

The Association of Redox Regulatory Drug Target Genes with Psychiatric Disorders: A Mendelian Randomization Study
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
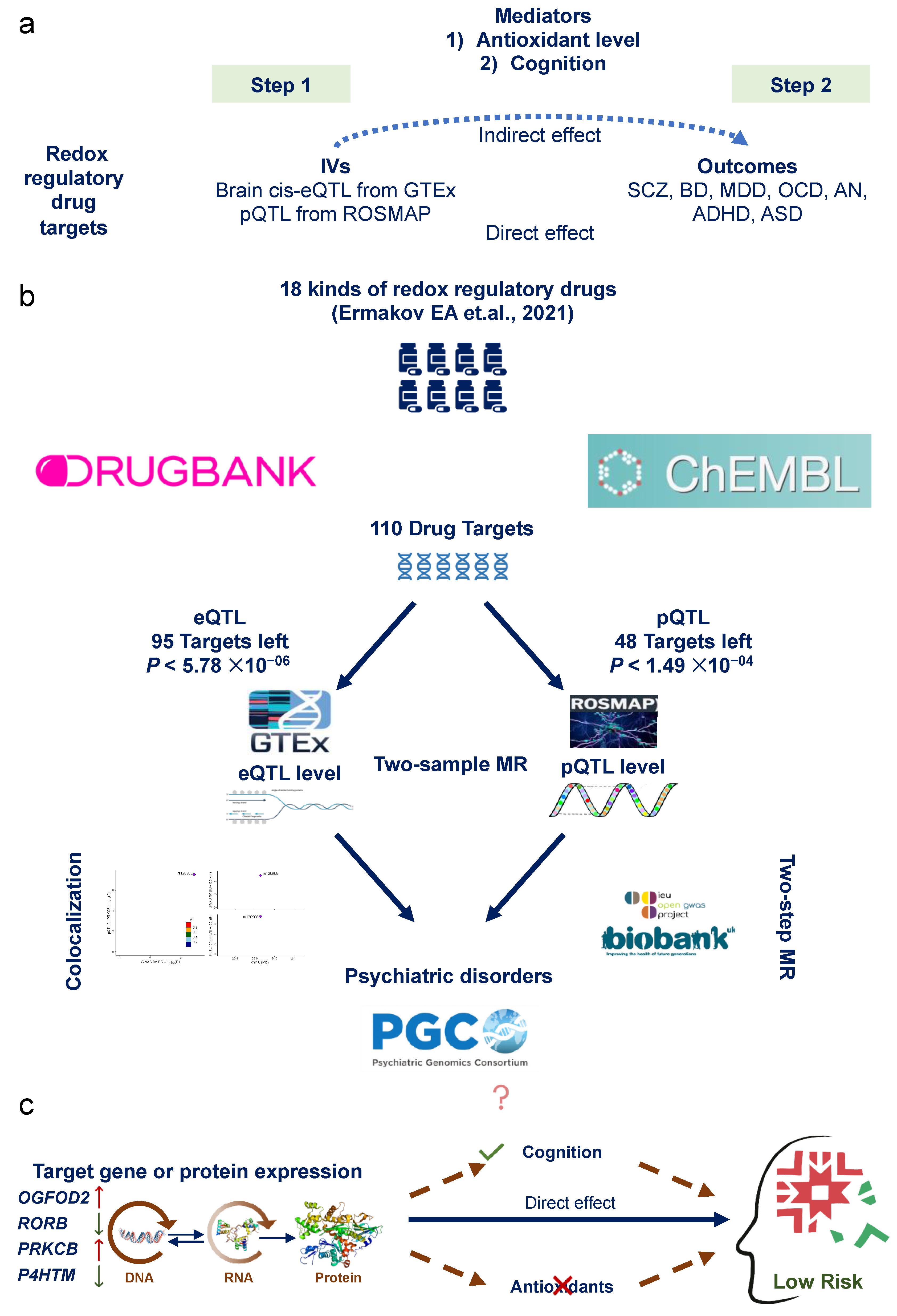
2.1. Study Design
2.2. Ethics Approval
2.3. Identification of Redox Regulatory Drug Target Genes
2.4. Genetic Instruments for Redox Regulatory Drug Target Genes
2.5. Outcome Data
2.6. Mendelian Randomization
2.7. Sensitivity Analyses
2.8. Two-Step Mendelian Randomization for Mediation Analysis
3. Results
3.1. Genetic Instrument Selection
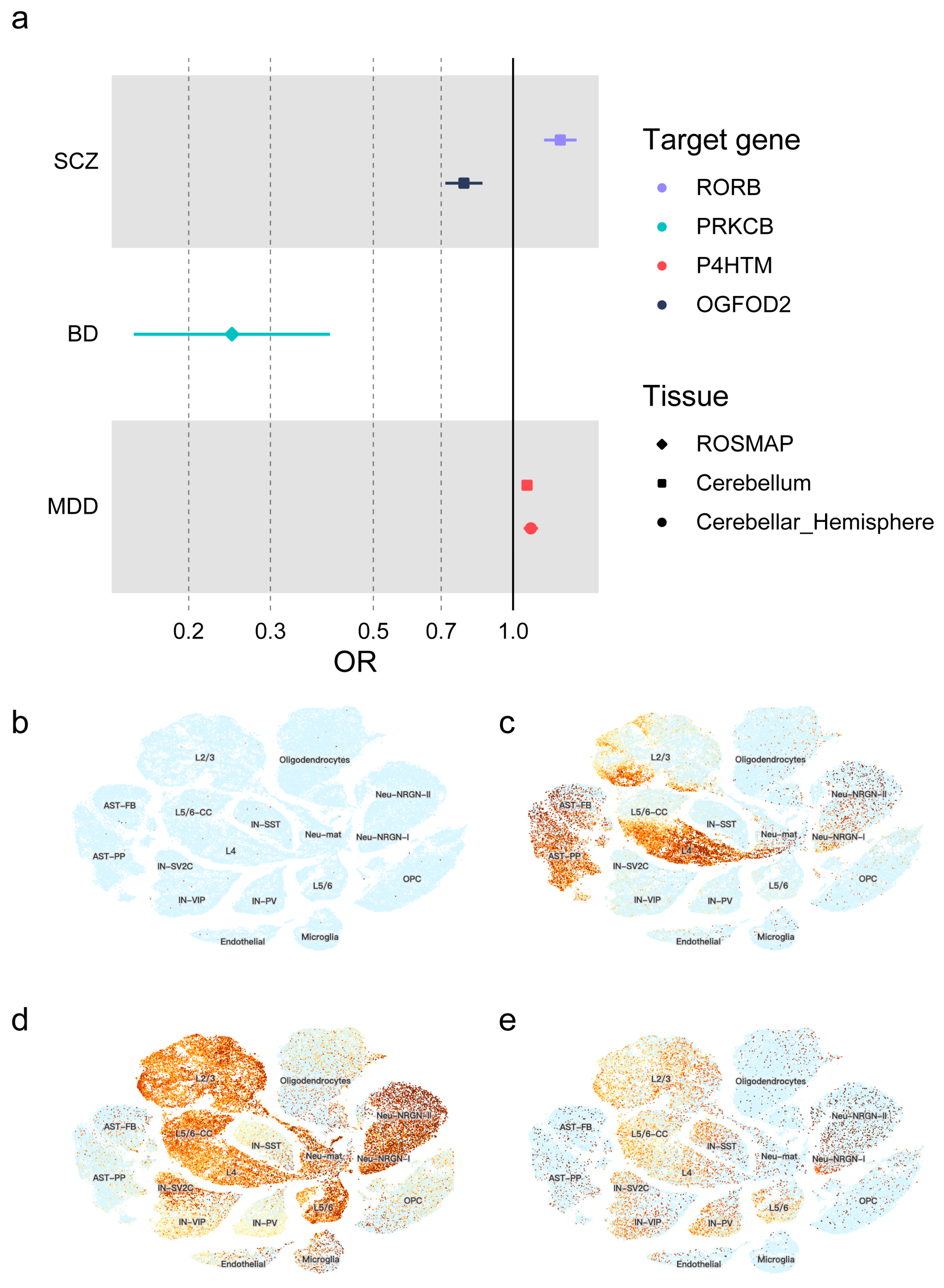
3.2. Causal Effects of Redox Regulatory Drug Targets on Psychiatric Disorders
3.3. Gene Expression Level in Single-Cell Clusters
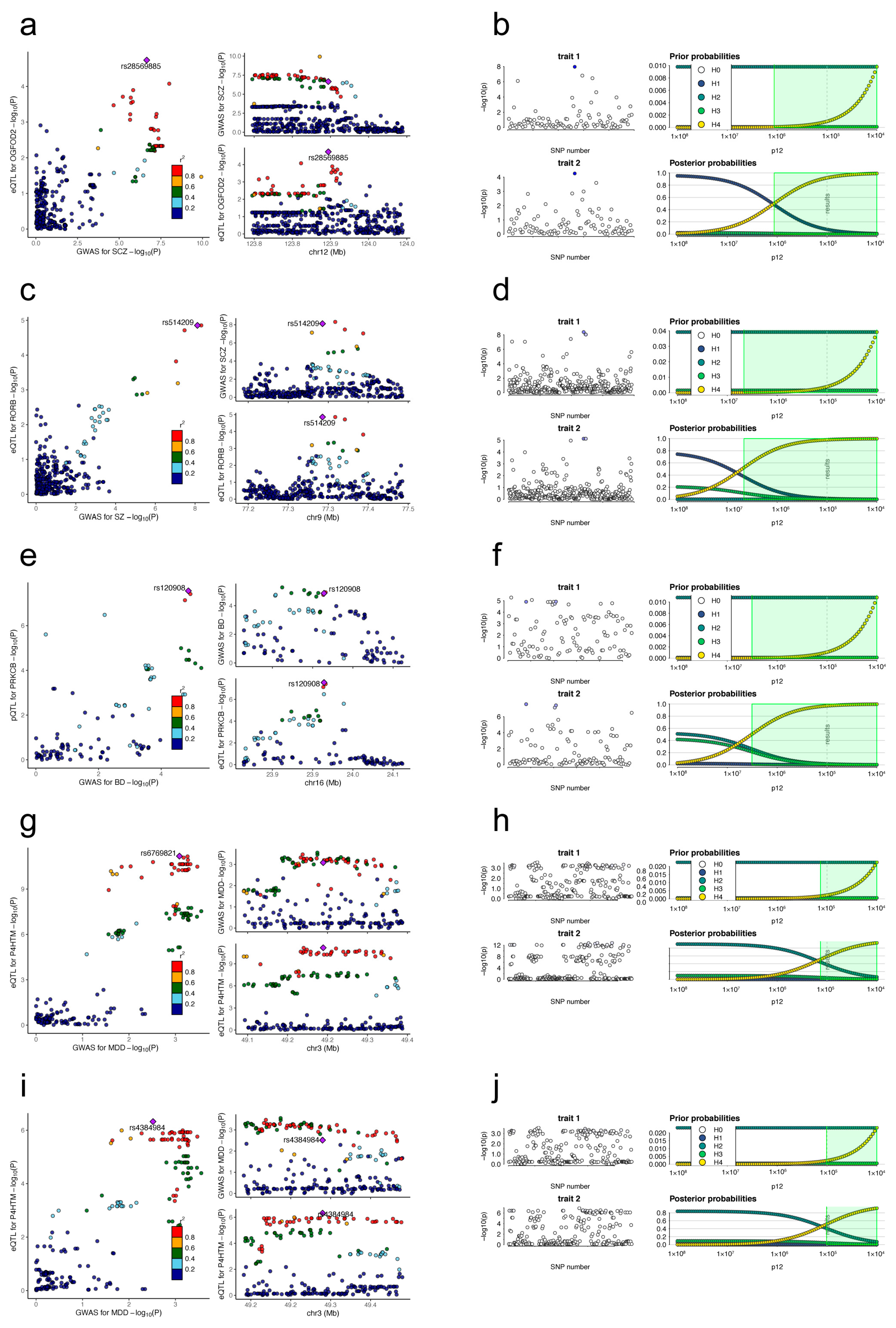
3.4. Colocalization Results
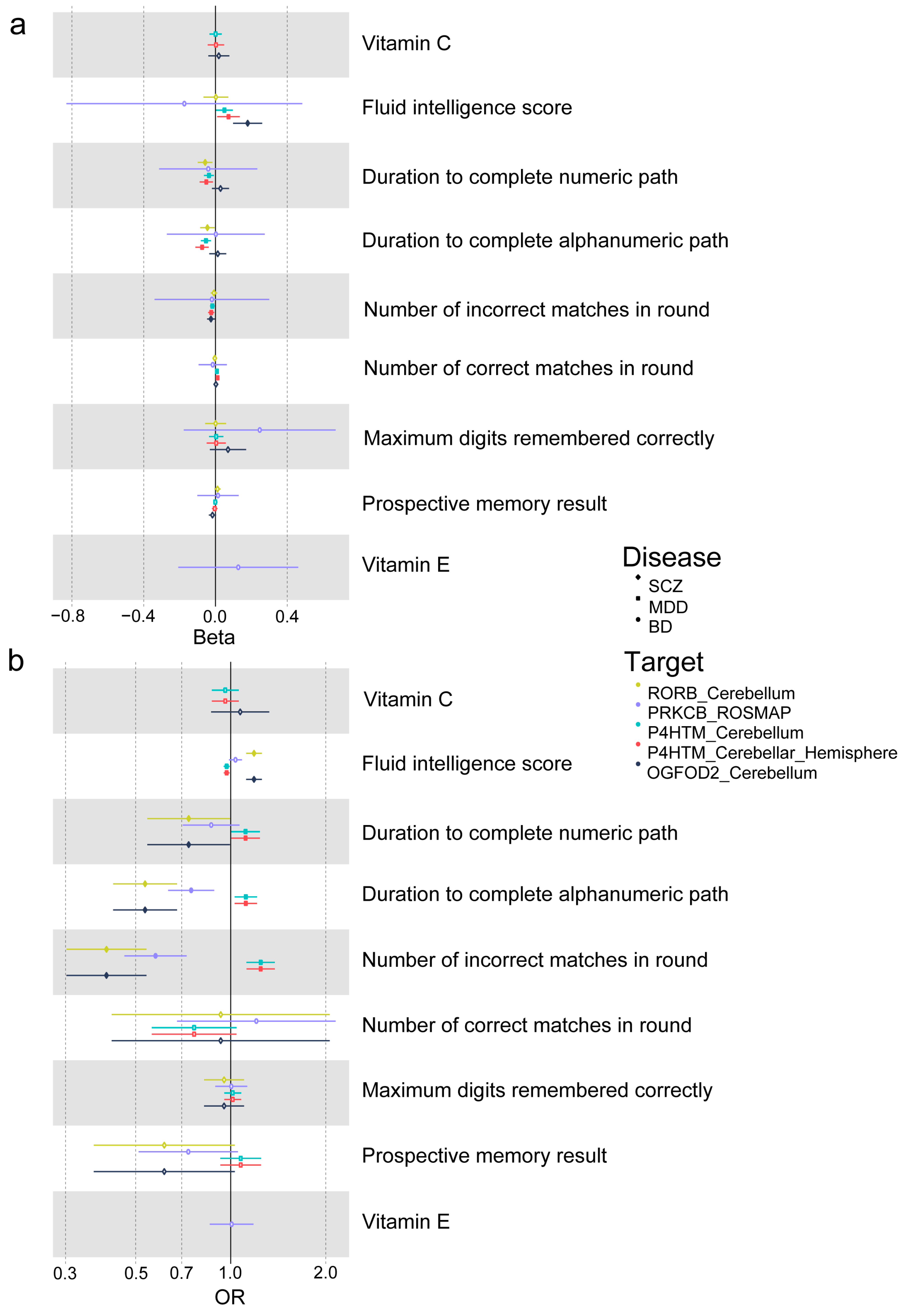
3.5. Two-Step MR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Marwaha, S.; Palmer, E.; Suppes, T.; Cons, E.; Young, A.H.; Upthegrove, R. Novel and emerging treatments for major depression. Lancet 2023, 401, 141–153. [Google Scholar] [CrossRef]
- Marder, S.R.; Umbricht, D. Negative symptoms in schizophrenia: Newly emerging measurements, pathways, and treatments. Schizophr. Res. 2023, 258, 71–77. [Google Scholar] [CrossRef]
- Paul, S.M.; Potter, W.Z. Finding new and better treatments for psychiatric disorders. Neuropsychopharmacology 2024, 49, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Sutkowy, P.; Woźniak, A.; Mila-Kierzenkowska, C.; Szewczyk-Golec, K.; Wesołowski, R.; Pawłowska, M.; Nuszkiewicz, J. Physical Activity vs. Redox Balance in the Brain: Brain Health, Aging and Diseases. Antioxidants 2021, 11, 95. [Google Scholar] [CrossRef] [PubMed]
- Lohr, J.B. Oxygen radicals and neuropsychiatric illness: Some speculations. Arch. Gen. Psychiatry 1991, 48, 1097–1106. [Google Scholar] [CrossRef] [PubMed]
- Pisoschi, A.M.; Pop, A. The role of antioxidants in the chemistry of oxidative stress: A review. Eur. J. Med. Chem. 2015, 97, 55–74. [Google Scholar] [CrossRef]
- Parikh, V.; Khan, M.M.; Mahadik, S.P. Differential effects of antipsychotics on expression of antioxidant enzymes and membrane lipid peroxidation in rat brain. J. Psychiatr. Res. 2003, 37, 43–51. [Google Scholar] [CrossRef]
- Zhang, M.; Zhao, Z.; He, L.; Wan, C. A meta-analysis of oxidative stress markers in schizophrenia. Sci. China Life Sci. 2010, 53, 112–124. [Google Scholar] [CrossRef]
- Fraguas, D.; Díaz-Caneja, C.M.; Ayora, M.; Hernández-Álvarez, F.; Rodríguez-Quiroga, A.; Recio, S.; Leza, J.C.; Arango, C. Oxidative Stress and Inflammation in First-Episode Psychosis: A Systematic Review and Meta-analysis. Schizophr. Bull. 2019, 45, 742–751. [Google Scholar] [CrossRef]
- Jiménez-Fernández, S.; Gurpegui, M.; Garrote-Rojas, D.; Gutiérrez-Rojas, L.; Carretero, M.D.; Correll, C.U. Oxidative stress parameters and antioxidants in patients with bipolar disorder: Results from a meta-analysis comparing patients, including stratification by polarity and euthymic status, with healthy controls. Bipolar Disord. 2021, 23, 117–129. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Fernández, S.; Gurpegui, M.; Díaz-Atienza, F.; Pérez-Costillas, L.; Gerstenberg, M.; Correll, C.U. Oxidative stress and antioxidant parameters in patients with major depressive disorder compared to healthy controls before and after antidepressant treatment: Results from a meta-analysis. J. Clin. Psychiatry 2015, 76, 1658–1667. [Google Scholar] [CrossRef]
- Maia, A.; Oliveira, J.; Lajnef, M.; Mallet, L.; Tamouza, R.; Leboyer, M.; Oliveira-Maia, A.J. Oxidative and nitrosative stress markers in obsessive-compulsive disorder: A systematic review and meta-analysis. Acta Psychiatr. Scand. 2019, 139, 420–433. [Google Scholar] [CrossRef] [PubMed]
- Solmi, M.; Veronese, N.; Luchini, C.; Manzato, E.; Sergi, G.; Favaro, A.; Santonastaso, P.; Correll, C.U. Oxidative Stress and Antioxidant Levels in Patients with Anorexia Nervosa after Oral Re-alimentation: A Systematic Review and Exploratory Meta-analysis. Eur. Eat. Disord. Rev. 2016, 24, 101–105. [Google Scholar] [CrossRef]
- Ermakov, E.A.; Dmitrieva, E.M.; Parshukova, D.A.; Kazantseva, D.V.; Vasilieva, A.R.; Smirnova, L.P. Oxidative Stress-Related Mechanisms in Schizophrenia Pathogenesis and New Treatment Perspectives. Oxidative Med. Cell. Longev. 2021, 2021, 8881770. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, P.V.; Dean, O.; Andreazza, A.C.; Berk, M.; Kapczinski, F. Antioxidant treatments for schizophrenia. Cochrane Database Syst. Rev. 2016, 2, Cd008919. [Google Scholar] [CrossRef]
- Rossetti, A.C.; Paladini, M.S.; Riva, M.A.; Molteni, R. Oxidation-reduction mechanisms in psychiatric disorders: A novel target for pharmacological intervention. Pharmacol. Ther. 2020, 210, 107520. [Google Scholar] [CrossRef]
- Lee, A.R.Y.B.; Tariq, A.; Lau, G.; Tok, N.W.K.; Tam, W.W.S.; Ho, C.S.H. Vitamin E, Alpha-Tocopherol, and Its Effects on Depression and Anxiety: A Systematic Review and Meta-Analysis. Nutrients 2022, 14, 656. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, Z.; Du, Y.; Shi, S.; Cheng, Y. Antioxidant interventions in autism spectrum disorders: A meta-analysis. Prog. Neuropsychopharmacol. Biol. Psychiatry 2022, 113, 110476. [Google Scholar] [CrossRef]
- Lu, Z.; Pu, C.; Zhang, Y.; Sun, Y.; Liao, Y.; Kang, Z.; Feng, X.; Yue, W. Oxidative Stress and Psychiatric Disorders: Evidence from the Bidirectional Mendelian Randomization Study. Antioxidants 2022, 11, 1386. [Google Scholar] [CrossRef]
- Gil Martínez, V.; Avedillo Salas, A.; Santander Ballestín, S. Vitamin Supplementation and Dementia: A Systematic Review. Nutrients 2022, 14, 1033. [Google Scholar] [CrossRef] [PubMed]
- Skrivankova, V.W.; Richmond, R.C.; Woolf, B.A.; Yarmolinsky, J.; Davies, N.M.; Swanson, S.A.; VanderWeele, T.J.; Higgins, J.P.; Timpson, N.J.; Dimou, N.; et al. Strengthening the Reporting of Observational Studies in Epidemiology Using Mendelian Randomization: The STROBE-MR Statement. JAMA 2021, 326, 1614–1621. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Cheng, Y.; Li, M.; Zhang, Z.; Li, T.; Luo, X.J. Genome-wide Mendelian randomization identifies actionable novel drug targets for psychiatric disorders. Neuropsychopharmacology 2023, 48, 270–280. [Google Scholar] [CrossRef]
- Bowden, J.; Del Greco, M.F.; Minelli, C.; Davey Smith, G.; Sheehan, N.A.; Thompson, J.R. Assessing the suitability of summary data for two-sample Mendelian randomization analyses using MR-Egger regression: The role of the I2 statistic. Int. J. Epidemiol. 2016, 45, 1961–1974. [Google Scholar] [CrossRef] [PubMed]
- Trubetskoy, V.; Pardiñas, A.F.; Qi, T.; Panagiotaropoulou, G.; Awasthi, S.; Bigdeli, T.B.; Bryois, J.; Chen, C.-Y.; Dennison, C.A.; Hall, L.S.; et al. Mapping genomic loci implicates genes and synaptic biology in schizophrenia. Nature 2022, 604, 502–508. [Google Scholar] [CrossRef] [PubMed]
- Mullins, N.; Forstner, A.J.; O’connell, K.S.; Coombes, B.; Coleman, J.R.I.; Qiao, Z.; Als, T.D.; Bigdeli, T.B.; Børte, S.; Bryois, J.; et al. Genome-wide association study of more than 40,000 bipolar disorder cases provides new insights into the underlying biology. Nat. Genet. 2021, 53, 817–829. [Google Scholar] [CrossRef]
- Howard, D.M.; Adams, M.J.; Clarke, T.-K.; Hafferty, J.D.; Gibson, J.; Shirali, M.; Coleman, J.R.I.; Hagenaars, S.P.; Ward, J.; Wigmore, E.M.; et al. Genome-wide meta-analysis of depression identifies 102 independent variants and highlights the importance of the prefrontal brain regions. Nat. Neurosci. 2019, 22, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Demontis, D.; Walters, R.K.; Martin, J.; Mattheisen, M.; Als, T.D.; Agerbo, E.; Baldursson, G.; Belliveau, R.; Bybjerg-Grauholm, J.; Bækvad-Hansen, M.; et al. Discovery of the first genome-wide significant risk loci for attention deficit/hyperactivity disorder. Nat. Genet. 2019, 51, 63–75. [Google Scholar] [CrossRef]
- Grove, J.; Ripke, S.; Als, T.D.; Mattheisen, M.; Walters, R.K.; Won, H.; Pallesen, J.; Agerbo, E.; Andreassen, O.A.; Anney, R.; et al. Identification of common genetic risk variants for autism spectrum disorder. Nat. Genet. 2019, 51, 431–444. [Google Scholar] [CrossRef]
- (OCGAS) IOCDFGCI-GaOCGAS. Revealing the complex genetic architecture of obsessive-compulsive disorder using meta-analysis. Mol. Psychiatry 2018, 23, 1181–1188. [Google Scholar] [CrossRef]
- Watson, H.J.; Yilmaz, Z.; Thornton, L.M.; Hübel, C.; Coleman, J.R.; Gaspar, H.A.; Bryois, J.; Hinney, A.; Leppä, V.M.; Mattheisen, M.; et al. Genome-wide association study identifies eight risk loci and implicates metabo-psychiatric origins for anorexia nervosa. Nat. Genet. 2019, 51, 1207–1214. [Google Scholar] [CrossRef]
- Velmeshev, D.; Schirmer, L.; Jung, D.; Haeussler, M.; Perez, Y.; Mayer, S.; Bhaduri, A.; Goyal, N.; Rowitch, D.H.; Kriegstein, A.R. Single-cell genomics identifies cell type-specific molecular changes in autism. Science 2019, 364, 685–689. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Gloudemans, M.J.; Rao, A.S.; Ingelsson, E.; Montgomery, S.B. Abundant associations with gene expression complicate GWAS follow-up. Nat. Genet. 2019, 51, 768–769. [Google Scholar] [CrossRef]
- Ma, C.; Li, Y.; Li, X.; Liu, J.; Luo, X.J. Identification of a functional SNP rs7304782 at schizophrenia risk locus 12q24.31 and validation of its association with schiz ophrenia in Chinese populations. Psychiatry Res. 2020, 294, 113491. [Google Scholar] [CrossRef] [PubMed]
- Masana, M.I.; Sumaya, I.C.; Becker-Andre, M.; Dubocovich, M.L. Behavioral characterization and modulation of circadian rhythms by light and melatonin in C3H/HeN mice homozygous for the RORbeta knockout. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 292, R2357–R2367. [Google Scholar] [CrossRef]
- Mansour, H.A.; Talkowski, M.E.; Wood, J.; Chowdari, K.V.; McClain, L.; Prasad, K.; Montrose, D.; Fagiolini, A.; Friedman, E.S.; Allen, M.H.; et al. Association study of 21 circadian genes with bipolar I disorder, schizoaffective disorder, and schizophrenia. Bipolar Disord. 2009, 11, 701–710. [Google Scholar] [CrossRef]
- Lin, R.; Taylor, B.V.; Simpson, S., Jr.; Charlesworth, J.; Ponsonby, A.L.; Pittas, F.; Dwyer, T.; van der Mei, I.A. Novel modulating effects of PKC family genes on the relationship between serum vitamin D and relapse in multiple sclerosis. J. Neurol. Neurosurg. Psychiatry 2014, 85, 399–404. [Google Scholar] [CrossRef]
- Serreze, D.V.; Fleming, S.A.; Chapman, H.D.; Richard, S.D.; Leiter, E.H.; Tisch, R.M. B lymphocytes are critical antigen-presenting cells for the initiation of T cell-mediated autoimmune diabetes in nonobese diabetic mice. J. Immunol. 1998, 161, 3912–3918. [Google Scholar] [CrossRef] [PubMed]
- Pandey, G.N.; Rizavi, H.S.; Ren, X. Protein and mRNA expression of protein kinase C (PKC) in the postmortem brain of bipolar and schizophrenic subjects. J. Psychiatr. Res. 2020, 130, 362–371. [Google Scholar] [CrossRef]
- Leinonen, H.; Koivisto, H.; Lipponen, H.-R.; Matilainen, A.; Salo, A.M.; Dimova, E.Y.; Hämäläinen, E.; Stavén, S.; Miettinen, P.; Myllyharju, J.; et al. Null mutation in P4h-tm leads to decreased fear and anxiety and increased social behavior in mice. Neuropharmacology 2019, 153, 63–72. [Google Scholar] [CrossRef]
- Wang, M.; Yan, H.; Tian, X.; Yue, W.; Liu, Y.; Fan, L.; Hu, K.; Sun, Y.; Zhao, Y.; Lou, J.; et al. Neuroimaging and multiomics reveal cross-scale circuit abnormalities in schizophrenia. Nat. Ment. Health 2023, 1, 633–654. [Google Scholar] [CrossRef]
- Farsi, Z.; Nicolella, A.; Simmons, S.K.; Aryal, S.; Shepard, N.; Brenner, K.; Lin, S.; Herzog, L.; Moran, S.P.; Stalnaker, K.J.; et al. Brain-region-specific changes in neurons and glia and dysregulation of dopamine signaling in Grin2a mutant mice. Neuron 2023, 111, 3378–3396.e9. [Google Scholar] [CrossRef] [PubMed]
- Hwang, H.; Szucs, M.J.; Ding, L.J.; Allen, A.; Ren, X.; Haensgen, H.; Gao, F.; Rhim, H.; Andrade, A.; Pan, J.Q.; et al. Neurogranin, Encoded by the Schizophrenia Risk Gene NRGN, Bidirectionally Modulates Synaptic Plasticity via Calmodulin-Dependent Regulation of the Neuronal Phosphoproteome. Biol. Psychiatry 2021, 89, 256–269. [Google Scholar] [CrossRef] [PubMed]
- Moretti, M.; Rodrigues, A.L.S. Functional role of ascorbic acid in the central nervous system: A focus on neurogenic and synaptogenic processes. Nutr. Neurosci. 2022, 25, 2431–2441. [Google Scholar] [CrossRef]
- Wambebe, C.; Sokomba, E. Some behavioural and EEG effects of ascorbic acid in rats. Psychopharmacology 1986, 89, 167–170. [Google Scholar] [CrossRef] [PubMed]
- Binfaré, R.W.; Rosa, A.O.; Lobato, K.R.; Santos, A.R.; Rodrigues, A.L. Ascorbic acid administration produces an antidepressant-like effect: Evidence for the involvement of monoaminergic neurotransmission. Prog. Neuropsychopharmacol. Biol. Psychiatry 2009, 33, 530–540. [Google Scholar] [CrossRef]
- Dai, F.; Yang, J.Y.; Gu, P.F.; Hou, Y.; Wu, C.F. Effect of drug-induced ascorbic acid release in the striatum and the nucleus accumbens in hippocampus-lesioned rats. Brain Res. 2006, 1125, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Gu, P.F.; Wu, C.F.; Yang, J.Y.; Shang, Y.; Hou, Y.; Bi, X.L.; Dai, F. Differential effects of drug-induced ascorbic acid release in the striatum and nucleus accumbens of freely moving rats. Neurosci. Lett. 2006, 399, 79–84. [Google Scholar] [CrossRef]
- Jomova, K.; Raptova, R.; Alomar, S.Y.; Alwasel, S.H.; Nepovimova, E.; Kuca, K.; Valko, M. Reactive oxygen species, toxicity, oxidative stress, and antioxidants: Chronic diseases and aging. Arch. Toxicol. 2023, 97, 2499–2574. [Google Scholar]
- La Torre, M.E.; Villano, I.; Monda, M.; Messina, A.; Cibelli, G.; Valenzano, A.; Pisanelli, D.; Panaro, M.A.; Tartaglia, N.; Ambrosi, A.; et al. Role of Vitamin E and the Orexin System in Neuroprotection. Brain Sci. 2021, 11, 1098. [Google Scholar] [CrossRef]
- Behl, C. Vitamin E protects neurons against oxidative cell death in vitro more effectively than 17-beta estradiol and induces the activity of the transcription factor NF-kappaB. J. Neural Transm. 2000, 107, 393–407. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, S.; Barbanta, A.; Ettinger, U.; Kumari, V. Pineal Abnormalities in Psychosis and Mood Disorders: A Systematic Review. Brain Sci. 2023, 13, 827. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Li, H.; Fu, X.; Cen, M.; Wu, J. Association of the Oxidative Balance Score and Cognitive Function and the Mediating Role of Oxidative Stress: Evidence from the National Health and Nutrition Examination Survey (NHANES) 2011–2014. J. Nutr. 2023, 153, 1974–1983. [Google Scholar] [CrossRef] [PubMed]
- Scapagnini, G.; Davinelli, S.; Drago, F.; De Lorenzo, A.; Oriani, G. Antioxidants as antidepressants: Fact or fiction? CNS Drugs 2012, 26, 477–490. [Google Scholar] [CrossRef] [PubMed]
- Kishi, T.; Sakuma, K.; Hatano, M.; Iwata, N. N-acetylcysteine for schizophrenia: A systematic review and meta-analysis. Psychiatry Clin. Neurosci. 2023, 77, 119–121. [Google Scholar] [CrossRef]
- Skrivankova, V.W.; Richmond, R.C.; Woolf, B.A.R.; Davies, N.M.; Swanson, S.A.; VanderWeele, T.J. Strengthening the Reporting of Observational Studies in Epidemiology using Mendelian Randomisation (STROBE-MR): Explanation and Elaboration. BMJ 2021, 375, n2233. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Targets | Tissues | Psychiatry Disorders | IVs | F | H0 | H1 | H2 | H3 | H4 |
|---|---|---|---|---|---|---|---|---|---|
| OGFOD2 | Cerebellum | Schizophrenia | rs28569885 | 19.45 | 1.11 × 10−3 | 7.69 × 10−2 | 3.83 × 10−5 | 1.74 × 10−3 | 9.20 × 10−1 |
| RORB | Cerebellum | Schizophrenia | rs514209 | 20.03 | 5.20 × 10−5 | 1.52 × 10−2 | 1.76 × 10−5 | 4.16 × 10−3 | 9.81 × 10−1 |
| PRKCB | DLPFC | Bipolar disorder | rs120908 | 54.54 | 6.82 × 10−4 | 6.02 × 10−3 | 1.57 × 10−2 | 1.29 × 10−2 | 9.70 × 10−1 |
| P4HTM | Cerebellum | Major depressive disorder | rs6769821 | 28.01 | 3.11 × 10−6 | 3.37 × 10−7 | 3.74 × 10−1 | 4.00 × 10−2 | 5.86 × 10−1 |
| P4HTM | Cerebellar Hemisphere | Major depressive disorder | rs4384984 | 30.85 | 2.91 × 10−2 | 3.28 × 10−3 | 4.03 × 10−1 | 4.49 × 10−2 | 5.20 × 10−1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, Z.; Yang, Y.; Zhao, G.; Zhang, Y.; Sun, Y.; Liao, Y.; Kang, Z.; Feng, X.; Sun, J.; Yue, W. The Association of Redox Regulatory Drug Target Genes with Psychiatric Disorders: A Mendelian Randomization Study. Antioxidants 2024, 13, 398. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox13040398
Lu Z, Yang Y, Zhao G, Zhang Y, Sun Y, Liao Y, Kang Z, Feng X, Sun J, Yue W. The Association of Redox Regulatory Drug Target Genes with Psychiatric Disorders: A Mendelian Randomization Study. Antioxidants. 2024; 13(4):398. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox13040398
Chicago/Turabian StyleLu, Zhe, Yang Yang, Guorui Zhao, Yuyanan Zhang, Yaoyao Sun, Yundan Liao, Zhewei Kang, Xiaoyang Feng, Junyuan Sun, and Weihua Yue. 2024. "The Association of Redox Regulatory Drug Target Genes with Psychiatric Disorders: A Mendelian Randomization Study" Antioxidants 13, no. 4: 398. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox13040398