
Identification of Crucial Polymethoxyflavones Tangeretin and 3,5,6,7,8,3′,4′-Heptamethoxyflavone and Evaluation of Their Contribution to Anticancer Effects of Pericarpium Citri Reticulatae ‘Chachi’ during Storage
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples and Reagents
2.2. Cell Culture
2.3. MTT Assay
2.4. Cell Cycle Analysis
2.5. Cell Apoptosis Analysis
2.6. Detection of Intracellular ROS Accumulation
2.7. Data Processing and Statistical Analysis
3. Results and Discussion
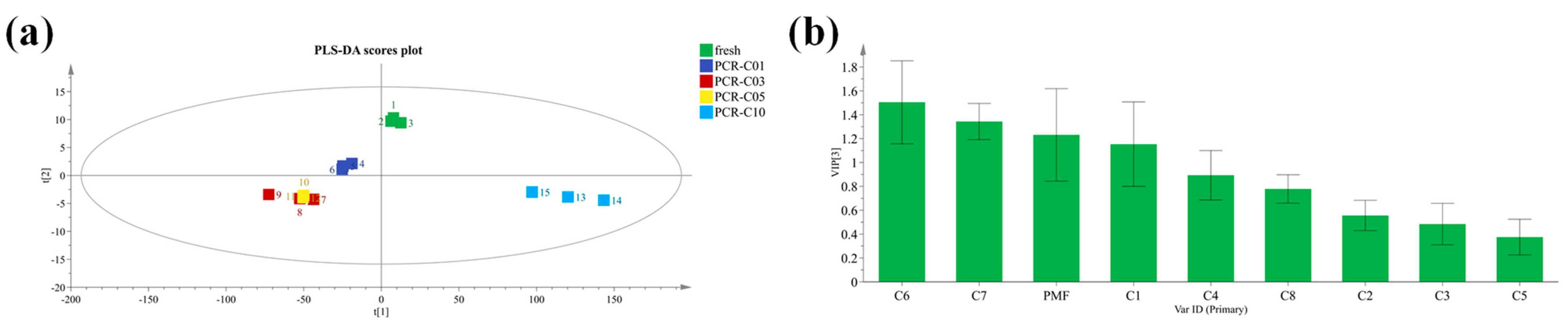
3.1. Multivariate Statistical Analysis of Bound and Free PMFs during PCR-C Storage
3.2. Screening of Important PMFs in PCR-C during Storage
3.3. PCR-CF Inhibits Cell Growth of A549
3.4. PCR-CF Induces Cell Cycle Arrest in A549 Cells
3.5. PCR-CF Induces Apoptosis in A549 Cells
3.6. PCR-CF Induces ROS Accumulation in A549 Cells
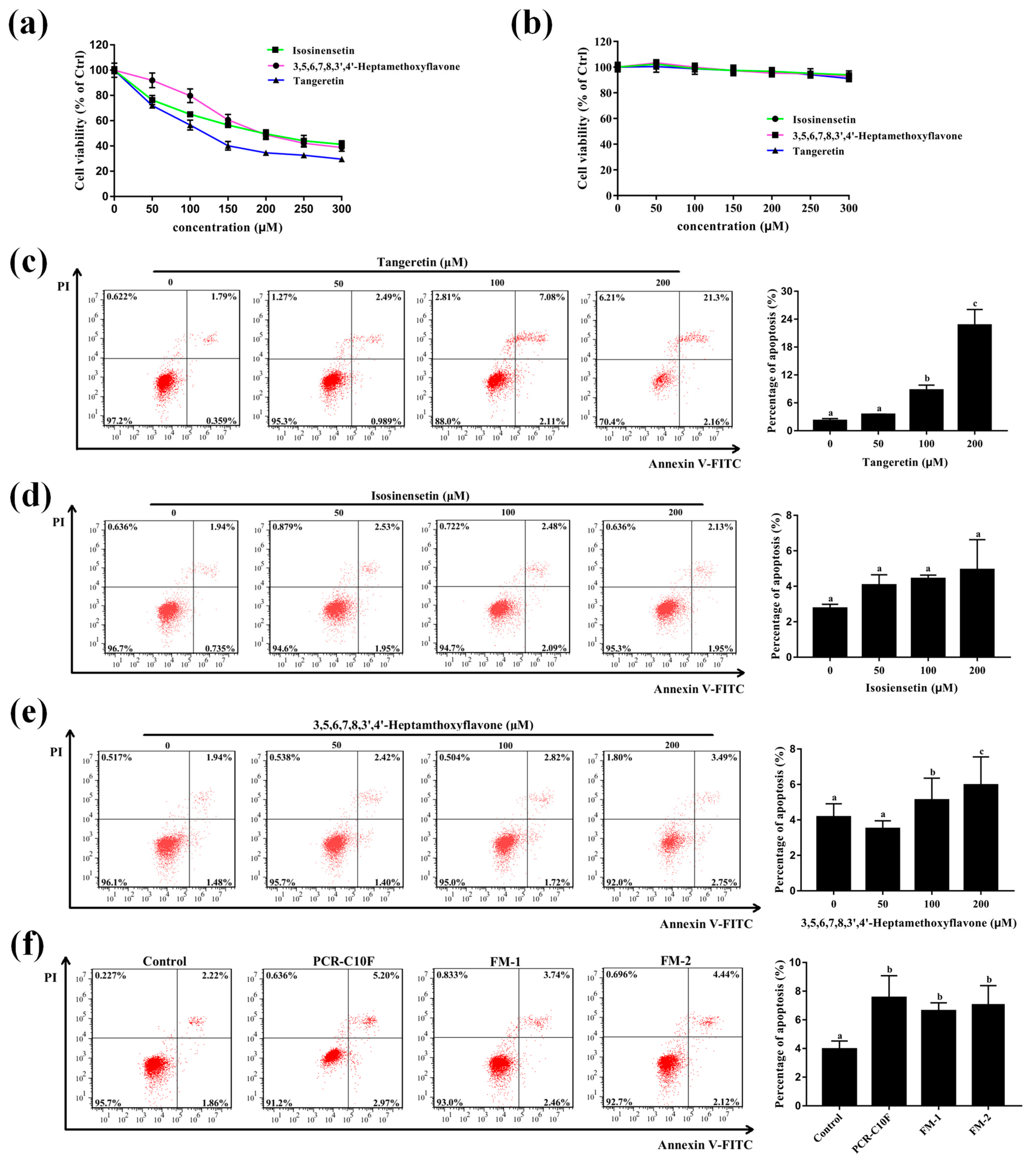
3.7. Important PMFs Inhibit the Cell Growth and Induce Apoptosis of A549 Cells
3.8. Crucial Active PMFs Induces ROS Accumulation in A549
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, H.; Qiu, N.; Ding, H.; Yao, R. Polyphenols contents and antioxidant capacity of 68 Chinese herbals suitable for medical or food uses. Food Res. Int. 2008, 41, 363–370. [Google Scholar] [CrossRef]
- Wei, D.; Ci, X.; Chu, X.; Wei, M.; Hua, S.; Deng, X. Hesperidin Suppresses Ovalbumin-Induced Airway Inflammation in a Mouse Allergic Asthma Model. Inflammation 2012, 35, 114–121. [Google Scholar] [CrossRef]
- Tang, X.; Zhao, H.; Jiang, W.; Zhang, S.; Guo, S.; Gao, X.; Yang, P.; Shi, L.; Liu, L. Pharmacokinetics and pharmacodynamics of citrus peel extract in lipopolysaccharide-induced acute lung injury combined with Pinelliae Rhizoma Praeparatum. Food Funct. 2018, 9, 5880–5890. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Huang, Q.; Zhang, Y.; Su, J.; Liu, J. Current Situation of the Long-stored TCM Research and Exploration of the Ideas. China Pharm. 2012, 23, 1433–1435. [Google Scholar]
- Fu, M.; Xu, Y.; Chen, Y.; Wu, J.; Yu, Y.; Zou, B.; An, K.; Xiao, G. Evaluation of bioactive flavonoids and antioxidant activity in Pericarpium Citri Reticulatae (Citrus reticulata ‘Chachi’) during storage. Food Chem. 2017, 230, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Luo, H.; Hu, P.; Yang, Y.; Wu, B.; Zheng, G. Evaluation of chemical components in Citri Reticulatae Pericarpium of different cultivars collected from different regions by GC-MS and HPLC. Food Sci. Nutr. 2018, 6, 400–416. [Google Scholar] [CrossRef] [PubMed]
- Youn, K.; Lee, S.; Jun, M. Discovery of Nobiletin from Citrus Peel as a Potent Inhibitor of beta-Amyloid Peptide Toxicity. Nutrients 2019, 11, 2684. [Google Scholar] [CrossRef]
- Yao, X.; Zhu, X.; Pan, S.; Fang, Y.; Jiang, F.; Phillips, G.O.; Xu, X. Antimicrobial activity of nobiletin and tangeretin against Pseudomonas. Food Chem. 2012, 132, 1883–1890. [Google Scholar] [CrossRef]
- Bian, X.; Xie, X.; Cai, J.; Zhao, Y.; Miao, W.; Chen, X.; Xiao, Y.; Li, N.; Wu, J.-L. Dynamic changes of phenolic acids and antioxidant activity of Citri Reticulatae Pericarpium during aging processes. Food Chem. 2022, 373, 131399. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Zeng, W.; Huang, K.-E.; Li, D.-X.; Chen, W.; Yu, X.-Q.; Ke, X.-H. Discrimination of Citrus reticulata Blanco and Citrus reticulata ‘Chachi’ as well as the Citrus reticulata ‘Chachi’ within different storage years using ultra high performance liquid chromatography quadrupole/time-of-flight mass spectrometry based metabolomics approach. J. Pharm. Biomed. Anal. 2019, 171, 218–231. [Google Scholar] [CrossRef]
- Duma, N.; Santana-Davila, R.; Molina, J.R. Non-Small Cell Lung Cancer: Epidemiology, Screening, Diagnosis, and Treatment. Mayo Clin. Proc. 2019, 94, 1623–1640. [Google Scholar] [CrossRef] [PubMed]
- Zarogoulidis, K.; Zarogoulidis, P.; Darwiche, K.; Boutsikou, E.; Machairiotis, N.; Tsakiridis, K.; Katsikogiannis, N.; Kougioumtzi, I.; Karapantzos, I.; Huang, H.; et al. Treatment of non-small cell lung cancer (NSCLC). J. Thorac. Dis. 2013, 5, S389–S396. [Google Scholar] [CrossRef] [PubMed]
- Lemjabbar-Alaoui, H.; Hassan, O.U.; Yang, Y.-W.; Buchanan, P. Lung cancer: Biology and treatment options. Biochim. Et Biophys. Acta-Rev. Cancer 2015, 1856, 189–210. [Google Scholar] [CrossRef]
- Duan, L.; Dou, L.-L.; Yu, K.-Y.; Guo, L.; Chen, B.-Z.; Li, P.; Liu, E.H. Polymethoxyflavones in peel of Citrus reticulata ‘Chachi’ and their biological activities. Food Chem. 2017, 234, 254–261. [Google Scholar] [CrossRef]
- Fu, M.; Zou, B.; An, K.; Yu, Y.; Tang, D.; Wu, J.; Xu, Y.; Ti, H. Anti- asthmatic activity of alkaloid compounds from Pericarpium Citri Reticulatae (Citrus reticulata ‘Chachi’). Food Funct. 2019, 10, 903–911. [Google Scholar] [CrossRef]
- Chen, X.-M.; Tait, A.R.; Kitts, D.D. Flavonoid composition of orange peel and its association with antioxidant and anti-inflammatory activities. Food Chem. 2017, 218, 15–21. [Google Scholar] [CrossRef]
- Yu, Q.; Tao, Y.; Huang, Y.; Zogona, D.; Wu, T.; Liu, R.; Pan, S.; Xu, X. Aged Pericarpium Citri Reticulatae ‘Chachi’ Attenuates Oxidative Damage Induced by tert-Butyl Hydroperoxide (t-BHP) in HepG2 Cells. Foods 2022, 11, 273. [Google Scholar] [CrossRef]
- Liang, S.; Wen, Z.; Tang, T.; Liu, Y.; Dang, F.; Xie, T.; Wu, H. Study on flavonoid and bioactivity features of the pericarp of Citri Reticulatae ‘chachi’ during storage. Arab. J. Chem. 2022, 15, 103653. [Google Scholar] [CrossRef]
- Choi, M.-Y.; Chai, C.; Park, J.H.; Lim, J.; Lee, J.; Kwon, S.W. Effects of storage period and heat treatment on phenolic compound composition in dried Citrus peels (Chenpi) and discrimination of Chenpi with different storage periods through targeted metabolomic study using HPLC-DAD analysis. J. Pharm. Biomed. Anal. 2011, 54, 638–645. [Google Scholar] [CrossRef]
- Wang, H.; Chen, G.; Fu, X.; Liu, R.-H. Effects of aging on the phytochemical profile and antioxidative activity of Pericarpium Citri Reticulatae ‘Chachiensis’. Rsc Adv. 2016, 6, 105272–105281. [Google Scholar] [CrossRef]
- Barker, M.; Rayens, W. Partial least squares for discrimination. J. Chemom. 2003, 17, 166–173. [Google Scholar] [CrossRef]
- Zheng, Y.; Zeng, X.; Peng, W.; Wu, Z.; Su, W. Study on the Discrimination between Citri Reticulatae Pericarpium Varieties Based on HS-SPME-GC-MS Combined with Multivariate Statistical Analyses. Molecules 2018, 23, 1235. [Google Scholar] [CrossRef]
- Lloyd, G.R.; Stone, N. Method for Identification of Spectral Targets in Discrete Frequency Infrared Spectroscopy for Clinical Diagnostics. Appl. Spectrosc. 2015, 69, 1066–1073. [Google Scholar] [CrossRef]
- Yang, M.; Jiang, Z.; Wen, M.; Wu, Z.; Zha, M.; Xu, W.; Zhang, L. Chemical Variation of Chenpi (Citrus Peels) and Corresponding Correlated Bioactive Compounds by LC-MS Metabolomics and Multibioassay Analysis. Front. Nutr. 2022, 9, 825381. [Google Scholar] [CrossRef]
- Liu, X.; Jiang, Q.; Liu, H.; Luo, S. Vitexin induces apoptosis through mitochondrial pathway and PI3K/Akt/mTOR signaling in human non-small cell lung cancer A549 cells. Biol. Res. 2019, 52, 7. [Google Scholar] [CrossRef]
- Ren, H.; Zhang, Y.-Y.; Li, Y.-L.; Bai, M.; Yan, Q.-L.; Huang, X.-X.; Cui, W.; Zhao, H.; Gu, L.; Liu, Q.; et al. Semisynthesis and Non-Small-Cell Lung Cancer Cytotoxicity Evaluation of Germacrane-Type Sesquiterpene Lactones from Elephantopus scaber. J. Nat. Prod. 2022, 85, 352–364. [Google Scholar] [CrossRef]
- Wang, J.; Li, X. Chamaejasmine induces apoptosis in human non-small-cell lung cancer A549 cells through increasing the Bax/Bcl-2 ratio, caspase-3 and activating the Fas/FasL. Minerva Med. 2021, 112, 413–415. [Google Scholar] [CrossRef]
- Song, M.; Charoensinphon, N.; Wu, X.; Zheng, J.; Gao, Z.; Xu, F.; Wang, M.; Xiao, H. Inhibitory Effects of Metabolites of 5-Demethylnobiletin on Human Nonsmall Cell Lung Cancer Cells. J. Agric. Food Chem. 2016, 64, 4943–4949. [Google Scholar] [CrossRef]
- Park, K.I.; Park, H.S.; Nagappan, A.; Hong, G.E.; Lee, D.H.; Kang, S.R.; Kim, J.A.; Zhang, J.; Kim, E.H.; Lee, W.S.; et al. Induction of the cell cycle arrest and apoptosis by flavonoids isolated from Korean Citrus aurantium L. in non-small-cell lung cancer cells. Food Chem. 2012, 135, 2728–2735. [Google Scholar] [CrossRef]
- Park, K.-I.; Park, H.-S.; Kim, M.-K.; Hong, G.-E.; Nagappan, A.; Lee, H.-J.; Yumnam, S.; Lee, W.-S.; Won, C.-K.; Shin, S.-C.; et al. Flavonoids identified from Korean Citrus aurantium L. inhibit Non-Small Cell Lung Cancer growth in vivo and in vitro. J. Funct. Foods 2014, 7, 287–297. [Google Scholar] [CrossRef]
- Ma, L.-L.; Wang, D.-w.; Yu, X.-D.; Zhou, Y.-L. Tangeretin induces cell cycle arrest and apoptosis through upregulation of PTEN expression in glioma cells. Biomed. Pharmacother. 2016, 81, 491–496. [Google Scholar] [CrossRef] [PubMed]
- Chin-Chen, C.; Shih-Ying, C.; Charng-Cherng, C.; Pin-Der, D. Antiproliferative effect of sweet orange peel and its bioactive compounds against human hepatoma cells, in vitro and in vivo. J. Funct. Foods 2017, 33, 363–375. [Google Scholar] [CrossRef]
- Woelfle, U.; Seelinger, G.; Bauer, G.; Meinke, M.C.; Lademann, J.; Schempp, C.M. Reactive Molecule Species and Antioxidative Mechanisms in Normal Skin and Skin Aging. Ski. Pharmacol. Physiol. 2014, 27, 316–332. [Google Scholar] [CrossRef] [PubMed]
- Menon, S.; Devi, S.K.S.; Santhiya, R.; Rajeshkumar, S.; Kumar, V.S. Selenium nanoparticles: A potent chemotherapeutic agent and an elucidation of its mechanism. Colloids Surf. B-Biointerfaces 2018, 170, 280–292. [Google Scholar] [CrossRef]
- Bossis, G.; Sarry, J.-E.; Kifagi, C.; Ristic, M.; Saland, E.; Vergez, F.; Salem, T.; Boutzen, H.; Baik, H.; Brockly, F.; et al. The ROS/SUMO Axis Contributes to the Response of Acute Myeloid Leukemia Cells to Chemotherapeutic Drugs. Cell Rep. 2014, 7, 1815–1823. [Google Scholar] [CrossRef]
- Fouzder, C.; Mukhuty, A.; Kundu, R. Kaempferol inhibits Nrf2 signalling pathway via downregulation of Nrf2 mRNA and induces apoptosis in NSCLC cells. Arch. Biochem. Biophys. 2021, 697, 108700. [Google Scholar] [CrossRef]
- Liang, F.; Fang, Y.; Cao, W.; Zhang, Z.; Pan, S.; Xu, X. Attenuation of tert-Butyl Hydroperoxide (t-BHP)-Induced Oxidative Damage in HepG2 Cells by Tangeretin: Relevance of the Nrf2-ARE and MAPK Signaling Pathways. J. Agric. Food Chem. 2018, 66, 6317–6325. [Google Scholar] [CrossRef]
- Kim, H.-I.; Jeong, Y.-U.; Kim, J.-H.; Park, Y.-J. 3,5,6,7,8,3,4-Heptamethoxyflavone, a Citrus Flavonoid, Inhibits Collagenase Activity and Induces Type I Procollagen Synthesis in HDFn Cells. Int. J. Mol. Sci. 2018, 19, 620. [Google Scholar] [CrossRef]
- Lee, Y.Y.; Lee, E.-J.; Park, J.-S.; Jang, S.-E.; Kim, D.-H.; Kim, H.-S. Anti-Inflammatory and Antioxidant Mechanism of Tangeretin in Activated Microglia. J. Neuroimmune Pharmacol. 2016, 11, 294–305. [Google Scholar] [CrossRef]
- Guo, X.-Q.; Cao, Y.-L.; Hao, F.; Yan, Z.-R.; Wang, M.-L.; Liu, X.-W. Tangeretin alters neuronal apoptosis and ameliorates the severity of seizures in experimental epilepsy-induced rats by modulating apoptotic protein expressions, regulating matrix metalloproteinases, and activating the PI3K/Akt cell survival pathway. Adv. Med. Sci. 2017, 62, 246–253. [Google Scholar] [CrossRef]
- Sawamoto, A.; Okuyama, S.; Amakura, Y.; Yoshimura, M.; Yamada, T.; Yokogoshi, H.; Nakajima, M.; Furukawa, Y. 3,5,6,7,8,3′,4′-Heptamethoxyflavone Ameliorates Depressive-Like Behavior and Hippocampal Neurochemical Changes in Chronic Unpredictable Mild Stressed Mice by Regulating the Brain-Derived Neurotrophic Factor: Requirement for ERK Activation. Int. J. Mol. Sci. 2017, 18, 2133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, J.-J.; Huang, C.-C.; Su, Y.-L.; Luo, H.-L.; Lee, N.-L.; Sung, M.-T.; Wu, Y.-J. Proteomics Analysis of Tangeretin-Induced Apoptosis through Mitochondrial Dysfunction in Bladder Cancer Cells. Int. J. Mol. Sci. 2019, 20, 1017. [Google Scholar] [CrossRef] [PubMed]
- Iwase, Y.; Takemura, Y.; Ju-ichi, M.; Yano, M.; Ito, C.; Furukawa, H.; Mukainaka, T.; Kuchide, M.; Tokuda, H.; Nishino, H. Cancer chemopreventive activity of 3,5,6,7,8,3′,4′-heptamethoxyflavone from the peel of citrus plants. Cancer Lett. 2001, 163, 7–9. [Google Scholar] [CrossRef]
- Chen, Y.-K.; Wang, H.-C.; Ho, C.-T.; Chen, H.-Y.; Li, S.; Chan, H.-L.; Chung, T.-W.; Tan, K.-T.; Li, Y.-R.; Lin, C.-C. 5-Demethylnobiletin promotes the formation of polymerized tubulin, leads to G2/M phase arrest and induces autophagy via JNK activation in human lung cancer cells. J. Nutr. Biochem. 2015, 26, 484–504. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Wu, Y.; Ma, Y.; Cheng, Y.; Song, G.; Zhang, F. Systematic analysis of the mechanism of aged citrus peel (Chenpi) in oral squamous cell carcinoma treatment via network pharmacology, molecular docking and experimental validation. J. Funct. Foods 2022, 91, 105012. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IC50 Values (µg/mL) | |||||
|---|---|---|---|---|---|
| Samples | Fresh-F | PCR-C01F | PCR-C03F | PCR-C05F | PCR-C10F |
| A549 | 379.4 | 213.7 | 253.6 | 208.2 | 190.2 |
| IC50 Values (µM) | |||
|---|---|---|---|
| Samples | Tangeretin | Isosinensetin | 3,5,6,7,8,3′,4′-Heptamethoxyflavone |
| A549 | 118.5 | 197.6 | 208.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tao, Y.; Yu, Q.; Huang, Y.; Liu, R.; Zhang, X.; Wu, T.; Pan, S.; Xu, X. Identification of Crucial Polymethoxyflavones Tangeretin and 3,5,6,7,8,3′,4′-Heptamethoxyflavone and Evaluation of Their Contribution to Anticancer Effects of Pericarpium Citri Reticulatae ‘Chachi’ during Storage. Antioxidants 2022, 11, 1922. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11101922
Tao Y, Yu Q, Huang Y, Liu R, Zhang X, Wu T, Pan S, Xu X. Identification of Crucial Polymethoxyflavones Tangeretin and 3,5,6,7,8,3′,4′-Heptamethoxyflavone and Evaluation of Their Contribution to Anticancer Effects of Pericarpium Citri Reticulatae ‘Chachi’ during Storage. Antioxidants. 2022; 11(10):1922. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11101922
Chicago/Turabian StyleTao, Yexing, Qian Yu, Yuting Huang, Ruiting Liu, Xiwen Zhang, Ting Wu, Siyi Pan, and Xiaoyun Xu. 2022. "Identification of Crucial Polymethoxyflavones Tangeretin and 3,5,6,7,8,3′,4′-Heptamethoxyflavone and Evaluation of Their Contribution to Anticancer Effects of Pericarpium Citri Reticulatae ‘Chachi’ during Storage" Antioxidants 11, no. 10: 1922. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11101922