GPX1 Localizes to the Nucleus in Prostate Epithelium and its Levels are not Associated with Prostate Cancer Recurrence


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Imaging
2.2. Source of Clinical Samples
2.3. Immunohistochemistry
2.4. VECTRA Quantitative Imaging Analysis
2.5. Data Analysis
3. Results
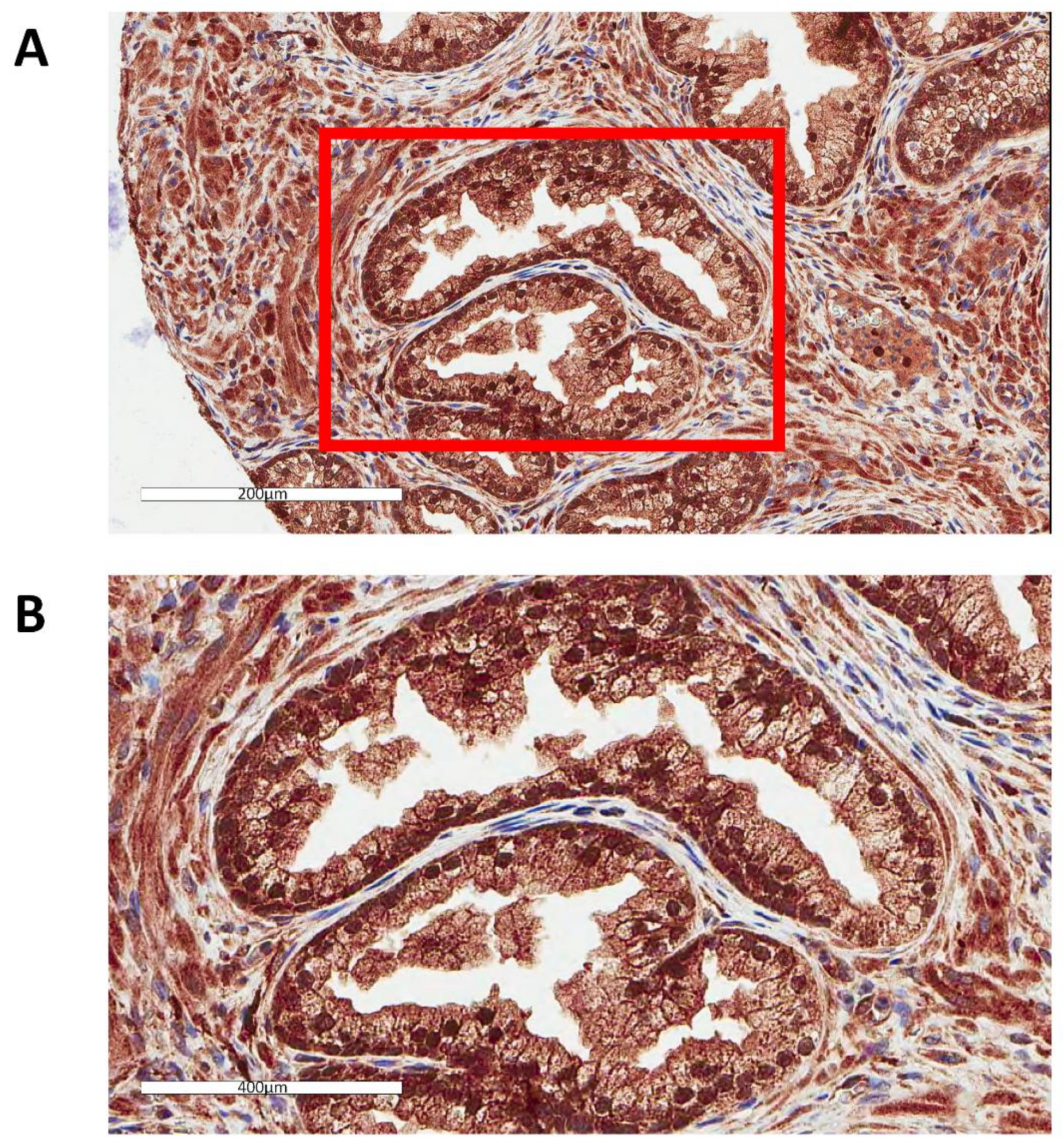
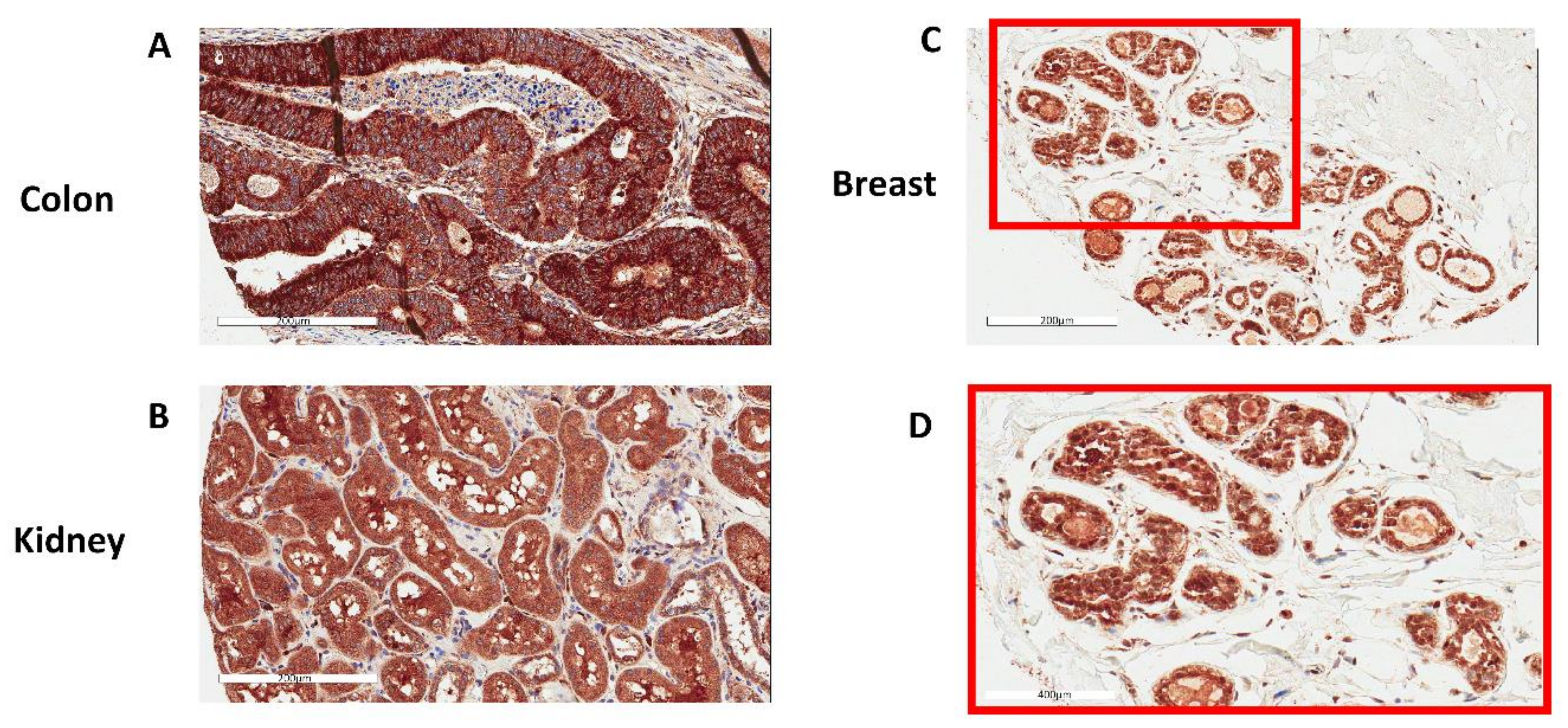
3.1. GPX1 is a Nuclear Protein in Benign Human Prostate Epithelial Cells
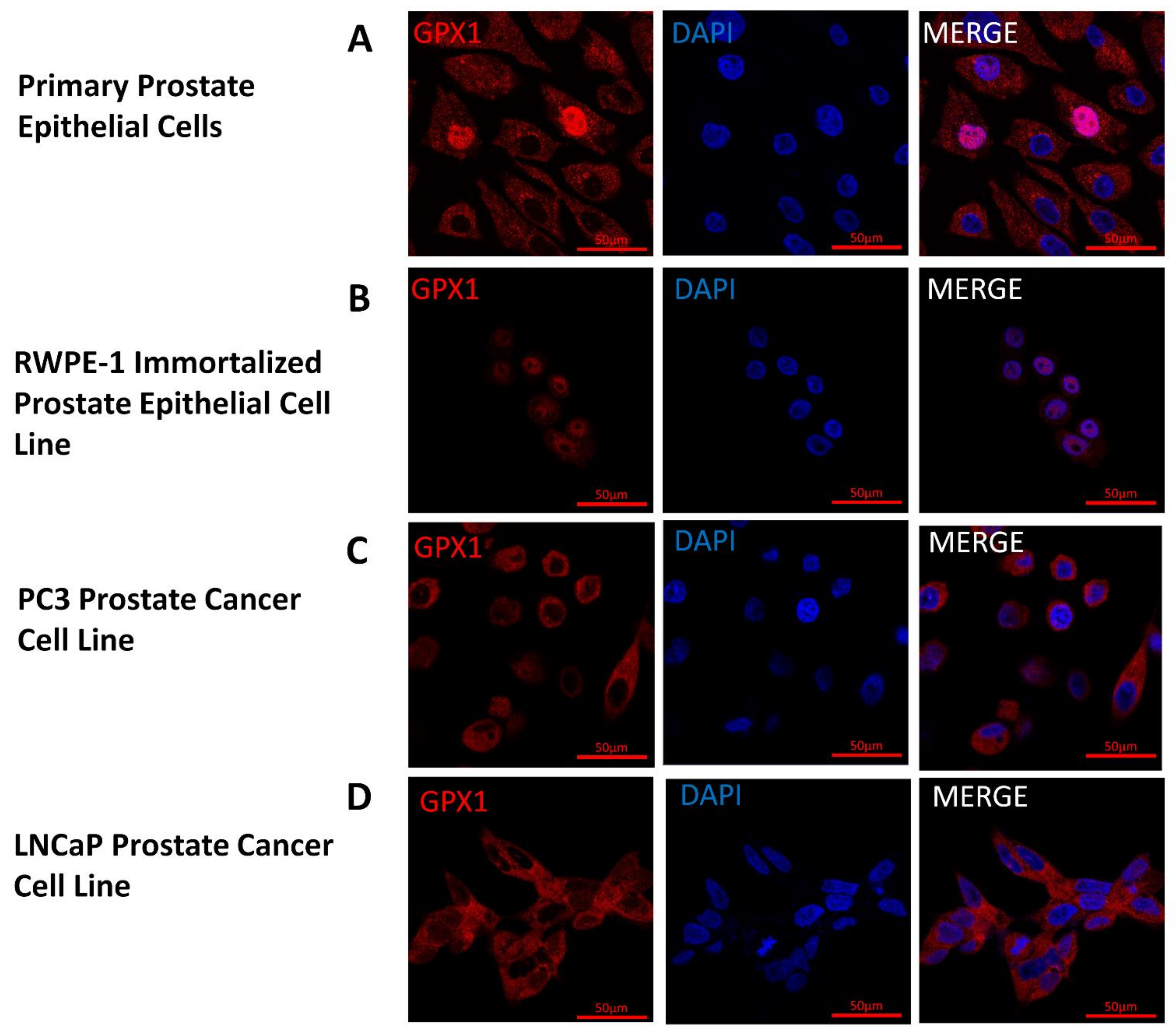
3.2. Nuclear Localization of GPX1 in Immortalized Human Prostate Cells but Not in Tumor-Derived Cell Lines
3.3. Neither Levels nor Cellular Location of GPX1 in Human Prostate Epithelium are Associated with Prostate Cancer Recurrence
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Behne, D.; Kyriakopoulos, A. Mammalian selenium-containing proteins. Annu. Rev. Nutr. 2001, 21, 453–473. [Google Scholar] [CrossRef] [PubMed]
- Labunskyy, V.M.; Hatfield, D.L.; Gladyshev, V.N. Selenoproteins: Molecular pathways and physiological roles. Physiol. Rev. 2014, 94, 739–777. [Google Scholar] [CrossRef] [PubMed]
- Driscoll, D.M.; Copeland, P.R. Mechanism and regulation of selenoprotein synthesis. Annu. Rev. Nutr. 2003, 23, 17–40. [Google Scholar] [CrossRef] [PubMed]
- Brigelius-Flohe, R.; Maiorino, M. Glutathione peroxidases. Biochim. Biophys. Acta 2013, 1830, 3289–3303. [Google Scholar] [CrossRef] [PubMed]
- Mills, G.C. Hemoglobin catabolism I. Glutathione peroxidase, an erythrocyte enzyme which protects hemoglobin from oxidative breakdown. J. Biol. Chem. 1957, 229, 189–197. [Google Scholar] [PubMed]
- Lubos, E.; Loscalzo, J.; Handy, D.E. Glutathione peroxidase-1 in health and disease: From molecular mechanisms to therapeutic opportunities. Antioxid. Redox Signal. 2011, 15, 1957–1997. [Google Scholar] [CrossRef] [PubMed]
- Ho, Y.S.; Magnenat, J.L.; Bronson, R.T.; Cao, J.; Gargano, M.; Sugawara, M.; Funk, C.D. Mice deficient in cellular glutathione peroxidase develop normally and show no increased sensitivity to hyperoxia. J. Biol. Chem. 1997, 272, 16644–16651. [Google Scholar] [CrossRef] [PubMed]
- Vinceti, M.; Filippini, T.; Del Giovane, C.; Dennert, G.; Zwahlen, M.; Brinkman, M.; Zeegers, M.P.; Horneber, M.; D’Amico, R.; Crespi, C.M. Selenium for preventing cancer. Cochrane Database Syst. Rev. 2018, 1, CD005195. [Google Scholar] [CrossRef] [PubMed]
- Sayehmiri, K.; Azami, M.; Mohammadi, Y.; Soleymani, A.; Tardeh, Z. The association between Selenium and Prostate Cancer: A Systematic Review and Meta-Analysis. Asian Pac. J. Cancer Prev. 2018, 19, 1431–1437. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.; Yuan, Y.; Vaishnav, A.; Tessel, M.A.; Nonn, L.; van Breemen, R.B. Effects of lycopene on protein expression in human primary prostatic epithelial cells. Cancer Prev. Res. 2013, 6, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Ekoue, D.N.; Ansong, E.; Liu, L.; Macias, V.; Deaton, R.; Lacher, C.; Picklo, M.; Nonn, L.; Gann, P.H.; Kajdacsy-Balla, A.; et al. Correlations of SELENOF and SELENOP genotypes with serum selenium levels and prostate cancer. Prostate 2018, 78, 279–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kajdacsy-Balla, A.; Geynisman, J.M.; Macias, V.; Setty, S.; Nanaji, N.M.; Berman, J.J.; Dobbin, K.; Melamed, J.; Kong, X.; Bosland, M.; et al. Practical aspects of planning, building, and interpreting tissue microarrays: The Cooperative Prostate Cancer Tissue Resource experience. J. Mol. Histol. 2007, 38, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Melamed, J.; Datta, M.W.; Becich, M.J.; Orenstein, J.M.; Dhir, R.; Silver, S.; Fidelia-Lambert, M.; Kadjacsy-Balla, A.; Macias, V.; Patel, A.; et al. The cooperative prostate cancer tissue resource: A specimen and data resource for cancer researchers. Clin. Cancer Res. 2004, 10, 4614–4621. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, D.R.; Kalyana-Sundaram, S.; Mahavisno, V.; Varambally, R.; Yu, J.; Briggs, B.B.; Barrette, T.R.; Anstet, M.J.; Kincead-Beal, C.; Kulkarni, P.; et al. Oncomine 3.0: Genes, pathways, and networks in a collection of 18,000 cancer gene expression profiles. Neoplasia 2007, 9, 166–180. [Google Scholar] [CrossRef] [PubMed]
- Erdem, O.; Eken, A.; Akay, C.; Arsova-Sarafinovska, Z.; Matevska, N.; Suturkova, L.; Erten, K.; Ozgok, Y.; Dimovski, A.; Sayal, A.; et al. Association of GPX1 polymorphism, GPX activity and prostate cancer risk. Hum. Exp. Toxicol. 2012, 31, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Arsova-Sarafinovska, Z.; Matevska, N.; Eken, A.; Petrovski, D.; Banev, S.; Dzikova, S.; Georgiev, V.; Sikole, A.; Erdem, O.; Sayal, A.; et al. Glutathione peroxidase 1 (GPX1) genetic polymorphism, erythrocyte GPX activity, and prostate cancer risk. Int. Urol. Nephrol. 2009, 41, 63. [Google Scholar] [CrossRef] [PubMed]
- Men, T.; Zhang, X.; Yang, J.; Shen, B.; Li, X.; Chen, D.; Wang, J. The rs1050450 C > T polymorphism of GPX1 is associated with the risk of bladder but not prostate cancer: Evidence from a meta-analysis. Tumor Biol. 2014, 35, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Parlaktas, B.S.; Atilgan, D.; Gencten, Y.; Benli, I.; Ozyurt, H.; Uluocak, N.; Erdemir, F. A pilot study of the association of manganese superoxide dismutase and glutathione peroxidase 1 single gene polymorphisms with prostate cancer and serum prostate specific antigen levels. Arch. Med. Sci. 2015, 11, 994–1000. [Google Scholar] [CrossRef] [PubMed]
- Liwei, L.; Wei, Z.; Ruifa, H.; Chunyu, L. Association between genetic variants in glutathione peroxidase 1 gene and risk of prostate cancer: A meta-analysis. Mol. Biol. Rep. 2012, 39, 8615–8619. [Google Scholar] [CrossRef] [PubMed]
- Gerstenberger, J.P.; Bauer, S.R.; Van Blarigan, E.L.; Sosa, E.; Song, X.; Witte, J.S.; Carroll, P.R.; Chan, J.M. Selenoprotein and antioxidant genes and the risk of high-grade prostate cancer and prostate cancer recurrence. Prostate 2015, 75, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Van Blarigan, E.L.; Ma, J.; Kenfield, S.A.; Stampfer, M.J.; Sesso, H.D.; Giovannucci, E.L.; Witte, J.S.; Erdman, J.W., Jr.; Chan, J.M.; Penney, K.L. Plasma antioxidants, genetic variation in SOD2, CAT, GPX1, GPX4, and prostate cancer survival. Cancer Epidemiol. Biomark. Prev. 2014, 23, 1037–1046. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.Y.; Neuhouser, M.L.; Barnett, M.; Hudson, M.; Kristal, A.R.; Thornquist, M.; King, I.B.; Goodman, G.E.; Ambrosone, C.B. Polymorphisms in oxidative stress-related genes are not associated with prostate cancer risk in heavy smokers. Cancer Epidemiol. Biomark. Prev. 2007, 16, 1115–1120. [Google Scholar] [CrossRef] [PubMed]
- Geybels, M.S.; van den Brandt, P.A.; Schouten, L.J.; van Schooten, F.J.; van Breda, S.G.; Rayman, M.P.; Green, F.R.; Verhage, B.A. Selenoprotein gene variants, toenail selenium levels, and risk for advanced prostate cancer. J. Natl. Cancer Inst. 2014, 106, dju003. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.M.; Darke, A.K.; Penney, K.L.; Tangen, C.M.; Goodman, P.J.; Lee, G.S.; Sun, T.; Peisch, S.; Tinianow, A.M.; Rae, J.M.; et al. Selenium- or Vitamin E-Related Gene Variants, Interaction with Supplementation, and Risk of High-Grade Prostate Cancer in SELECT. Cancer Epidemiol. Biomark. Prev. 2016, 25, 1050–1058. [Google Scholar] [CrossRef] [PubMed]
- Kote-Jarai, Z.; Durocher, F.; Edwards, S.M.; Hamoudi, R.; Jackson, R.A.; Ardern-Jones, A.; Murkin, A.; Dearnaley, D.P.; Kirby, R.; Houlston, R.; et al. Association between the GCG polymorphism of the selenium dependent GPX1 gene and the risk of young onset prostate cancer. Prostate Cancer Prostatic Dis. 2002, 5, 189–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshimura, S.; Komatsu, N.; Watanabe, K. Purification and immunohistochemical localization of rat liver glutathione peroxidase. Biochim. Biophys. Acta 1980, 621, 130–137. [Google Scholar] [CrossRef]
- Asayama, K.; Yokota, S.; Dobashi, K.; Hayashibe, H.; Kawaoi, A.; Nakazawa, S. Purification and immunoelectron microscopic localization of cellular glutathione peroxidase in rat hepatocytes: Quantitative analysis by postembedding method. Histochemistry 1994, 102, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Soboll, S.; Grundel, S.; Harris, J.; Kolb-Bachofen, V.; Ketterer, B.; Sies, H. The content of glutathione and glutathione S-transferases and the glutathione peroxidase activity in rat liver nuclei determined by a non-aqueous technique of cell fractionation. Biochem. J. 1995, 311, 889–894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Utsunomiya, H.; Komatsu, N.; Yoshimura, S.; Tsutsumi, Y.; Watanabe, K. Exact ultrastructural localization of glutathione peroxidase in normal rat hepatocytes: Advantages of microwave fixation. J. Histochem. Cytochem. 1991, 39, 1167–1174. [Google Scholar] [CrossRef] [PubMed]
- Oberley, T.D.; Zhong, W.; Szweda, L.I.; Oberley, L.W. Localization of antioxidant enzymes and oxidative damage products in normal and malignant prostate epithelium. Prostate 2000, 44, 144–155. [Google Scholar]
- Oberley, T.D.; Oberley, L.W.; Slattery, A.F.; Elwell, J.H. Immunohistochemical localization of glutathione-S-transferase and glutathione peroxidase in adult Syrian hamster tissues and during kidney development. Am. J. Pathol. 1991, 139, 355–369. [Google Scholar] [PubMed]
- Zhang, X.; Zhang, L.; Zhu, J.H.; Cheng, W.H. Nuclear selenoproteins and genome maintenance. IUBMB Life 2016, 68, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Fried, H.; Kutay, U. Nucleocytoplasmic transport: Taking an inventory. Cell. Mol. Life Sci. 2003, 60, 1659–1688. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Conditional Logistic Regression of Prostate Cancer Recurrence GPX Analysis | ||
|---|---|---|
| Odds Ratio (95% CI) | p-Value 1 | |
| GPX in Whole Cell | ||
| Second Quartile | 0.980 (0.501,1.916) | 0.9519 |
| Third Quartile | 0.671 (0.338,1.332) | 0.2541 |
| Fourth Quartile | 0.876 (0.446,1.724) | 0.7025 |
| GPX in Cytoplasm | ||
| Second Quartile | 0.976 (0.506,1.883) | 0.9423 |
| Third Quartile | 0.946 (0.471,1.903) | 0.8773 |
| Fourth Quartile | 1.076 (0.560,2.067) | 0.8252 |
| GPX in Nucleus | ||
| Second Quartile | 0.617 (0.320,1.188) | 0.1487 |
| Third Quartile | 0.499 (0.240,1.037) | 0.0625 |
| Fourth Quartile | 0.747 (0.386,1.445) | 0.3862 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ekoue, D.N.; Ansong, E.; Hong, L.K.; Nonn, L.; Macias, V.; Deaton, R.; Rupnow, R.; Gann, P.H.; Kajdacsy-Balla, A.; Diamond, A.M. GPX1 Localizes to the Nucleus in Prostate Epithelium and its Levels are not Associated with Prostate Cancer Recurrence. Antioxidants 2018, 7, 167. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox7110167
Ekoue DN, Ansong E, Hong LK, Nonn L, Macias V, Deaton R, Rupnow R, Gann PH, Kajdacsy-Balla A, Diamond AM. GPX1 Localizes to the Nucleus in Prostate Epithelium and its Levels are not Associated with Prostate Cancer Recurrence. Antioxidants. 2018; 7(11):167. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox7110167
Chicago/Turabian StyleEkoue, Dede N., Emmanuel Ansong, Lenny K. Hong, Larisa Nonn, Virgilia Macias, Ryan Deaton, Rawan Rupnow, Peter H. Gann, Andre Kajdacsy-Balla, and Alan M. Diamond. 2018. "GPX1 Localizes to the Nucleus in Prostate Epithelium and its Levels are not Associated with Prostate Cancer Recurrence" Antioxidants 7, no. 11: 167. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox7110167