Can Plant Phenolic Compounds Protect the Skin from Airborne Particulate Matter?


1
Department of Molecular Medicine, School of Medicine, Kyungpook National University, Daegu 41944, Korea
2
BK21 Plus KNU Biomedical Convergence Program, Kyungpook National University, Daegu 41944, Korea
3
Cell and Matrix Research Institute, Kyungpook National University, Daegu 41944, Korea
Antioxidants 2019, 8(9), 379; https://doi.org/10.3390/antiox8090379
Submission received: 19 August 2019
/
Revised: 4 September 2019
/
Accepted: 4 September 2019
/
Published: 6 September 2019
(This article belongs to the Special Issue Antioxidants in Cosmetics)
Abstract
:The skin is directly exposed to the polluted atmospheric environment, and skin diseases, such as atopic dermatitis and acne vulgaris, can be induced or exacerbated by airborne particulate matter (PM). PM can also promote premature skin aging with its accompanying functional and morphological changes. PM-induced skin diseases and premature skin aging are largely mediated by reactive oxygen species (ROS), and the harmful effects of PM may be ameliorated by safe and effective natural antioxidants. Experimental studies have shown that the extracts and phenolic compounds derived from many plants, such as cocoa, green tea, grape, pomegranate, and some marine algae, have antioxidant and anti-inflammatory effects on PM-exposed cells. The phenolic compounds can decrease the levels of ROS in cells and/or enhance cellular antioxidant capacity and, thereby, can attenuate PM-induced oxidative damage to nucleic acids, proteins, and lipids. They also lower the levels of cytokines, chemokines, cell adhesion molecules, prostaglandins, and matrix metalloproteinases implicated in cellular inflammatory responses to PM. Although there is still much research to be done, current studies in this field suggest that plant-derived phenolic compounds may have a protective effect on skin exposed to high levels of air pollution.
1. Introduction
Airborne particulate matter (PM) can damage skin cells by generating excessive reactive oxygen species (ROS) and free radicals through chemical and biological processes [1]. Alleviation or attenuation of these initial processes may help prevent the skin damage associated with air pollution [2]. As such, dermatologists and cosmetologists are engaged in uncovering safe and effective natural antioxidants to treat or prevent PM-induced skin diseases and premature aging [3].
Plant phenolic compounds, such as phenolic acids, flavonoids, stilbenoids, ellagitannins, and phlorotannins, are known to reduce the levels of cellular ROS by scavenging them directly or inhibiting their production [4]. Phenolic compounds can also enhance cells’ antioxidant capacity by stimulating the expression of enzymes involved in oxygen metabolism and xenobiotic detoxification [5,6]. Moreover, they have been shown to suppress multiple signaling pathways linked to inflammatory reactions [7,8].
However, whether and how plant phenolic compounds could protect the skin from PM still remains an open question to be answered with scientific evidence. Keeping this question in mind, we reviewed recent studies that report the protective effects of plant-derived phenolic compounds against oxidative stress in skin cells exposed to PM.
2. PM and Its Harmful Effects
PM is a mixture of solid particles and liquid contaminants and contains inorganic and organic pollutants produced directly or indirectly from a variety of natural, artificial, and industrial sources, at sizes small enough to remain suspended in the atmosphere [9,10]. PM with a diameter of less than 10 µm (PM10) or less than 2.5 µm (PM2.5) can have major impacts on health [11,12]. PM exposure increases the incidence of inflammatory diseases in the lung, heart, blood vessels, and brain [13,14,15,16]. PM can also directly attack the eyes, causing inflammatory diseases [17].
Human skin protects the body from air pollutants, but exposure to pollutants over a prolonged or repeated period can have a serious negative impact on the skin itself [18,19,20]. Patients with imperfect skin barriers are even more susceptible to PM because of increased absorption thereof through the percutaneous tract [21,22,23]. PM can damage the skin barrier itself as well, enhancing the subsequent absorption of pollutants [24].
In a highly polluted atmosphere, disrupted immune function can induce or exacerbate skin diseases such as atopic dermatitis [22,25] and acne vulgaris [26,27]. A continuous inflammatory response due to PM exposure has also been associated with premature skin aging [28,29] and hyperpigmentation [30]. Simultaneous PM and ultraviolet radiation (UV) exposure exerts synergistic negative effects on the skin, associated with photo-aging and cancer [31,32].
3. Toxicity Mechanisms of PM
PM is composed of several different toxic compounds, such as transition metals and polycyclic aromatic hydrocarbons, with each constituent instigating harmful effects through different pathways and mechanisms [11,12,33]. PM can cause oxidative damage to nucleic acids, proteins, and lipids inside cells [34,35]. It also induces inflammatory reactions, disrupting skin homeostasis and barrier function [36,37]. Intracellular ROS are an inevitable byproduct of normal metabolism in oxygen-breathing organisms [38], with the mitochondrial electron transport system and oxygen-dependent metabolic enzymes as the major intracellular sources of ROS [39].
NADPH oxidases (NOX)-1, -2, and -4 are upregulated by PM, contributing to ROS production in cells exposed to PM [40,41,42,43]. PM-induced ROS production is intracellular calcium-dependent [44,45], and dual oxidase (DUOX)-2 which requires intracellular calcium for activation has been shown to regulate ROS production in keratinocytes exposed to PM10 [46] or house dust [47]. The impact of ROS on skin biology can be further increased by the Fenton reaction catalyzed by transition metals contained in PM or by the photochemical, photodynamic reactions mediated by UV in the skin [1,48].
PM can activate signaling pathways involving mitogen-activated protein (MAP) kinases such as extracellular signal-regulated kinase (ERK), c-Jun N-terminal kinase (JNK), and p38 MAP kinase [49,50]. PM also stimulates signaling pathways which lead to the activation of the redox-sensitive transcription factor, nuclear factor (NF)-κB [45,51]. PM has been shown to increase mRNA and protein levels of many inflammatory cytokines, including tumor necrosis factor (TNF)-α, interleukin (IL)-1α, IL-1β, IL-6, IL-8, and IL-18, in cells and in reconstructed human epidermis [50,52,53,54]. PM also induces the expression of chemokines, such as monocyte chemoattractant protein (MCP)-1, and cell adhesion molecules, such as intercellular adhesion molecule (ICAM)-1, and vascular cell adhesion molecule (VCAM)-1 [55,56]. PM has been implicated in the activation of matrix metalloproteinase (MMP)-1,-2, and -9 that are involved in the degradation of extracellular matrix components, including collagen [23,37,50,54,57].
PM has been shown to upregulate the production of prostaglandin (PG) E2, an eicosanoid mediator of inflammation [36,58]. In the prostaglandin synthetic pathway, arachidonic acid is converted to PGG2 by cyclooxygenase (COX)-1 or -2 and then spontaneously to PGH2 [59], and PGH2 conversion to PGE2 is catalyzed by microsomal prostaglandin E2 synthase (mPGES) 1 and 2 and cytosolic prostaglandin E2 synthase (cPGES) [60]. PM increased the expression of COX-2 by facilitating the activation of MAP kinases such as ERK, JNK, and p38 MAP kinase in keratinocytes [36,61]. Cellular mRNA levels of COX-1 and -2 and mPGES-1 and -2 have been shown to be upregulated by PM in keratinocytes [62].
Aryl hydrocarbon receptor (AhR) is known to regulate gene expression associated with air pollution toxicity [18,63]. AhR mediates the PM effects that increase ROS production, NOX activity, COX-2 expression, and PGE2 production and decreases the expression of filaggrin that physically aggregates keratin bundles and thereby enhances skin barrier function in human keratinocytes [36,64].
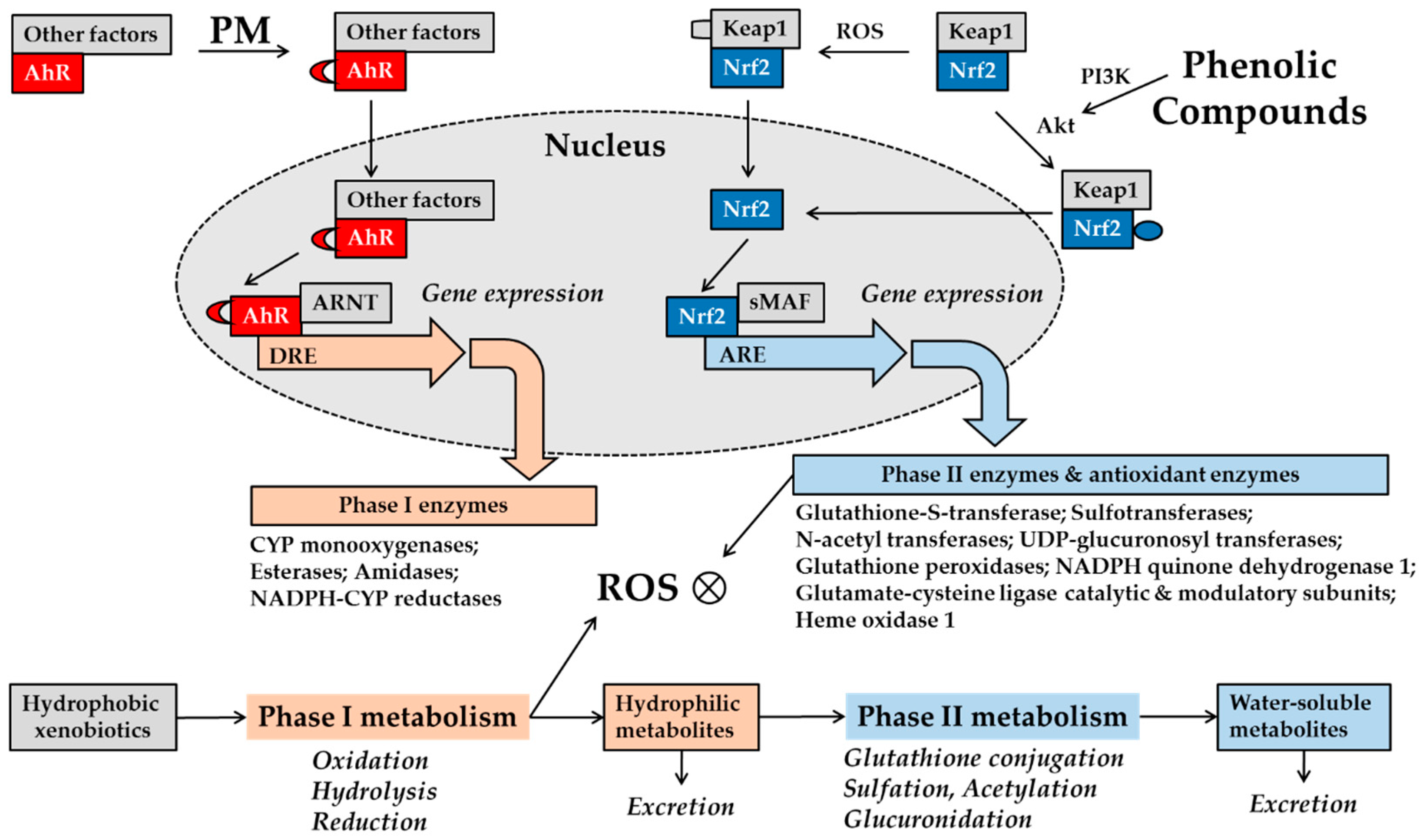
AhR in the cytosol detects xenobiotics, such as drugs and contaminants, and regulates the expression of enzymes involved in phase I metabolism of foreign substances, including cytochrome P450 (CYP)-dependent redox enzymes [65,66]. CYP enzyme activity inevitably generates ROS as byproducts, and xenobiotics themselves can generate ROS by co-opting cellular pathways such as calcium (Ca2+) signaling. Cells also possess a system to detect and defend their oxidative status, which involves nuclear factor erythroid 2-related factor 2 (Nrf2) [67]. The Nrf2 system mainly induces the expression of phase II metabolic enzymes, adding glutathione, glucuronic acid, and other hydrophilic molecules to the foreign substances to enhance their clearance from the cells. The system can also induce antioxidant enzymes involved in the removal of ROS and repair of oxidative damage. The AhR and Nrf2 systems are closely related to one another and thus are compared side by side in Figure 1.
4. Protective Effects of Plant Phenolic Compounds
Recent studies have reported that extracts and phenolic compounds derived from different plants attenuated cellular oxidative stress induced by PM. These studies are listed in Table 1 and discussed briefly in this section.
4.1. Cocoa Extracts and Flavan-3-ols and Derivatives
The flavonoid group of plant phenolic compounds comprises flavonols, flavones, flavans, flavanonols, flavanones, flavan-3-ols (catechins), isoflavone, and anthocyanidins [85]. Flavonoids demonstrate antioxidant, antimicrobial, and other biological activities depending on their chemical structure, and uptake of certain flavonoids has been associated with benefits to human health [86,87].
Cocoa (Theobroma cacao) is a rich source of flavan-3-ols and procyanidins [88,89]. The cocoa products, cacao and chocolate, have been consumed by humans since ancient times, and their health benefits are supported by modern clinical evidence [90]. In 2012, Villarreal-Calderon et al. reported the effects of chocolate administration in mice housed in polluted urban air conditions [81]. Administration of chocolate was associated with a significant downregulation of cytokines such as TNF-α, IL-6, and IL-1β, and alleviation of myocardial inflammation. Chocolate was suggested to confer neuroprotective effects against air pollution in humans [43]. A human study also showed that a one-week oral administration of 4–6 g of high-flavanol cocoa per day reduced erythema formation due to UV exposure [91]. It has been suggested that cocoa-derived phenolic compounds can have positive effects on skin health [92], but their effects on skin exposed to air pollution have yet to be directly addressed.
4.2. Green Tea Extracts and Catechins with Galloyl Residues
Green tea (Camellia sinensis) is an easily accessible source of dietary antioxidants thought to positively impact human health and help prevent disease [4]. Green tea is a rich source of catechins with galloyl residues [93], and green tea extract is frequently used as an active ingredient in cosmetics [94,95].
In a recent study by Seok et al., (−)-epigallocatechin gallate (EGCG) rescued the viability of human epidermal keratinocytes exposed to PM [75]. It also lowered ROS and mRNA expression of NOX-1, NOX-2, TNF-α, IL-1β, IL-6, IL-8, and MMP-1 stimulated by PM. The protein levels of TNF-α, IL-1β, IL-6, and IL-8 were increased by PM, and these changes were mitigated by (−)-EGCG. In another study by Wang et al., (−)-EGCG decreased intracellular ROS and increased cell viability in dermal fibroblasts exposed to PM [76]. (−)-EGCG also restored collagen synthesis while decreasing the expression of MMPs via NF-κB, activator protein 1 (AP-1) and MAP kinase signaling pathways. These results suggest that (−)-EGCG may attenuate the oxidative stress and inflammatory responses of these cells caused by PM.
4.3. Other Flavonoid-Rich Plant Extracts and Other Flavonoids
Camellia japonica leaf extracts contain phenolic compounds such as quercetin, rutin, and other flavonoids [96,97]. C. japonica flower extract directly scavenges ROS in vitro and induces cellular antioxidant enzymes such as superoxide dismutase, catalase, and glutathione peroxidase in human keratinocyte HaCaT cells [98]. C. japonica flower extract inhibited urban dust-induced ROS generation, AhR activation, and MMP-1 expression in human dermal fibroblasts [84]. In an ex vivo test on human skin explants, urban dust increased the levels of lipid peroxidation and MMP-1 expression and decreased the levels of collagen; however, these changes were prevented by pretreatment of the tissue with C. japonica flower extract [84].
Astragalus mongholicus extract and its component, formononetin, rescued the expression of keratins that had been impaired by PM exposure in HaCaT keratinocytes and in a 3D reconstructed human skin model [80]. PM increased the levels of p53, Bcl2-associated X protein (Bax), cleaved caspase-3 and poly (ADP-ribose) polymerase (PARP), all involved in apoptotic cell pathways; these changes were ameliorated by A. mongholicus extract and formononetin, suggesting protective effects against polluting agents.
Eupafolin, a flavone found in Phyla nodiflora, has been shown to inhibit COX-2 expression and PGE2 production in HaCaT cells exposed to PM [70]. It also suppressed PM-induced intracellular generation of ROS, activation of NOX, activation of MAP kinases such as ERK, JNK, and p38, and activation NF-κB. Topical treatment with eupafolin lowered COX-2 expression in vivo in the epidermal keratinocytes of PM-treated mice. Eupafolin nanoparticles, prepared by precipitation using the nanoparticle carriers Eudragit E100 and polyvinyl alcohol, showed enhanced aqueous solubility and skin penetration compared with free eupafolin. Eupafolin nanoparticles also showed superior antioxidant and anti-inflammatory properties than those of free eupafolin in HaCaT cells [71].
Afzelin is a flavonol glycoside (kaempferol 3-O-rhamnoside) found in Thesium chinense that inhibited mRNA expression and protein secretion of IL-1α, but not IL-1β and IL-6 in HaCaT cells exposed to PM [79]. It suppressed PM‑induced intracellular ROS generation and activation of p38 MAP kinase and of the transcription factor AP‑1 components c‑Fos and c‑Jun.
4.4. Grape Extracts and Resveratrol
Grapes (Vitis vinifera) are an easily accessible source of bioactive polyphenols including stilbenoids and proanthocyanidins [89,99,100,101]. Extracts derived from grape seeds and other grape components have shown favorable effects on human skin health and beauty [102,103]. Resveratrol, a stilbenoid compound contained in grapes, has been shown to have antioxidant, antimelanogenic, anti-inflammatory, antimicrobial, anticancer, and cardioprotective effects [104,105].
Resveratrol antioxidant and anti-inflammatory activities have been shown in human fibroblast-like synoviocytes exposed to PM [73]. Resveratrol reduced PM-induced COX-2 expression and PGE2 production, the upregulation thereof having been implicated in many inflammatory diseases. Resveratrol also reduced PM-induced NOX activity, ROS generation, and phosphorylation of Akt, ERK1/2, and p38 MAP kinase and attenuated PM-enhanced NF-κB p65 phosphorylation and NF-κB promoter activity. These results suggest that resveratrol may help combat PM-induced synovial joint inflammation in rheumatoid arthritis.
Choi et al. showed that resveratrol restored survival rates in human epidermal keratinocytes exposed to PM [74]. Resveratrol also alleviated PM10-induced ROS production and IL-6 expression at the mRNA and protein levels. Of interest, resveratrol had no effect on the mRNA levels of TNF-α or IL-8, and even enhanced IL-1β mRNA in PM10-exposed cells. This seemingly peculiar property of resveratrol occurs because the compound can increase or decrease inflammatory reactions and inhibit or promote apoptosis, depending on the cell type and microenvironment [106,107,108,109].
4.5. Pomegranate Extracts and Ellagitannins
Pomegranate (Punica granatum) is rich in polyphenols, the major components thereof being ellagitannins such as ellagic acid and punicalagin (a mixture of α and β anomers) [110,111], which have potent free-radical scavenging properties [112].
In a study by Park et al., pomegranate peel extract lowered the levels of ROS production and the expression of inflammatory cytokines such as TNF-α, IL-1β, and MCP-1 in PM-exposed THP-1 human acute monocytic leukemia cells [55]. The extract also reduced the expression of ICAM-1, but not VCAM-1, in monocytes and attenuated the adhesion of monocytes to endothelial cells in vitro [55]. Punicalagin and ellagic acid showed comparably attenuated PM-induced monocyte adhesion to endothelial cells, but the latter showed significant cytotoxicity.
In a subsequent study, punicalagin rescued the viability of PM-exposed human epidermal keratinocytes and attenuated ROS production therein [75]. It also suppressed PM-induced expression of NOX-1 and NOX-2 involved in ROS production and of TNF-α, IL-1β, IL-6, IL-8, and MMP-1 involved in inflammatory reactions. These results support the anti-inflammatory activity of punicalagin previously demonstrated in different experimental conditions [113,114,115].
4.6. Marine Algae Extracts and Phlorotannins
Marine algae have emerged as a relatively new resource for bioactive cosmetic ingredients [116,117]. Brown algae are unique in containing phlorotannins, such as eckol and dieckol, produced by the polymerization of phloroglucinol [118].
Ecklonia cava is a perennial brown macroalga and is found in the coastal area of East Asia [119]. It is a rich source of phlorotannins with a range of bioactivities [120]. The total phenolic content of E. cava extract is relatively high compared to that of many other marine algae [77]. E. cava extract and its constituents, eckol and dieckol, attenuated cellular lipid peroxidation in HaCaT keratinocytes exposed to PM [77]. Eckol and dieckol also downregulated the mRNA and protein levels of inflammatory cytokines, such as TNF-α, IL-1β, IL-6, and IL-8, in human epidermal keratinocytes exposed to PM [77].
E. cava extract attenuated PGE2 production in HaCaT cells exposed to varying concentrations of PM10 [62]. Dieckol purified from E. cava extract also exhibited similar effects. Both E. cava extract and dieckol suppressed the expression of the COX-1, COX-2, mPGES-1, and mPGES-2 genes involved in PGE2 synthesis. In a 3D reconstructed skin model, dieckol also attenuated the morphological changes induced by PM.
In a study by Zhen et al., eckol has been shown to decrease the levels of ROS and inhibit oxidative damage to lipids, proteins, and DNA in PM-exposed HaCaT keratinocytes [78]. Eckol has been shown to attenuate PM-induced cell apoptosis by modulating MAP kinase signaling pathways [78].
Diphlorethohydroxycarmalol is a phlorotannin found in the brown alga Ishige okamurae [121] and has been shown to attenuate ROS formation, oxidative modifications to DNA, lipids, and proteins, endoplasmic reticulum (ER) stress, autophagy, and apoptosis induced by PM in human HaCaT keratinocytes [83]. It also attenuated oxidative damage and inflammatory reactions in HR-1 mice [83].
5. Mechanisms of Action of Plant Phenolic Compounds
Phenolic compounds in the plant kingdom demonstrate many different chemical structures but often have common chemical and biological activities. Some differences do, however, depend on the structure. A common characteristic is the presentation of phenolic hydroxyl groups that easily donate electrons and confer antioxidant activity. Phenolic compounds can scavenge different kinds of ROS or free radicals and, after these reactions, they become relatively stable radicals that react among themselves, terminating oxidative chain reactions. Many phenolic compounds activate signal transduction systems, such as PI3K- and Akt-dependent pathways that enhance cell survival mechanisms. In this case, the transcription factor Nrf2 is activated through phosphorylation, and the expression of downstream genes increases; as a result, the cell’s antioxidant capacity and resistance to oxidative stress are increased.
Table 2 summarizes studies providing an experimental basis for the chemical and biological properties of the various phenolic compounds. Based on the evidence, plant-derived phenolic compounds are assumed to confer antioxidant and anti-inflammatory activities via multiple mechanisms, including direct scavenging of ROS or reducing ROS generation in cells and enhancing cellular antioxidant capacity via Nrf2 activation. Through these direct and indirect antioxidant actions, the phenolic compounds can suppress the activation of redox-sensitive signaling pathways which lead to the gene expression of cytokines, chemokines, cell adhesion molecules, prostaglandins, and matrix metalloproteinases involved in the inflammatory response of cells exposed to PM.
Phenolic compounds can inhibit the catalytic activity of protein kinases involved in MAP kinase and NF-κB pathways [141]. In addition, they can inhibit the catalytic activity of metabolic enzymes involved in prostaglandin synthesis [142,143]. The catalytic activities of NOXs and MMPs can also be inhibited by certain phenolic compounds [144,145,146]. Therefore, it is considered that certain phenolic compounds can impart anti-inflammatory activity in cells exposed to PM through direct inhibition of target enzyme reactions, in addition to the other mechanisms mentioned above.
It has been reported that several phenolic compounds modulate AhR activity in different directions depending on the individual phenolic compounds tested and the specific experimental conditions, resulting in the increase or decrease of downstream gene expression [147,148,149,150,151]. Since the activation of AhR is a necessary process for the removal of toxic substances, suppressing this process may not necessarily be beneficial for cells. Nonetheless, either activation or inhibition of AhR-mediated gene expression is not considered a common mechanism of action of phenolic compounds, limiting its utility as a strategy for reducing excessive oxidative stress due to PM in cells.
6. Clinical Trials
PM toxicity to the skin has been evaluated indirectly by comparing the skin of residents from two regions with different levels of air pollution [152,153]. No standardized method has been established for validating the antipollution efficacy of cosmetic products on human skin. A study reported the protective efficacy of antioxidant-containing cosmetics on human skin exposed to air pollution [154]. Milani et al. tested a cosmetic serum containing Deschampsia antartica extract, ferulic acid, and vitamin C on the face of 20 women living in a high-pollution area in Rome [154]. The study was conducted over four weeks, and the results showed the product significantly improved skin barrier function as determined by transepidermal water loss (−19%) and enhanced skin brightness as determined by individual typology angles in color (+7%). The product also reduced oxidative stress as determined by squalene peroxide/squalene ratio (−16%) in sebum samples taken from the forehead with a swabbing method.
Due to ethical issues, the direct application of toxic air pollutants to human skin is not permitted, and few studies have applied these types of methods to evaluate the efficacy of cosmetics [155,156]. Giacomelli et al. evaluated the human skin protective effects of a multicomponent powder consisting of grape seed, green tea, and oak wood extracts, against metal deposition in a polluted environment [156]. Each hemi-face of 30 healthy volunteers was treated with a cream formulation of the multicomponent powder or a placebo cream. Following a six-hour exposure in a polluted area, stratum corneum samples of face skin were obtained using the tape-stripping method and analyzed for heavy metal content. There was a significant increase over base line in the metal content of the skin on the hemi-faces treated with the placebo cream, and these changes were not observed on the hemi-faces treated with the multicomponent powder.
Although these studies are promising, there remains a need for the development of standardized human testing methods to yield data that are objective, universal, and comparable to verify the antipollution effects of products used on human skin. Skin protection with plant-derived extracts and phenolic compounds must also be validated through standardized human testing.
7. Conclusions and Future Directions
Recent studies have revealed at least some of the molecular mechanisms that underlie the negative effects of PM on the skin and suggest that oxidative stress and the expression of inflammatory mediators are at the center of this phenomenon. It has also been suggested that the physicochemical and biological processes associated with the adverse effects of PM could be inhibited by plant-derived phenolic compounds with antioxidant and anti-inflammatory effects, by the direct scavenging of ROS or the enhancement of cellular antioxidant capacity via Nrf2 activation. These compounds could be promising active ingredients to address the air pollution that exacerbates skin diseases and promotes premature skin aging, as illustrated in Figure 2. To date, however, most studies have been limited to laboratory experiments using cultured epidermal keratinocytes or dermal fibroblasts, and direct clinical evidence awaits future studies.
The ideal approach for the prevention and alleviation of the adverse effects of PM on the skin is to reduce exposure to air pollution. Experts recommend that outdoor activity be minimized in highly polluted areas and conditions and that individuals protect their skin and body by wearing protective clothing. Dermatological and cosmetological approaches are a secondary approach to attenuate PM-induced deleterious effects on the skin. The composition of PM varies depending on collection place and time but commonly includes inorganic substances such as heavy metals and organic substances such as polycyclic aromatic compounds. It is important to find the right type and concentration of phenolic compounds to protect the skin under different conditions, depending on the composition and concentration of specific harmful PM. It must also be remembered that certain phenolic compounds have prooxidant side effects depending on the circumstances [157].
It is believed that various natural antioxidants such as phenolic compounds, carotenoids, and vitamins can help prevent skin inflammation, premature aging, and pigmentation disorders [105,158,159,160]. The clinical efficacy of the antioxidants will depend not only on their activity potency but also on the reach of their action targets [161,162]. Therefore, it is important to select potent antioxidants that have high skin absorption and target reachability. In addition, there is a need for the development of techniques to enhance the transport and efficacy of selected compounds using advanced formulation techniques such as liposomes, microemulsions, and nanoparticles [71,163,164,165].
Although there remains much research to be done, current studies in this field have produced a consensus that vegetable polyphenols are likely to have positive effects on the skin exposed to air pollution. In the future, it is hoped that this research will be expanded and direct evidence pursued to select plant-derived extracts and phenol compounds that can help protect human skin health and beauty.
Funding
This research was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (NRF-2016R1 D1A1B03932501), Republic of Korea.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mikrut, M.; Regiel-Futyra, A.; Samek, L.; Macyk, W.; Stochel, G.; van Eldik, R. Generation of hydroxyl radicals and singlet oxygen by particulate matter and its inorganic components. Environ. Pollut. 2018, 238, 638–646. [Google Scholar] [CrossRef]
- Rajagopalan, P.; Jain, A.P.; Nanjappa, V.; Patel, K.; Mangalaparthi, K.K.; Babu, N.; Cavusoglu, N.; Roy, N.; Soeur, J.; Breton, L.; et al. Proteome-wide changes in primary skin keratinocytes exposed to diesel particulate extract—A role for antioxidants in skin health. J. Dermatol. Sci. 2018, 91, 239–249. [Google Scholar] [CrossRef]
- Burke, K.E. Mechanisms of aging and development—A new understanding of environmental damage to the skin and prevention with topical antioxidants. Mech. Ageing Dev. 2018, 172, 123–130. [Google Scholar] [CrossRef]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxidative Med. Cell. Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef]
- Scapagnini, G.; Vasto, S.; Abraham, N.G.; Caruso, C.; Zella, D.; Fabio, G. Modulation of Nrf2/ARE pathway by food polyphenols: A nutritional neuroprotective strategy for cognitive and neurodegenerative disorders. Mol. Neurobiol. 2011, 44, 192–201. [Google Scholar] [CrossRef]
- Nabavi, S.F.; Barber, A.J.; Spagnuolo, C.; Russo, G.L.; Daglia, M.; Nabavi, S.M.; Sobarzo-Sanchez, E. Nrf2 as molecular target for polyphenols: A novel therapeutic strategy in diabetic retinopathy. Crit. Rev. Clin. Lab. Sci. 2016, 53, 293–312. [Google Scholar] [CrossRef]
- Sarkar, S.; Mazumder, S.; Saha, S.J.; Bandyopadhyay, U. Management of Inflammation by Natural Polyphenols: A Comprehensive Mechanistic Update. Curr. Med. Chem. 2016, 23, 1657–1695. [Google Scholar] [CrossRef]
- Hussain, T.; Tan, B.; Yin, Y.; Blachier, F.; Tossou, M.C.; Rahu, N. Oxidative Stress and Inflammation: What Polyphenols Can Do for Us? Oxid. Med. Cell. Longev. 2016, 2016, 7432797. [Google Scholar] [CrossRef]
- Sierra-Vargas, M.P.; Teran, L.M. Air pollution: Impact and prevention. Respirology 2012, 17, 1031–1038. [Google Scholar] [CrossRef]
- Jia, Y.Y.; Wang, Q.; Liu, T. Toxicity Research of PM2.5 Compositions In Vitro. Int. J. Environ. Res. Public Health 2017, 14, 232. [Google Scholar] [CrossRef]
- Sacks, J.D.; Stanek, L.W.; Luben, T.J.; Johns, D.O.; Buckley, B.J.; Brown, J.S.; Ross, M. Particulate matter-induced health effects: Who is susceptible? Environ. Health Perspect. 2012, 119, 446–454. [Google Scholar] [CrossRef]
- Anderson, J.O.; Thundiyil, J.G.; Stolbach, A. Clearing the air: A review of the effects of particulate matter air pollution on human health. J. Med. Toxicol. 2012, 8, 166–175. [Google Scholar] [CrossRef]
- Badyda, A.J.; Grellier, J.; Dabrowiecki, P. Ambient PM2.5 Exposure and Mortality Due to Lung Cancer and Cardiopulmonary Diseases in Polish Cities. Respir. Treat. Prev. 2017, 944, 9–17. [Google Scholar]
- Guo, L.; Zhu, N.; Guo, Z.; Li, G.K.; Chen, C.; Sang, N.; Yao, Q.C. Particulate matter (PM10) exposure induces endothelial dysfunction and inflammation in rat brain. J. Hazard. Mater. 2012, 213, 28–37. [Google Scholar] [CrossRef]
- Larrieu, S.; Lefranc, A.; Gault, G.; Chatignoux, E.; Couvy, F.; Jouves, B.; Filleul, L. Are the short-term effects of air pollution restricted to cardiorespiratory diseases? Am. J. Epidemiol. 2009, 169, 1201–1208. [Google Scholar] [CrossRef]
- Zanobetti, A.; Schwartz, J.; Samoli, E.; Gryparis, A.; Touloumi, G.; Peacock, J.; Anderson, R.H.; Le Tertre, A.; Bobros, J.; Celko, M.; et al. The temporal pattern of respiratory and heart disease mortality in response to air pollution. Environ. Health Perspect. 2003, 111, 1188–1193. [Google Scholar] [CrossRef]
- Hwang, S.H.; Choi, Y.H.; Paik, H.J.; Wee, W.R.; Kim, M.K.; Kim, D.H. Potential Importance of Ozone in the Association Between Outdoor Air Pollution and Dry Eye Disease in South Korea. JAMA Ophthalmol. 2016, 134, 503–510. [Google Scholar] [CrossRef] [Green Version]
- Mancebo, S.E.; Wang, S.Q. Recognizing the impact of ambient air pollution on skin health. J. Eur. Acad. Dermatol. Venereol. 2015, 29, 2326–2332. [Google Scholar] [CrossRef] [Green Version]
- Puri, P.; Nandar, S.K.; Kathuria, S.; Ramesh, V. Effects of air pollution on the skin: A review. Indian J. Dermatol. Venereol. Leprol. 2017, 83, 415–423. [Google Scholar]
- Parrado, C.; Mercado-Saenz, S.; Perez-Davo, A.; Gilaberte, Y.; Gonzalez, S.; Juarranz, A. Environmental Stressors on Skin Aging. Mechanistic Insights. Front. Pharmacol. 2019, 10, 759. [Google Scholar] [CrossRef]
- Bakke, J.V.; Wieslander, G.; Norback, D.; Moen, B.E. Eczema increases susceptibility to PM10 in office indoor environments. Arch. Environ. Occup. Health 2012, 67, 15–21. [Google Scholar] [CrossRef]
- Ahn, K. The role of air pollutants in atopic dermatitis. J. Allergy Clin. Immunol. 2014, 134, 993–999. [Google Scholar] [CrossRef]
- Jin, S.P.; Li, Z.; Choi, E.K.; Lee, S.; Kim, Y.K.; Seo, E.Y.; Chung, J.H.; Cho, S. Urban particulate matter in air pollution penetrates into the barrier-disrupted skin and produces ROS-dependent cutaneous inflammatory response in vivo. J. Dermatol. Sci. 2018, 91, 175–183. [Google Scholar] [CrossRef]
- Pan, T.L.; Wang, P.W.; Aljuffali, I.A.; Huang, C.T.; Lee, C.W.; Fang, J.Y. The impact of urban particulate pollution on skin barrier function and the subsequent drug absorption. J. Dermatol. Sci. 2015, 78, 51–60. [Google Scholar] [CrossRef]
- Song, S.; Lee, K.; Lee, Y.M.; Lee, J.H.; Il Lee, S.; Yu, S.D.; Paek, D. Acute health effects of urban fine and ultrafine particles on children with atopic dermatitis. Environ. Res. 2011, 111, 394–399. [Google Scholar] [CrossRef]
- Krutmann, J.; Moyal, D.; Liu, W.; Kandahari, S.; Lee, G.S.; Nopadon, N.; Xiang, L.F.; Seite, S. Pollution and acne: Is there a link? Clin. Cosmet. Investig. Dermatol. 2017, 10, 199–204. [Google Scholar] [CrossRef]
- Liu, W.; Pan, X.; Vierkotter, A.; Guo, Q.; Wang, X.; Wang, Q.; Seite, S.; Moyal, D.; Schikowski, T.; Krutmann, J. A Time-Series Study of the Effect of Air Pollution on Outpatient Visits for Acne Vulgaris in Beijing. Skin Pharmacol. Physiol. 2018, 31, 107–113. [Google Scholar] [CrossRef]
- Vierkotter, A.; Schikowski, T.; Ranft, U.; Sugiri, D.; Matsui, M.; Kramer, U.; Krutmann, J. Airborne particle exposure and extrinsic skin aging. J. Investig. Dermatol. 2010, 130, 2719–2726. [Google Scholar] [CrossRef]
- Park, S.Y.; Byun, E.J.; Lee, J.D.; Kim, S.; Kim, H.S. Air Pollution, Autophagy, and Skin Aging: Impact of Particulate Matter (PM10) on Human Dermal Fibroblasts. Int. J. Mol. Sci. 2018, 19, 2727. [Google Scholar] [CrossRef]
- Roberts, W.E. Pollution as a risk factor for the development of melasma and other skin disorders of facial hyperpigmentation is there a case to be made? J. Drugs Dermatol. 2015, 14, 337–341. [Google Scholar]
- Soeur, J.; Belaidi, J.P.; Chollet, C.; Denat, L.; Dimitrov, A.; Jones, C.; Perez, P.; Zanini, M.; Zobiri, O.; Mezzache, S.; et al. Photo-pollution stress in skin: Traces of pollutants (PAH and particulate matter) impair redox homeostasis in keratinocytes exposed to UVA1. J. Dermatol. Sci. 2017, 86, 162–169. [Google Scholar] [CrossRef]
- Datzmann, T.; Markevych, I.; Trautmann, F.; Heinrich, J.; Schmitt, J.; Tesch, F. Outdoor air pollution, green space, and cancer incidence in Saxony: A semi-individual cohort study. BMC Public Health 2018, 18, 715. [Google Scholar] [CrossRef]
- Magnani, N.D.; Muresan, X.M.; Belmonte, G.; Cervellati, F.; Sticozzi, C.; Pecorelli, A.; Miracco, C.; Marchini, T.; Evelson, P.; Valacchi, G. Skin Damage Mechanisms Related to Airborne Particulate Matter Exposure. Toxicol. Sci. 2016, 149, 227–236. [Google Scholar] [CrossRef]
- Piao, M.J.; Ahn, M.J.; Kang, K.A.; Ryu, Y.S.; Hyun, Y.J.; Shilnikova, K.; Zhen, A.X.; Jeong, J.W.; Choi, Y.H.; Kang, H.K.; et al. Particulate matter 2.5 damages skin cells by inducing oxidative stress, subcellular organelle dysfunction, and apoptosis. Arch. Toxicol. 2018, 92, 2077–2091. [Google Scholar] [CrossRef] [Green Version]
- Pecorelli, A.; Woodby, B.; Prieux, R.; Valacchi, G. Involvement of 4-hydroxy-2-nonenal in pollution-induced skin damage. Biofactors 2019. [Google Scholar] [CrossRef]
- Lee, C.W.; Lin, Z.C.; Hu, S.C.; Chiang, Y.C.; Hsu, L.F.; Lin, Y.C.; Lee, I.T.; Tsai, M.H.; Fang, J.Y. Urban particulate matter down-regulates filaggrin via COX2 expression/PGE2 production leading to skin barrier dysfunction. Sci. Rep. 2016, 6, 27995. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.E.; Cho, D.; Park, H.J. Air pollution and skin diseases: Adverse effects of airborne particulate matter on various skin diseases. Life Sci. 2016, 152, 126–134. [Google Scholar] [CrossRef]
- Lushchak, V.I. Free radicals, reactive oxygen species, oxidative stress and its classification. Chem. Biol. Interact. 2014, 224, 164–175. [Google Scholar] [CrossRef]
- Sarniak, A.; Lipinska, J.; Tytman, K.; Lipinska, S. Endogenous mechanisms of reactive oxygen species (ROS) generation. Postepy Hig. Med. Doswiadczalnej 2016, 70, 1150–1165. [Google Scholar] [CrossRef]
- Cho, D.Y.; Le, W.; Bravo, D.T.; Hwang, P.H.; Illek, B.; Fischer, H.; Nayak, J.V. Air pollutants cause release of hydrogen peroxide and interleukin-8 in a human primary nasal tissue culture model. Int. Forum Allergy Rhinol. 2014, 4, 966–971. [Google Scholar] [CrossRef]
- Bedard, K.; Krause, K.H. The NOX family of ROS-generating NADPH oxidases: Physiology and pathophysiology. Physiol. Rev. 2007, 87, 245–313. [Google Scholar] [CrossRef]
- Lassegue, B.; Griendling, K.K. NADPH oxidases: Functions and pathologies in the vasculature. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 653–661. [Google Scholar] [CrossRef]
- Calderon-Garciduenas, L.; San Juan Chavez, V.; Vacaseydel-Aceves, N.B.; Calderon-Sanchez, R.; Macias-Escobedo, E.; Frias, C.; Giacometto, M.; Velasquez, L.; Felix-Villarreal, R.; Martin, J.D.; et al. Chocolate, Air Pollution and Children’s Neuroprotection: What Cognition Tools should be at Hand to Evaluate Interventions? Front. Pharmacol. 2016, 7, 232. [Google Scholar] [CrossRef]
- Lee, D.U.; Ji, M.J.; Kang, J.Y.; Kyung, S.Y.; Hong, J.H. Dust particles-induced intracellular Ca2+ signaling and reactive Oxygen species in lung fibroblast cell line MRC5. Korean J. Physiol. Pharmacol. 2017, 21, 327–334. [Google Scholar] [CrossRef]
- Donaldson, K.; Stone, V.; Borm, P.J.A.; Jimenez, L.A.; Gilmour, P.S.; Schins, R.P.F.; Knaapen, A.M.; Rahman, I.; Faux, S.P.; Brown, D.M.; et al. Oxidative stress and calcium signaling in the adverse effects of environmental particles (PM10). Free Radic. Biol. Med. 2003, 34, 1369–1382. [Google Scholar] [CrossRef]
- Seok, J.K.; Cho, M.A.; Ha, J.W.; Boo, Y.C. Role of Dual Oxidase 2 in Reactive Oxygen Species Production Induced by Airborne Particulate Matter PM10 in Human Epidermal Keratinocytes. J. Soc. Cosmet. Sci. Korea 2018, 45, 57–67. [Google Scholar]
- Ko, E.; Choi, H.; Park, K.N.; Park, J.Y.; Lee, T.R.; Shin, D.W.; Bae, Y.S. Dual oxidase 2 is essential for house dust mite-induced pro-inflammatory cytokine production in human keratinocytes. Exp. Dermatol. 2015, 24, 936–941. [Google Scholar] [CrossRef]
- Dumax-Vorzet, A.F.; Tate, M.; Walmsley, R.; Elder, R.H.; Povey, A.C. Cytotoxicity and genotoxicity of urban particulate matter in mammalian cells. Mutagenesis 2015, 30, 621–633. [Google Scholar] [CrossRef] [Green Version]
- Xiao, X.; Wang, R.; Cao, L.; Shen, Z.X.; Cao, Y.X. The Role of MAPK Pathways in Airborne Fine Particulate Matter-Induced Upregulation of Endothelin Receptors in Rat Basilar Arteries. Toxicol. Sci. 2016, 149, 213–226. [Google Scholar] [CrossRef]
- Kim, M.; Kim, J.H.; Jeong, G.J.; Park, K.Y.; Lee, M.K.; Seo, S.J. Particulate matter induces pro-inflammatory cytokines via phosphorylation of p38 MAPK possibly leading to dermal inflammaging. Exp. Dermatol. 2019, 28, 809–815. [Google Scholar] [CrossRef]
- Ryu, Y.S.; Kang, K.A.; Piao, M.J.; Ahn, M.J.; Yi, J.M.; Hyun, Y.M.; Kim, S.H.; Ko, M.K.; Park, C.O.; Hyun, J.W. Particulate matter induces inflammatory cytokine production via activation of NFkappaB by TLR5-NOX4-ROS signaling in human skin keratinocyte and mouse skin. Redox Biol. 2019, 21, 101080. [Google Scholar] [CrossRef]
- Romani, A.; Cervellati, C.; Muresan, X.M.; Belmonte, G.; Pecorelli, A.; Cervellati, F.; Benedusi, M.; Evelson, P.; Valacchi, G. Keratinocytes oxidative damage mechanisms related to airbone particle matter exposure. Mech. Ageing Dev. 2018, 172, 86–95. [Google Scholar] [CrossRef]
- Verdin, A.; Cazier, F.; Fitoussi, R.; Blanchet, N.; Vie, K.; Courcot, D.; Momas, I.; Seta, N.; Achard, S. An in vitro model to evaluate the impact of environmental fine particles (PM0.3-2.5) on skin damage. Toxicol. Lett. 2019, 305, 94–102. [Google Scholar] [CrossRef]
- Lecas, S.; Boursier, E.; Fitoussi, R.; Vie, K.; Momas, I.; Seta, N.; Achard, S. In vitro model adapted to the study of skin ageing induced by air pollution. Toxicol. Lett. 2016, 259, 60–68. [Google Scholar] [CrossRef]
- Park, S.; Seok, J.K.; Kwak, J.Y.; Suh, H.J.; Kim, Y.M.; Boo, Y.C. Anti-Inflammatory Effects of Pomegranate Peel Extract in THP-1 Cells Exposed to Particulate Matter PM10. Evid. Based Complement. Altern. Med. 2016, 2016, 6836080. [Google Scholar] [CrossRef]
- Montiel-Davalos, A.; Silva Sanchez, G.J.; Huerta-Garcia, E.; Rueda-Romero, C.; Soca Chafre, G.; Mitre-Aguilar, I.B.; Alfaro-Moreno, E.; Pedraza-Chaverri, J.; Lopez-Marure, R. Curcumin inhibits activation induced by urban particulate material or titanium dioxide nanoparticles in primary human endothelial cells. PLoS ONE 2017, 12, e0188169. [Google Scholar] [CrossRef]
- Morales-Barcenas, R.; Chirino, Y.I.; Sanchez-Perez, Y.; Osornio-Vargas, A.R.; Melendez-Zajgla, J.; Rosas, I.; Garcia-Cuellar, C.M. Particulate matter (PM10) induces metalloprotease activity and invasion in airway epithelial cells. Toxicol. Lett. 2015, 237, 167–173. [Google Scholar] [CrossRef]
- Fernando, I.P.S.; Jayawardena, T.U.; Sanjeewa, K.K.A.; Wang, L.; Jeon, Y.J.; Lee, W.W. Anti-inflammatory potential of alginic acid from Sargassum horneri against urban aerosol-induced inflammatory responses in keratinocytes and macrophages. Ecotoxicol. Environ. Saf. 2018, 160, 24–31. [Google Scholar] [CrossRef]
- Ferrer, M.D.; Busquests-Cortes, C.; Capo, X.; Tejada, S.; Tur, J.A.; Pons, A.; Sureda, A. Cyclooxygenase-2 inhibitors as a therapeutic target in inflammatory diseases. Curr. Med. Chem. 2018. [Google Scholar] [CrossRef]
- Hara, S. Prostaglandin terminal synthases as novel therapeutic targets. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2017, 93, 703–723. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Chen, M.K.; Li, K.; Hu, C.; Lu, M.H.; Jie, S.T. Eupafolin nanoparticle improves acute renal injury induced by LPS through inhibiting ROS and inflammation. Biomed. Pharmacother. 2017, 85, 704–711. [Google Scholar] [CrossRef]
- Ha, J.W.; Song, H.; Hong, S.S.; Boo, Y.C. Marine Alga Ecklonia cava Extract and Dieckol Attenuate Prostaglandin E2 Production in HaCaT Keratinocytes Exposed to Airborne Particulate Matter. Antioxidants 2019, 8, 190. [Google Scholar] [CrossRef]
- Hidaka, T.; Ogawa, E.; Kobayashi, E.H.; Suzuki, T.; Funayama, R.; Nagashima, T.; Fujimura, T.; Aiba, S.; Nakayama, K.; Okuyama, R.; et al. The aryl hydrocarbon receptor AhR links atopic dermatitis and air pollution via induction of the neurotrophic factor artemin. Nat. Immunol. 2017, 18, 64–73. [Google Scholar] [CrossRef]
- Yokota, M.; Yahagi, S.; Masaki, H. Ethyl 2,4-dicarboethoxy pantothenate, a derivative of pantothenic acid, prevents cellular damage initiated by environmental pollutants through Nrf2 activation. J. Dermatol. Sci. 2018, 92, 162–171. [Google Scholar] [CrossRef] [Green Version]
- Dietrich, C. Antioxidant Functions of the Aryl Hydrocarbon Receptor. Stem. Cells Int. 2016, 2016, 7943495. [Google Scholar] [CrossRef]
- Bock, K.W.; Kohle, C. The mammalian aryl hydrocarbon (Ah) receptor: From mediator of dioxin toxicity toward physiological functions in skin and liver. Biol. Chem. 2009, 390, 1225–1235. [Google Scholar] [CrossRef]
- Yamamoto, M.; Kensler, T.W.; Motohashi, H. The KEAP1-NRF2 System: A Thiol-Based Sensor-Effector Apparatus for Maintaining Redox Homeostasis. Physiol. Rev. 2018, 98, 1169–1203. [Google Scholar] [CrossRef]
- Croom, E. Metabolism of xenobiotics of human environments. Prog. Mol. Biol. Transl. Sci. 2012, 112, 31–88. [Google Scholar]
- McMillan, B.J.; Bradfield, C.A. The aryl hydrocarbon receptor sans xenobiotics: Endogenous function in genetic model systems. Mol. Pharmacol. 2007, 72, 487–498. [Google Scholar] [CrossRef]
- Lee, C.W.; Lin, Z.C.; Hsu, L.F.; Fang, J.Y.; Chiang, Y.C.; Tsai, M.H.; Lee, M.H.; Li, S.Y.; Hu, S.C.; Lee, I.T.; et al. Eupafolin ameliorates COX-2 expression and PGE2 production in particulate pollutants-exposed human keratinocytes through ROS/MAPKs pathways. J. Ethnopharmacol. 2016, 189, 300–309. [Google Scholar] [CrossRef]
- Lin, Z.C.; Lee, C.W.; Tsai, M.H.; Ko, H.H.; Fang, J.Y.; Chiang, Y.C.; Liang, C.J.; Hsu, L.F.; Hu, S.C.; Yen, F.L. Eupafolin nanoparticles protect HaCaT keratinocytes from particulate matter-induced inflammation and oxidative stress. Int. J. Nanomed. 2016, 11, 3907–3926. [Google Scholar]
- Huang, P.H.; Tseng, C.H.; Lin, C.Y.; Lee, C.W.; Yen, F.L. Preparation, characterizations and anti-pollutant activity of 7,3′,4′-trihydroxyisoflavone nanoparticles in particulate matter-induced HaCaT keratinocytes. Int. J. Nanomed. 2018, 13, 3279–3293. [Google Scholar] [CrossRef]
- Tsai, M.H.; Hsu, L.F.; Lee, C.W.; Chiang, Y.C.; Lee, M.H.; How, J.M.; Wu, C.M.; Huang, C.L.; Lee, I.T. Resveratrol inhibits urban particulate matter-induced COX-2/PGE2 release in human fibroblast-like synoviocytes via the inhibition of activation of NADPH oxidase/ROS/NF-kappaB. Int. J. Biochem. Cell Biol. 2017, 88, 113–123. [Google Scholar] [CrossRef]
- Choi, M.A.; Seok, J.K.; Lee, J.-W.; Lee, S.Y.; Kim, Y.M.; Boo, Y.C. Effects of Resveratrol and Resveratryl Triacetate on The Inflammatory Responses of Human Epidermal Keratinocytes Exposed to Airborne Particulate Matter PM10. J. Soc. Cosmet. Sci. Korea 2018, 44, 249–258. [Google Scholar]
- Seok, J.K.; Lee, J.W.; Kim, Y.M.; Boo, Y.C. Punicalagin and (-)-Epigallocatechin-3-Gallate Rescue Cell Viability and Attenuate Inflammatory Responses of Human Epidermal Keratinocytes Exposed to Airborne Particulate Matter PM10. Skin Pharmacol. Physiol. 2018, 31, 134–143. [Google Scholar] [CrossRef]
- Wang, L.; Lee, W.; Cui, Y.R.; Ahn, G.; Jeon, Y.J. Protective effect of green tea catechin against urban fine dust particle-induced skin aging by regulation of NF-kappaB, AP-1, and MAPKs signaling pathways. Environ. Pollut. 2019, 252, 1318–1324. [Google Scholar] [CrossRef]
- Lee, J.W.; Seok, J.K.; Boo, Y.C. Ecklonia cava Extract and Dieckol Attenuate Cellular Lipid Peroxidation in Keratinocytes Exposed to PM10. Evid. Based Complement. Altern. Med. 2018, 2018, 8248323. [Google Scholar] [CrossRef]
- Zhen, A.X.; Hyun, Y.J.; Piao, M.J.; Fernando, P.; Kang, K.A.; Ahn, M.J.; Yi, J.M.; Kang, H.K.; Koh, Y.S.; Lee, N.H.; et al. Eckol Inhibits Particulate Matter 2.5-Induced Skin Keratinocyte Damage via MAPK Signaling Pathway. Mar. Drugs 2019, 17, 444. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, M.; Kim, J.M.; Lee, M.K.; Seo, S.J.; Park, K.Y. Afzelin suppresses proinflammatory responses in particulate matter-exposed human keratinocytes. Int. J. Mol. Med. 2019, 43, 2516–2522. [Google Scholar] [CrossRef]
- Nguyen, L.T.H.; Nguyen, U.T.; Kim, Y.H.; Shin, H.M.; Yang, I.J. Astragali Radix and its compound formononetin ameliorate diesel particulate matter-induced skin barrier disruption by regulation of keratinocyte proliferation and apoptosis. J. Ethnopharmacol. 2019, 228, 132–141. [Google Scholar] [CrossRef]
- Villarreal-Calderon, R.; Reed, W.; Palacios-Moreno, J.; Keefe, S.; Herritt, L.; Brooks, D.; Torres-Jardon, R.; Calderon-Garciduenas, L. Urban air pollution produces up-regulation of myocardial inflammatory genes and dark chocolate provides cardioprotection. Exp. Toxicol. Pathol. 2012, 64, 297–306. [Google Scholar] [CrossRef] [Green Version]
- Saputri, R.K.; Setiawan, B.; Nugrahenny, D.; Kania, N.; Wahyuni, E.S.; Widodo, M.A. The effects of Eucheuma cottonii on alveolar macrophages and malondialdehyde levels in bronchoalveolar lavage fluid in chronically particulate matter 10 coal dust-exposed rats. Iran. J. Basic Med. Sci. 2014, 17, 541–545. [Google Scholar]
- Zhen, A.X.; Piao, M.J.; Hyun, Y.J.; Kang, K.A.; Madushan Fernando, P.D.S.; Cho, S.J.; Ahn, M.J.; Hyun, J.W. Diphlorethohydroxycarmalol Attenuates Fine Particulate Matter-Induced Subcellular Skin Dysfunction. Mar. Drugs 2019, 17, 95. [Google Scholar] [CrossRef]
- Kim, M.; Son, D.; Shin, S.; Park, D.; Byun, S.; Jung, E. Protective effects of Camellia japonica flower extract against urban air pollutants. BMC Complement. Altern. Med. 2019, 19, 30. [Google Scholar] [CrossRef]
- Kumar, S.; Pandey, A.K. Chemistry and biological activities of flavonoids: An overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef]
- Cazarolli, L.H.; Zanatta, L.; Alberton, E.H.; Figueiredo, M.S.; Folador, P.; Damazio, R.G.; Pizzolatti, M.G.; Silva, F.R. Flavonoids: Prospective drug candidates. Mini Rev. Med. Chem. 2008, 8, 1429–1440. [Google Scholar] [CrossRef]
- Terahara, N. Flavonoids in foods: A review. Nat. Prod. Commun. 2015, 10, 521–528. [Google Scholar] [CrossRef]
- Wollgast, J.; Anklam, E. Review on polyphenols in Theobroma cacao: Changes in composition during the manufacture of chocolate and methodology for identification and quantification. Food Res. Int. 2000, 33, 423–447. [Google Scholar] [CrossRef]
- Cadiz-Gurrea, M.D.; Borras-Linares, I.; Lozano-Sanchez, J.; Joven, J.; Fernandez-Arroyo, S.; Segura-Carretero, A. Cocoa and Grape Seed Byproducts as a Source of Antioxidant and Anti-Inflammatory Proanthocyanidins. Int. J. Mol. Sci. 2017, 18, 376. [Google Scholar] [CrossRef]
- Dillinger, T.L.; Barriga, P.; Escarcega, S.; Jimenez, M.; Lowe, D.S.; Grivetti, L.E. Food of the gods: Cure for humanity? A cultural history of the medicinal and ritual use of chocolate. J. Nutr. 2000, 130, 2057s–2072s. [Google Scholar] [CrossRef]
- Calzavara-Pinton, P.; Calzavara-Pinton, I.; Arisi, M.; Rossi, M.T.; Scapagnini, G.; Davinelli, S.; Venturini, M. Cutaneous Photoprotective Activity of a Short-term Ingestion of High-Flavanol Cocoa: A Nutritional Intervention Study. Photochem. Photobiol. 2019, 95, 1029–1034. [Google Scholar] [CrossRef]
- Choi, K.S.; Kundu, J.K.; Chun, K.S.; Na, H.K.; Surh, Y.J. Rutin inhibits UVB radiation-induced expression of COX-2 and iNOS in hairless mouse skin: p38 MAP kinase and JNK as potential targets. Arch. Biochem. Biophys. 2014, 559, 38–45. [Google Scholar] [CrossRef]
- Ito, H.; Gonthier, M.P.; Manach, C.; Morand, C.; Mennen, L.; Remesy, C.; Scalbert, A. Polyphenol levels in human urine after intake of six different polyphenol-rich beverages. Br. J. Nutr. 2005, 94, 500–509. [Google Scholar] [CrossRef]
- Hsu, S. Green tea and the skin. J. Am. Acad. Dermatol. 2005, 52, 1049–1059. [Google Scholar] [CrossRef]
- Roh, E.; Kim, J.E.; Kwon, J.Y.; Park, J.S.; Bode, A.M.; Dong, Z.; Lee, K.W. Molecular mechanisms of green tea polyphenols with protective effects against skin photoaging. Crit. Rev. Food Sci. Nutr. 2017, 57, 1631–1637. [Google Scholar] [CrossRef]
- Onodera, K.; Hanashiro, K.; Yasumoto, T. Camellianoside, a novel antioxidant glycoside from the leaves of Camellia japonica. Biosci. Biotechnol. Biochem. 2006, 70, 1995–1998. [Google Scholar] [CrossRef]
- Yoon, I.S.; Park, D.H.; Kim, J.E.; Yoo, J.C.; Bae, M.S.; Oh, D.S.; Shim, J.H.; Choi, C.Y.; An, K.W.; Kim, E.I.; et al. Identification of the biologically active constituents of Camellia japonica leaf and anti-hyperuricemic effect in vitro and in vivo. Int. J. Mol. Med. 2017, 39, 1613–1620. [Google Scholar] [CrossRef] [Green Version]
- Piao, M.J.; Yoo, E.S.; Koh, Y.S.; Kang, H.K.; Kim, J.; Kim, Y.J.; Kang, H.H.; Hyun, J.W. Antioxidant effects of the ethanol extract from flower of Camellia japonica via scavenging of reactive oxygen species and induction of antioxidant enzymes. Int. J. Mol. Sci. 2011, 12, 2618–2630. [Google Scholar] [CrossRef]
- Nile, S.H.; Kim, S.H.; Ko, E.Y.; Park, S.W. Polyphenolic contents and antioxidant properties of different grape (V. vinifera, V. labrusca, and V. hybrid) cultivars. BioMed Res. Int. 2013, 2013, 718065. [Google Scholar] [CrossRef]
- Nassiri-Asl, M.; Hosseinzadeh, H. Review of the pharmacological effects of Vitis vinifera (Grape) and its bioactive compounds. Phytother. Res. 2009, 23, 1197–1204. [Google Scholar] [CrossRef]
- Falchi, M.; Bertelli, A.; Lo Scalzo, R.; Morassut, M.; Morelli, R.; Das, S.; Cui, J.; Das, D.K. Comparison of cardioprotective abilities between the flesh and skin of grapes. J. Agric. Food Chem. 2006, 54, 6613–6622. [Google Scholar] [CrossRef]
- Yamakoshi, J.; Otsuka, F.; Sano, A.; Tokutake, S.; Saito, M.; Kikuchi, M.; Kubota, Y. Lightening effect on ultraviolet-induced pigmentation of guinea pig skin by oral administration of a proanthocyanidin-rich extract from grape seeds. Pigment. Cell Res. 2003, 16, 629–638. [Google Scholar] [CrossRef]
- Khanna, S.; Venojarvi, M.; Roy, S.; Sharma, N.; Trikha, P.; Bagchi, D.; Bagchi, M.; Sen, C.K. Dermal wound healing properties of redox-active grape seed proanthocyanidins. Free. Radic. Biol. Med. 2002, 33, 1089–1096. [Google Scholar] [CrossRef]
- Rauf, A.; Imran, M.; Suleria, H.A.R.; Ahmad, B.; Peters, D.G.; Mubarak, M.S. A comprehensive review of the health perspectives of resveratrol. Food Funct. 2017, 8, 4284–4305. [Google Scholar] [CrossRef]
- Boo, Y.C. Human Skin Lightening Efficacy of Resveratrol and Its Analogs: From in Vitro Studies to Cosmetic Applications. Antioxidants 2019, 8, 332. [Google Scholar] [CrossRef]
- Schwager, J.; Richard, N.; Widmer, F.; Raederstorff, D. Resveratrol distinctively modulates the inflammatory profiles of immune and endothelial cells. BMC Complement. Altern. Med. 2017, 17, 309. [Google Scholar] [CrossRef]
- Cignarella, A.; Minici, C.; Bolego, C.; Pinna, C.; Sanvito, P.; Gaion, R.M.; Puglisi, L. Potential pro-inflammatory action of resveratrol in vascular smooth muscle cells from normal and diabetic rats. Nutr. Metab. Cardiovasc. Dis. 2006, 16, 322–329. [Google Scholar] [CrossRef]
- Walker, J.; Schueller, K.; Schaefer, L.M.; Pignitter, M.; Esefelder, L.; Somoza, V. Resveratrol and its metabolites inhibit pro-inflammatory effects of lipopolysaccharides in U-937 macrophages in plasma-representative concentrations. Food Funct. 2014, 5, 74–84. [Google Scholar] [CrossRef]
- Baarine, M.; Thandapilly, S.J.; Louis, X.L.; Mazue, F.; Yu, L.; Delmas, D.; Netticadan, T.; Lizard, G.; Latruffe, N. Pro-apoptotic versus anti-apoptotic properties of dietary resveratrol on tumoral and normal cardiac cells. Genes Nutr. 2011, 6, 161–169. [Google Scholar] [CrossRef] [Green Version]
- Ambigaipalan, P.; de Camargo, A.C.; Shahidi, F. Phenolic Compounds of Pomegranate Byproducts (Outer Skin, Mesocarp, Divider Membrane) and Their Antioxidant Activities. J. Agric. Food Chem. 2016, 64, 6584–6604. [Google Scholar] [CrossRef]
- Singh, R.P.; Chidambara Murthy, K.N.; Jayaprakasha, G.K. Studies on the antioxidant activity of pomegranate (Punica granatum) peel and seed extracts using in vitro models. J. Agric. Food Chem. 2002, 50, 81–86. [Google Scholar] [CrossRef]
- Kulkarni, A.P.; Mahal, H.S.; Kapoor, S.; Aradhya, S.M. In vitro studies on the binding, antioxidant, and cytotoxic actions of punicalagin. J. Agric. Food Chem. 2007, 55, 1491–1500. [Google Scholar] [CrossRef]
- Jean-Gilles, D.; Li, L.; Vaidyanathan, V.G.; King, R.; Cho, B.; Worthen, D.R.; Chichester, C.O., III; Seeram, N.P. Inhibitory effects of polyphenol punicalagin on type-II collagen degradation in vitro and inflammation in vivo. Chem. Biol. Interact. 2013, 205, 90–99. [Google Scholar] [CrossRef]
- Lin, C.C.; Hsu, Y.F.; Lin, T.C. Effects of punicalagin and punicalin on carrageenan-induced inflammation in rats. Am. J. Chin. Med. 1999, 27, 371–376. [Google Scholar] [CrossRef]
- Xu, X.; Yin, P.; Wan, C.; Chong, X.; Liu, M.; Cheng, P.; Chen, J.; Liu, F.; Xu, J. Punicalagin inhibits inflammation in LPS-induced RAW264.7 macrophages via the suppression of TLR4-mediated MAPKs and NF-kappaB activation. Inflammation 2014, 37, 956–965. [Google Scholar] [CrossRef]
- Kiuru, P.; D’Auria, M.V.; Muller, C.D.; Tammela, P.; Vuorela, H.; Yli-Kauhaluoma, J. Exploring marine resources for bioactive compounds. Planta Med. 2014, 80, 1234–1246. [Google Scholar] [CrossRef]
- Kwak, J.Y.; Seok, J.K.; Suh, H.J.; Choi, Y.H.; Hong, S.S.; Kim, D.S.; Boo, Y.C. Antimelanogenic effects of luteolin 7-sulfate isolated from Phyllospadix iwatensis Makino. Br. J. Dermatol. 2016, 175, 501–511. [Google Scholar] [CrossRef]
- Li, Y.; Qian, Z.J.; Ryu, B.; Lee, S.H.; Kim, M.M.; Kim, S.K. Chemical components and its antioxidant properties in vitro: An edible marine brown alga, Ecklonia cava. Bioorg. Med. Chem 2009, 17, 1963–1973. [Google Scholar] [CrossRef]
- Kim, M.M.; Ta, Q.V.; Mendis, E.; Rajapakse, N.; Jung, W.K.; Byun, H.G.; Jeon, Y.J.; Kim, S.K. Phlorotannins in Ecklonia cava extract inhibit matrix metalloproteinase activity. Life Sci. 2006, 79, 1436–1443. [Google Scholar] [CrossRef]
- Wijesinghe, W.A.; Jeon, Y.J. Exploiting biological activities of brown seaweed Ecklonia cava for potential industrial applications: A review. Int. J. Food Sci. Nutr. 2012, 63, 225–235. [Google Scholar] [CrossRef]
- Heo, S.J.; Kim, J.P.; Jung, W.K.; Lee, N.H.; Kang, H.S.; Jun, E.M.; Park, S.H.; Kang, S.M.; Lee, Y.J.; Park, P.J.; et al. Identification of chemical structure and free radical scavenging activity of diphlorethohydroxycarmalol isolated from a brown alga, Ishige okamurae. J. Microbiol. Biotechnol. 2008, 18, 676–681. [Google Scholar]
- Saint-Cricq De Gaulejac, N.; Provost, C.; Vivas, N. Comparative study of polyphenol scavenging activities assessed by different methods. J. Agric. Food Chem. 1999, 47, 425–431. [Google Scholar] [CrossRef]
- Rodriguez-Ramiro, I.; Ramos, S.; Bravo, L.; Goya, L.; Martin, M.A. Procyanidin B2 induces Nrf2 translocation and glutathione S-transferase P1 expression via ERKs and p38-MAPK pathways and protect human colonic cells against oxidative stress. Eur. J. Nutr. 2012, 51, 881–892. [Google Scholar] [CrossRef]
- Song, J.H.; Lee, H.J.; Kang, K.S. Procyanidin C1 Activates the Nrf2/HO-1 Signaling Pathway to Prevent Glutamate-Induced Apoptotic HT22 Cell Death. Int. J. Mol. Sci. 2019, 20, 142. [Google Scholar] [CrossRef]
- An, Z.; Qi, Y.; Huang, D.; Gu, X.; Tian, Y.; Li, P.; Li, H.; Zhang, Y. EGCG inhibits Cd2+ -induced apoptosis through scavenging ROS rather than chelating Cd2+ in HL-7702 cells. Toxicol. Mech. Methods 2014, 24, 259–267. [Google Scholar] [CrossRef]
- Na, H.K.; Surh, Y.J. Modulation of Nrf2-mediated antioxidant and detoxifying enzyme induction by the green tea polyphenol EGCG. Food Chem. Toxicol. 2008, 46, 1271–1278. [Google Scholar] [CrossRef]
- Mahal, H.S.; Mukherjee, T. Scavenging of reactive oxygen radicals by resveratrol: Antioxidant effect. Res. Chem. Intermed. 2006, 32, 59–71. [Google Scholar] [CrossRef]
- Holthoff, J.H.; Woodling, K.A.; Doerge, D.R.; Burns, S.T.; Hinson, J.A.; Mayeux, P.R. Resveratrol, a dietary polyphenolic phytoalexin, is a functional scavenger of peroxynitrite. Biochem. Pharmacol. 2010, 80, 1260–1265. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Oh, J.; Averilla, J.N.; Kim, H.J.; Kim, J.S.; Kim, J.S. Grape Peel Extract and Resveratrol Inhibit Wrinkle Formation in Mice Model Through Activation of Nrf2/HO-1 Signaling Pathway. J. Food Sci. 2019, 84, 1600–1608. [Google Scholar] [CrossRef]
- Aherne, S.A.; O’Brien, N.M. Mechanism of protection by the flavonoids, quercetin and rutin, against tert-butylhydroperoxide- and menadione-induced DNA single strand breaks in Caco-2 cells. Free Radic. Biol. Med. 2000, 29, 507–514. [Google Scholar] [CrossRef]
- Lopez-Lopez, G.; Moreno, L.; Cogolludo, A.; Galisteo, M.; Ibarra, M.; Duarte, J.; Lodi, F.; Tamargo, J.; Perez-Vizcaino, F. Nitric oxide (NO) scavenging and NO protecting effects of quercetin and their biological significance in vascular smooth muscle. Mol. Pharmacol. 2004, 65, 851–859. [Google Scholar] [CrossRef]
- Schadich, E.; Hlavac, J.; Volna, T.; Varanasi, L.; Hajduch, M.; Dzubak, P. Effects of Ginger Phenylpropanoids and Quercetin on Nrf2-ARE Pathway in Human BJ. Fibroblasts and HaCaT Keratinocytes. Biomed. Res. Int. 2016, 2016, 2173275. [Google Scholar] [CrossRef]
- Sthijns, M.; Schiffers, P.M.; Janssen, G.M.; Lemmens, K.J.A.; Ides, B.; Vangrieken, P.; Bouwman, F.G.; Mariman, E.C.; Pader, I.; Arner, E.S.J.; et al. Rutin protects against H2O2-triggered impaired relaxation of placental arterioles and induces Nrf2-mediated adaptation in Human Umbilical Vein Endothelial Cells exposed to oxidative stress. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 1177–1189. [Google Scholar] [CrossRef]
- Xu, X.; Li, H.; Hou, X.; Li, D.; He, S.; Wan, C.; Yin, P.; Liu, M.; Liu, F.; Xu, J. Punicalagin Induces Nrf2/HO-1 Expression via Upregulation of PI3K/AKT Pathway and Inhibits LPS-Induced Oxidative Stress in RAW264.7 Macrophages. Mediat. Inflamm. 2015, 2015, 380218. [Google Scholar] [CrossRef]
- Priyadarsini, K.I.; Khopde, S.M.; Kumar, S.S.; Mohan, H. Free radical studies of ellagic acid, a natural phenolic antioxidant. J. Agric. Food Chem. 2002, 50, 2200–2206. [Google Scholar] [CrossRef]
- Ding, Y.; Zhang, B.; Zhou, K.; Chen, M.; Wang, M.; Jia, Y.; Song, Y.; Li, Y.; Wen, A. Dietary ellagic acid improves oxidant-induced endothelial dysfunction and atherosclerosis: Role of Nrf2 activation. Int. J. Cardiol. 2014, 175, 508–514. [Google Scholar] [CrossRef]
- Cui, Y.; Amarsanaa, K.; Lee, J.H.; Rhim, J.K.; Kwon, J.M.; Kim, S.H.; Park, J.M.; Jung, S.C.; Eun, S.Y. Neuroprotective mechanisms of dieckol against glutamate toxicity through reactive oxygen species scavenging and nuclear factor-like 2/heme oxygenase-1 pathway. Korean J. Physiol. Pharmacol. 2019, 23, 121–130. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.S.; Lee, B.; Park, K.E.; Utsuki, T.; Shin, T.; Oh, C.W.; Kim, H.R. Dieckol enhances the expression of antioxidant and detoxifying enzymes by the activation of Nrf2-MAPK signalling pathway in HepG2 cells. Food Chem. 2015, 174, 538–546. [Google Scholar] [CrossRef]
- Kang, K.A.; Lee, K.H.; Chae, S.; Zhang, R.; Jung, M.S.; Lee, Y.; Kim, S.Y.; Kim, H.S.; Joo, H.G.; Park, J.W.; et al. Eckol isolated from Ecklonia cava attenuates oxidative stress induced cell damage in lung fibroblast cells. FEBS Lett. 2005, 579, 6295–6304. [Google Scholar] [CrossRef]
- Kim, K.C.; Kang, K.A.; Zhang, R.; Piao, M.J.; Kim, G.Y.; Kang, M.Y.; Lee, S.J.; Lee, N.H.; Surh, Y.J.; Hyun, J.W. Up-regulation of Nrf2-mediated heme oxygenase-1 expression by eckol, a phlorotannin compound, through activation of Erk and PI3K/Akt. Int. J. Biochem. Cell Biol. 2010, 42, 297–305. [Google Scholar] [CrossRef]
- Guerra, B.; Issinger, O.G. Natural Compounds and Derivatives as Ser/Thr Protein Kinase Modulators and Inhibitors. Pharmaceuticals 2019, 12, 4. [Google Scholar] [CrossRef]
- Li, Y.; Frenz, C.M.; Li, Z.; Chen, M.; Wang, Y.; Li, F.; Luo, C.; Sun, J.; Bohlin, L.; Li, Z.; et al. Virtual and in vitro bioassay screening of phytochemical inhibitors from flavonoids and isoflavones against xanthine oxidase and cyclooxygenase-2 for gout treatment. Chem. Biol. Drug Des. 2013, 81, 537–544. [Google Scholar] [CrossRef]
- Desai, S.J.; Prickril, B.; Rasooly, A. Mechanisms of Phytonutrient Modulation of Cyclooxygenase-2 (COX-2) and Inflammation Related to Cancer. Nutr. Cancer 2018, 70, 350–375. [Google Scholar] [CrossRef]
- Yousefian, M.; Shakour, N.; Hosseinzadeh, H.; Hayes, A.W.; Hadizadeh, F.; Karimi, G. The natural phenolic compounds as modulators of NADPH oxidases in hypertension. Phytomedicine 2019, 55, 200–213. [Google Scholar] [CrossRef]
- Thomas, N.V.; Manivasagan, P.; Kim, S.K. Potential matrix metalloproteinase inhibitors from edible marine algae: A review. Environ. Toxicol. Pharmacol. 2014, 37, 1090–1100. [Google Scholar] [CrossRef]
- Crasci, L.; Basile, L.; Panico, A.; Puglia, C.; Bonina, F.P.; Basile, P.M.; Rizza, L.; Guccione, S. Correlating In Vitro Target-Oriented Screening and Docking: Inhibition of Matrix Metalloproteinases Activities by Flavonoids. Planta Med. 2017, 83, 901–911. [Google Scholar] [CrossRef] [Green Version]
- Revel, A.; Raanani, H.; Younglai, E.; Xu, J.; Han, R.; Savouret, J.F.; Casper, R.F. Resveratrol, a natural aryl hydrocarbon receptor antagonist, protects sperm from DNA damage and apoptosis caused by benzo(a)pyrene. Reprod. Toxicol. 2001, 15, 479–486. [Google Scholar] [CrossRef]
- Palermo, C.M.; Westlake, C.A.; Gasiewicz, T.A. Epigallocatechin gallate inhibits aryl hydrocarbon receptor gene transcription through an indirect mechanism involving binding to a 90 kDa heat shock protein. Biochemistry 2005, 44, 5041–5052. [Google Scholar] [CrossRef]
- Potapovich, A.I.; Lulli, D.; Fidanza, P.; Kostyuk, V.A.; De Luca, C.; Pastore, S.; Korkina, L.G. Plant polyphenols differentially modulate inflammatory responses of human keratinocytes by interfering with activation of transcription factors NFkappaB and AhR and EGFR-ERK pathway. Toxicol. Appl. Pharmacol. 2011, 255, 138–149. [Google Scholar] [CrossRef]
- Mohammadi-Bardbori, A.; Bengtsson, J.; Rannug, U.; Rannug, A.; Wincent, E. Quercetin, resveratrol, and curcumin are indirect activators of the aryl hydrocarbon receptor (AHR). Chem. Res. Toxicol. 2012, 25, 1878–1884. [Google Scholar] [CrossRef]
- Vrba, J.; Kren, V.; Vacek, J.; Papouskova, B.; Ulrichova, J. Quercetin, quercetin glycosides and taxifolin differ in their ability to induce AhR activation and CYP1A1 expression in HepG2 cells. Phytother. Res. 2012, 26, 1746–1752. [Google Scholar] [CrossRef]
- Lefebvre, M.A.; Pham, D.M.; Boussouira, B.; Qiu, H.; Ye, C.; Long, X.; Chen, R.; Gu, W.; Laurent, A.; Nguyen, Q.L. Consequences of urban pollution upon skin status. A controlled study in Shanghai area. Int. J. Cosmet. Sci. 2016, 38, 217–223. [Google Scholar] [CrossRef]
- Lefebvre, M.A.; Pham, D.M.; Boussouira, B.; Bernard, D.; Camus, C.; Nguyen, Q.L. Evaluation of the impact of urban pollution on the quality of skin: A multicentre study in Mexico. Int. J. Cosmet. Sci. 2015, 37, 329–338. [Google Scholar] [CrossRef]
- Milani, M.; Hashtroody, B.; Piacentini, M.; Celleno, L. Skin protective effects of an antipollution, antioxidant serum containing Deschampsia antartica extract, ferulic acid and vitamin C: A controlled single-blind, prospective trial in women living in urbanized, high air pollution area. Clin. Cosmet. Investig. Dermatol. 2019, 12, 393–399. [Google Scholar] [CrossRef]
- Curpen, S.; Francois-Newton, V.; Moga, A.; Hosenally, M.; Petkar, G.; Soobramaney, V.; Ruchaia, B.; Lutchmanen Kolanthan, V.; Roheemun, N.; Sokeechand, B.N.; et al. A novel method for evaluating the effect of pollution on the human skin under controlled conditions. Skin Res. Technol. 2019. [Google Scholar] [CrossRef]
- Giacomelli, L.; Togni, S.; Meneghin, M.; Eggenhoffner, R.; Maramaldi, G. In vivo validation of the multicomponent powder (Vitachelox((R))) against the deposition of polluting ions. Clin. Cosmet. Investig. Dermatol. 2018, 11, 109–113. [Google Scholar] [CrossRef]
- Eghbaliferiz, S.; Iranshahi, M. Prooxidant Activity of Polyphenols, Flavonoids, Anthocyanins and Carotenoids: Updated Review of Mechanisms and Catalyzing Metals. Phytother. Res. 2016, 30, 1379–1391. [Google Scholar] [CrossRef]
- Davinelli, S.; Nielsen, M.E.; Scapagnini, G. Astaxanthin in Skin Health, Repair, and Disease: A Comprehensive Review. Nutrients 2018, 10, 522. [Google Scholar] [CrossRef]
- Davinelli, S.; Bertoglio, J.C.; Polimeni, A.; Scapagnini, G. Cytoprotective Polyphenols Against Chronological Skin Aging and Cutaneous Photodamage. Curr. Pharm. Des. 2018, 24, 99–105. [Google Scholar] [CrossRef]
- Boo, Y.C. p-Coumaric Acid as An Active Ingredient in Cosmetics: A Review Focusing on its Antimelanogenic Effects. Antioxidants 2019, 8, 275. [Google Scholar] [CrossRef]
- Aljuffali, I.A.; Hsu, C.Y.; Lin, Y.K.; Fang, J.Y. Cutaneous delivery of natural antioxidants: The enhancement approaches. Curr. Pharm. Des. 2015, 21, 2745–2757. [Google Scholar] [CrossRef]
- Ohara, M.; Ohyama, Y. Delivery and application of dietary polyphenols to target organs, tissues and intracellular organelles. Curr. Drug Metab. 2014, 15, 37–47. [Google Scholar] [CrossRef]
- Nan, W.H.; Ding, L.; Chen, H.J.; Khan, F.U.; Yu, L.; Sui, X.B.; Shi, X.J. Topical Use of Quercetin-Loaded Chitosan Nanoparticles Against Ultraviolet B Radiation. Front. Pharm. 2018, 9, 826. [Google Scholar] [CrossRef]
- Liu, D.; Hu, H.; Lin, Z.; Chen, D.; Zhu, Y.; Hou, S.; Shi, X. Quercetin deformable liposome: Preparation and efficacy against ultraviolet B induced skin damages in vitro and in vivo. J. Photochem. Photobiol. B 2013, 127, 8–17. [Google Scholar] [CrossRef]
- Vicentini, F.T.; Simi, T.R.; Del Ciampo, J.O.; Wolga, N.O.; Pitol, D.L.; Iyomasa, M.M.; Bentley, M.V.; Fonseca, M.J. Quercetin in w/o microemulsion: In vitro and in vivo skin penetration and efficacy against UVB-induced skin damages evaluated in vivo. Eur. J. Pharm. Biopharm. 2008, 69, 948–957. [Google Scholar] [CrossRef]
Figure 1.
Schematic of aryl hydrocarbon receptor (AhR) and nuclear factor erythroid 2-related factor 2 (Nrf2) in metabolism phases I and II of xenobiotics. Xenobiotics, including drugs and pollutants from external sources, normally undergo phase I metabolism followed by phase II metabolism inside cells [68]. These processes render hydrophobic molecules progressively more water-soluble for excretion from the cells. Phase I metabolism oxidizes, hydrolyzes, or reduces substrates, and phase II metabolism involves conjugation reactions with glutathione, sulfate, acetate, and glucuronic acid. Organic molecules bind to AhR in a complex with protein factors in the cytosol, which promotes its nuclear translocation. In the nucleus, these factors are replaced by a homologous nuclear protein, AhR nuclear translocator (ARNT), leading to the formation of the liganded AhR–ARNT dimer, which binds to the dioxin response element (DER) on the promoter of target genes encoding phase I enzymes [69]. Nrf2 is kept at low levels, because its binding to Kelch-like ECH-associated protein 1 (Keap1) enhances ubiquitination by cullin and subsequent proteasomal degradation [67]. Under oxidative stress, certain cysteine residues of Keap1 can be modified, resulting in the release of Nrf2, which can then enter the nucleus, heterodimerize with small musculo-aponeurotic fibrosarcoma protein (sMAF), and bind to the antioxidant response element (ARE) on the promoters of target genes encoding phase II enzymes and antioxidant enzymes. Organic pollutants contained in particulate matter (PM) can stimulate AhR-dependent gene expression [18,63]. PM-induced activation of AhR can cause an excessive phase I metabolism and an increase of ROS, a condition that might be rescued by enhancing cell antioxidant capacity via an Nrf2-dependent mechanism. Many phenolic compounds derived from plants can activate Nrf2 via phosphoinositide 3-kinases (PI3K)- and Akt (protein kinase B)-dependent mechanisms, increasing downstream gene expression of antioxidant enzymes [5].
Figure 1.
Schematic of aryl hydrocarbon receptor (AhR) and nuclear factor erythroid 2-related factor 2 (Nrf2) in metabolism phases I and II of xenobiotics. Xenobiotics, including drugs and pollutants from external sources, normally undergo phase I metabolism followed by phase II metabolism inside cells [68]. These processes render hydrophobic molecules progressively more water-soluble for excretion from the cells. Phase I metabolism oxidizes, hydrolyzes, or reduces substrates, and phase II metabolism involves conjugation reactions with glutathione, sulfate, acetate, and glucuronic acid. Organic molecules bind to AhR in a complex with protein factors in the cytosol, which promotes its nuclear translocation. In the nucleus, these factors are replaced by a homologous nuclear protein, AhR nuclear translocator (ARNT), leading to the formation of the liganded AhR–ARNT dimer, which binds to the dioxin response element (DER) on the promoter of target genes encoding phase I enzymes [69]. Nrf2 is kept at low levels, because its binding to Kelch-like ECH-associated protein 1 (Keap1) enhances ubiquitination by cullin and subsequent proteasomal degradation [67]. Under oxidative stress, certain cysteine residues of Keap1 can be modified, resulting in the release of Nrf2, which can then enter the nucleus, heterodimerize with small musculo-aponeurotic fibrosarcoma protein (sMAF), and bind to the antioxidant response element (ARE) on the promoters of target genes encoding phase II enzymes and antioxidant enzymes. Organic pollutants contained in particulate matter (PM) can stimulate AhR-dependent gene expression [18,63]. PM-induced activation of AhR can cause an excessive phase I metabolism and an increase of ROS, a condition that might be rescued by enhancing cell antioxidant capacity via an Nrf2-dependent mechanism. Many phenolic compounds derived from plants can activate Nrf2 via phosphoinositide 3-kinases (PI3K)- and Akt (protein kinase B)-dependent mechanisms, increasing downstream gene expression of antioxidant enzymes [5].

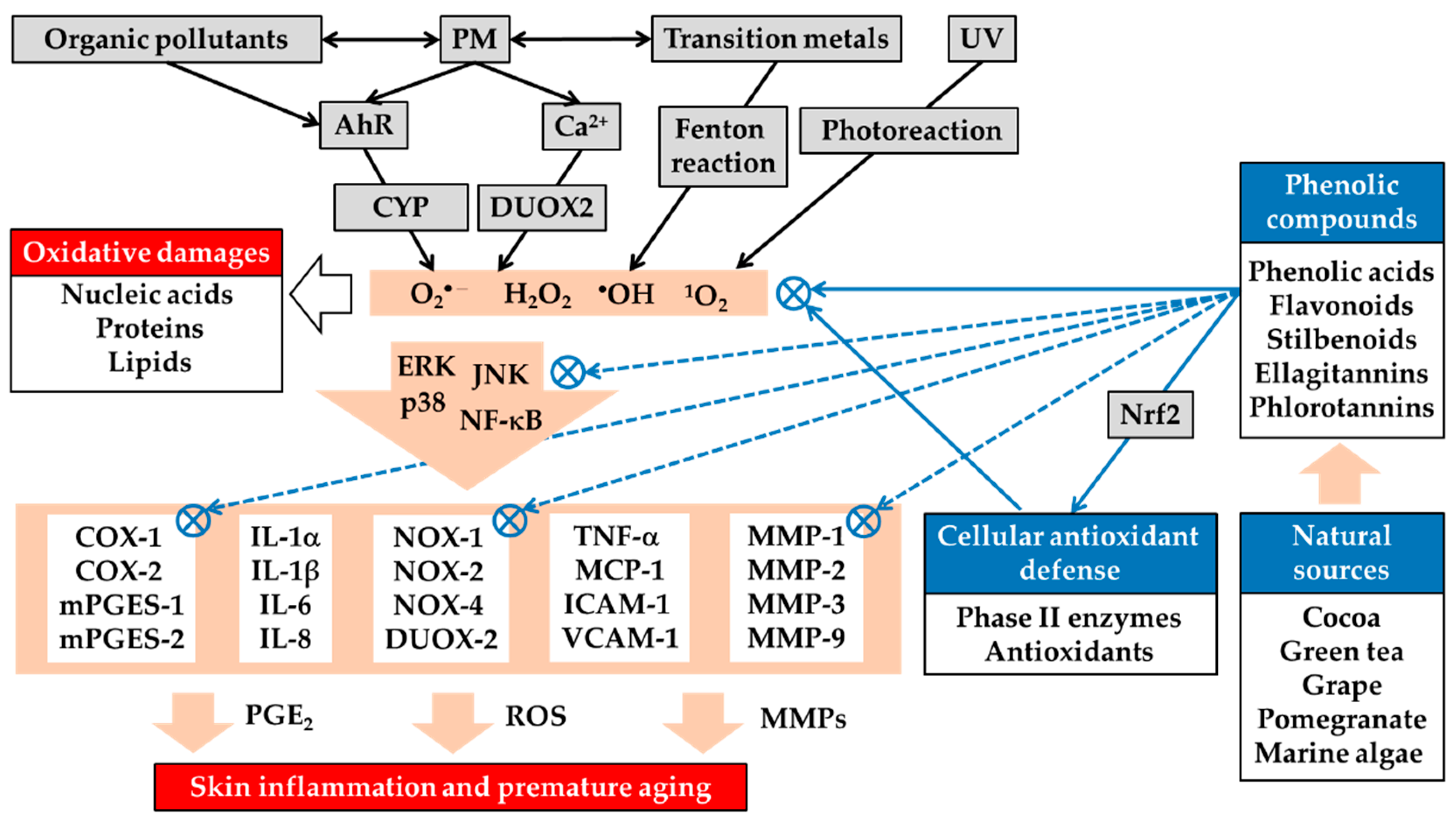
Figure 2.
Airborne PM-induced oxidative stress in the skin and its attenuation by plant-derived phenolic antioxidants. PM stimulates the generation of ROS, such as superoxide anion radical (O2•−), hydrogen peroxide (H2O2), hydroxyl radical (•OH), and singlet oxygen (1O2), by mechanisms that include the AhR/cytochrome P-450 (CYP) pathway, calcium (Ca2+), and dual oxidase 2 (DUOX2)-dependent mechanisms, as well as Fenton reaction catalyzed by transition metals and photochemical or photodynamic reactions resulting from ultraviolet radiation (UV). PM-induced ROS can cause oxidative damage to nucleic acids, proteins, and lipids and activate multiple signaling pathways, such as the extracellular signal-regulated kinase (ERK), c-Jun N-terminal kinase (JNK), p38 mitogen-activated protein (MAP) kinase, and nuclear factor (NF)-κB, linked to gene expression of inflammatory mediators, such as cytokines, chemokines, cell adhesion molecules, and matrix metalloproteinases (MMPs), and synthesis of prostaglandin (PG) E2. Many plants, such as cocoa, green tea, grape, pomegranate, and some marine algae, are rich sources of phenolic acids, flavonoids, stilbenoids, ellagitannins, phlorotannins, and other phenolic compounds. These phenolic compounds can directly scavenge ROS from various sources and can enhance the cellular antioxidant capacity by Nrf2-dependent mechanisms. Through these direct and indirect antioxidant activities, the phenolic compounds can suppress the redox-sensitive signaling pathways leading to the gene expression of inflammatory mediators. Phenolic compounds can also directly inhibit the catalytic activity of protein kinases in the signaling pathways and other metabolic enzymes involved in ROS generation, prostaglandin synthesis, and extracellular matrix degradation. Plant-derived phenolic antioxidants can thereby alleviate oxidative stress and inflammatory skin diseases, such as atopic dermatitis and acne vulgaris, and premature skin aging caused by PM. Some phenolic compounds can increase or decrease AhR-dependent downstream gene expression, and can have positive or negative effects on skin health. NOX, NADPH oxidase; COX, cyclooxygenase; mPGES, microsomal prostaglandin E2 synthase; IL, interleukin; TNF, tumor necrosis factor; MCP, monocyte chemoattractant protein; ICAM, intercellular adhesion molecule; VCAM, vascular cell adhesion molecule.
Figure 2.
Airborne PM-induced oxidative stress in the skin and its attenuation by plant-derived phenolic antioxidants. PM stimulates the generation of ROS, such as superoxide anion radical (O2•−), hydrogen peroxide (H2O2), hydroxyl radical (•OH), and singlet oxygen (1O2), by mechanisms that include the AhR/cytochrome P-450 (CYP) pathway, calcium (Ca2+), and dual oxidase 2 (DUOX2)-dependent mechanisms, as well as Fenton reaction catalyzed by transition metals and photochemical or photodynamic reactions resulting from ultraviolet radiation (UV). PM-induced ROS can cause oxidative damage to nucleic acids, proteins, and lipids and activate multiple signaling pathways, such as the extracellular signal-regulated kinase (ERK), c-Jun N-terminal kinase (JNK), p38 mitogen-activated protein (MAP) kinase, and nuclear factor (NF)-κB, linked to gene expression of inflammatory mediators, such as cytokines, chemokines, cell adhesion molecules, and matrix metalloproteinases (MMPs), and synthesis of prostaglandin (PG) E2. Many plants, such as cocoa, green tea, grape, pomegranate, and some marine algae, are rich sources of phenolic acids, flavonoids, stilbenoids, ellagitannins, phlorotannins, and other phenolic compounds. These phenolic compounds can directly scavenge ROS from various sources and can enhance the cellular antioxidant capacity by Nrf2-dependent mechanisms. Through these direct and indirect antioxidant activities, the phenolic compounds can suppress the redox-sensitive signaling pathways leading to the gene expression of inflammatory mediators. Phenolic compounds can also directly inhibit the catalytic activity of protein kinases in the signaling pathways and other metabolic enzymes involved in ROS generation, prostaglandin synthesis, and extracellular matrix degradation. Plant-derived phenolic antioxidants can thereby alleviate oxidative stress and inflammatory skin diseases, such as atopic dermatitis and acne vulgaris, and premature skin aging caused by PM. Some phenolic compounds can increase or decrease AhR-dependent downstream gene expression, and can have positive or negative effects on skin health. NOX, NADPH oxidase; COX, cyclooxygenase; mPGES, microsomal prostaglandin E2 synthase; IL, interleukin; TNF, tumor necrosis factor; MCP, monocyte chemoattractant protein; ICAM, intercellular adhesion molecule; VCAM, vascular cell adhesion molecule.


{kind=link}
{kind=link}
Table 1.
Protective effects of plant extracts and phenolic compounds against oxidative stress and inflammation induced by airborne PM.
Table 1.
Protective effects of plant extracts and phenolic compounds against oxidative stress and inflammation induced by airborne PM.
| Studies | Models | Targets | Materials | Literature |
|---|---|---|---|---|
| In vitro | HaCaT keratinocytes | Skin inflammation | Eupafolin from Phyla nodiflora | Lee et al., 2016 [70] |
| HaCaT keratinocytes | Skin inflammation | Eupafolin nanoparticles | Lin et al., 2016 [71] | |
| HaCaT keratinocytes | Skin inflammation | Nanoparticles containing 7,3′,4′-trihydroxyisoflavone | Huang et al., 2018 [72] | |
| Fibroblast-like synoviocytes | Rheumatoid arthritis | Resveratrol | Tsai et al., 2017 [73] | |
| HaCaT keratinocytes, 3D-skin models | Skin inflammation | Resveratrol, Resveratryl triacetate | Choi et al., 2018 [74] | |
| Monocytic THP-1 cells, EA.hy926 endothelial cells | Inflammation | Punica granatum extract, Ellagic acid, Punicalagin | Park et al., 2016 [55] | |
| Human epidermal Keratinocytes | Skin inflammation | Punicalagin, (−)-Epigallocatechin gallate. | Seok et al., 2018 [75] | |
| Human dermal fibroblasts | Skin inflammation | (−)-Epigallocatechin gallate | Wang et al., 2019 [76] | |
| HaCaT keratinocytes | Skin inflammation | Ecklonia cava extract, Eckol, Dieckol | Lee et al., 2018 [77]. | |
| HaCaT keratinocytes, 3D-skin models | Skin inflammation | E. cava extract, Dieckol | Ha et al., 2019 [62] | |
| HaCaT keratinocytes | Skin inflammation | Eckol | Zhen et al., 2019 [78] | |
| HaCaT keratinocytes | Skin inflammation | Afzelin from Thesium chinense | Kim et al., 2019 [79] | |
| HaCaT keratinocytes, 3D-skin models | Skin inflammation | Formononetin from Astragalus mongholicus. | Nguyen et al., 2019 [80] | |
| In vivo | Mice | Cardiac inflammation | Chocolate | Villarreal-Calderon et al., 2012 [81] |
| Rats | Bronchial inflammation | Eucheuma cottonii extract | Saputri et al., 2014 [82] | |
| HaCaT keratinocytes, HR-1 hairless mice | Skin inflammation | Diphlorethohydroxycarmalol from Ishige okamurae | Zhen et al., 2019 [83] | |
| Ex vivo | HaCaT keratinocytes, Human skin explants | Skin inflammation | Camellia japonica extract | Kim et al., 2019 [84] |
Table 2.
Potential mechanisms of antioxidant action for plant-derived phenolic compounds.
| Phenolic Compounds | (1) Scavenges ROS or Reduces Their Generation | (2) Enhances Cellular Antioxidant Capacity by Activating Nrf2 |
|---|---|---|
| Procyanidins | Saint-Cricq De Gaulejac et al., 1999 [122] | Rodriguez-Ramiro et al., 2012 [123] Song et al., 2019 [124] |
| (−)-EGCG | An et al., 2014 [125] | Na et al., 2008 [126] |
| Resveratrol | Mahal et al., 2006 [127] Holthoff et al., 2010 [128] | Kim et al., 2019 [129] |
| Quercetin | Aherne et al., 2000 [130] Lopez-Lopez et al., 2004 [131] | Schadich et al., 2016 [132] |
| Rutin | Aherne et al., 2000 [130] | Sthijns et al., 2017 [133] |
| Punicalagin | Kulkarni et al., 2007 [112] | Xu et al., 2015 [134] |
| Ellagic acid | Priyadarsini et al., 2002 [135] | Ding et al., 2014 [136] |
| Dieckol | Cui et al., 2019 [137] | Lee et al., 2015 [138] |
| Eckol | Kang et al., 2005 [139] | Kim et al., 2010 [140] |
Abbreviations: ROS, reactive oxygen species; Nrf2, nuclear factor erythroid 2-related factor 2; EGCG, epigallocatechin gallate.
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Boo, Y.C. Can Plant Phenolic Compounds Protect the Skin from Airborne Particulate Matter? Antioxidants 2019, 8, 379. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox8090379
AMA Style
Boo YC. Can Plant Phenolic Compounds Protect the Skin from Airborne Particulate Matter? Antioxidants. 2019; 8(9):379. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox8090379
Chicago/Turabian StyleBoo, Yong Chool. 2019. "Can Plant Phenolic Compounds Protect the Skin from Airborne Particulate Matter?" Antioxidants 8, no. 9: 379. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox8090379
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.