
A Pathway-Based Genetic Score for Oxidative Stress: An Indicator of Host Vulnerability to Phthalate-Associated Adverse Neurodevelopment

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Cohort
2.2. Phthalate Measurement
2.3. Child Neurodevelopmental Outcomes
2.4. Genotyping
2.5. Genetic Pathway Function Score for Oxidative Stress
2.6. Statistical Analysis
3. Results
3.1. Association between Phthalate Daily Intake and Neurodevelopmental Outcomes
3.2. Association between gPFSox and Neurodevelopmental Outcomes
3.3. Interplay between gPFSox and Prenatal Phthalate Exposure against Neurodevelopmental Outcomes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Centers for Disease Control and Prevention. Prevalence of Autism Spectrum Disorders—Autism and Developmental Disabilities Monitoring Network, Six Sites, United States, 2000; Morbidity and Mortality Weekly Report; Surveillance summaries; Centers for Disease Control and Prevention: Washington, DC, USA, 2007; Volume 56, pp. 1–11. [Google Scholar]
- Baio, J.; Wiggins, L.; Christensen, D.L.; Maenner, M.J.; Daniels, J.; Warren, Z.; Kurzius-Spencer, M.; Zahorodny, W.; Robinson Rosenberg, C.; White, T.; et al. Prevalence of Autism Spectrum Disorder among Children Aged 8 Years-Autism and Developmental Disabilities Monitoring Network, 11 Sites, United States, 2014; Morbidity and Mortality Weekly Report; Surveillance summaries; Centers for Disease Control and Prevention: Washington, DC, USA, 2018; Volume 67, pp. 1–23. [Google Scholar]
- Atladottir, H.O.; Gyllenberg, D.; Langridge, A.; Sandin, S.; Hansen, S.N.; Leonard, H.; Gissler, M.; Reichenberg, A.; Schendel, D.E.; Bourke, J. The increasing prevalence of reported diagnoses of childhood psychiatric disorders: A descriptive multinational comparison. Eur. Child Adolesc. Psychiatry 2015, 24, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Kuo, H.-T.; Muo, C.-H.; Chang, Y.-T.; Lin, C.-K. Change in prevalence status for children with developmental delay in taiwan: A nationwide population-based retrospective study. Neuropsychiatr. Dis. Treat. 2015, 11, 1541. [Google Scholar] [PubMed] [Green Version]
- Boyle, C.A.; Boulet, S.; Schieve, L.A.; Cohen, R.A.; Blumberg, S.J.; Yeargin-Allsopp, M.; Visser, S.; Kogan, M.D. Trends in the prevalence of developmental disabilities in us children, 1997–2008. Pediatrics 2011, 127, 1034–1042. [Google Scholar] [CrossRef] [Green Version]
- Matson, J.L.; Kozlowski, A.M. The increasing prevalence of autism spectrum disorders. Res. Autism Spectr. Disord. 2011, 5, 418–425. [Google Scholar] [CrossRef]
- Miguel, P.M.; Pereira, L.O.; Silveira, P.P.; Meaney, M.J. Early environmental influences on the development of children’s brain structure and function. Dev. Med. Child Neurol. 2019, 61, 1127–1133. [Google Scholar] [CrossRef]
- Lucaccioni, L.; Trevisani, V.; Passini, E.; Righi, B.; Plessi, C.; Predieri, B.; Iughetti, L. Perinatal exposure to phthalates: From endocrine to neurodevelopment effects. Int. J. Mol. Sci. 2021, 22, 4063. [Google Scholar] [CrossRef]
- WHO. United Nations Environment Programme. State of the Science of Endocrine Disrupting Chemicals-2012; World Health Organization: Geneva, Switzerland, 2015. [Google Scholar]
- WHO. Toxicological and Health Aspects of Bisphenol A; Report of Joint FAO/WHO Expert Meeting, 2–5 November 2010; Report of Stakeholder Meeting on Bisphenol A, 1 November 2010, Ottawa, Canada; World Health Organization: Geneva, Switzerland, 2011. [Google Scholar]
- Benjamin, S.; Masai, E.; Kamimura, N.; Takahashi, K.; Anderson, R.C.; Faisal, P.A. Phthalates impact human health: Epidemiological evidences and plausible mechanism of action. J. Hazard. Mater. 2017, 340, 360–383. [Google Scholar] [CrossRef]
- Gore, A.C.; Chappell, V.A.; Fenton, S.E.; Flaws, J.A.; Nadal, A.; Prins, G.S.; Toppari, J.; Zoeller, R.T. Edc-2: The endocrine society’s second scientific statement on endocrine-disrupting chemicals. Endocr. Rev. 2015, 36, E1–E150. [Google Scholar] [CrossRef]
- Net, S.; Sempéré, R.; Delmont, A.; Paluselli, A.; Ouddane, B. Occurrence, fate, behavior and ecotoxicological state of phthalates in different environmental matrices. Environ. Sci. Technol. 2015, 49, 4019–4035. [Google Scholar] [CrossRef]
- Bräuner, E.V.; Uldbjerg, C.S.; Lim, Y.H.; Gregersen, L.S.; Krause, M.; Frederiksen, H.; Andersson, A.M. Presence of parabens, phenols and phthalates in paired maternal serum, urine and amniotic fluid. Environ. Int. 2022, 158, 106987. [Google Scholar] [CrossRef]
- Sugeng, E.J.; Symeonides, C.; O’Hely, M.; Vuillermin, P.; Sly, P.D.; Vijayasarathy, S.; Thompson, K.; Pezic, A.; Mueller, J.F.; Ponsonby, A.-L. Predictors with regard to ingestion, inhalation and dermal absorption of estimated phthalate daily intakes in pregnant women: The barwon infant study. Environ. Int. 2020, 139, 105700. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Chen, W.; Zhu, H.; Wang, C. Determination of free and total phthalates in commercial whole milk products in different packaging materials by gas chromatography-mass spectrometry. J. Dairy Sci. 2015, 98, 8278–8284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serrano, S.E.; Karr, C.J.; Seixas, N.S.; Nguyen, R.H.; Barrett, E.S.; Janssen, S.; Redmon, B.; Swan, S.H.; Sathyanarayana, S. Dietary phthalate exposure in pregnant women and the impact of consumer practices. Int. J. Environ. Res. Public Health 2014, 11, 6193–6215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Castro, M.I.; Olea-Serrano, M.F.; Rivas-Velasco, A.M.; Medina-Rivero, E.; Ordoñez-Acevedo, L.G.; De León-Rodríguez, A. Phthalates and bisphenols migration in mexican food cans and plastic food containers. Bull. Environ. Contam. Toxicol. 2011, 86, 627–631. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Zhou, F.; Wang, Y.; Ning, Y.; Yang, J.Y.; Zhou, Y.K. Phthalate levels and related factors in children aged 6–12 years. Environ. Pollut. 2017, 220, 990–996. [Google Scholar] [CrossRef] [PubMed]
- Buckley, J.P.; Palmieri, R.T.; Matuszewski, J.M.; Herring, A.H.; Baird, D.D.; Hartmann, K.E.; Hoppin, J.A. Consumer product exposures associated with urinary phthalate levels in pregnant women. J. Expo. Sci. Environ. Epidemiol. 2012, 22, 468–475. [Google Scholar] [CrossRef]
- Parlett, L.E.; Calafat, A.M.; Swan, S.H. Women’s exposure to phthalates in relation to use of personal care products. J. Expo. Sci. Environ. Epidemiol. 2013, 23, 197–206. [Google Scholar] [CrossRef]
- Eales, J.; Bethel, A.; Galloway, T.; Hopkinson, P.; Morrissey, K.; Short, R.E.; Garside, R. Human health impacts of exposure to phthalate plasticizers: An overview of reviews. Environ. Int. 2022, 158, 106903. [Google Scholar] [CrossRef]
- Lee, D.W.; Kim, M.S.; Lim, Y.H.; Lee, N.; Hong, Y.C. Prenatal and postnatal exposure to di-(2-ethylhexyl) phthalate and neurodevelopmental outcomes: A systematic review and meta-analysis. Environ. Res. 2018, 167, 558–566. [Google Scholar] [CrossRef]
- Radke, E.G.; Braun, J.M.; Nachman, R.M.; Cooper, G.S. Phthalate exposure and neurodevelopment: A systematic review and meta-analysis of human epidemiological evidence. Environ. Int. 2020, 137, 105408. [Google Scholar] [CrossRef]
- Ejaredar, M.; Nyanza, E.C.; Ten Eycke, K.; Dewey, D. Phthalate exposure and childrens neurodevelopment: A systematic review. Environ. Res. 2015, 142, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Chen, X.Z.; Huang, X.; Wang, M.; Wu, J. The association between prenatal exposure to phthalates and cognition and neurobehavior of children-evidence from birth cohorts. Neurotoxicology 2019, 73, 199–212. [Google Scholar] [CrossRef] [PubMed]
- Miodovnik, A.; Engel, S.M.; Zhu, C.; Ye, X.; Soorya, L.V.; Silva, M.J.; Calafat, A.M.; Wolff, M.S. Endocrine disruptors and childhood social impairment. Neurotoxicology 2011, 32, 261–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braun, J.M.; Kalkbrenner, A.E.; Just, A.C.; Yolton, K.; Calafat, A.M.; Sjödin, A.; Hauser, R.; Webster, G.M.; Chen, A.; Lanphear, B.P. Gestational exposure to endocrine-disrupting chemicals and reciprocal social, repetitive, and stereotypic behaviors in 4-and 5-year-old children: The home study. Environ. Health Perspect. 2014, 122, 513–520. [Google Scholar] [CrossRef]
- Haggerty, D.K.; Strakovsky, R.S.; Talge, N.M.; Carignan, C.C.; Glazier-Essalmi, A.N.; Ingersoll, B.R.; Karthikraj, R.; Kannan, K.; Paneth, N.S.; Ruden, D.M. Prenatal phthalate exposures and autism spectrum disorder symptoms in low-risk children. Neurotoxicol. Teratol. 2021, 83, 106947. [Google Scholar] [CrossRef]
- Oulhote, Y.; Lanphear, B.; Braun, J.M.; Webster, G.M.; Arbuckle, T.E.; Etzel, T.; Forget-Dubois, N.; Seguin, J.R.; Bouchard, M.F.; MacFarlane, A.J.E. Gestational exposures to phthalates and folic acid, and autistic traits in canadian children. Environ. Health Perspect. 2020, 128, 027004. [Google Scholar] [CrossRef]
- Ponsonby, A.-L.; Symeonides, C.; Saffery, R.; Mueller, J.F.; O’Hely, M.; Sly, P.D.; Wardrop, N.; Pezic, A.; Mansell, T.; Collier, F. Prenatal phthalate exposure, oxidative stress-related genetic vulnerability and early life neurodevelopment: A birth cohort study. Neurotoxicology 2020, 80, 20–28. [Google Scholar] [CrossRef]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative stress: Harms and benefits for human health. Oxidative Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef]
- Ferguson, K.K.; Chen, Y.-H.; VanderWeele, T.J.; McElrath, T.F.; Meeker, J.D.; Mukherjee, B. Mediation of the relationship between maternal phthalate exposure and preterm birth by oxidative stress with repeated measurements across pregnancy. Environ. Health Perspect. 2017, 125, 488–494. [Google Scholar] [CrossRef]
- Wu, Y.; Li, J.; Yan, B.; Zhu, Y.; Liu, X.; Chen, M.; Li, D.; Lee, C.-C.; Yang, X.; Ma, P. Oral exposure to dibutyl phthalate exacerbates chronic lymphocytic thyroiditis through oxidative stress in female wistar rats. Sci. Rep. 2017, 7, 15469. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Li, K.; Zuo, H.; Yuan, Y.; Sun, Y.; Yang, X. Primary neuronal-astrocytic co-culture platform for neurotoxicity assessment of di-(2-ethylhexyl) phthalate. J. Environ. Sci. 2014, 26, 1145–1153. [Google Scholar] [CrossRef]
- Wang, X.; Michaelis, E.K. Selective neuronal vulnerability to oxidative stress in the brain. Front. Aging Neurosci. 2010, 2, 12. [Google Scholar] [CrossRef] [PubMed]
- Kushima, I.; Aleksic, B.; Nakatochi, M.; Shimamura, T.; Okada, T.; Uno, Y.; Morikawa, M.; Ishizuka, K.; Shiino, T.; Kimura, H.; et al. Comparative analyses of copy-number variation in autism spectrum disorder and schizophrenia reveal etiological overlap and biological insights. Cell Rep. 2018, 24, 2838–2856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, J.; Cai, J.J.; Ji, G.; Sham, P.C. Commonality in dysregulated expression of gene sets in cortical brains of individuals with autism, schizophrenia, and bipolar disorder. Transl. Psychiatry 2019, 9, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lintas, C.; Sacco, R.; Persico, A.M. Genome-wide expression studies in autism spectrum disorder, rett syndrome, and down syndrome. Neurobiol. Dis. 2012, 45, 57–68. [Google Scholar] [CrossRef]
- Bowers, K.; Li, Q.; Bressler, J.; Avramopoulos, D.; Newschaffer, C.; Fallin, M.D. Glutathione pathway gene variation and risk of autism spectrum disorders. J. Neurodev. Disord. 2011, 3, 132–143. [Google Scholar] [CrossRef] [Green Version]
- Heslop, C.L.; Tebbutt, S.J.; Podder, M.; Ruan, J.; Hill, J.S. Combined polymorphisms in oxidative stress genes predict coronary artery disease and oxidative stress in coronary angiography patients. Ann. Hum. Genet. 2012, 76, 435–447. [Google Scholar] [CrossRef]
- Bind, M.-A.; Coull, B.; Suh, H.; Wright, R.; Baccarelli, A.; Vokonas, P.; Schwartz, J. A novel genetic score approach using instruments to investigate interactions between pathways and environment: Application to air pollution. PLoS ONE 2014, 9, e96000. [Google Scholar]
- Dudbridge, F. Power and predictive accuracy of polygenic risk scores. PLoS Genet. 2013, 9, e1003348. [Google Scholar] [CrossRef]
- Dudbridge, F. Polygenic epidemiology. Genet. Epidemiol. 2016, 40, 268–272. [Google Scholar] [CrossRef] [Green Version]
- Chasioti, D.; Yan, J.; Nho, K.; Saykin, A.J. Progress in polygenic composite scores in alzheimer’s and other complex diseases. Trends Genet. 2019, 35, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Patel, K.P.; Teng, A.K.; Berens, A.J.; Lachance, J. Genetic disease risks can be misestimated across global populations. Genome Biol. 2018, 19, 179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De La Vega, F.M.; Bustamante, C.D. Polygenic risk scores: A biased prediction? Genome Med. 2018, 10, 100. [Google Scholar] [CrossRef] [PubMed]
- Vuillermin, P.; Saffery, R.; Allen, K.J.; Carlin, J.B.; Tang, M.L.; Ranganathan, S.; Burgner, D.; Dwyer, T.; Collier, F.; Jachno, K. Cohort profile: The barwon infant study. Int. J. Epidemiol. 2015, 44, 1148–1160. [Google Scholar] [CrossRef] [PubMed]
- Heffernan, A.; Gomez-Ramos, M.; Symeonides, C.; Hare, D.; Vijayasarathy, S.; Thompson, K.; Mueller, J.; Ponsonby, A.; Sly, P.; BIS Investigator Group. Harmonizing analytical chemistry and clinical epidemiology for human biomonitoring studies. A case-study of plastic product chemicals in urine. Chemosphere 2020, 238, 124631. [Google Scholar] [CrossRef]
- Adibi, J.J.; Whyatt, R.M.; Williams, P.L.; Calafat, A.M.; Camann, D.; Herrick, R.; Nelson, H.; Bhat, H.K.; Perera, F.P.; Silva, M.J. Characterization of phthalate exposure among pregnant women assessed by repeat air and urine samples. Environ. Health Perspect. 2008, 116, 467–473. [Google Scholar] [CrossRef]
- Suzuki, Y.; Niwa, M.; Yoshinaga, J.; Watanabe, C.; Mizumoto, Y.; Serizawa, S.; Shiraishi, H. Exposure assessment of phthalate esters in japanese pregnant women by using urinary metabolite analysis. Environ. Health Prev. Med. 2009, 14, 180–187. [Google Scholar] [CrossRef] [Green Version]
- Achenbach, T.M.; Rescorla, L. Achenbach System of Empirically Based Assessment; Mental Measurements Yearbook; EBSCOhost: Ipswich, MA, USA, 2007. [Google Scholar]
- Achenbach, T.M.; Rescorla, L. Manual for ASEBA Preschool Forms and Profiles; University of Vermont, Research Centre for Children, Youth and Families: Burlington, VT, USA, 2000. [Google Scholar]
- Bayley, N. Bayley Scales of Infant and Toddler Development: Bayley-III; Harcourt Assessment; Psych. Corporation: San Antonio, TX, USA, 2006. [Google Scholar]
- McCarthy, S.; Das, S.; Kretzschmar, W.; Delaneau, O.; Wood, A.R.; Teumer, A.; Kang, H.M.; Fuchsberger, C.; Danecek, P.; Sharp, K.; et al. A reference panel of 64,976 haplotypes for genotype imputation. Nat. Genet. 2016, 48, 1279–1283. [Google Scholar]
- The Haplotype Reference Consortium. Available online: http://www.haplotype-reference-consortium.org/ (accessed on 24 March 2022).
- Slenter, D.N.; Kutmon, M.; Hanspers, K.; Riutta, A.; Windsor, J.; Nunes, N.; Mélius, J.; Cirillo, E.; Coort, S.L.; Digles, D.; et al. Wikipathways: A multifaceted pathway database bridging metabolomics to other omics research. Nucleic Acids Res. 2018, 46, D661–D667. [Google Scholar] [CrossRef]
- Martens, M.; Ammar, A.; Riutta, A.; Waagmeester, A.; Slenter, D.N.; Hanspers, K.; Miller, R.A.; Digles, D.; Lopes, E.N.; Ehrhart, F.; et al. Wikipathways: Connecting communities. Nucleic Acids Res. 2020, 49, D613–D621. [Google Scholar] [CrossRef]
- Morel, Y.; Barouki, R. Repression of gene expression by oxidative stress. Biochem. J. 1999, 342, 481–496. [Google Scholar] [CrossRef] [PubMed]
- Wikipathways. Available online: https://www.wikipathways.org/index.php/WikiPathways (accessed on 14 March 2022).
- Lonsdale, J.; Thomas, J.; Salvatore, M.; Phillips, R.; Lo, E.; Shad, S.; Hasz, R.; Walters, G.; Garcia, F.; Young, N.; et al. The genotype-tissue expression (gtex) project. Nat. Genet. 2013, 45, 580–585. [Google Scholar] [CrossRef] [PubMed]
- Gtex Portal. Available online: https://gtexportal.org/home/ (accessed on 26 March 2022).
- VanderWeele, T.J. Reconsidering the denominator of the attributable proportion for interaction. Eur. J. Epidemiol. 2013, 28, 779–784. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; VanderWeele, T.J. Additive interaction in the presence of a mismeasured outcome. Am. J. Epidemiol. 2015, 181, 81–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pham, C.; Symeonides, C.; O’Hely, M.; Sly, P.D.; Knibbs, L.D.; Thomson, S.; Vuillermin, P.; Saffery, R.; Ponsonby, A.L. Early life environmental factors associated with autism spectrum disorder symptoms in children at age 2 years: A birth cohort study. Autism Int. J. Res. Pract. 2022; advance online publication. [Google Scholar] [CrossRef]
- Linseman, D.A. Targeting oxidative stress for neuroprotection. Antioxid. Redox Signal. 2009, 11, 421–424. [Google Scholar] [CrossRef]
- Newton, D.F.; Naiberg, M.R.; Goldstein, B.I. Oxidative stress and cognition amongst adults without dementia or stroke: Implications for mechanistic and therapeutic research in psychiatric disorders. Psychiatry Res. 2015, 227, 127–134. [Google Scholar] [CrossRef]
- Carter, C.J.; Blizard, R.A. Autism genes are selectively targeted by environmental pollutants including pesticides, heavy metals, bisphenol a, phthalates and many others in food, cosmetics or household products. Neurochem. Int. 2016, 101, 83–109. [Google Scholar] [CrossRef]
- Mandic-Maravic, V.; Pljesa-Ercegovac, M.; Mitkovic-Voncina, M.; Savic-Radojevic, A.; Lecic-Tosevski, D.; Simic, T.; Pejovic-Milovancevic, M. Impaired redox control in autism spectrum disorders: Could it be the x in gxe? Curr. Psychiatry Rep. 2017, 19, 52. [Google Scholar] [CrossRef]
- Ferguson, K.; McElrath, T.; Chen, Y.; Mukherjee, B.; Meeker, J. Urinary phthalate metabolites and biomarkers of oxidative stress in pregnant women: A repeated measures analysis. Environ. Health Perspect. 2015, 123, 210–216. [Google Scholar] [CrossRef] [Green Version]
- Thomson, S.; Drummond, K.; O’Hely, M.; Symeonides, C.; Chandran, C.; Mansell, T.; Sly, P.; Ponsonby, A.-L.; BIS Investigator Group. Increased maternal non-oxidative energy metabolism mediates association between prenatal DEHP exposure and offspring ASD symptoms: A birth cohort study. Environ. Health Perspect. 2022; under review. [Google Scholar]
- Barnham, K.J.; Masters, C.L.; Bush, A.I. Neurodegenerative diseases and oxidative stress. Nat. Rev. Drug Discov. 2004, 3, 205–214. [Google Scholar] [CrossRef]
- Valavanidis, A.; Vlachogianni, T.; Fiotakis, C. 8-hydroxy-2′-deoxyguanosine (8-ohdg): A critical biomarker of oxidative stress and carcinogenesis. J. Environ. Sci. Health C Environ. Carcinog. Ecotoxicol. Rev. 2009, 27, 120–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graille, M.; Wild, P.; Sauvain, J.; Hemmendinger, M.; Canu, I.G.; Hopf, N. Urinary 8-isoprostane as a biomarker for oxidative stress. A systematic review and meta-analysis. Toxicol. Lett. 2020, 328, 19–27. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Gene | eQTL SNP | Effect of SNP on Gene Expression | Inferred Effect of SNP on Oxidative Stress | |
|---|---|---|---|---|
| Pro-oxidant genes | XDH | rs45498201 | increase | pro-oxidant |
| NOX4 | rs10830296 | reduction | antioxidant | |
| NFIX | rs149677133 | reduction | antioxidant | |
| CYP1A1 | rs2470890 | increase | pro-oxidant | |
| Antioxidant genes | MAPK10 | rs80320648 | reduce | pro-oxidant |
| NFKB1 | rs28573147 | increase | antioxidant | |
| SP1 | rs35437931 | increase | antioxidant | |
| FOS | rs79713290 | increase | antioxidant | |
| CAT | rs12793666 | increase | antioxidant | |
| GPX1 | rs17650792 | reduction | pro-oxidant | |
| SOD1 | rs4998557 | increase | antioxidant | |
| SOD2 | rs5746105 | increase | antioxidant |
| Full Cohort (N = 1074) | Participants with Any Neurodevelopment Data (N = 868) | |||||
|---|---|---|---|---|---|---|
| N | Mean (SD) or GM {GSD} or % [n] | N | Mean (SD) or GM {GSD} or % [n] | |||
| Genotype | ||||||
| Oxidative stress genetic score | 1031 | 6.8 | (1.0) | 850 | 6.8 | (1.0) |
| Phthalate daily intakes (µg/kg bw/day) | ||||||
| DEP daily intake | 847 | 1.6 | {3.8} | 756 | 1.6 | {3.8} |
| DBPs daily intake | 847 | 1.9 | {2.0} | 756 | 1.9 | {2.0} |
| DEHP (oxidized) daily intake | 847 | 1.6 | {2.1} | 756 | 1.6 | {2.1} |
| Σ phthalate daily intake | 847 | 6.3 | {2.2} | 756 | 6.3 | {2.2} |
| DEP daily intake top quintile (≥4.459) | 847 | 20.2% | [171] | 756 | 20.8% | [157] |
| DBPs daily intake top quintile (≥3.362) | 847 | 20.1% | [170] | 756 | 20.5% | [155] |
| DEHP (oxidized) daily intake top quintile (≥2.614) | 847 | 20.1% | [170] | 756 | 19.8% | [150] |
| Σ phthalate daily intake top quintile (≥11.043) | 847 | 20.1% | [170] | 756 | 20.4% | [154] |
| Gestational age at maternal urine collection (weeks) | 847 | 36.3 | (0.7) | 756 | 36.3 | (0.7) |
| Demographic and household factors | ||||||
| Maternal age at conception (years) | 1074 | 31.3 | (4.8) | 868 | 31.8 | (4.5) |
| Paternal age at conception (years) | 1024 | 33.5 | (5.9) | 830 | 33.8 | (5.6) |
| British/European ancestry (all 4 grandparents) | 1060 | 73.0% | [774] | 861 | 73.8% | [635] |
| Maternal university-level education | 1068 | 51.3% | [548] | 865 | 55.8% | [483] |
| Parental marital status (married) | 1071 | 70.4% | [754] | 868 | 74.1% | [643] |
| Older siblings of child living at home (one or more) | 1072 | 55.0% | [590] | 865 | 55.2% | [478] |
| Prenatal, perinatal, and postnatal factors | ||||||
| Gestational age at birth (weeks) | 1074 | 39.4 | (1.5) | 868 | 39.4 | (1.5) |
| Child sex at birth (male) | 1074 | 51.7% | [555] | 868 | 52.6% | [457] |
| Child neurodevelopment | ||||||
| Bayley-III Cognitive Scale raw score | 678 | 71.1 | (4.1) | |||
| Bayley-III Cognitive Scale scaled score | 678 | 10.8 | (2.1) | |||
| Bayley-III Cognitive Scale raw score <70 | 678 | 34.7% | [235] | |||
| Child age at Bayley-III assessment (months) | 678 | 29.4 | (1.7) | |||
| CBCL autism spectrum problems (T-score above 50) | 676 | 36.8% | [249] | |||
| CBCL attention-deficit hyperactivity problems (T-score above 65) | 676 | 5.2% | [35] | |||
| Child age at CBCL assessment (months) | 677 | 29.5 | (1.8) | |||
| Autism spectrum disorder doctor diagnosis | 791 | 1.4% | [11] | |||
| Parent-reported autistic traits | 791 | 4.9% | [39] | |||
| Child age at 4-year review (months) | 791 | 49.9 | (3.1) | |||
| Bayley-III Cognition | Above-Median CBCL Autism Spectrum Problems | ASD Diagnosis | ASD Traits | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Adj. Mean Difference (95% CI) | p Value | YES: Mean (SD) or % [n] | NO: Mean (SD) or % [n] | AOR (95% CI) | p Value | YES: Mean (SD) or % [n] | NO: Mean (SD) or % [n] | AOR (95% CI) | p Value | YES: Mean (SD) or % [n] | NO: Mean (SD) or % [n] | AOR (95% CI) | p Value | |
| gPFSox (Per 2 additional pro-oxidant alleles) | −0.20 (−0.50, 0.10) | 0.197 | 6.8 (1.0) | 6.7 (0.9) | 1.10 (0.94, 1.30) | 0.250 | 7.5 (1.2) | 6.8 (1.0) | 2.10 (1.12, 4.11) | 0.024 | 7.1 (0.9) | 6.8 (1.0) | 1.42 (1.02, 2.01) | 0.041 |
| gPFSox (Top quintile vs. rest) | −0.49 (−1.31, 0.29) | 0.231 | 17.0% [41] | 14.8% [62] | 1.11 (0.68, 1.71) | 0.637 | 36.4% [4] | 17.7% [135] | 2.56 (0.74, 9.03) | 0.140 | 21.1% [8] | 17.8% [131] | 1.20 (0.54, 2.67) | 0.654 |
| Bayley-III Cognition | Above-Median CBCL Autism Spectrum Problems | ASD Diagnosis | ASD Traits | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Gene-Phthalate Subgroup | N | Mean (SD) | % [n] 1 | N | % [n] 2 | N | % [n] 3 | N | % [n] 4 | |
| DEP | GloPlo | 396 | 71.4 (3.9) | 32.6 [129] | 391 | 36.1 [141] | 438 | 1.4 [6] | 438 | 5.0 [22] |
| GloPhi | 110 | 71.2 (4.2) | 31.8 [35] | 111 | 36.0 [40] | 120 | 0.8 [1] | 120 | 5.0 [6] | |
| GhiPlo | 82 | 71.6 (3.9) | 29.3 [24] | 82 | 39.0 [32] | 100 | 2.0 [2] | 100 | 6.0 [6] | |
| GhiPhi | 18 | 67.6 (7.6) | 72.2 [13] | 13 | 46.2 [6] | 19 | 0.0 [0] | 19 | 0.0 [0] | |
| DBPs | GloPlo | 403 | 71.2 (4.1) | 34.0 [137] | 398 | 34.2 [136] | 444 | 1.4 [6] | 444 | 5.0 [22] |
| GloPhi | 103 | 71.7 (3.3) | 26.2 [27] | 104 | 43.3 [45] | 114 | 0.9 [1] | 114 | 5.3 [6] | |
| GhiPlo | 74 | 71.5 (4) | 32.4 [24] | 73 | 34.2 [25] | 91 | 1.1 [1] | 91 | 3.3 [3] | |
| GhiPhi | 26 | 69.3 (7) | 50.0 [13] | 22 | 59.1 [13] | 28 | 3.6 [1] | 28 | 10.7 [3] | |
| DEHP | GloPlo | 408 | 71.1 (3.9) | 34.6 [141] | 397 | 35.5 [141] | 447 | 1.3 [6] | 447 | 5.4 [24] |
| GloPhi | 98 | 72.1 (3.8) | 23.5 [23] | 105 | 38.1 [40] | 111 | 0.9 [1] | 111 | 3.6 [4] | |
| GhiPlo | 81 | 71.3 (4.1) | 35.8 [29] | 75 | 40.0 [30] | 91 | 0.0 [0] | 91 | 2.2 [2] | |
| GhiPhi | 19 | 69.3 (7.7) | 42.1 [8] | 20 | 40.0 [8] | 28 | 7.1 [2] | 28 | 14.3 [4] | |
| Σ phthalates | GloPlo | 401 | 71.3 (3.9) | 33.7 [135] | 397 | 36.0 [143] | 440 | 1.1 [5] | 440 | 4.3 [19] |
| GloPhi | 105 | 71.6 (4) | 27.6 [29] | 105 | 36.2 [38] | 118 | 1.7 [2] | 118 | 7.6 [9] | |
| GhiPlo | 83 | 71.5 (4.1) | 32.5 [27] | 82 | 39.0 [32] | 97 | 0.0 [0] | 97 | 3.1 [3] | |
| GhiPhi | 17 | 68.2 (7.7) | 58.8 [10] | 13 | 46.2 [6] | 22 | 9.1 [2] | 22 | 13.6 [3] | |
| G | P | Glo Phi | Ghi Plo | Ghi Phi | Additive Interaction | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| gPFSox | Phthalate | |||||||||
| Adj Mean Difference (95% CI) | p Value | Adj Mean Difference (95% CI) | p Value | Adj Mean Difference (95% CI) | p Value | AP (95% CI) | p Value | |||
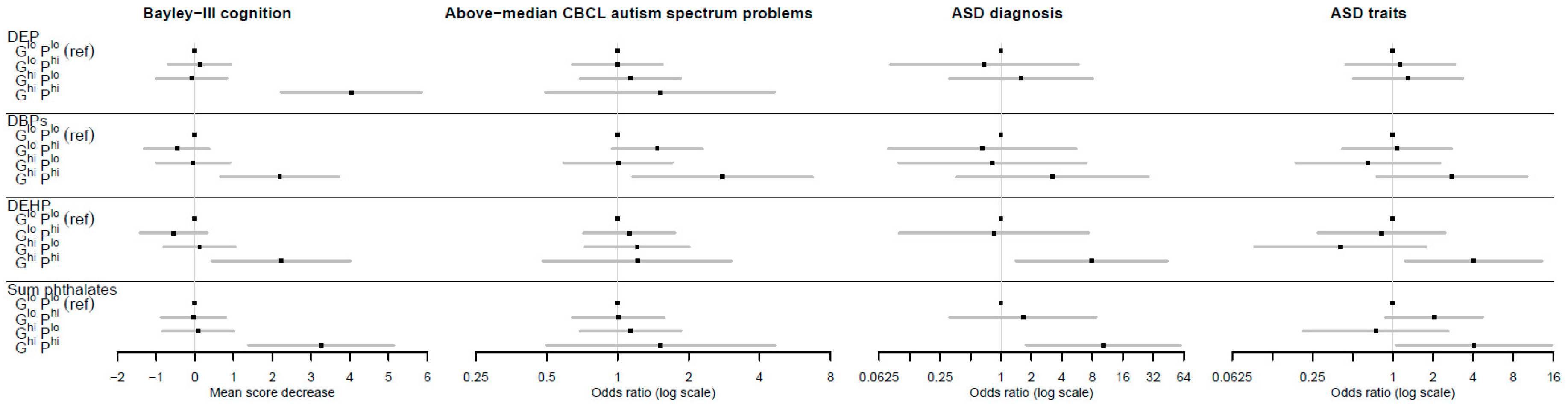
| Bayley-III cognition a | DEP | −0.12 (−0.93, 0.69) | 0.766 | 0.09 (−0.83, 1.00) | 0.852 | −4.04 (−5.85, −2.22) | <0.0001 | 0.82 (0.62, 1.01) | <0.0001 | |
| DBPs | 0.46 (−0.37, 1.30) | 0.276 | 0.04 (−0.91, 1.00) | 0.927 | −2.19 (−3.72, −0.66) | 0.005 | 0.71 (0.43, 1.03) | <0.0001 | ||
| DEHP | 0.55 (−0.32, 1.42) | 0.212 | −0.12 (−1.04, 0.80) | 0.796 | −2.23 (−4.01, −0.45) | 0.014 | 0.30 (−0.62, 1.16) | 0.511 | ||
| Σ phthalates | 0.04 (−0.79, 0.88) | 0.922 | −0.08 (−1.00, 0.83) | 0.856 | −3.26 (−5.13, −1.39) | 0.001 | 0.72 (0.39, 1.10) | <0.0001 | ||
| AOR (95% CI) | p value | AOR (95% CI) | p value | AOR (95% CI) | p value | |||||
| Above-median CBCL autism spectrum problems | DEP | 1.00 (0.64, 1.55) | 0.993 | 1.13 (0.69, 1.85) | 0.615 | 1.51 (0.50, 4.62) | 0.468 | 0.04 (−1.21, 1.34) | 0.952 | |
| DBPs | 1.47 (0.95, 2.28) | 0.086 | 1.00 (0.59, 1.70) | 0.986 | 2.78 (1.16, 6.69) | 0.022 | 0.43 (−0.17, 1.03) | 0.146 | ||
| DEHP | 1.12 (0.72, 1.75) | 0.625 | 1.21 (0.73, 2.01) | 0.465 | 1.21 (0.48, 3.04) | 0.682 | −0.10 (−1.28, 1.14) | 0.874 | ||
| Σ phthalates | 1.01 (0.64, 1.58) | 0.977 | 1.13 (0.70, 1.85) | 0.613 | 1.52 (0.50, 4.63) | 0.46 | 0.28 (−0.61, 1.24) | 0.554 | ||
| ASD diagnosis | DEP | 0.69 (0.08, 5.83) | 0.729 | 1.58 (0.31, 8.02) | 0.583 | --- | −0.26 (−3.48, 2.91) | 0.872 | ||
| DBPs | 0.65 (0.08, 5.49) | 0.693 | 0.82 (0.10, 6.93) | 0.854 | 3.23 (0.37, 28.61) | 0.291 | 0.86 (0.08, 1.62) | 0.025 | ||
| DEHP | 0.85 (0.88, 0.10) | 0.884 | --- | 7.84 (1.40, 43.98) | 0.019 | 0.89 (0.62, 1.16) | <0.0001 | |||
| Σ phthalates | 1.65 (0.31, 8.70) | 0.554 | --- | 10.24 (1.76, 59.48) | 0.01 | 0.84 (0.51, 1.17) | <0.0001 | |||
| ASD traits | DEP | 1.14 (0.44, 2.90) | 0.791 | 1.30 (0.51, 3.36) | 0.582 | --- | −0.44 (−0.24, 1.36) | 0.632 | ||
| DBPs | 1.07 (0.42, 2.75) | 0.881 | 0.65 (0.19, 2.26) | 0.502 | 2.77 (0.71, 10.11) | 0.122 | 0.74 (0.21, 1.25) | 0.009 | ||
| DEHP | 0.82 (0.27, 2.46) | 0.724 | 0.40 (0.09, 1.76) | 0.226 | 4.01 (1.23, 13.01) | 0.021 | 0.94 (0.66, 1.22) | <0.0001 | ||
| Σ phthalates | 2.04 (0.88, 4.70) | 0.095 | 0.75 (0.21, 2.60) | 0.645 | 4.06 (1.06, 15.61) | 0.041 | 0.56 (−0.13, 1.26) | 0.114 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tanner, S.; Thomson, S.; Drummond, K.; O’Hely, M.; Symeonides, C.; Mansell, T.; Saffery, R.; Sly, P.D.; Collier, F.; Burgner, D.; et al. A Pathway-Based Genetic Score for Oxidative Stress: An Indicator of Host Vulnerability to Phthalate-Associated Adverse Neurodevelopment. Antioxidants 2022, 11, 659. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11040659
Tanner S, Thomson S, Drummond K, O’Hely M, Symeonides C, Mansell T, Saffery R, Sly PD, Collier F, Burgner D, et al. A Pathway-Based Genetic Score for Oxidative Stress: An Indicator of Host Vulnerability to Phthalate-Associated Adverse Neurodevelopment. Antioxidants. 2022; 11(4):659. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11040659
Chicago/Turabian StyleTanner, Samuel, Sarah Thomson, Katherine Drummond, Martin O’Hely, Christos Symeonides, Toby Mansell, Richard Saffery, Peter D. Sly, Fiona Collier, David Burgner, and et al. 2022. "A Pathway-Based Genetic Score for Oxidative Stress: An Indicator of Host Vulnerability to Phthalate-Associated Adverse Neurodevelopment" Antioxidants 11, no. 4: 659. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11040659