
Effect of Extracellular Matrix Stiffness on Candesartan Efficacy in Anti-Fibrosis and Antioxidation

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Assessment of Cardiac Functions of Rats
2.3. Staining Masson’s Trichrome
2.4. Measurement of Young’s Modulus of Myocardial Tissue
2.5. Measurement of Brain Natriuretic Peptide (BNP) and Cardiac Troponin T (c-TnT)
2.6. Immunohistochemical Analysis of AT1R, p-FAK, FAK and NOX2
2.7. Preparation and Characterization of Polyacrylamide (PA) Gels
2.8. Isolation and Purified Culture of Neonatal Rat Cardiac Fibroblasts
2.9. Immunofluorescence Staining of α-Smooth Muscle Actin, p-FAK and AT1R
2.10. Western Blotting
2.11. SECM Measurements of Extracellular GSH Levels
2.12. Statistical Analysis
3. Results
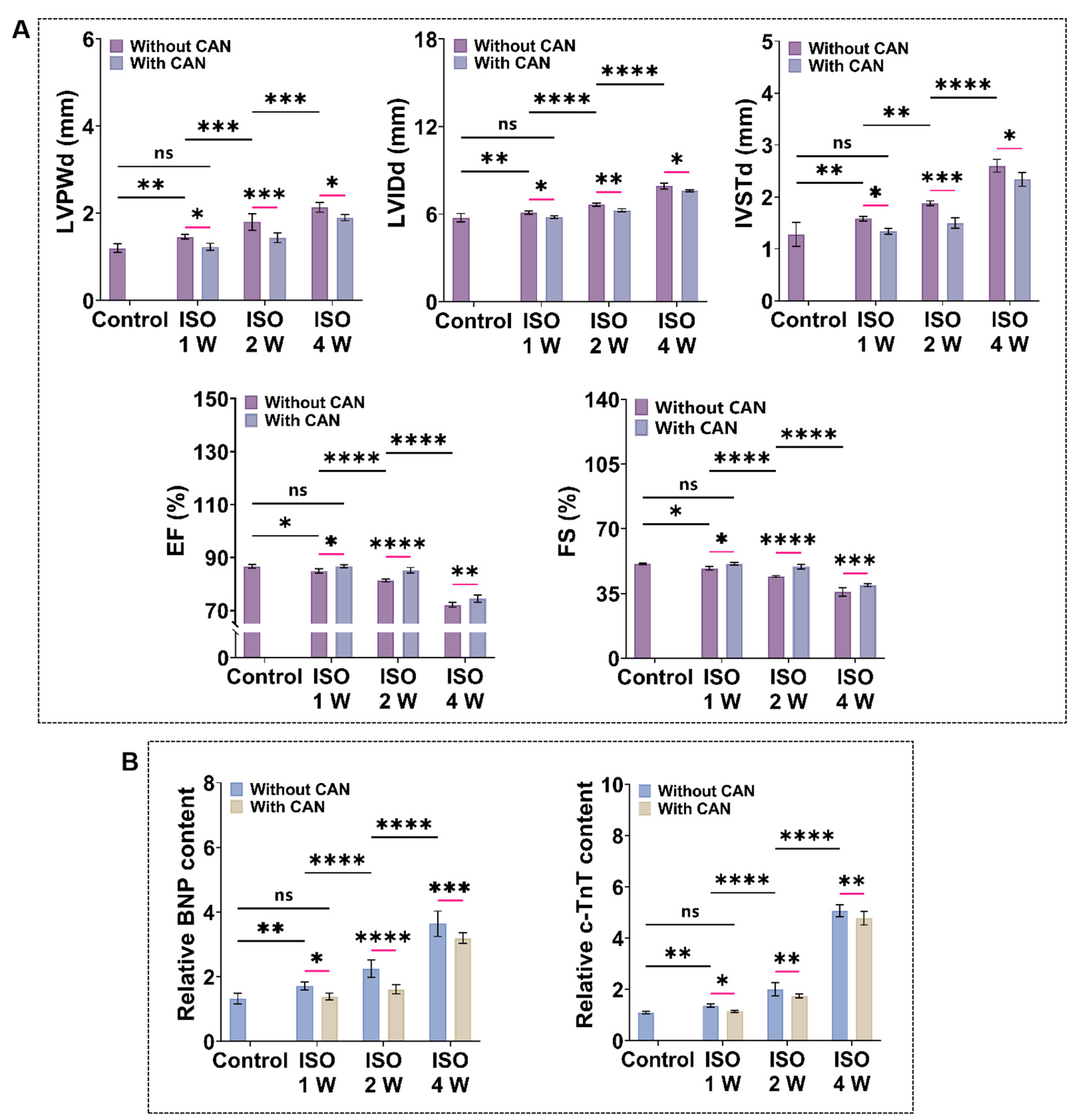
3.1. CAN Treatment Results in Decreased Myocardial Stiffness and Collagen Content in Myocardial Fibrosis Rats
3.2. CAN Treatment Improves the Cardiac Function of Myocardial Fibrosis Rats
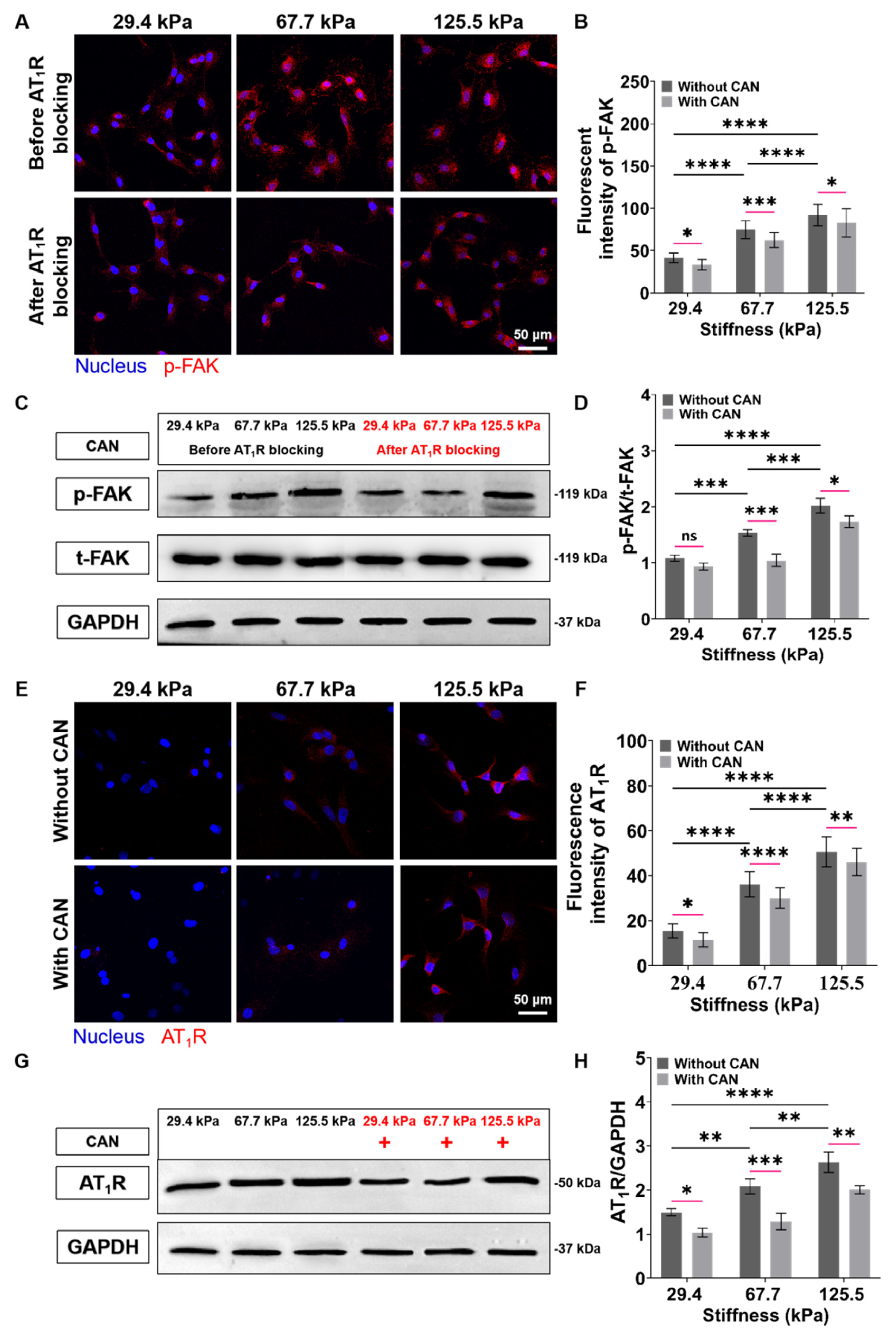
3.3. CAN Treatment Decreases the Expressions of AT1R and p-FAK of Myocardial Fibrosis Rats
3.4. CAN Treatment Ameliorates the Oxidative Stress State of Myocardial Fibrosis Rats
3.5. CAN Treatment Inhibits CF Migration and Proliferation under Different ECM Stiffness
3.6. CAN Treatment Inhibits CF Activation and Collagen Protein Production under Different ECM Stiffness
3.7. CAN Treatment Ameliorates Redox Imbalance of CFs under Different ECM Stiffness
3.8. ECM Stiffness-Dependent Anti-Fibrosis and Antioxidant Efficacy of CAN by Regulation of AT1R and p-FAK
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- de Boer, R.A.; De Keulenaer, G.; Bauersachs, J.; Brutsaert, D.; Cleland, J.G.; Diez, J.; Du, X.J.; Ford, P.; Heinzel, F.R.; Lipson, K.E.; et al. Towards better definition, quantification and treatment of fibrosis in heart failure. A scientific roadmap by the Committee of Translational Research of the Heart Failure Association (HFA) of the European Society of Cardiology. Eur. J. Heart Fail. 2019, 21, 272–285. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.G.; Yuan, Y.P.; Wu, H.M.; Zhang, X.; Tang, Q.Z. Cardiac fibrosis: New insights into the pathogenesis. Int. J. Biol. Sci. 2018, 14, 1645–1657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, J.; Zhang, Q.; Zhu, T.; Zhang, Y.; Liu, B.; Xu, J.; Zhao, H. Substrate stiffness-regulated matrix metalloproteinase output in myocardial cells and cardiac fibroblasts: Implications for myocardial fibrosis. Acta Biomater. 2014, 10, 2463–2472. [Google Scholar] [CrossRef] [PubMed]
- Mouton, A.J.; Rivera, O.J.; Lindsey, M.L. Myocardial infarction remodeling that progresses to heart failure: A signaling misunderstanding. Am. J. Physiol. Heart Circ. Physiol. 2018, 315, H71–H79. [Google Scholar] [CrossRef] [PubMed]
- Yong, K.W.; Li, Y.; Liu, F.; Gao, B.; Lu, T.J.; Wan Abas, W.A.; Wan Safwani, W.K.; Pingguan-Murphy, B.; Ma, Y.; Xu, F.; et al. Paracrine Effects of Adipose-Derived Stem Cells on Matrix Stiffness-Induced Cardiac Myofibroblast Differentiation via Angiotensin II Type 1 Receptor and Smad7. Sci. Rep. 2016, 6, 33067. [Google Scholar] [CrossRef]
- Li, R.; Frangogiannis, N.G. Integrins in cardiac fibrosis. J. Mol. Cell. Cardiol. 2022, 172, 1–13. [Google Scholar] [CrossRef]
- Li, Y.; Lang, J.; Ye, Z.; Wang, M.; Yang, Y.; Guo, X.; Zhuang, J.; Zhang, J.; Xu, F.; Li, F. Effect of Substrate Stiffness on Redox State of Single Cardiomyocyte: A Scanning Electrochemical Microscopy Study. Anal. Chem. 2020, 92, 4771–4779. [Google Scholar] [CrossRef]
- Yang, H.; Cheam, N.M.J.; Cao, H.; Lee, M.K.H.; Sze, S.K.; Tan, N.S.; Tay, C.Y. Materials Stiffness-Dependent Redox Metabolic Reprogramming of Mesenchymal Stem Cells for Secretome-Based Therapeutic Angiogenesis. Adv. Health Mater. 2019, 8, e1900929. [Google Scholar] [CrossRef]
- MacKenna, D.; Summerour, S.R.; Villarreal, F.J. Role of mechanical factors in modulating cardiac fibroblast function and extracellular matrix synthesis. Cardiovasc. Res. 2000, 46, 257–263. [Google Scholar] [CrossRef] [Green Version]
- Kanoldt, V.; Kluger, C.; Barz, C.; Schweizer, A.L.; Ramanujam, D.; Windgasse, L.; Engelhardt, S.; Chrostek-Grashoff, A.; Grashoff, C. Metavinculin modulates force transduction in cell adhesion sites. Nat. Commun. 2020, 11, 6403. [Google Scholar] [CrossRef]
- Mehta, P.K.; Griendling, K.K. Angiotensin II cell signaling: Physiological and pathological effects in the cardiovascular system. Am. J. Physiol. Cell Physiol. 2007, 292, C82–C97. [Google Scholar] [CrossRef] [PubMed]
- Tapial Martínez, P.; López Navajas, P.; Lietha, D. FAK Structure and Regulation by Membrane Interactions and Force in Focal Adhesions. Biomolecules 2020, 10, 179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brancaccio, M.; Hirsch, E.; Notte, A.; Selvetella, G.; Lembo, G.; Tarone, G. Integrin signalling: The tug-of-war in heart hypertrophy. Cardiovasc. Res. 2006, 70, 422–433. [Google Scholar] [CrossRef]
- Carragher, N.O.; Westhoff, M.A.; Fincham, V.J.; Schaller, M.D.; Frame, M.C. A novel role for FAK as a protease-targeting adaptor protein: Regulation by p42 ERK and Src. Curr. Biol. 2003, 13, 1442–1450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bond, M.; Sala-Newby, G.B.; Newby, A.C. Focal adhesion kinase (FAK)-dependent regulation of S-phase kinase-associated protein-2 (Skp-2) stability. A novel mechanism regulating smooth muscle cell proliferation. J. Biol. Chem. 2004, 279, 37304–37310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chauhan, A.; Khan, T. Focal adhesion kinase—An emerging viable target in cancer and development of focal adhesion kinase inhibitors. Chem. Biol. Drug Des. 2021, 97, 774–794. [Google Scholar] [CrossRef]
- Vadali, K.; Cai, X.; Schaller, M.D. Focal adhesion kinase: An essential kinase in the regulation of cardiovascular functions. IUBMB Life 2007, 59, 709–716. [Google Scholar] [CrossRef]
- Wei-Wei, L.; Wen-Wen, Z.; Jin-Jian, L.; Yi-Tao, W.; Xiu-Ping, C. Cucurbitacin B suppresses metastasis mediated by reactive oxygen species (ROS) via focal adhesion kinase (FAK) in breast cancer MDA-MB-231 cells. Chin. J. Nat. Med. 2018, 16, 10–19. [Google Scholar] [CrossRef]
- Ansell, T.K.; Noble, P.B.; Mitchell, H.W.; McFawn, P.K. Pharmacological bronchodilation is partially mediated by reduced airway wall stiffness. Br. J. Pharmacol. 2014, 171, 4376–4384. [Google Scholar] [CrossRef] [Green Version]
- Feng, J.; Tang, Y.; Xu, Y.; Sun, Q.; Liao, F.; Han, D. Substrate stiffness influences the outcome of antitumor drug screening in vitro. Clin. Hemorheol. Microcirc. 2013, 55, 121–131. [Google Scholar] [CrossRef]
- Gleiter, C.H.; Jägle, C.; Gresser, U.; Mörike, K. Candesartan. Cardiovasc. Drug Rev. 2004, 22, 263–284. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, D.; Kosugi, K.; Waki, H.; Yamamoto, K.; Tsujino, T.; Masuyama, T. Role of activated renin-angiotensin sys-tem in myocardial fibrosis and left ventricular diastolic dysfunction in diabetic patients—Reversal by chronic angio-tensin II type 1A receptor blockade. Circ. J. 2007, 71, 524–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tharaux, P.L.; Chatziantoniou, C.; Fakhouri, F.; Dussaule, J.C. Angiotensin II activates collagen I gene through a mechanism involving the MAP/ER kinase pathway. Hypertension 2000, 36, 330–336. [Google Scholar] [CrossRef] [Green Version]
- McKelvie, R.S. Candesartan for the management of heart failure: More than an alternative. Expert Opin. Pharmacother. 2006, 7, 1945–1956. [Google Scholar] [CrossRef] [PubMed]
- Onishi, K.; Dohi, K.; Koji, T.; Funabiki, K.; Kitamura, T.; Imanaka-Yoshida, K.; Ito, M.; Nobori, T.; Nakano, T. Candesartan prevents myocardial fibrosis during progression of congestive heart failure. J. Cardiovasc. Pharmacol. 2004, 43, 860–867. [Google Scholar] [CrossRef] [PubMed]
- National Research Council Committee for the Update of the Guide for the Care and Use of Laboratory Animals; Use of Laboratory Animals. The National Academies Collection: Reports funded by National Institutes of Health. In Guide for the Care and Use of Laboratory Animals; National Academies Press (US) Copyright © 2011, National Academy of Sciences: Washington, DC, USA, 2011. [Google Scholar] [CrossRef]
- Trappmann, B.; Gautrot, J.E.; Connelly, J.T.; Strange, D.G.; Li, Y.; Oyen, M.L.; Cohen Stuart, M.A.; Boehm, H.; Li, B.; Vogel, V.; et al. Extracellular-matrix tethering regulates stem-cell fate. Nat. Mater. 2012, 11, 642–649. [Google Scholar] [CrossRef]
- Sanzari, I.; Humphrey, E.J.; Dinelli, F.; Terracciano, C.M.; Prodromakis, T. Effect of patterned polyacrylamide hydrogel on morphology and orientation of cultured NRVMs. Sci. Rep. 2018, 8, 11991. [Google Scholar] [CrossRef]
- Lang, J.; Li, Y.; Ye, Z.; Yang, Y.; Xu, F.; Huang, G.; Zhang, J.; Li, F. Investigating the Effect of Substrate Stiffness on the Redox State of Cardiac Fibroblasts Using Scanning Electrochemical Microscopy. Anal. Chem. 2021, 93, 5797–5804. [Google Scholar] [CrossRef]
- Brandes, R.P.; Weissmann, N.; Schröder, K. Redox-mediated signal transduction by cardiovascular Nox NADPH oxidases. J. Mol. Cell. Cardiol. 2014, 73, 70–79. [Google Scholar] [CrossRef]
- Goldfracht, I.; Efraim, Y.; Shinnawi, R.; Kovalev, E.; Huber, I.; Gepstein, A.; Arbel, G.; Shaheen, N.; Tiburcy, M.; Zimmermann, W.H.; et al. Engineered heart tissue models from hiPSC-derived cardiomyocytes and cardiac ECM for disease modeling and drug testing applications. Acta Biomater. 2019, 92, 145–159. [Google Scholar] [CrossRef]
- Hofbauer, P.; Jahnel, S.M.; Mendjan, S. In vitro models of the human heart. Development 2021, 148, dev199672. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.H.; Vincent, L.G.; Fuhrmann, A.; Choi, Y.S.; Hribar, K.C.; Taylor-Weiner, H.; Chen, S.; Engler, A.J. Interplay of matrix stiffness and protein tethering in stem cell differentiation. Nat. Mater. 2014, 13, 979–987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinz, B.; McCulloch, C.A.; Coelho, N.M. Mechanical regulation of myofibroblast phenoconversion and collagen contraction. Exp. Cell Res. 2019, 379, 119–128. [Google Scholar] [CrossRef]
- Hinz, B. Myofibroblasts. Exp. Eye Res. 2016, 142, 56–70. [Google Scholar] [CrossRef] [PubMed]
- Hori, Y.; Yoshioka, K.; Kanai, K.; Hoshi, F.; Itoh, N.; Higuchi, S. Spironolactone decreases isoproterenol-induced ventricular fibrosis and matrix metalloproteinase-2 in rats. Biol. Pharm. Bull. 2011, 34, 61–65. [Google Scholar] [CrossRef] [Green Version]
- Teuber, J.P.; Essandoh, K.; Hummel, S.L.; Madamanchi, N.R.; Brody, M.J. NADPH Oxidases in Diastolic Dysfunction and Heart Failure with Preserved Ejection Fraction. Antioxidants 2022, 11, 1822. [Google Scholar] [CrossRef]
- Liu, T.; Sun, L.; Zhang, Y.; Wang, Y.; Zheng, J. Imbalanced GSH/ROS and sequential cell death. J. Biochem. Mol. Toxicol. 2022, 36, e22942. [Google Scholar] [CrossRef]
- Franco, R.; Cidlowski, J.A. Glutathione efflux and cell death. Antioxid. Redox Signal. 2012, 17, 1694–1713. [Google Scholar] [CrossRef] [Green Version]
- López-Blanch, R.; Salvador-Palmer, R.; Estrela, J.M.; Obrador, E. An Intercellular Flow of Glutathione Regulated by Interleukin 6 Links Astrocytes and the Liver in the Pathophysiology of Amyotrophic Lateral Sclerosis. Antioxidants 2021, 10, 2007. [Google Scholar] [CrossRef]
- Takahashi, Y.; Kumatani, A.; Shiku, H.; Matsue, T. Scanning Probe Microscopy for Nanoscale Electrochemical Imaging. Anal. Chem. 2017, 89, 342–357. [Google Scholar] [CrossRef]
- Zhang, J.J.; Zhu, T.; Lang, J.X.; Fu, W.X.; Li, F. Recent advances of scanning electrochemical microscopy and scanning ion conductance microscopy for single-cell analysis. Curr. Opin. Electrochem. 2020, 22, 178–185. [Google Scholar] [CrossRef]
- Li, Y.; Ye, Z.; Zhang, J.; Zhao, Y.; Zhu, T.; Song, J.; Xu, F.; Li, F. In Situ and Quantitative Monitoring of Cardiac Tissues Using Programmable Scanning Electrochemical Microscopy. Anal. Chem. 2022, 94, 10515–10523. [Google Scholar] [CrossRef] [PubMed]
- Hirano, Y.; Kodama, M.; Shibuya, M.; Maki, Y.; Komatsu, Y. Analysis of beat fluctuations and oxygen consumption in cardiomyocytes by scanning electrochemical microscopy. Anal. Biochem. 2014, 447, 39–42. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, V.Y.; Morgan, A.E.; Kim, J.; Tafreshi, R.; Wallace, A.W.; Guccione, J.M.; Weinsaft, J.W.; Ge, L.; Ratcliffe, M.B. Finite-element based optimization of left ventricular passive stiffness in normal volunteers and patients after myocardial infarction: Utility of an inverse deformation gradient calculation of regional diastolic strain. J. Mech. Behav. Biomed. Mater. 2021, 119, 104431. [Google Scholar] [CrossRef]
- Liao, F.; Li, M.; Han, D.; Cao, J.; Chen, K. Biomechanopharmacology: A new borderline discipline. Trends Pharmacol. Sci. 2006, 27, 287–289. [Google Scholar] [CrossRef]
- Yong, A.S.; Pennings, G.J.; Chang, M.; Hamzah, A.; Chung, T.; Qi, M.; Brieger, D.; Behnia, M.; Krilis, S.A.; Ng, M.K.; et al. Intracoronary shear-related up-regulation of platelet P-selectin and platelet-monocyte aggregation despite the use of aspirin and clopidogrel. Blood 2011, 117, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Q.; Chu, Z.; Li, X.; Kang, H.; Yang, X.; Fan, Y. The Effect of Fluid Shear Stress on the In Vitro Release Kinetics of Sirolimus from PLGA Films. Polymers 2017, 9, 618. [Google Scholar] [CrossRef] [Green Version]
- Jackson, S.P.; Schoenwaelder, S.M. Antithrombotic phosphoinositide 3-kinase β inhibitors in humans: A ‘shear’ delight! J. Thromb. Haemost. 2012, 10, 2123–2126. [Google Scholar] [CrossRef] [Green Version]
- Rahaman, S.O.; Grove, L.M.; Paruchuri, S.; Southern, B.D.; Abraham, S.; Niese, K.A.; Scheraga, R.G.; Ghosh, S.; Thodeti, C.K.; Zhang, D.X.; et al. TRPV4 mediates myofibroblast differentiation and pulmonary fibrosis in mice. J. Clin. Investig. 2014, 124, 5225–5238. [Google Scholar] [CrossRef]
- DiMichele, L.A.; Doherty, J.T.; Rojas, M.; Beggs, H.E.; Reichardt, L.F.; Mack, C.P.; Taylor, J.M. Myocyte-restricted focal adhesion kinase deletion attenuates pressure overload-induced hypertrophy. Circ. Res. 2006, 99, 636–645. [Google Scholar] [CrossRef] [Green Version]
- Clemente, C.F.; Tornatore, T.F.; Theizen, T.H.; Deckmann, A.C.; Pereira, T.C.; Lopes-Cendes, I.; Souza, J.R.; Franchini, K.G. Targeting focal adhesion kinase with small interfering RNA prevents and reverses load-induced cardiac hypertrophy in mice. Circ. Res. 2007, 101, 1339–1348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakai, M.; Yoshimura, R. Mechanotransduction-Targeting Drugs Attenuate Stiffness-Induced Hepatic Stellate Cell Activation in Vitro. Biol. Pharm. Bull. 2021, 44, 416–421. [Google Scholar] [CrossRef] [PubMed]
- Trevelin, S.C.; Zampetaki, A.; Sawyer, G.; Ivetic, A.; Brewer, A.C.; Smyth, L.A.; Marelli-Berg, F.; Köchl, R.; Lechler, R.I.; Shah, A.M.; et al. Nox2-deficient Tregs improve heart transplant outcomes via their increased graft recruitment and enhanced potency. JCI Insight 2021, 6, e149301. [Google Scholar] [CrossRef]
- Liu, R.M.; Pravia, K.A.G. Oxidative stress and glutathione in TGF-beta-mediated fibrogenesis. Free Radic. Biol. Med. 2010, 48, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mills, B.J.; Weiss, M.M.; Lang, C.A.; Liu, M.C.; Ziegler, C. Blood glutathione and cysteine changes in cardiovascular disease. J. Lab. Clin. Med. 2000, 135, 396–401. [Google Scholar] [CrossRef] [PubMed]
- Chaiswing, L.; Oberley, T.D. Extracellular/microenvironmental redox state. Antioxid. Redox Signal. 2010, 13, 449–465. [Google Scholar] [CrossRef]
- Urbano, R.L.; Swaminathan, S.; Clyne, A.M. Stiff Substrates Enhance Endothelial Oxidative Stress in Response to Protein Kinase C Activation. Appl. Bionics Biomech. 2019, 2019, 6578492. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | ISO (Day) | CAN (Day) |
|---|---|---|
| From day 1–day 7 | / |
| From day 1–day 14 | / |
| From day 1–day 28 | / |
| From day 1–day 7 | From day 1–day 28 |
| From day 1–day 14 | From day 1–day 28 |
| From day 1–day 28 | From day 1–day 28 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, T.; Song, J.; Gao, B.; Zhang, J.; Li, Y.; Ye, Z.; Zhao, Y.; Guo, X.; Xu, F.; Li, F. Effect of Extracellular Matrix Stiffness on Candesartan Efficacy in Anti-Fibrosis and Antioxidation. Antioxidants 2023, 12, 679. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox12030679
Zhu T, Song J, Gao B, Zhang J, Li Y, Ye Z, Zhao Y, Guo X, Xu F, Li F. Effect of Extracellular Matrix Stiffness on Candesartan Efficacy in Anti-Fibrosis and Antioxidation. Antioxidants. 2023; 12(3):679. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox12030679
Chicago/Turabian StyleZhu, Tong, Jingjing Song, Bin Gao, Junjie Zhang, Yabei Li, Zhaoyang Ye, Yuxiang Zhao, Xiaogang Guo, Feng Xu, and Fei Li. 2023. "Effect of Extracellular Matrix Stiffness on Candesartan Efficacy in Anti-Fibrosis and Antioxidation" Antioxidants 12, no. 3: 679. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox12030679