
Spatial Distribution, Antioxidant Capacity, and Spore Germination-Promoting Effect of Bibenzyls from Marchantia polymorpha
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Plant Samples
2.2. Sample Preparation of MSI and AFADESI-MSI Analysis
2.3. LC–MS/MS analysis
2.4. RT-Q-PCR Analysis
2.5. Determination of Antioxidant Activity of Crude Extract in Different Tissues of M. polymorpha
2.5.1. Preparation of Methanol Extracts
2.5.2. Total Phenolic Acid Content (TPC) Determination
2.5.3. DPPH Radical Scavenging
2.5.4. ABTS Radical Scavenging
2.5.5. Ferric Reducing Antioxidant Power (FRAP)
2.6. Expression Patterns of STCS1s and TPC Response to UV Treatment
2.7. Spore Germination Rate Assay
2.8. Statistical Analysis
3. Results and Discussion
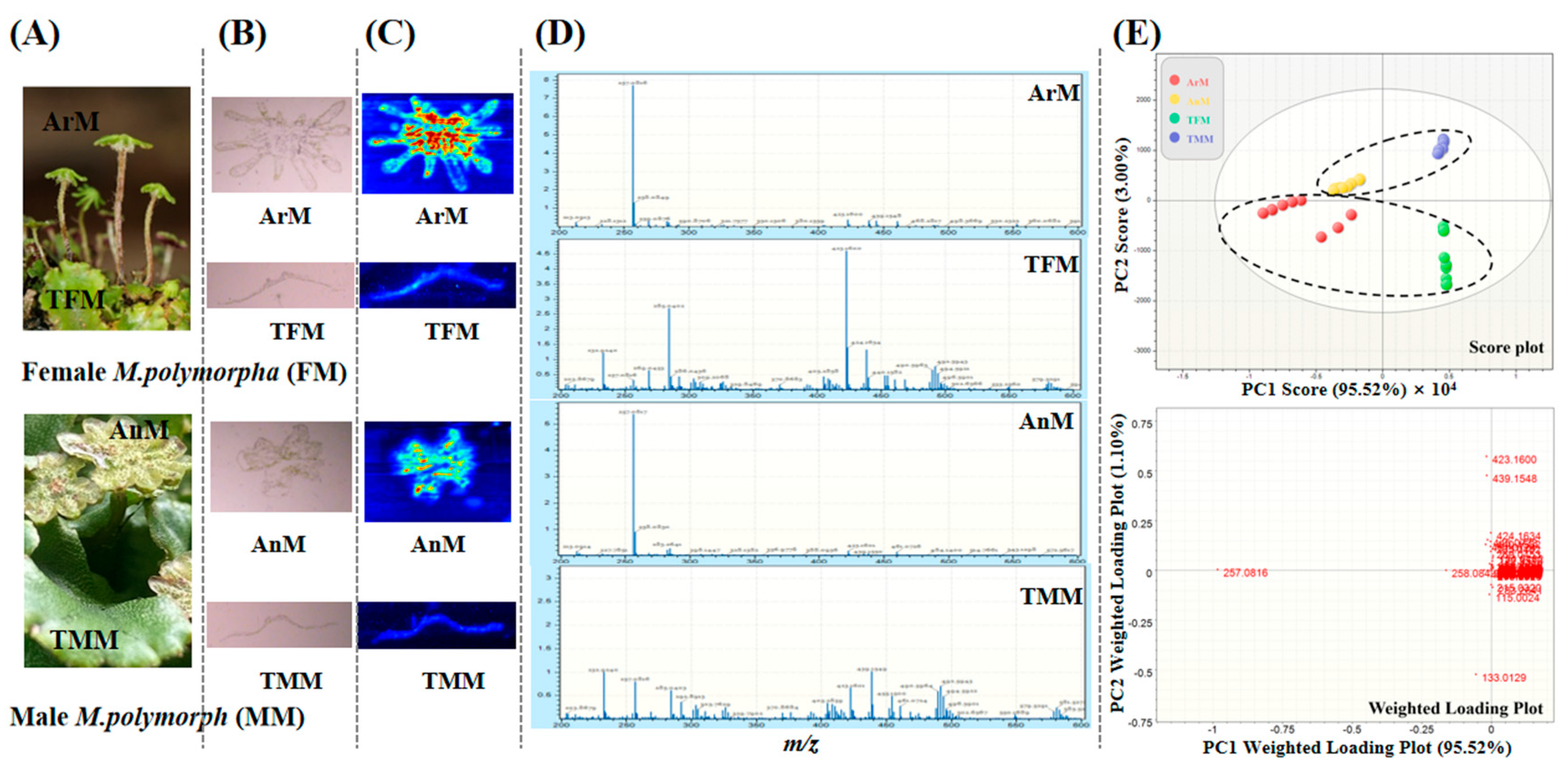
3.1. Data Mining of Characteristic Metabolites from M. polymorpha Using AFADESI
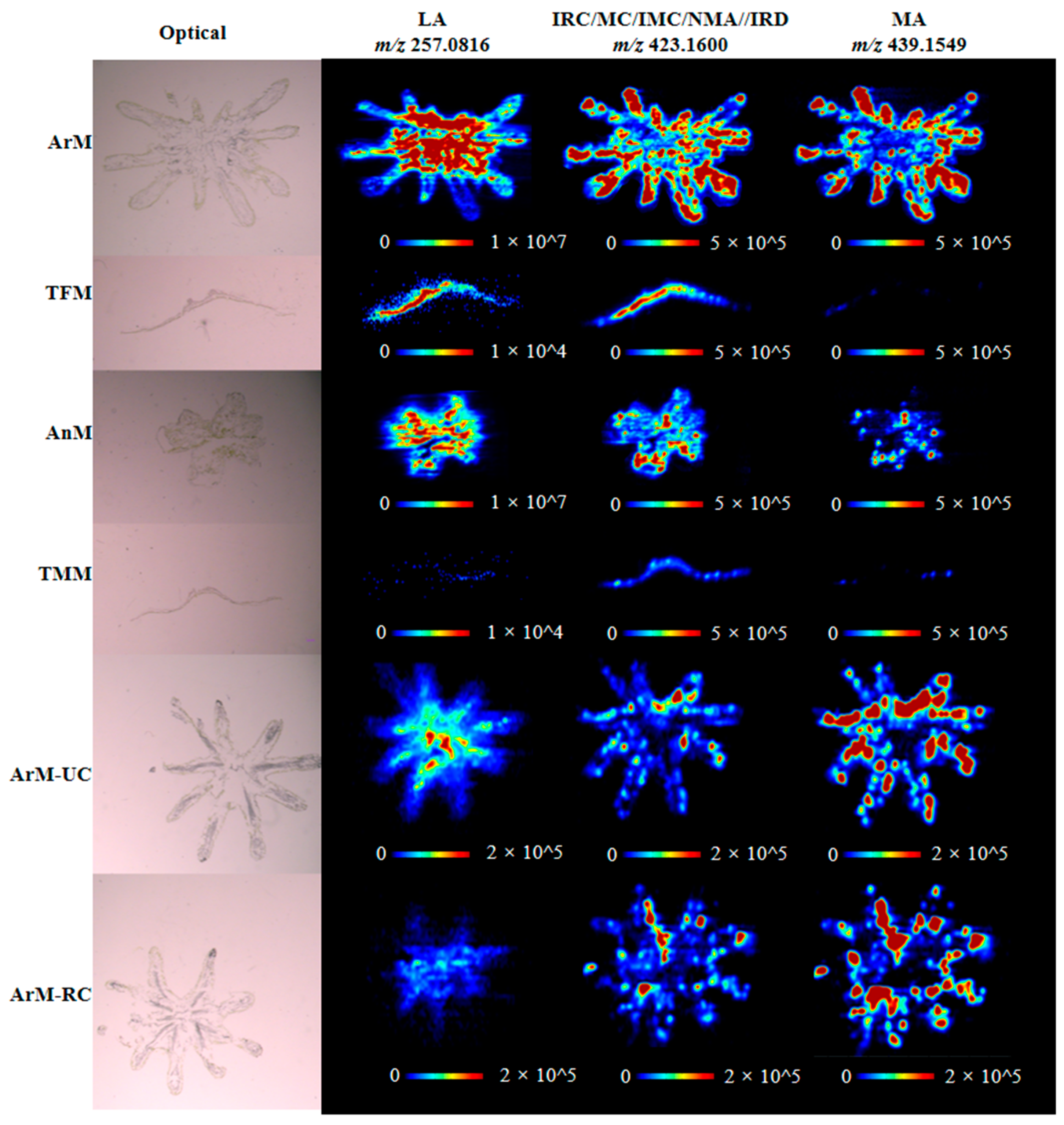
3.2. Spatial Distribution of Bibenzyls in M. polymorpha
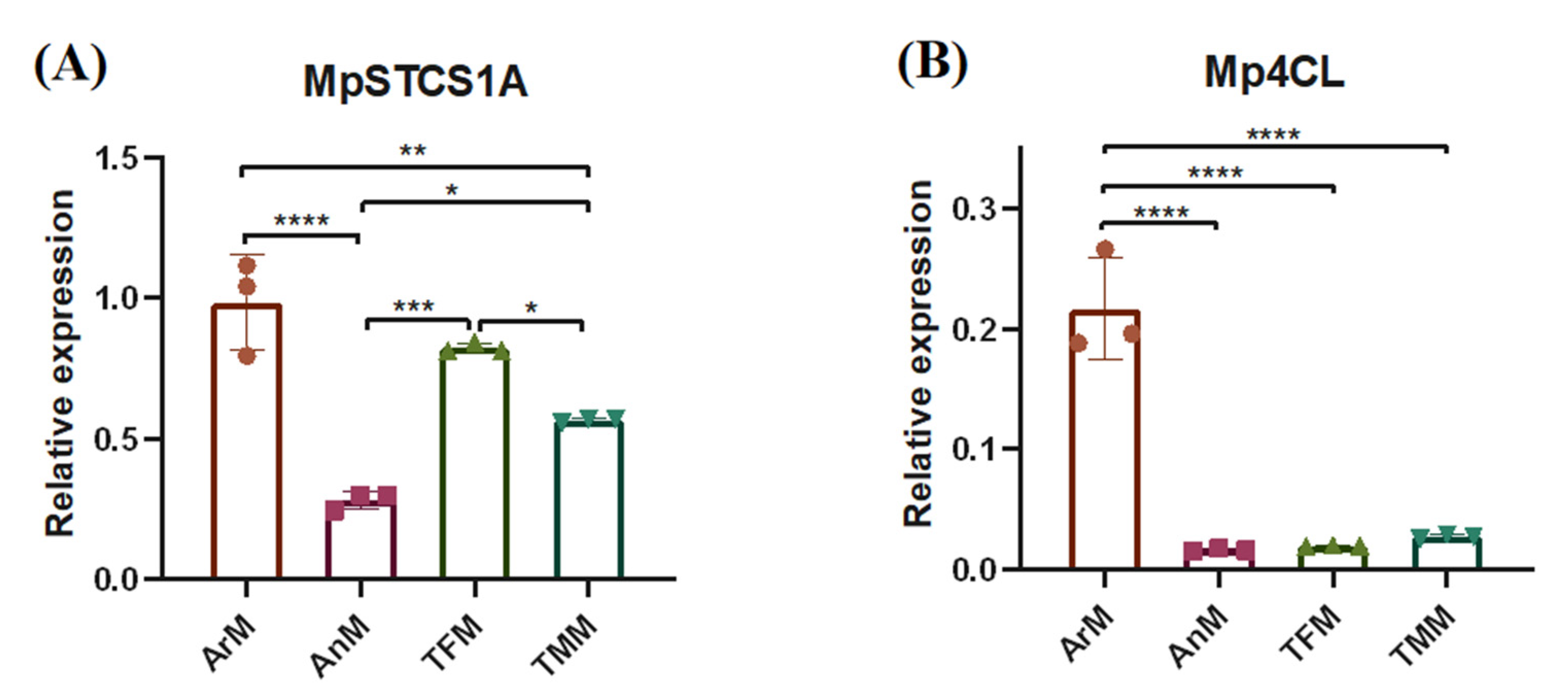
3.3. Expression Patterns of Key Bibenzyl Biosysthesis Genes in Four Parts of M. polymorpha
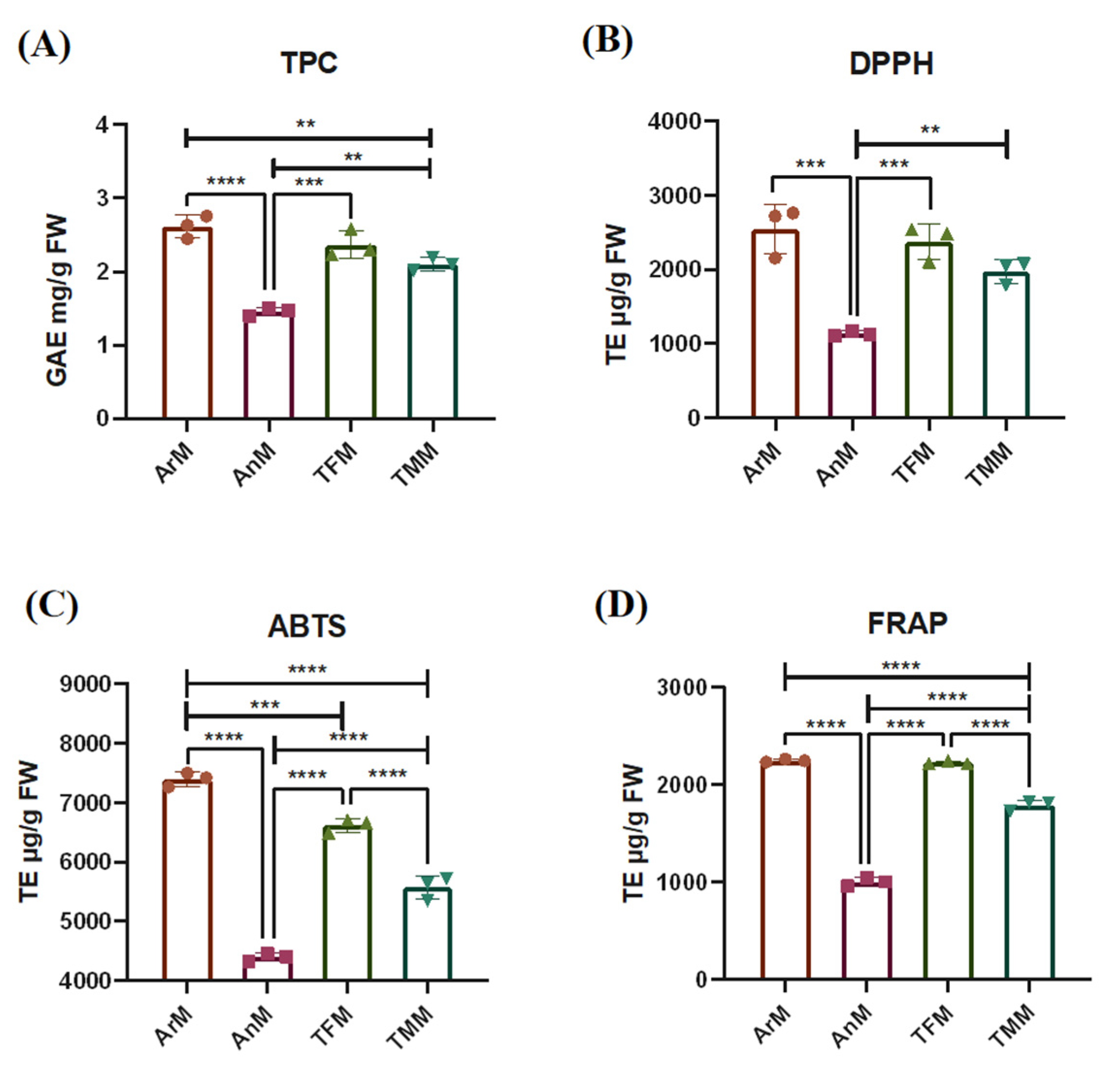
3.4. Antioxidant Activity Evaluation of Four Parts from M. polymorpha
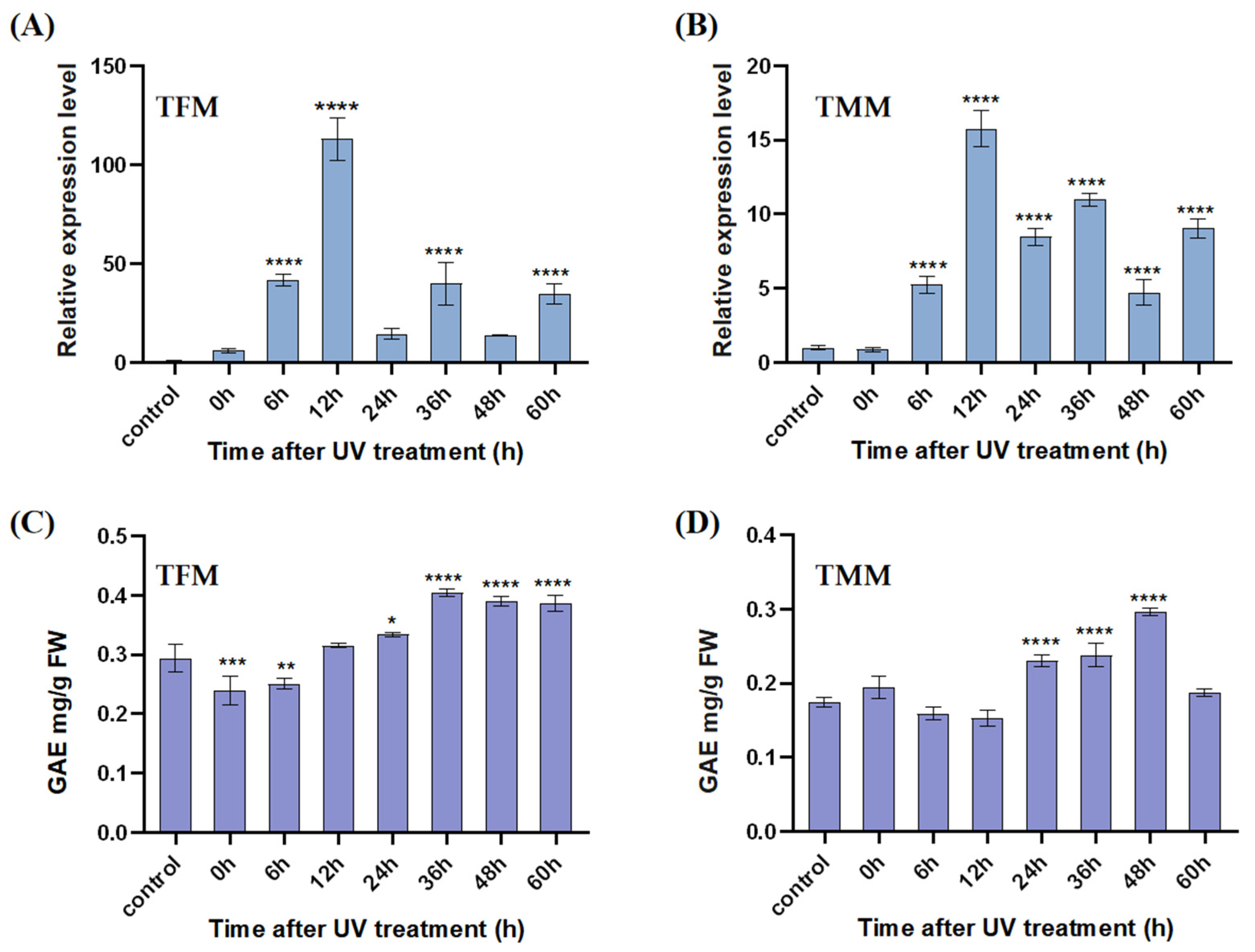
3.5. Analysis of Gene Expression Patterns and TPC Responses after UV Treatment
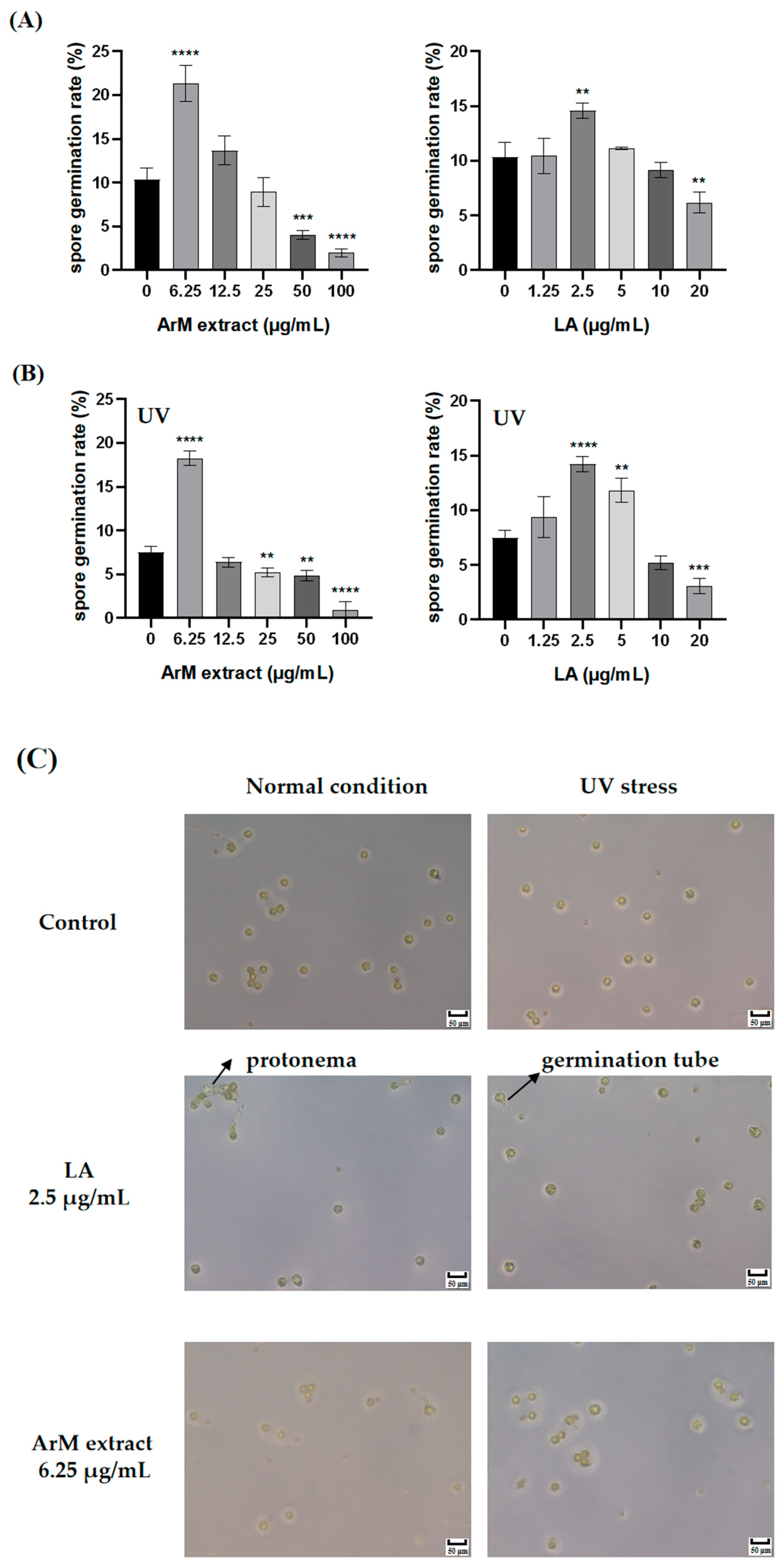
3.6. Promotion Effect of LA and Extraction of ArM on Spore Germination under Normal Conditions and UV Stress
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wellman, C.H.; Osterloff, P.L.; Mohiuddin, U. Fragments of the earliest land plants. Nature 2003, 425, 282–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowman, J.L.; Kohchi, T.; Yamato, K.T. Insights into land plant evolution garnered from the Marchantia polymorpha genome. Cell 2017, 171, 287–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huttunen, S.; Lappalainen, N.M.; Turunen, J. UV-absorbing compounds in subarctic herbarium bryophytes. Environ. Pollut. 2005, 133, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Hori, K.; Maruyama, F.; Fujisawa, T.; Togashi, T.; Yamamoto, N.; Seo, M.; Sato, S.; Yamada, T.; Mori, H.; Tajima, N.; et al. Klebsormidium flaccidum genome reveals primary factors for plant terrestrial adaptation. Nat. Commun. 2014, 5, 3978. [Google Scholar] [CrossRef] [Green Version]
- Zhu, T.T.; Sun, C.J.; Liu, X.Y.; Zhang, J.Z.; Hou, X.B.; Ni, R.; Zhang, J.; Cheng, A.X.; Lou, H.X. Interaction of PKR with STCS1: An indispensable step in the biosynthesis of lunularic acid in Marchantia polymorpha. New Phytol. 2022. [Google Scholar] [CrossRef]
- Ko, F.N.; Liao, C.H.; Wu, C.L. Marchantinquinone, isolated from Reboulia hemisphaerica, as inhibitor of lipid peroxidation and as free radical scavenger. Chem. Biol. Interact. 1995, 98, 131–143. [Google Scholar] [CrossRef]
- Schmid, M.W.; Giraldo-Fonseca, A.; Rövekamp, M.; Vekamp, M.; Smetanin, D.; Bowman, J.L.; Grossniklaus, U. Extensive epigenetic reprogramming during the life cycle of Marchantia polymorpha. Genome Biol. 2018, 19, 9. [Google Scholar] [CrossRef]
- Iwasaki, M.; Kajiwara, T.; Yasui, Y.; Yoshitake, Y.; Miyazaki, M.; Kawamura, S.; Suetsugu, N.; Nishihama, R.; Yamaoka, S.; Wanke, D.; et al. Identification of the sex-determining factor in the liverwort Marchantia polymorpha reveals unique evolution of sex chromosomes in a haploid system. Curr. Biol. 2021, 24, 5522–5532. [Google Scholar] [CrossRef]
- Hansen, R.L.; Lee, Y.J. High-spatial resolution mass spectrometr imaging: Toward single cell metabolomics in plant tissues. Chem. Rec. 2018, 18, 65–77. [Google Scholar] [CrossRef]
- Spraker, J.E.; Luu, G.T.; Sanchez, L.M. Imaging mass spectrometry for natural products discovery: A review of ionization methods. Nat. Prod. Rep. 2020, 37, 150–162. [Google Scholar] [CrossRef]
- Spengler, B. Mass spectrometry imaging of biomolecular information. Anal. Chem. 2015, 87, 64–82. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Du, Q.; Song, X.; Gao, S.; He, J. Evaluation of the tumor-targeting efficiency and intratumor heterogeneity of anticancer drugs using quantitative mass spectrometry imaging. Theranostics 2020, 10, 2621–2630. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Mao, X.; Sun, C.; Luo, Z.; Song, X.; Li, X.; Zhang, R.; Lv, Y.; Chen, J.; He, J.; et al. A graphical data processing pipeline for mass spectrometry imaging-based spatially resolved metabolomics on tumor heterogeneity. Anal. Chim. Acta 2019, 1077, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Gambino, G.; Perrone, I.; Gribaudo, I. A Rapid and effective method for RNA extraction from different tissues of grapevine and other woody plants. Phytochem. Anal. PCA 2008, 19, 520–525. [Google Scholar] [CrossRef]
- Afonso, S.; Oliveira, I.V.; Meyer, A.S.; Aires, A.; Saavedra, M.J.; Gonçalves, B. Phenolic profile and bioactive potential of stems and seed kernels of sweet cherry fruit. Antioxidants 2020, 9, 1295. [Google Scholar] [CrossRef]
- Tadashi, N.; Akeo, K.; Masamitsu, W. Photoinduction of spore germination in Marchantia polymorpha L. is mediated by photosynthesis. Plant Cell Physiol. 1999, 10, 1014–1020. [Google Scholar]
- Cerrato, A.; Aita, S.E.; Capriotti, A.L.; Cavaliere, C.; Montone, C.M.; Piovesana, S.; Laganà, A. Fully automatized detection of phosphocholine-containing lipids through an isotopically labeled buffer modification workflow. Anal. Chem. 2021, 93, 15042–15048. [Google Scholar] [CrossRef]
- Tang, Y.C.; Liu, Y.J.; He, G.R.; Cao, Y.W.; Bi, M.M.; Song, M.; Yang, P.P.; Xu, L.F.; Ming, J. Comprehensive analysis of secondary metabolites in the extracts from different lily bulbs and their antioxidant ability. Antioxidants 2021, 10, 1634. [Google Scholar] [CrossRef]
- Yoshikawa, H.; Ichiki, Y.; Sakakibara, K.; Tamura, H.; Suiko, M. The biological and structural similarity between lunularic acid and abscisic acid. Biosci. Biotechnol. Biochem. 2002, 66, 840–846. [Google Scholar] [CrossRef] [Green Version]
- Asakawa, Y.; Ludwiczuk, A.; Nagashima, F. Phytochemical and biological studies of bryophytes. Phytochemistry 2013, 91, 52–80. [Google Scholar] [CrossRef]
- Sugano, S.S.; Shirakawa, M.; Takagi, J.; Matsuda, Y.; Shimada, T.; Hara-Nishimura, I.; Kohchi, T. CRISPR/Cas9-mediated targeted mutagenesis in the liverwort Marchantia polymorpha L. Plant Cell Physiol. 2014, 55, 475–481. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.-Z.; Wang, C.; Zhu, T.-T.; Fu, J.; Tan, H.; Zhang, C.-M.; Cheng, A.-X.; Lou, H.-X. Spatial Distribution, Antioxidant Capacity, and Spore Germination-Promoting Effect of Bibenzyls from Marchantia polymorpha. Antioxidants 2022, 11, 2157. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11112157
Zhang J-Z, Wang C, Zhu T-T, Fu J, Tan H, Zhang C-M, Cheng A-X, Lou H-X. Spatial Distribution, Antioxidant Capacity, and Spore Germination-Promoting Effect of Bibenzyls from Marchantia polymorpha. Antioxidants. 2022; 11(11):2157. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11112157
Chicago/Turabian StyleZhang, Jiao-Zhen, Chan Wang, Ting-Ting Zhu, Jie Fu, Hui Tan, Cheng-Min Zhang, Ai-Xia Cheng, and Hong-Xiang Lou. 2022. "Spatial Distribution, Antioxidant Capacity, and Spore Germination-Promoting Effect of Bibenzyls from Marchantia polymorpha" Antioxidants 11, no. 11: 2157. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11112157