Salicylic Acid and Melatonin Alleviate the Effects of Heat Stress on Essential Oil Composition and Antioxidant Enzyme Activity in Mentha × piperita and Mentha arvensis L.


 , ,
, ,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material, Culture, and Treatment
2.2. Relative Water Content (RWC)
2.3. Antioxidant Enzyme Activity
2.4. Isolation of Essential Oils
2.5. GC and GC-MS Analysis
2.6. Statistical Analysis
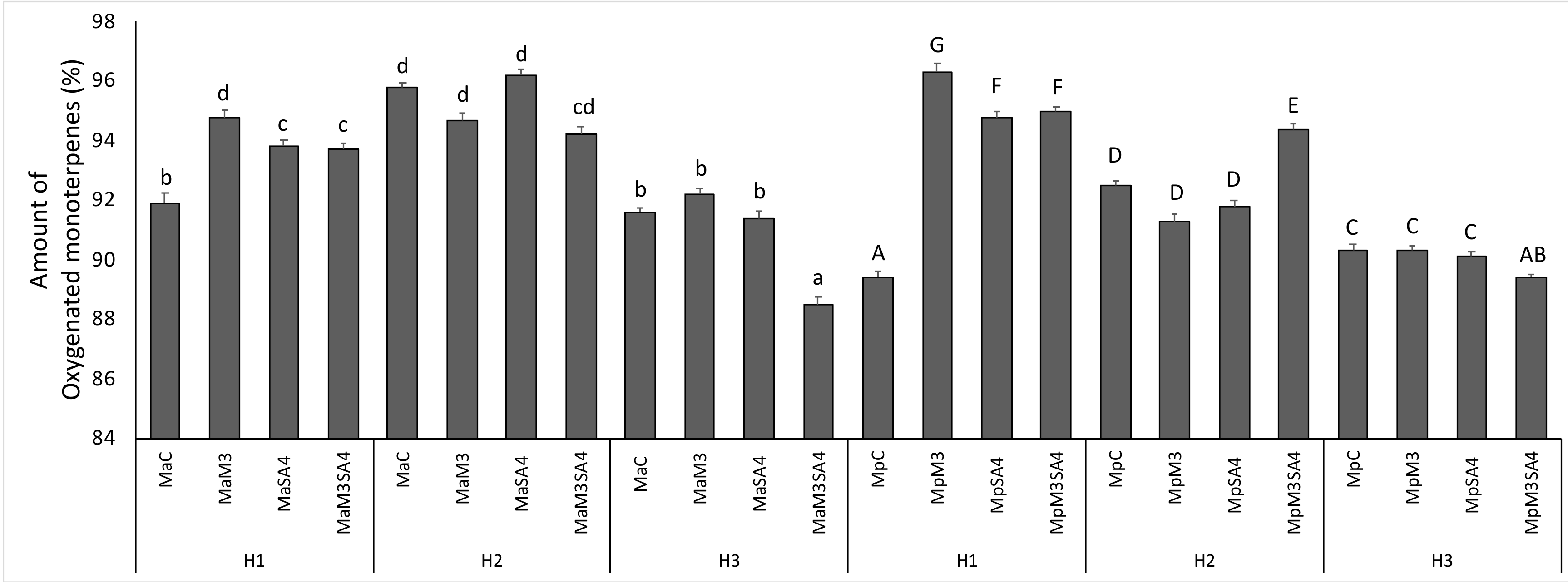
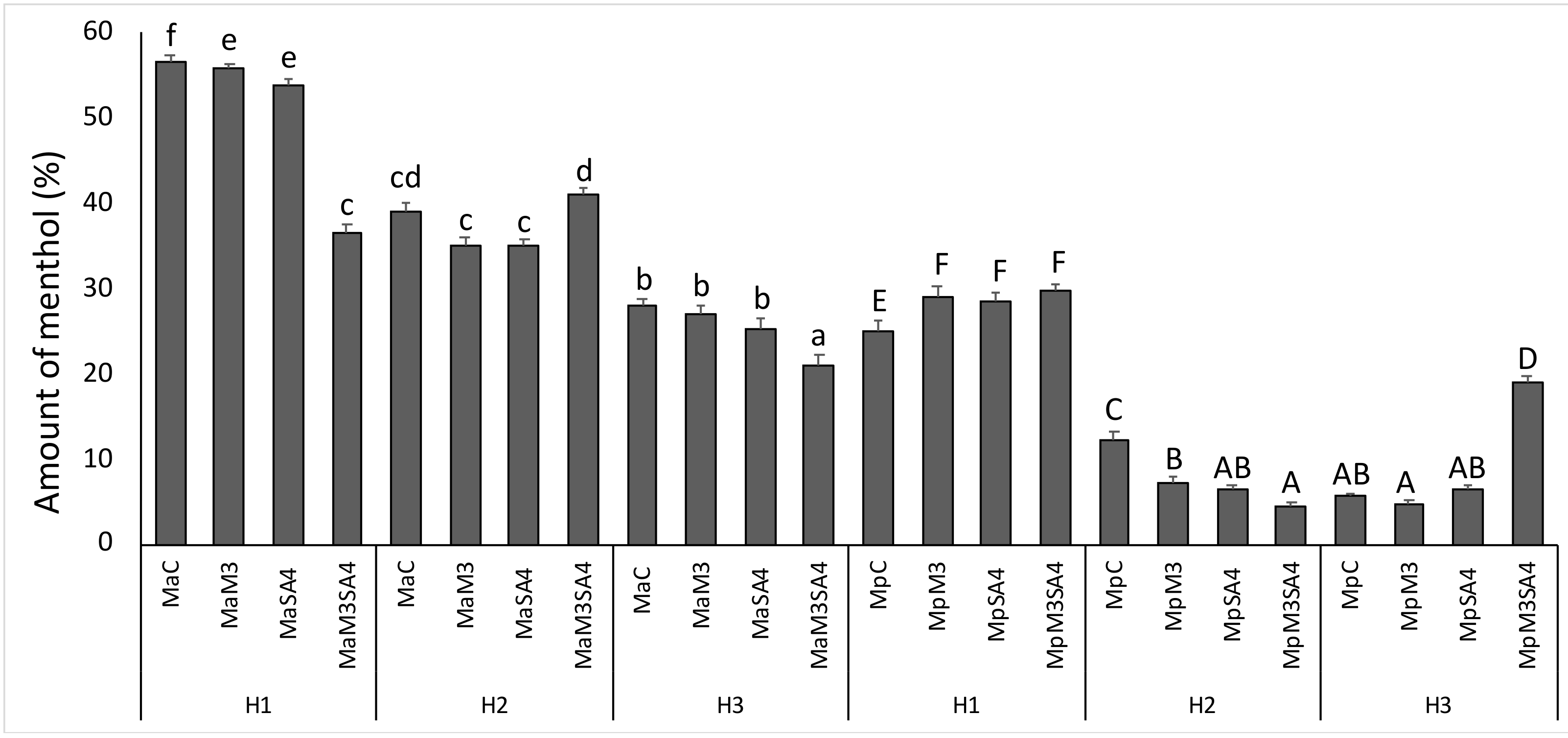
3. Results
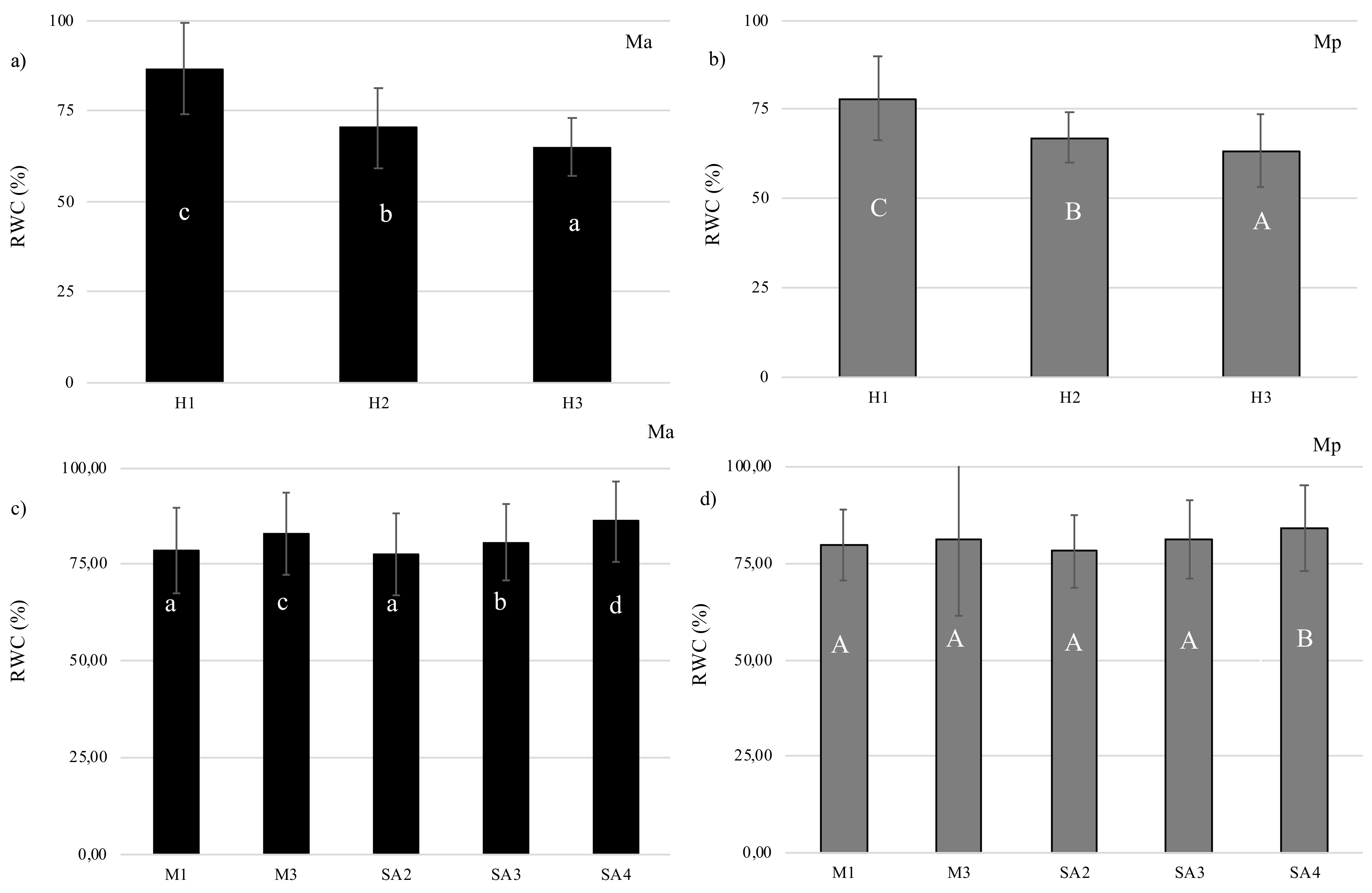
3.1. Relative Water Content (RWC)
3.2. Antioxidant Enzyme Activity
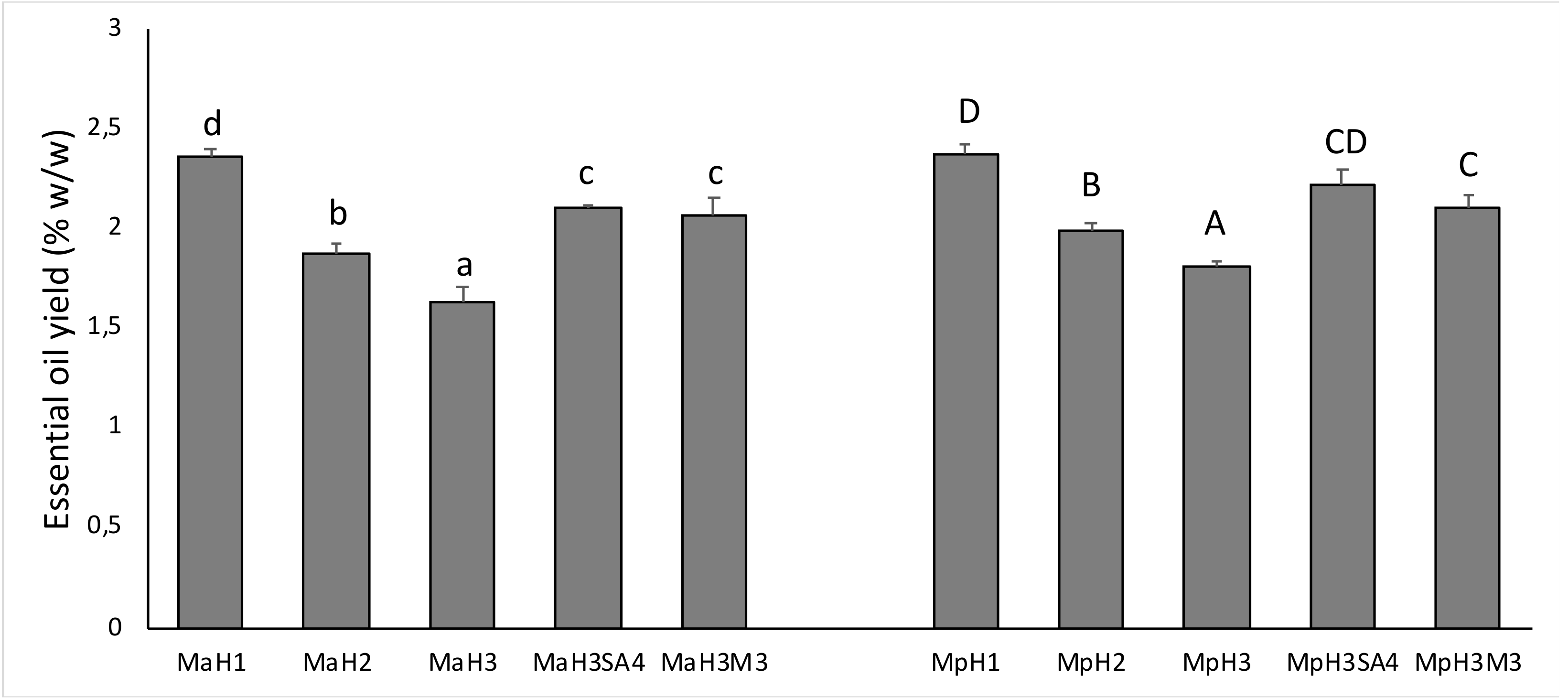
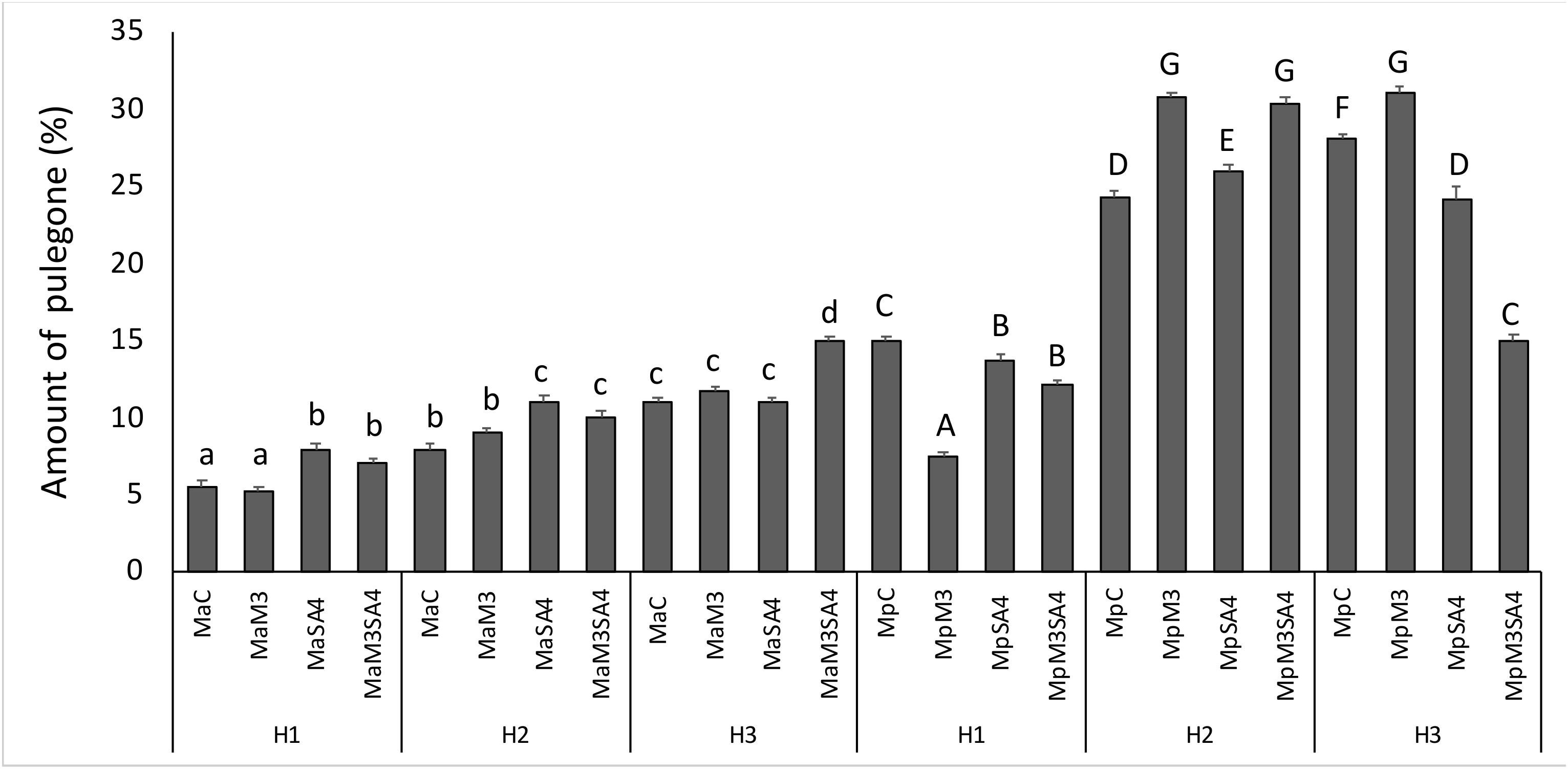
3.3. Essential Oil Yield
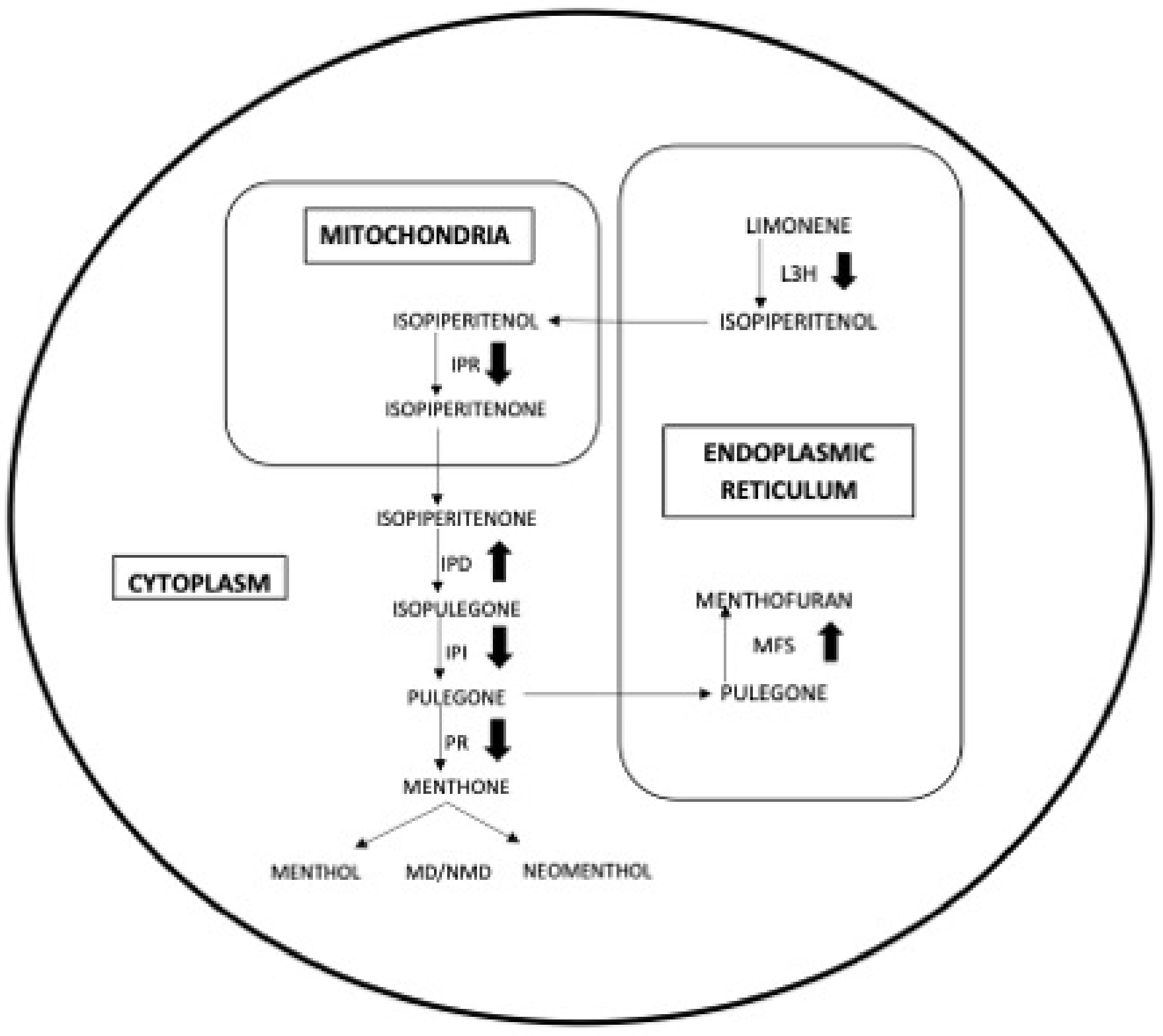
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lawrence, B.M. Mint: The Genus Mentha; CRC Press: Boca Raton, FL, USA, 2006; ISBN 978-0-429-12587-4. [Google Scholar]
- Kokkini, S.; Karousou, R.; Hanlidou, E. HERBS|Herbs of the Labiatae. In Encyclopedia of Food Sciences and Nutrition; Academic Press: Cambridge, MA, USA, 2003; pp. 3082–3090. ISBN 978-0-12-227055-0. [Google Scholar]
- Taneja, S.C.; Chandra, S. 20—Mint. In Handbook of Herbs and Spices (Second Edition); Peter, K.V., Ed.; Woodhead Publishing Series in Food Science, Technology and Nutrition; Woodhead Publishing: Sawston/Cambridge, UK, 2012; pp. 366–387. ISBN 978-0-85709-039-3. [Google Scholar]
- Mander, L.; Liu, H.W. Comprehensive Natural Products II: Chemistry and Biology; Elsevier: Amsterdam, The Netherlands, 2010; ISBN 978-0-08-045382-8. [Google Scholar]
- Fletcher, R.S.; Slimmon, T.; McAuley, C.Y.; Kott, L.S. Heat stress reduces the accumulation of rosmarinic acid and the total antioxidant capacity in spearmint (Mentha spicata L). J. Sci. Food Agric. 2005, 85, 2429–2436. [Google Scholar] [CrossRef]
- Smirnoff, N. The role of active oxygen in the response of plants to water deficit and desiccation. New Phytol. 1993, 125, 27–58. [Google Scholar] [CrossRef]
- Gille, G.; Sigler, K. Oxidative stress and living cell. Folia Microbiol. 1995, 40, 131–152. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Van Breusegem, F. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef]
- Moghaddam, N.M.; Arvin, M.J.; Nezhad, G.R.K.; Maghsoudi, K. Effect of salicylic acid on growth and forage and grain yield of maize under drought stress in field conditions. Seed Plant Prod. J. 2011, 272, 41–55. [Google Scholar]
- Shirasu, K.; Nakajima, H.; Rajasekhar, V.K.; Dixon, R.A.; Lamb, C. Salicylic acid potentiates an agonist-dependent gain control that amplifies pathogen signals in the activation of defense mechanisms. Plant Cell 1997, 9, 261–270. [Google Scholar]
- Knörzer, O.C.; Lederer, B.; Durner, J.; Böger, P. Antioxidative defense activation in soybean cells. Physiol. Plant. 1999, 107, 294–302. [Google Scholar] [CrossRef]
- Calvo, J.R.; González-Yanes, C.; Maldonado, M.D. The role of melatonin in the cells of the innate immunity: A review. J. Pineal Res. 2013, 55, 103–120. [Google Scholar] [CrossRef]
- Manchester, L.C.; Coto-Montes, A.; Boga, J.A.; Andersen, L.P.H.; Zhou, Z.; Galano, A.; Vriend, J.; Tan, D.-X.; Reiter, R.J. Melatonin: An ancient molecule that makes oxygen metabolically tolerable. J. Pineal Res. 2015, 59, 403–419. [Google Scholar] [CrossRef]
- Wang, P.; Sun, X.; Li, C.; Wei, Z.; Liang, D.; Ma, F. Long-term exogenous application of melatonin delays drought-induced leaf senescence in apple. J. Pineal Res. 2013, 54, 292–302. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Sun, Q.; Zhang, H.; Cao, Y.; Weeda, S.; Ren, S.; Guo, Y.-D. Roles of melatonin in abiotic stress resistance in plants. J. Exp. Bot. 2015, 66, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Zhang, Y.; Feng, Z.; Bai, Q.; He, J.; Wang, Y. Effects of Melatonin on Antioxidant Capacity in Naked Oat Seedlings under Drought Stress. Molecules 2018, 23, 1580. [Google Scholar] [CrossRef]
- Nawaz, M.A.; Huang, Y.; Bie, Z.; Ahmed, W.; Reiter, R.J.; Niu, M.; Hameed, S. Melatonin: Current Status and Future Perspectives in Plant Science. Front. Plant Sci. 2016, 6, 1230. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin: Plant growth regulator and/or biostimulator during stress? Trends Plant Sci. 2014, 19, 789–797. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin and its relationship to plant hormones. Ann. Bot. 2018, 121, 195–207. [Google Scholar] [CrossRef]
- Tan, D.-X.; Hardeland, R.; Manchester, L.C.; Paredes, S.D.; Korkmaz, A.; Sainz, R.M.; Mayo, J.C.; Fuentes-Broto, L.; Reiter, R.J. The changing biological roles of melatonin during evolution: From an antioxidant to signals of darkness, sexual selection and fitness. Biol. Rev. 2010, 85, 607–623. [Google Scholar] [CrossRef]
- Heydari, M.; Zanfardino, A.; Taleei, A.; Bushehri, A.A.S.; Hadian, J.; Maresca, V.; Sorbo, S.; Napoli, M.D.; Varcamonti, M.; Basile, A.; et al. Effect of Heat Stress on Yield, Monoterpene Content and Antibacterial Activity of Essential Oils of Mentha x piperita var. Mitcham and Mentha arvensis var. piperascens. Molecules 2018, 23, 1903. [Google Scholar] [CrossRef]
- Dhopte, A.M.; Livera-M, M. Principles and Techniques for Plant Scientist [s]; Agrobios: Jodhpur, India, 2002. [Google Scholar]
- Arjenaki, F.G.; Morshedi, A.; Jabbari, R. Evaluation of Drought Stress on Relative Water Content, Chlorophyll Content and Mineral Elements of Wheat (Triticum aestivum L.) Varieties. Int. J. Agric. Crop Sci. 2012, 4, 726–729. [Google Scholar]
- Maresca, V.; Fusaro, L.; Sorbo, S.; Siciliano, A.; Loppi, S.; Paoli, L.; Monaci, F.; Karam, E.A.; Piscopo, M.; Guida, M.; et al. Functional and structural biomarkers to monitor heavy metal pollution of one of the most contaminated freshwater sites in Southern Europe. Ecotoxicol. Environ. Saf. 2018, 163, 665–673. [Google Scholar] [CrossRef]
- Russo, A.; Formisano, C.; Rigano, D.; Cardile, V.; Arnold, N.A.; Senatore, F. Comparative phytochemical profile and antiproliferative activity on human melanoma cells of essential oils of three lebanese Salvia species. Ind. Crop. Prod. 2016, 83, 492–499. [Google Scholar] [CrossRef]
- Rigano, D.; Arnold, N.A.; Conforti, F.; Menichini, F.; Formisano, C.; Piozzi, F.; Senatore, F. Characterisation of the essential oil of Nepeta glomerata Montbret et Aucher ex Bentham from Lebanon and its biological activities. Nat. Prod. Res. 2011, 25, 614–626. [Google Scholar] [CrossRef] [PubMed]
- Schonfeld, M.A.; Johnson, R.C.; Carver, B.F.; Mornhinweg, D.W. Water Relations in Winter Wheat as Drought Resistance Indicators. Crop Sci. 1988, 28, 526–531. [Google Scholar] [CrossRef]
- Jiang, Y.; Huang, B. Drought and Heat Stress Injury to Two Cool-Season Turfgrasses in Relation to Antioxidant Metabolism and Lipid Peroxidation. Crop Sci. 2001, 41, 436–442. [Google Scholar] [CrossRef]
- Gunasekera, D.; Berkowitz, G.A. Evaluation of contrasting cellular-level acclimation responses to leaf water deficits in three wheat genotypes. Plant Sci. 1992, 86, 1–12. [Google Scholar] [CrossRef]
- Alizadeh, A. Soil, Water, Plants Relationship; Emam Reza University Press: Mashhad, Iran, 2002; Volume 3. [Google Scholar]
- Çoban, Ö.; Göktürk Baydar, N. Brassinosteroid effects on some physical and biochemical properties and secondary metabolite accumulation in peppermint (Mentha piperita L.) under salt stress. Ind. Crop. Prod. 2016, 86, 251–258. [Google Scholar]
- He, Y.; Liu, Y.; Cao, W.; Huai, M.; Xu, B.; Huang, B. Effects of Salicylic Acid on Heat Tolerance Associated with Antioxidant Metabolism in Kentucky Bluegrass. Crop Sci. 2005, 45, 988–995. [Google Scholar] [CrossRef]
- Edreva, A.; Velikova, V.; Tsonev, T.; Dagnon, S.; Gürel, A.; Aktaş, L.; Gesheva, E. Stress-protective role of secondary metabolites: Diversity of functions and mechanisms. Gen. Appl. Plant Physiol. 2008, 34, 67–78. [Google Scholar]
- Cohen, S.D.; Kennedy, J.A. Plant metabolism and the environment: Implications for managing phenolics. Crit. Rev. Food Sci. Nutr. 2010, 50, 620–643. [Google Scholar] [CrossRef]
- Klessig, D.F.; Malamy, J. The salicylic acid signal in plants. Plant Mol. Biol. 1994, 26, 1439–1458. [Google Scholar] [CrossRef]
- Raskin, I. Role of Salicylic Acid in Plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1992, 43, 439–463. [Google Scholar] [CrossRef]
- Zhang, H.-J.; Zhang, N.; Yang, R.-C.; Wang, L.; Sun, Q.-Q.; Li, D.-B.; Cao, Y.-Y.; Weeda, S.; Zhao, B.; Ren, S.; et al. Melatonin promotes seed germination under high salinity by regulating antioxidant systems, ABA and GA4 interaction in cucumber (Cucumis sativus L.). J. Pineal Res. 2014, 57, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Bajwa, V.S.; Shukla, M.R.; Sherif, S.M.; Murch, S.J.; Saxena, P.K. Role of melatonin in alleviating cold stress in Arabidopsis thaliana. J. Pineal Res. 2014, 56, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Saharkhiz, M.J.; Goudarzi, T. Foliar Application of Salicylic acid Changes Essential oil Content and Chemical Compositions of Peppermint (Mentha piperita L.). J. Essent. Oil Bear. Plants 2014, 17, 435–440. [Google Scholar] [CrossRef]
- Croteau, R.B.; Davis, E.M.; Ringer, K.L.; Wildung, M.R. (−)-Menthol biosynthesis and molecular genetics. Naturwissenschaften 2005, 92, 562–577. [Google Scholar] [CrossRef]
- Xu, X.; Sun, Y.; Guo, X.; Sun, B.; Zhang, J. Effects of exogenous melatonin on ascorbate metabolism system in cucumber seedlings under high temperature stress. Yingyong Shengtai Xuebao 2010, 21, 2580–2586. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Temperature | Hormons | CAT | GST | POX | SOD | |
|---|---|---|---|---|---|---|
| Ma | H1 | C | 3.188 ± 0.89 a | 1.02 ± 0.08 a | 19.39 ± 1.25 a | 6.88 ± 0.21 a |
| Ma | M3 | 34.76 ± 1.94 a | 1.78 ± 0.08 c | 126.42 ± 1.25 c | 12.98 ± 1.03 b | |
| Ma | SA4 | 42.56 ± 2.05 a | 1.49 ± 0.06 b | 98.12 ± 3.69 c | 10.33 ± 1.05 bc | |
| Ma | M3SA4 | 66.42 ± 2.45 d | 2.54 ± 0.06 d | 226.77 ± 5.11 d | 18.45 ± 1.13 bc | |
| Ma | H2 | C | 9.46 ± 1.83 b | 1.52 ± 0.03 d | 22.73± 1.04 a | 9.98 ± 1.01 c |
| Ma | M3 | 63.86 ± 2.95 c | 2.01 ± 0.08 e | 245.13 ± 9.46 d | 43.79 ± 1.13 d | |
| Ma | SA4 | 98.73 ± 2.83 b | 2.67 ± 0.08 h | 108.42 ± 3.14 f | 19.07 ± 1.16 e | |
| Ma | M3SA4 | 134.52 ± 2.54 e | 4.29 ± 0.08 i | 269.67 ± 1.22 g | 54.53 ± 1.51 f | |
| Ma | H3 | C | 19.61 ± 1.67 d | 2.28 ± 0.02 f | 4.96 ±1.45 b | 16.88 ± 0.71 d |
| Ma | M3 | 92.37± 2.04 e | 2.67 ± 0.04 g | 316.91± 7.70 e | 49.07 ± 0.73 g | |
| Ma | SA4 | 129.04 ± 3.75 e | 3.74 ± 0.06 l | 146.95 ± 4.22 h | 49.12 ± 0.87 g | |
| Ma | M3SA4 | 130.33 ± 2.16 f | 4.61 ± 0.09 m | 491.12 ± 11.77 i | 58.77 ± 0.89 h | |
| Mp | H1 | C | 9.53 ± 1.33 a | 0.92 ±0.04 a | 11.49 ± 1.25 a | 5.64 ± 0.55 a |
| Mp | M3 | 10.51 ± 1.07 a | 1.70 ± 0.03 c | 61.50 ± 1.42 c | 18.61 ± 1.62 b | |
| Mp | SA4 | 12.54 ± 1.46 a | 1.57 ± 0.07 b | 66.26 ± 2.38 c | 20.61 ± 0.83 bc | |
| Mp | M3SA4 | 41.15 ± 2.03 d | 1.93 ± 0.1 d | 103.57 ±5.45 d | 20.54 ± 0.53 bc | |
| Mp | H2 | C | 19.42 ± 1.47 b | 1.93 ± 0.03 d | 15.19 ± 1.52 a | 22.06 ± 0.90 c |
| Mp | M3 | 29.54 ± 1.19 c | 2.28 ± 0.07 e | 95.12 ± 1.20 d | 28.75 ± 2.23 d | |
| Mp | SA4 | 22.39 ± 1.54 b | 3.12 ± 0.08 h | 154.46 ± 4.56 f | 33.48 ± 0.82 e | |
| Mp | M3SA4 | 54.49 ± 2.50 e | 3.74 ± 0.07 i | 343.70 ± 6.38 g | 51.21 ± 1.31 f | |
| Mp | H3 | C | 42.75 ± 1.6 d | 2.63 ± 0.04 f | 36.79 ± 0.82 b | 29.81 ± 1.19 d |
| Mp | M3 | 53.12 ± 2.04 e | 2.99 ± 0.08 g | 138.90 ± 7.00 e | 56.40 ± 1.47 g | |
| Mp | SA4 | 51.76 ± 2.26 e | 6.13 ± 0.08 l | 417.25 ± 13.36 h | 54.50 ± 1.52 g | |
| Mp | M3SA4 | 130.33 ± 2.16 f | 6.52 ± 0.08 m | 466.18 ± 16.72 i | 66.61 ± 1.27 h |
| Menthofuran | Menthol | Pulegone | Menthone | |
|---|---|---|---|---|
| Menthofuran | 1 | −0.459 ** | 0.345 * | −0.527 ** |
| Menthol | 1 | −0.912 ** | −0.054 | |
| Pulegone | 1 | 0.009 | ||
| Menthone | 1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haydari, M.; Maresca, V.; Rigano, D.; Taleei, A.; Shahnejat-Bushehri, A.A.; Hadian, J.; Sorbo, S.; Guida, M.; Manna, C.; Piscopo, M.; et al. Salicylic Acid and Melatonin Alleviate the Effects of Heat Stress on Essential Oil Composition and Antioxidant Enzyme Activity in Mentha × piperita and Mentha arvensis L. Antioxidants 2019, 8, 547. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox8110547
Haydari M, Maresca V, Rigano D, Taleei A, Shahnejat-Bushehri AA, Hadian J, Sorbo S, Guida M, Manna C, Piscopo M, et al. Salicylic Acid and Melatonin Alleviate the Effects of Heat Stress on Essential Oil Composition and Antioxidant Enzyme Activity in Mentha × piperita and Mentha arvensis L. Antioxidants. 2019; 8(11):547. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox8110547
Chicago/Turabian StyleHaydari, Milad, Viviana Maresca, Daniela Rigano, Alireza Taleei, Ali Akbar Shahnejat-Bushehri, Javad Hadian, Sergio Sorbo, Marco Guida, Caterina Manna, Marina Piscopo, and et al. 2019. "Salicylic Acid and Melatonin Alleviate the Effects of Heat Stress on Essential Oil Composition and Antioxidant Enzyme Activity in Mentha × piperita and Mentha arvensis L." Antioxidants 8, no. 11: 547. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox8110547