.20) | 0.02 0.01 | 0.57 | 0.47 | 0.05 |
| anteiso C17:0 | 0.65 | 0.90 (0.73) | 1.05 (0.58) | 0.55 (0.60) | 0.10 0.05 | 0.70 | <0.001 | 0.19 |
| ∑BCFA 2 | 1.95 | 2.54 (2.27) | 2.22 (1.73) | 1.71 (1.87) | 0.14 0.07 | 0.08 | <0.001 | 0.26 |
| ∑OBCFA 3 | 4.15 | 4.92 (4.51) | 4.16 (3.94) | 3.94 (3.93) | 0.22 0.19 | 0.11 | 0.02 | 0.03 |
| ∑SFA 4 | 66.7 | 65.4 (66.1) | 60.3 (67.1) | 69.5 (66.9) | 1.3 1.3 | 0.52 | <0.001 | 0.01 |
| Monounsaturated FA (MUFA) | ||||||||
| C18:1 trans-9 | 0.41 | 0.44 (0.45) | 0.45 (0.37) | 0.37 (0.39) | 0.04 0.04 | 0.55 | 0.15 | 0.71 |
| C18:1 cis-9 (oleic acid) | 18.1 | 19.3 (18.3) | 23.5 (18.3) | 17.5 (17.9) | 1.4 1.2 | 0.63 | <0.01 | 0.04 |
| C18:1 trans-11 (vaccenic acid) | 0.97 | 1.05 (0.98) | 1.12 (0.93) | 0.90 (1.02) | 0.08 0.08 | 0.74 | 0.08 | 0.49 |
| C24:1 | 0.27 | 0.60 (0.24) | 1.01 (0.29) | 0.13 (0.25) | 0.21 0.08 | 0.99 | <0.01 | 0.17 |
| ∑MUFA 5 | 25.1 | 26.9 (25.3) | 31.2 (25.1) | 24.0 (25.3) | 1.2 1.0 | 0.85 | <0.001 | 0.01 |
| Polyunsaturated FA (PUFA) | ||||||||
| C18:2 trans-9,12 | 0.38 | 0.36 (0.54) | 0.28 (0.21) | 0.34 (0.36) | 0.06 0.06 | 0.42 | 0.51 | 0.52 |
| C18:2 cis-9,12 (linoleic acid) | 2.48 | 2.38 (2.92) | 2.35 (2.09) | 2.25 (2.36) | 0.19 0.20 | 0.38 | 1.00 | 0.89 |
| C18:3 cis-9 (α-linolenic acid) | 0.32 | 0.34 (0.35) | 0.33 (0.30) | 0.27 (0.31) | 0.03 0.03 | 0.24 | 0.27 | 0.88 |
| C20:3 | 0.15 | 0.28 (0.11) | 0.16 (0.18) | 0.11 (0.15) | 0.05 0.05 | 0.27 | 0.06 | 0.20 |
| C22:2 (Docosadienoic acid) | 0.07 | 0.17 (0.08) | 0.19 (0.06) | 0.13 (0.08) | 0.02 0.02 | 0.06 | <0.01 | 0.98 |
| C20:5 (Eicosapentaenoic acid, EPA) | 0.11 | 0.30 (0.09) | 0.52 (0.12) | 0.08 (0.14) | 0.14 0.03 | 0.86 | 0.03 | 0.29 |
| C22:6 (Docosahexaenoic acid, DHA) | 0.28 | 0.47 (0.28) | 0.60 (0.28) | 0.18 (0.28) | 0.11 0.06 | 0.69 | <0.01 | 0.33 |
| ∑n-3 PUFA 6 | 0.87 | 1.38 (0.84) | 1.57 (0.88) | 0.64 (0.88) | 025 0.12 | 0.65 | <0.01 | 0.50 |
| ∑n-6 PUFA 7 | 3.02 | 3.01 (3.63) | 2.94 (2.45) | 2.82 (2.95) | 0.23 0.21 | 0.49 | 0.81 | 0.97 |
| n-6/n-3 ratio 8 | 3.81 | 2.68 (4.60) | 3.14 (3.23) | 4.15 (3.48) | 0.49 0.45 | 0.50 | 0.04 | 0.66 |
| ∑PUFA 9 | 3.83 | 4.38 (4.47) | 4.51 (3.33) | 3.47 (3.83) | 0.35 0.25 | 0.53 | 0.02 | 0.65 |
| ∑Unsaturated FA (UFA) 10 | 29.1 | 31.2 (29.7) | 35.9 (28.4) | 27.5 (29.2) | 1.2 1.0 | 0.98 | <0.001 | 0.01 |
| ∑Other FA | 6.69 | 7.51 (6.73) | 7.00 (6.31) | 6.06 (7.03) | 0.47 0.29 | 0.24 | <0.07 | 0.68 |
| Atherogenicity index (AI) 11 | 2.91 | 2.51 (2.87) | 1.77 (2.95) | 3.12 (2.92) | 0.17 0.18 | 0.87 | <0.001 | <0.01 |
| h/H ratio 12 | 0.41 | 0.47 (0.42) | 0.70 (0.40) | 0.38 (0.40) | 0.05 0.03 | 0.50 | <0.001 | <0.01 |
| Spreadability index (SI) 13 | 2.46 | 2.06 (2.35) | 1.55 (2.55) | 2.43 (2.45) | 0.18 0.24 | 0.44 | <0.01 | 0.09 |
| PC1 | PC2 | PC3 | PC4 | |
|---|---|---|---|---|
| C10:0 | −0.502 | 0.317 | −0.190 | 0.293 |
| C12:0 | −0.680 | 0.349 | −0.214 | 0.119 |
| iso C13:0 | −0.301 | 0.429 | −0.557 | 0.285 |
| C14:0 | −0.807 | 0.295 | −0.027 | −0.120 |
| iso C15:0 | −0.061 | 0.392 | −0.795 | 0.132 |
| C15:0 | −0.630 | 0.369 | 0.120 | −0.574 |
| C16:0 (palmitic acid) | −0.909 | 0.133 | 0.096 | −0.092 |
| anteiso C17:0 | 0.751 | 0.501 | 0.144 | −0.155 |
| C17:0 | 0.242 | 0.004 | −0.037 | −0.854 |
| C18:0 (stearic acid) | 0.634 | −0.574 | −0.046 | −0.176 |
| C18:1 trans-11 (vaccenic acid) | 0.182 | −0.425 | −0.474 | −0.180 |
| C18:1 cis-9 (oleic acid) | 0.540 | −0.766 | −0.256 | −0.104 |
| C20:0 | 0.755 | 0.452 | 0.325 | 0.085 |
| C21:0 | 0.658 | 0.410 | 0.423 | 0.147 |
| C22:0 | 0.727 | 0.426 | 0.370 | 0.087 |
| C22:2 (Docosadienoic acid) | 0.542 | 0.088 | 0.026 | 0.264 |
| C20:5 (Eicosapentaenoic acid, EPA) | 0.716 | 0.440 | 0.121 | 0.011 |
| C22:6 (Docosahexaenoic acid, DHA) | 0.775 | 0.517 | 0.016 | 0.094 |
| ∑BCFA 1 | 0.575 | 0.648 | −0.420 | −0.096 |
| ∑OBCFA 2 | 0.144 | 0.750 | −0.331 | −0.530 |
| ∑SFA 3 | −0.870 | 0.247 | 0.166 | −0.213 |
| ∑MUFA 4 | 0.700 | −0.638 | −0.247 | −0.096 |
| ∑PUFA 5 | 0.681 | 0.509 | −0.115 | 0.020 |
| ∑UFA 6 | 0.816 | −0.466 | −0.258 | −0.084 |
| ∑isoFA | 0.049 | 0.401 | −0.857 | 0.066 |
| ∑antisoFA | 0.734 | 0.567 | 0.078 | −0.178 |
| ∑PUFAn-3 7 | 0.777 | 0.512 | 0.086 | 0.059 |
| Atherogenicity index (AI) 8 | −0.911 | 0.335 | 0.144 | −0.078 |
| h/H ratio 9 | 0.840 | −0.452 | −0.145 | −0.077 |
| Eigenvalue | 12.381 | 6.159 | 3.043 | 1.898 |
| Variability (%) | 42.694 | 21.237 | 10.492 | 6.544 |
| Cumulative % | 42.694 | 63.931 | 74.423 | 80.967 |
| Item | Fresh Amla Fruit Doses Supplemented g/Day | Treatment by Dose, p | ||||||
|---|---|---|---|---|---|---|---|---|
| 0 | 200 | 400 | 600 | SE | Linear | Quadratic | Cubic | |
| Serum blood chemistry | ||||||||
| β-hydroxybutyrate | 0.22 | 0.27 (0.18) | 0.25 (0.26) | 0.29 (0.23) | 0.03 0.04 | 0.26 | 0.89 | 0.35 |
| Glucose | 70.2 | 85.6 (67.0) | 84.5 (71.4) | 76.5 (72.0) | 4.1 3.8 | 0.27 | <0.01 | 0.61 |
| Non-esterified fatty acid | 0.30 | 0.31 (0.37) | 0.23 (0.30) | 0.33 (0.23) | 0.04 0.05 | 0.88 | 0.28 | 0.23 |
| Triglycerides | 0.12 | 0.12 (0.11) | 0.10 (0.13) | 0.11 (0.13) | 0.02 0.02 | 0.44 | 0.68 | 0.36 |
| Total cholesterol | 5.17 | 5.47 (5.00) | 5.95 (5.36) | 4.51 (5.16) | 0.34 0.42 | 0.26 | 0.01 | 0.19 |
| Total protein | 55.3 | 58.2 (53.1) | 65.9 (56.7) | 43.8 (56.3) | 2.3 2.5 | 0.01 | <0.001 | 0.01 |
| Albumin (A) | 22.8 | 20.5 (21.2) | 31.4 (24.1) | 21.8 (23.2) | 1.2 1.5 | 0.14 | 0.01 | <0.001 |
| Globulin (G) | 32.5 | 37.8 (31.9) | 34.4 (32.5) | 19.4 (33.1) | 2.3 2.4 | <0.001 | <0.001 | 0.78 |
| A/G ratio | 0.74 | 0.56 (0.68) | 1.06 (0.79) | 1.21 (0.74) | 0.10 0.17 | <0.001 | 0.13 | 0.06 |
| Aspartate aminotransferase | 10.7 | 9.38 (11.0) | 9.79 (10.8) | 9.79 (10.3) | 0.44 0.50 | 0.23 | 0.15 | 0.31 |
| Gamma-Glutamyl Transferase | 13.7 | 16.2 (11.0) | 16.0 (15.2) | 17.1 (14.7) | 1.5 1.6 | 0.12 | 0.65 | 0.57 |
| Alanine aminotransferase | 11.0 | 14.8 (10.5) | 11.4 (11.0) | 12.0 (11.5) | 1.4 1.8 | 0.94 | 0.28 | 0.11 |
| Plasma blood chemistry | ||||||||
| Creatine | 59.4 | 60.0 (58.8) | 59.1 (59.0) | 59.9 (61.0) | 2.9 2.5 | 0.96 | 0.98 | 0.83 |
| Lactic acid | 0.93 | 1.10 (1.01) | 0.85 (0.79) | 0.90 (0.98) | 0.07 0.07 | 0.30 | 0.41 | 0.04 |
| Uric acid | 47.0 | 50.9 (46.1) | 50.1 (46.9) | 50.7 (49.2) | 2.3 4.7 | 0.27 | 0.48 | 0.60 |
| Item | Fresh Amla Fruit Doses Supplemented g/Day | Treatment by Dose, p | ||||||
|---|---|---|---|---|---|---|---|---|
| 0 | 200 | 400 | 600 | SE | Linear | Quadratic | Cubic | |
| Plasma | ||||||||
| Malondialdehyde (MDA) | 12.6 | 12.1 (14.0) | 12.8 (12.5) | 11.6 (13.7) | 1.1 0.97 | 0.61 | 0.76 | 0.57 |
| Nitric oxide (NO) | 10.5 | 10.8(9.55) | 10.5(11.8) | 6.07(10.1) | 1.1 1.2 | <0.01 | 0.04 | 0.51 |
| Superoxide dismutase (SOD) | 22.4 | 20.5(22.2) | 25.2(22.1) | 24.1(22.7) | 0.92 0.78 | 0.01 | 0.63 | <0.01 |
| Milk | ||||||||
| Total antioxidant capacity (TAC) | 0.97 | 1.00 (0.95) | 1.09 (1.02) | 1.09 (1.05) | 0.05 0.06 | <0.01 | 0.65 | 0.32 |
| ferric reducing-antioxidant power (FRAP) | 0.62 | 0.49 (0.53) | 0.75 (0.65) | 0.57 (0.68) | 0.03 0.04 | 0.43 | 0.45 | <0.001 |
| Flavonoid | 42.9 | 63.0 (44.5) | 66.7 (48.7) | 36.7 (35.5) | 3.1 3.5 | 0.32 | <0.001 | 0.32 |
| Ascorbic | 0.03 | 0.04 (0.03) | 0.06 (0.03) | 0.03 (0.03) | 0.01 0.01 | 0.49 | 0.01 | 0.15 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tilahun, M.; Zhao, L.; Sun, L.; Shen, Y.; Ma, L.; Callaway, T.R.; Xu, J.; Bu, D. Fresh Phyllanthus emblica (Amla) Fruit Supplementation Enhances Milk Fatty Acid Profiles and the Antioxidant Capacities of Milk and Blood in Dairy Cows. Antioxidants 2022, 11, 485. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11030485
Tilahun M, Zhao L, Sun L, Shen Y, Ma L, Callaway TR, Xu J, Bu D. Fresh Phyllanthus emblica (Amla) Fruit Supplementation Enhances Milk Fatty Acid Profiles and the Antioxidant Capacities of Milk and Blood in Dairy Cows. Antioxidants. 2022; 11(3):485. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11030485
Chicago/Turabian StyleTilahun, Mekonnen, Liansheng Zhao, Lingling Sun, Yifan Shen, Lu Ma, Todd R. Callaway, Jianchu Xu, and Dengpan Bu. 2022. "Fresh Phyllanthus emblica (Amla) Fruit Supplementation Enhances Milk Fatty Acid Profiles and the Antioxidant Capacities of Milk and Blood in Dairy Cows" Antioxidants 11, no. 3: 485. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11030485