Grown to be Blue—Antioxidant Properties and Health Effects of Colored Vegetables. Part I: Root Vegetables

 , , , , ,
, , , , ,  and
and 
Abstract
:1. Introduction
2. Main Colored Root Vegetables
2.1. Potato
2.2. Sweet Potato
2.3. Carrot
2.4. Beet Root
2.5. Yam
2.6. Onion
2.7. Other Root Vegetables
3. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
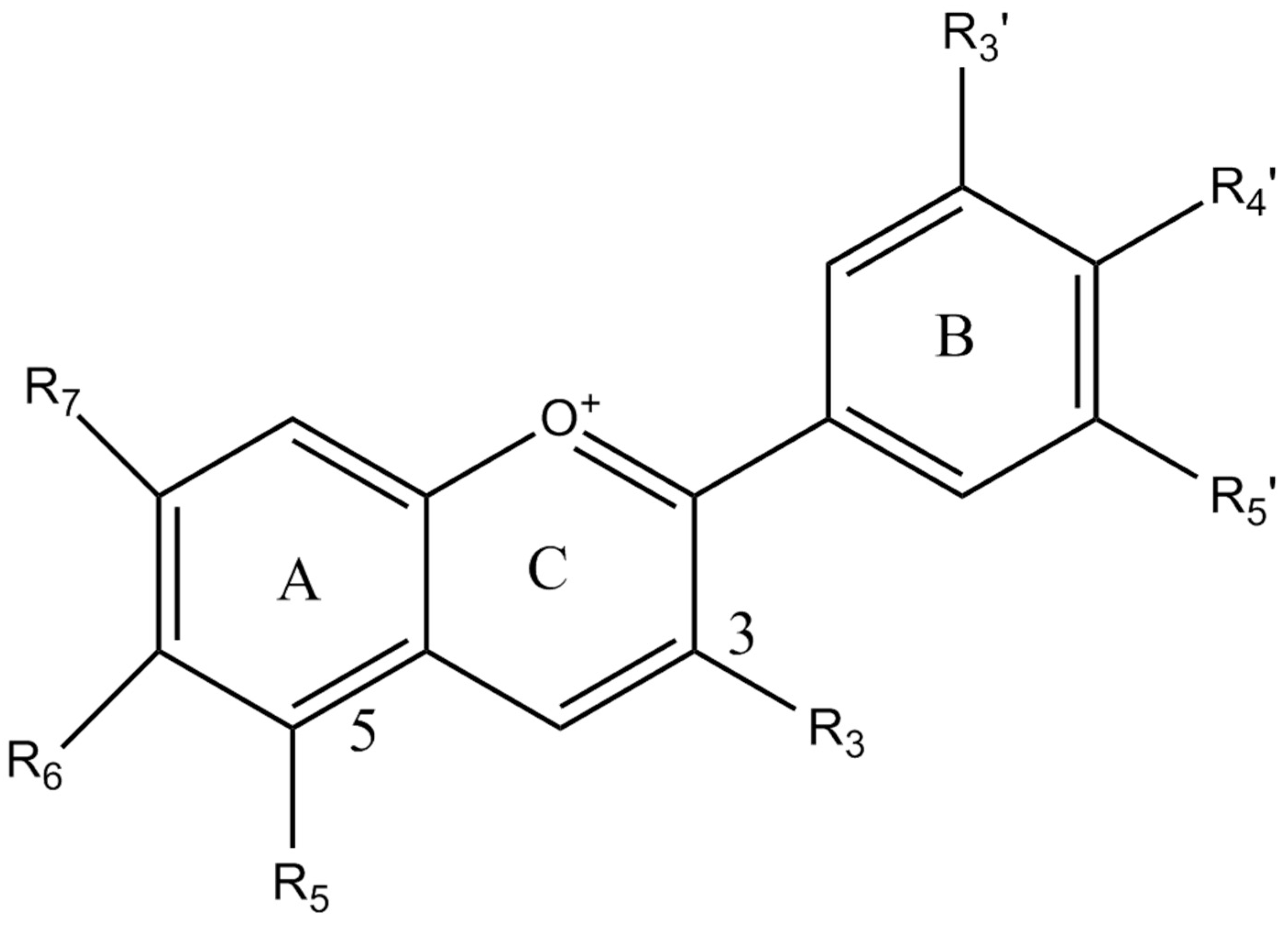
- Sasaki, N.; Nishizaki, Y.; Ozeki, Y.; Miyahara, T. The role of acyl-glucose in anthocyanin modifications. Molecules 2014, 19, 18747–18766. [Google Scholar] [CrossRef] [PubMed]
- Kyriacou, M.C.; Rouphael, Y. Towards a new definition of quality for fresh fruits and vegetables. Sci. Hortic. (Amst.) 2018, 234, 463–469. [Google Scholar] [CrossRef]
- Chaves-Silva, S.; dos Santos, A.L.; Chalfun-Júnior, A.; Zhao, J.; Peres, L.E.P.; Benedito, V.A. Understanding the genetic regulation of anthocyanin biosynthesis in plants—Tools for breeding purple varieties of fruits and vegetables. Phytochemistry 2018, 153, 11–27. [Google Scholar] [CrossRef] [PubMed]
- Menzies, I.J.; Youard, L.W.; Lord, J.M.; Carpenter, K.L.; van Klink, J.W.; Perry, N.B.; Schaefer, H.M.; Gould, K.S. Leaf colour polymorphisms: A balance between plant defence and photosynthesis. J. Ecol. 2016, 104, 104–113. [Google Scholar] [CrossRef]
- Khandagale, K.; Gawande, S. Genetics of bulb colour variation and flavonoids in onion. J. Hortic. Sci. Biotechnol. 2019, 94, 522–532. [Google Scholar] [CrossRef]
- Wrolstad, R.E.; Culver, C.A. Alternatives to Those Artificial FD&C Food Colorants. Annu. Rev. Food Sci. Technol. 2012, 3, 59–77. [Google Scholar]
- Giusti, M.M.; Wrolstad, R.E. Acylated anthocyanins from edible sources and their applications in food systems. Biochem. Eng. J. 2003, 14, 217–225. [Google Scholar] [CrossRef]
- Oki, T.; Masuda, M.; Furuta, S.; Nishiba, Y.; Terahara, N.; Suda, I. Involvement of anthocyanins and other phenolic compounds in radical-scavenging activity of purple-fleshed sweet potato cultivars. J. Food Sci. 2002, 67, 1752–1756. [Google Scholar] [CrossRef]
- Newsome, A.G.; Culver, C.A.; Van Breemen, R.B. Nature’s palette: The search for natural blue colorants. J. Agric. Food Chem. 2014, 62, 6498–6511. [Google Scholar] [CrossRef]
- Tang, Y.; Cai, W.; Xu, B. Profiles of phenolics, carotenoids and antioxidative capacities of thermal processed white, yellow, orange and purple sweet potatoes grown in Guilin, China. Food Sci. Hum. Wellness 2015, 4, 123–132. [Google Scholar] [CrossRef] [Green Version]
- Singh, B.K.; Koley, T.K.; Maurya, A.; Singh, P.M.; Singh, B. Phytochemical and antioxidative potential of orange, red, yellow, rainbow and black coloured tropical carrots (Daucus carota subsp. sativus Schubl. & Martens). Physiol. Mol. Biol. Plants 2018, 24, 899–907. [Google Scholar] [PubMed]
- Rytel, E.; Tajner-czopek, A.; Kita, A.; Aniołowska, M.; Kucharska, A.Z.; Hamouz, K. Content of polyphenols in coloured and yellow fleshed potatoes during dices processing. Food Chem. 2014, 161, 224–229. [Google Scholar] [CrossRef] [PubMed]
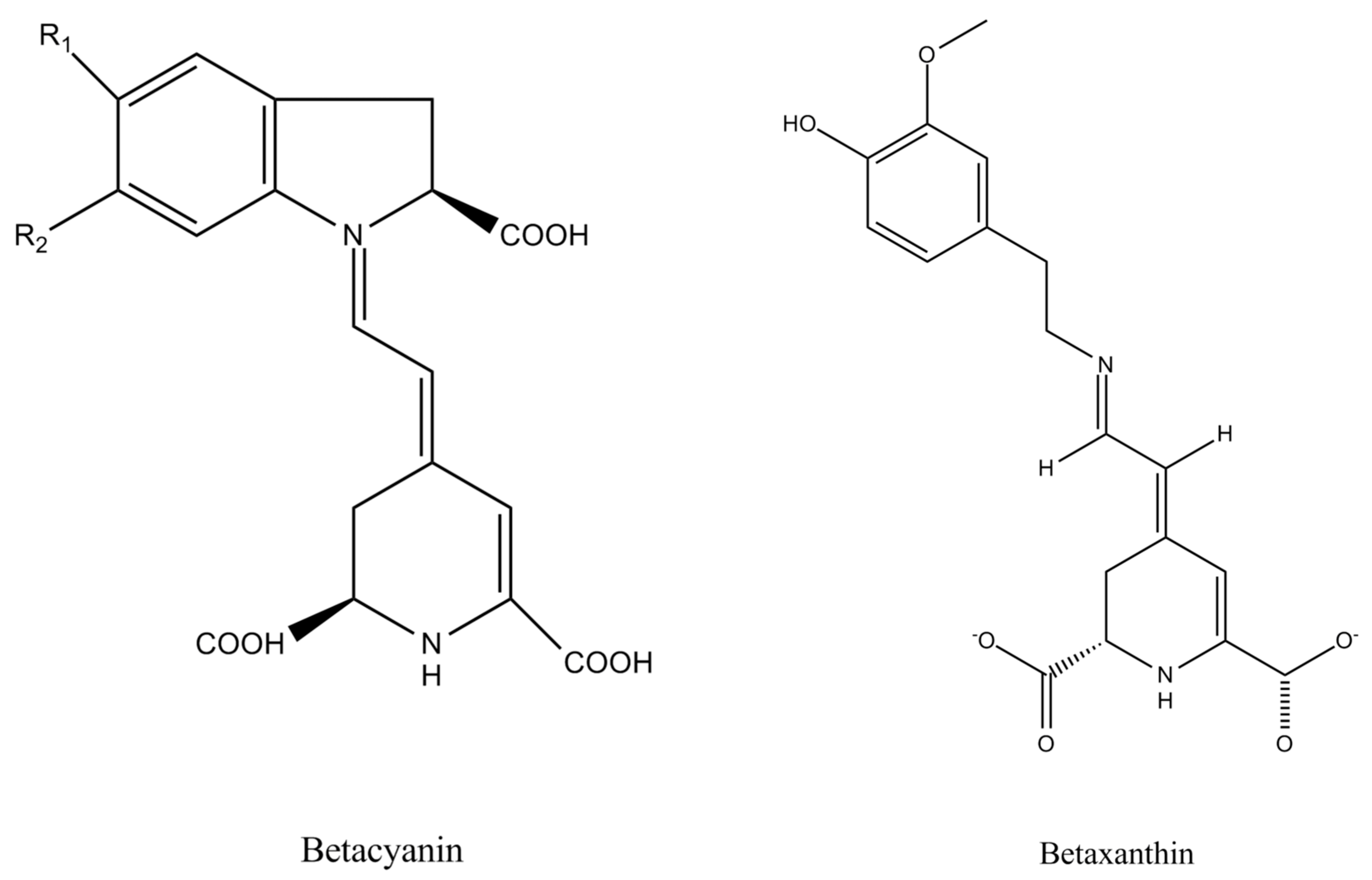
- Khan, M.I.; Giridhar, P. Plant betalains: Chemistry and biochemistry. Phytochemistry 2015, 117, 267–295. [Google Scholar] [CrossRef] [PubMed]
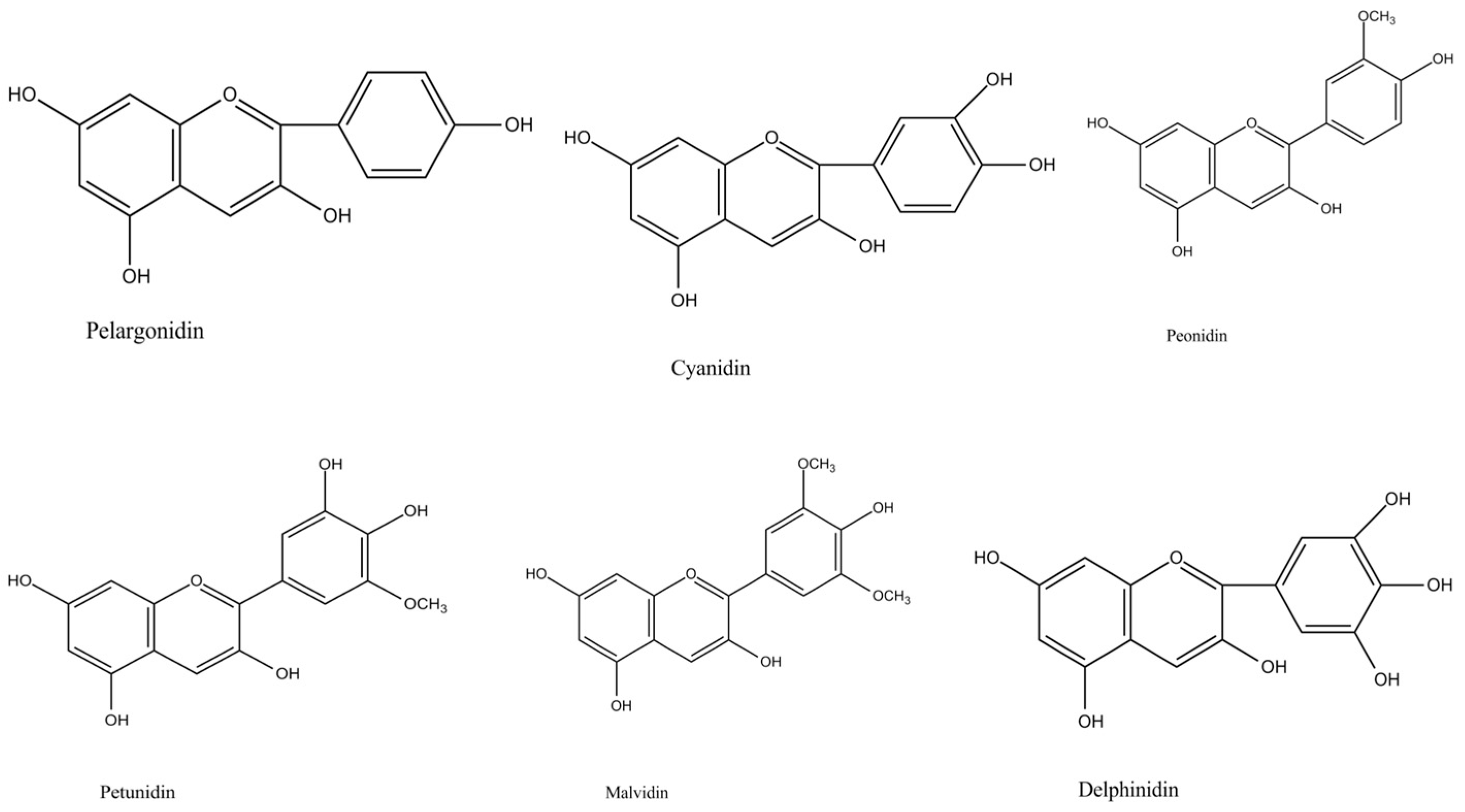
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and anthocyanins: Colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
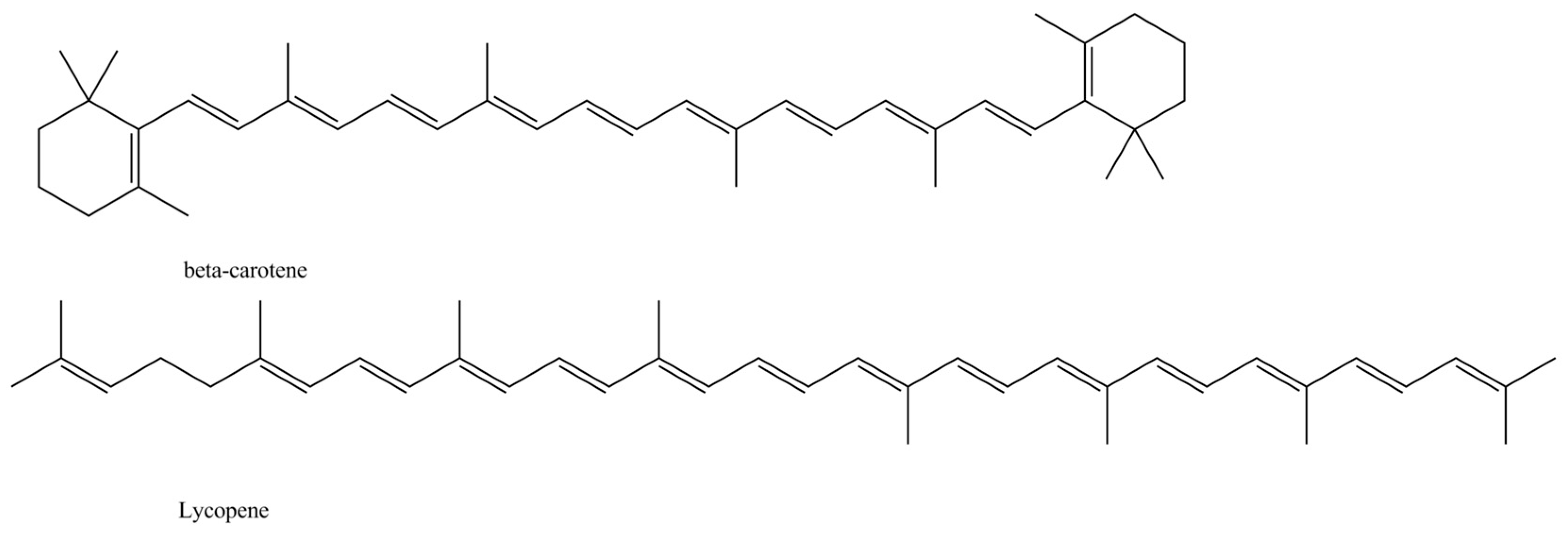
- Merhan, O. The Biochemistry and Antioxidant Properties of Carotenoids. In Carotenoids; Cvetkovic, D., Nikolic, G., Eds.; IntechOpen: London, UK, 2016; pp. 51–66. [Google Scholar]
- Young, A.J.; Lowe, G.L. Carotenoids—Antioxidant properties. Antioxidants 2018, 7, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Indra, V.K. Phytochemicals in Plant Cell Cultures; Vasil, I.K., Ed.; Elsevier: Amsterdam, The Netherlands, 1988. [Google Scholar]
- Rahimi, P.; Abedimanesh, S.; Mesbah-Namin, S.A.; Ostadrahimi, A. Betalains, the nature-inspired pigments, in health and diseases. Crit. Rev. Food Sci. Nutr. 2019, 59, 2949–2978. [Google Scholar] [CrossRef]
- Miguel, M.G. Betalains in some species of the amaranthaceae family: A review. Antioxidants 2018, 7, 53. [Google Scholar] [CrossRef] [Green Version]
- Belhadj Slimen, I.; Najar, T.; Abderrabba, M. Chemical and antioxidant properties of betalains. J. Agric. Food Chem. 2017, 65, 675–689. [Google Scholar] [CrossRef]
- Yin, L.; Chen, T.; Li, Y.; Fu, S.; Li, L.; Xu, M.; Niu, Y. A comparative study on total anthocyanin content, composition of anthocyanidin, total phenolic content and antioxidant activity of pigmented potato peel and flesh. Food Sci. Technol. Res. 2016, 22, 219–226. [Google Scholar] [CrossRef] [Green Version]
- Oertel, A.; Matros, A.; Hartmann, A.; Arapitsas, P.; Dehmer, K.J.; Martens, S.; Mock, H.P. Metabolite profiling of red and blue potatoes revealed cultivar and tissue specific patterns for anthocyanins and other polyphenols. Planta 2017, 246, 281–297. [Google Scholar] [CrossRef]
- Kita, A.; Bąkowska-Barczak, A.; Hamouz, K.; Kułakowska, K.; Lisińska, G. The effect of frying on anthocyanin stability and antioxidant activity of crisps from red- and purple-fleshed potatoes (Solanum tuberosum L.). J. Food Compos. Anal. 2013, 32, 169–175. [Google Scholar] [CrossRef]
- Nemś, A.; Pęksa, A.; Kucharska, A.Z.; Sokół-ŁĘtowska, A.; Kita, A.; Drozdz, W.; Hamouz, K. Anthocyanin and antioxidant activity of snacks with coloured potato. Food Chem. 2015, 172, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Kotíková, Z.; Šulc, M.; Lachman, J.; Pivec, V.; Orsák, M.; Hamouz, K. Carotenoid profile and retention in yellow-, purple- and red-fleshed potatoes after thermal processing. Food Chem. 2016, 197, 992–1001. [Google Scholar] [CrossRef] [PubMed]
- Ge, J.; Hu, Y.; Wang, H.; Huang, Y.; Zhang, P.; Liao, Z.; Chen, M. Profiling of anthocyanins in transgenic purple-fleshed sweet potatoes by HPLC-MS/MS. J. Sci. Food Agric. 2017, 97, 4995–5003. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Deng, L.; Chen, J.; Zhou, S.; Liu, S.; Fu, Y.; Yang, C.; Liao, Z.; Chen, M. An analytical pipeline to compare and characterise the anthocyanin antioxidant activities of purple sweet potato cultivars. Food Chem. 2016, 194, 46–54. [Google Scholar] [CrossRef]
- Pacheco, M.T.; Escribano-Bailón, M.T.; Moreno, F.J.; Villamiel, M.; Dueñas, M. Determination by HPLC-DAD-ESI/MSn of phenolic compounds in Andean tubers grown in Ecuador. J. Food Compos. Anal. 2019, 84, 103258. [Google Scholar] [CrossRef]
- Wang, A.; Li, R.; Ren, L.; Gao, X.; Zhang, Y.; Ma, Z.; Ma, D.; Luo, Y. A comparative metabolomics study of flavonoids in sweet potato with different flesh colors (Ipomoea batatas (L.) Lam). Food Chem. 2018, 260, 124–134. [Google Scholar] [CrossRef]
- Zhang, J.L.; Luo, C.L.; Zhou, Q.; Zhang, Z.C. Isolation and identification of two major acylated anthocyanins from purple sweet potato (Ipomoea batatas L. cultivar Eshu No. 8) by UPLC-QTOF-MS/MS and NMR. Int. J. Food Sci. Technol. 2018, 53, 1932–1941. [Google Scholar] [CrossRef]
- Gizir, A.M.; Turker, N.; Artuvan, E. Pressurized acidified water extraction of black carrot [Daucus carota ssp. sativus var. atrorubens Alef.] anthocyanins. Eur. Food Res. Technol. 2008, 226, 363–370. [Google Scholar] [CrossRef]
- Kirca, A.; Özkan, M.; Cemeroǧlu, B. Effects of temperature, solid content and pH on the stability of black carrot anthocyanins. Food Chem. 2007, 101, 212–218. [Google Scholar] [CrossRef]
- Türker, N.; Erdoğdu, F. Effects of pH and temperature of extraction medium on effective diffusion coefficient of anthocynanin pigments of black carrot (Daucus carota var. L.). J. Food Eng. 2006, 76, 579–583. [Google Scholar] [CrossRef]
- Montilla, E.C.; Arzaba, M.R.; Hillebrand, S.; Winterhalter, P. Anthocyanin composition of black carrot (Daucus carota ssp. sativus var. atrorubens Alef.) cultivars antonina, beta sweet, deep purple, and purple haze. J. Agric. Food Chem. 2011, 59, 3385–3390. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Herrera, P.; Pérez-Rodríguez, M.L.; Aguilera-Delgado, T.; Labari-Reyes, M.J.; Olmedilla-Alonso, B.; Camara, M.; de Pascual-Teresa, S. Anthocyanin profile of red fruits and black carrot juices, purees and concentrates by HPLC-DAD-ESI/MS-QTOF. Int. J. Food Sci. Technol. 2016, 51, 2290–2300. [Google Scholar] [CrossRef]
- Schwarz, M.; Wray, V.; Winterhalter, P. Isolation and identification of novel pyranoanthocyanins from black carrot (Daucus carota L.) juice. J. Agric. Food Chem. 2004, 52, 5095–5101. [Google Scholar] [CrossRef] [PubMed]
- Kaur, A.; Sogi, D.S. Effect of osmotic dehydration on physico-chemical properties and pigment content of carrot (Daucus carota L.) during preserve manufacture. J. Food Process. Preserv. 2017, 41, e13153. [Google Scholar] [CrossRef]
- Mayer-Miebach, E.; Behsnilian, D.; Regier, M.; Schuchmann, H.P. Thermal processing of carrots: Lycopene stability and isomerisation with regard to antioxidant potential. Food Res. Int. 2005, 38, 1103–1108. [Google Scholar] [CrossRef]
- Nemzer, B.; Pietrzkowski, Z.; Spórna, A.; Stalica, P.; Thresher, W.; Michałowski, T.; Wybraniec, S. Betalainic and nutritional profiles of pigment-enriched red beet root (Beta vulgaris L.) dried extracts. Food Chem. 2011, 127, 42–53. [Google Scholar] [CrossRef]
- Downham, A.; Collins, P. Colouring our foods in the last and next millennium. Int. J. Food Sci. Technol. 2000, 35, 5–22. [Google Scholar] [CrossRef]
- Moreno, D.A.; García-Viguera, C.; Gil, J.I.; Gil-Izquierdo, A. Betalains in the era of global agri-food science, technology and nutritional health. Phytochem. Rev. 2008, 7, 261–280. [Google Scholar] [CrossRef]
- Lee, E.J.; An, D.; Nguyen, C.T.T.; Patil, B.S.; Kim, J.; Yoo, K.S. Betalain and betaine composition of greenhouse- or field-produced beetroot (Beta vulgaris L.) and inhibition of HepG2 cell proliferation. J. Agric. Food Chem. 2014, 62, 1324–1331. [Google Scholar] [CrossRef]
- Moriya, C.; Hosoya, T.; Agawa, S.; Sugiyama, Y.; Kozone, I.; Shin-Ya, K.; Terahara, N.; Kumazawa, S. New acylated anthocyanins from purple yam and their antioxidant activity. Biosci. Biotechnol. Biochem. 2015, 79, 1484–1492. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.M.; Yan, R.X.; Zhang, P.T.; Han, X.Y.; Wang, L. Anthocyanin accumulation rate and the biosynthesis related gene expression in Dioscorea alata. Biol. Plant. 2015, 59, 325–330. [Google Scholar] [CrossRef]
- Ramos-Escudero, F.; Santos-Buelga, C.; Pérez-Alonso, J.J.; Yáñez, J.A.; Dueñas, M. HPLC-DAD-ESI/MS identification of anthocyanins in Dioscorea trifida L. yam tubers (purple sachapapa). Eur. Food Res. Technol. 2010, 230, 745–752. [Google Scholar] [CrossRef]
- Fang, Z.; Wu, D.; Yü, D.; Ye, X.; Liu, D.; Chen, J. Phenolic compounds in Chinese purple yam and changes during vacuum frying. Food Chem. 2011, 128, 943–948. [Google Scholar] [CrossRef]
- Price, E.J.; Bhattacharjee, R.; Lopez-Montes, A.; Fraser, P.D. Carotenoid profiling of yams: Clarity, comparisons and diversity. Food Chem. 2018, 259, 130–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frond, A.D.; Iuhas, C.I.; Stirbu, I.; Leopold, L.; Socaci, S.; Andreea, S.; Ayvaz, H.; Andreea, S.; Mihai, S.; Diaconeasa, Z.; et al. Phytochemical characterization of five edible purple-reddish vegetables: Anthocyanins, flavonoids, and phenolic acid derivatives. Molecules 2019, 24, 1536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, T.; Sun, G.; Cao, L.; Li, J.; Wang, L. Rheological behavior of film-forming solutions and film properties from Artemisia sphaerocephala Krasch. gum and purple onion peel extract. Food Hydrocoll. 2018, 82, 124–134. [Google Scholar] [CrossRef]
- Koley, T.K.; Khan, Z.; Oulkar, D.; Singh, B.K.; Maurya, A.; Singh, B.; Banerjee, K. High resolution LC-MS characterization of phenolic compounds and the evaluation of antioxidant properties of a tropical purple radish genotype. Arab. J. Chem. 2017. [Google Scholar] [CrossRef]
- Lin, L.Z.; Sun, J.; Chen, P.; Harnly, J.A. LC-PDA-ESI/MSn identification of new anthocyanins in purple bordeaux radish (Raphanus sativus L. Variety). J. Agric. Food Chem. 2011, 59, 6616–6627. [Google Scholar] [CrossRef]
- Park, N.I.; Xu, H.; Li, X.; Jang, I.H.; Park, S.; Ahn, G.H.; Lim, Y.P.; Kim, S.J.; Park, S.U. Anthocyanin accumulation and expression of anthocyanin biosynthetic genes in radish (Raphanus sativus L.). J. Agric. Food Chem. 2011, 59, 6034–6039. [Google Scholar] [CrossRef]
- Tatsuzawa, F.; Toki, K.; Saito, N.; Shinoda, K.; Shigihara, A.; Honda, T. Anthocyanin occurrence in the root peels, petioles and flowers of red radish (Raphanus sativus L.). Dye. Pigment. 2008, 79, 83–88. [Google Scholar] [CrossRef]
- Park, C.H.; Yeo, H.J.; Kim, N.S.; Eun, P.Y.; Kim, S.-J.; Arasu, M.V.; Al-Dhabi, N.A.; Park, S.-Y.; Kim, J.K.; Park, S.U. Metabolic profiling of pale green and purple kohlrabi (Brassica oleracea var. gongylodes). Appl. Biol. Chem. 2017, 60, 249–257. [Google Scholar] [CrossRef]
- Zhang, Y.; Hu, Z.; Zhu, M.; Zhu, Z.; Wang, Z.; Tian, S.; Chen, G. Anthocyanin Accumulation and Molecular Analysis of Correlated Genes in Purple Kohlrabi (Brassica oleracea var. gongylodes L.). J. Agric. Food Chem. 2015, 63, 4160–4169. [Google Scholar] [CrossRef] [PubMed]
- Park, W.T.; Kim, J.K.; Park, S.; Lee, S.W.; Li, X.; Kim, Y.B.; Uddin, M.R.; Park, N.I.; Kim, S.J.; Park, S.U. Metabolic profiling of glucosinolates, anthocyanins, carotenoids, and other secondary metabolites in kohlrabi (Brassica oleracea var. gongylodes). J. Agric. Food Chem. 2012, 60, 8111–8116. [Google Scholar] [CrossRef] [PubMed]
- Rahim, M.A.; Robin, A.H.K.; Natarajan, S.; Jung, H.J.; Lee, J.; Kim, H.R.; Kim, H.T.; Park, J.I.; Nou, I.S. Identification and characterization of anthocyanin biosynthesis-related genes in kohlrabi. Appl. Biochem. Biotechnol. 2018, 184, 1120–1141. [Google Scholar] [CrossRef] [PubMed]
- Kaushal, P.; Kumar, V.; Sharma, H.K. Utilization of taro (Colocasia esculenta): A review. J. Food Sci. Technol. 2015, 52, 27–40. [Google Scholar] [CrossRef]
- Ziarani, M.G.; Moradi, R.; Lashgari, N.; Kruger, H.G. Metal-Free Synthetic Organic Dyes; Elsevier: London, UK, 2018; ISBN 978-0-12-815647-6. [Google Scholar]
- Pires, T.C.S.P.; Barros, L.; Santos-Buelga, C.; Ferreira, I.C.F.R. Edible flowers: Emerging components in the diet. Trends Food Sci. Technol. 2019, 93, 244–258. [Google Scholar] [CrossRef]
- Amchova, P.; Kotolova, H.; Ruda-Kucerova, J. Health safety issues of synthetic food colorants. Regul. Toxicol. Pharmacol. 2015, 73, 914–922. [Google Scholar] [CrossRef]
- El-Wahab, H.M.F.A.; Moram, G.S.E.D. Toxic effects of some synthetic food colorants and/or flavor additives on male rats. Toxicol. Ind. Health 2013, 29, 224–232. [Google Scholar] [CrossRef]
- Rohrig, B. The chemistry of food coloring: Eating with your eyes. ChemMatters 2015, 2015, 5–7. [Google Scholar]
- Martins, N.; Roriz, C.L.; Morales, P.; Barros, L.; Ferreira, I.C.F.R. Food colorants: Challenges, opportunities and current desires of agro-industries to ensure consumer expectations and regulatory practices. Trends Food Sci. Technol. 2016, 52, 1–15. [Google Scholar] [CrossRef] [Green Version]
- De Pascual-Teresa, S.; Moreno, D.A.; García-Viguera, C. Flavanols and anthocyanins in cardiovascular health: A review of current evidence. Int. J. Mol. Sci. 2010, 11, 1679–1703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horbowicz, M.; Grzesiuk, A.; DĘBski, H.; Kosson, R. Anthocyanins of fruits and vegetables - Their occurrence, analysis and role in human. Veg. Crop. Res. Bull. 2008, 68, 5–22. [Google Scholar] [CrossRef]
- Mazewski, C.; Liang, K.; Gonzalez de Mejia, E. Comparison of the effect of chemical composition of anthocyanin-rich plant extracts on colon cancer cell proliferation and their potential mechanism of action using in vitro, in silico, and biochemical assays. Food Chem. 2018, 242, 378–388. [Google Scholar] [CrossRef]
- Sestari, I.; Zsögön, A.; Rehder, G.G.; de Lira Teixeira, L.; Hassimotto, N.M.A.; Purgatto, E.; Benedito, V.A.; Peres, L.E.P. Near-isogenic lines enhancing ascorbic acid, anthocyanin and carotenoid content in tomato (Solanum lycopersicum L. cv Micro-Tom) as a tool to produce nutrient-rich fruits. Sci. Hortic. (Amst.) 2014, 175, 111–120. [Google Scholar] [CrossRef]
- Pérez-Gregorio, M.R.; García-Falcón, M.S.; Simal-Gándara, J. Flavonoids changes in fresh-cut onions during storage in different packaging systems. Food Chem. 2011, 124, 652–658. [Google Scholar] [CrossRef]
- Pérez-Gregorio, M.R.; Regueiro, J.; Simal-Gándara, J.; Rodrigues, A.S.; Almeida, D.P.F. Increasing the added-value of onions as a source of antioxidant flavonoids: A Critical Review. Crit. Rev. Food Sci. Nutr. 2014, 54, 1050–1062. [Google Scholar] [CrossRef]
- Agcam, E.; Akyıldız, A.; Balasubramaniam, V.M. Optimization of anthocyanins extraction from black carrot pomace with thermosonication. Food Chem. 2017, 237, 461–470. [Google Scholar] [CrossRef]
- McGill, A.E.J. The potential effects of demands for natural and safe foods on global food security. Trends Food Sci. Technol. 2009, 20, 402–406. [Google Scholar] [CrossRef]
- Yusuf, M.; Shabbir, M.; Mohammad, F. Natural colorants: historical, processing and sustainable prospects. Nat. Prod. Bioprospect. 2017, 7, 123–145. [Google Scholar] [CrossRef] [Green Version]
- Dusuki, N.J.S.; Abu Bakar, M.F.; Abu Bakar, F.I.; Ismail, N.A.; Azman, M.I. Proximate composition and antioxidant potential of selected tubers peel. Food Res. 2019, 4, 121–126. [Google Scholar] [CrossRef]
- Yahia, E.M.; Barrera, A. Antioxidant capacity and correlation with phenolic compounds and carotenoids in 40 horticultural commodities. Acta Hortic. 2010, 877, 1215–1220. [Google Scholar] [CrossRef]
- Hamouz, K.; Lachman, J.; Pazderů, K.; Tomášek, J.; Hejtmánková, K.; Pivec, V. Differences in anthocyanin content and antioxidant activity of potato tubers with different flesh colour. Plant Soil Environ. 2011, 57, 478–485. [Google Scholar] [CrossRef] [Green Version]
- Sass-Kiss, A.; Kiss, J.; Milotay, P.; Kerek, M.M.; Toth-Markus, M. Differences in anthocyanin and carotenoid content of fruits and vegetables. Food Res. Int. 2005, 38, 1023–1029. [Google Scholar] [CrossRef]
- Oliveira, H.; Roma-Rodrigues, C.; Santos, A.; Veigas, B.; Brás, N.; Faria, A.; Calhau, C.; de Freitas, V.; Baptista, P.V.; Mateus, N.; et al. GLUT1 and GLUT3 involvement in anthocyanin gastric transport-Nanobased targeted approach. Sci. Rep. 2019, 9, 789. [Google Scholar] [CrossRef]
- Reyes, L.E.; Miller, J.C.J.; Cisneros-Zevallos, L. Environmental conditions influence the content and yield of anthocyanins and total phenolics in purple- and red-flesh potatoes during tuber development. Am. J. Potato Res. 2004, 81, 187–193. [Google Scholar] [CrossRef]
- Gu, K.D.; Wang, C.K.; Hu, D.G.; Hao, Y.J. How do anthocyanins paint our horticultural products? Sci. Hortic. (Amst.) 2019, 249, 257–262. [Google Scholar] [CrossRef]
- Leong, S.Y.; Oey, I. Effects of processing on anthocyanins, carotenoids and vitamin C in summer fruits and vegetables. Food Chem. 2012, 133, 1577–1587. [Google Scholar] [CrossRef]
- Espinosa-Acosta, G.; Ramos-Jacques, A.L.; Molina, G.A.; Maya-Cornejo, J.; Esparza, R.; Hernandez-Martinez, A.R.; Sánchez-González, I.; Estevez, M. Stability analysis of anthocyanins using alcoholic extracts from black carrot (Daucus carota ssp. Sativus var. Atrorubens Alef.). Molecules 2018, 23, 2744. [Google Scholar]
- Ersus, S.; Yurdagel, U. Microencapsulation of anthocyanin pigments of black carrot (Daucus carota L.) by spray drier. J. Food Eng. 2007, 80, 805–812. [Google Scholar] [CrossRef]
- Braga, A.R.C.; Murador, D.C.; de Souza Mesquita, L.M.; de Rosso, V.V. Bioavailability of anthocyanins: Gaps in knowledge, challenges and future research. J. Food Compos. Anal. 2018, 68, 31–40. [Google Scholar] [CrossRef]
- Takahata, Y.; Kai, Y.; Tanaka, M.; Nakayama, H.; Yoshinaga, M. Enlargement of the variances in amount and composition of anthocyanin pigments in sweetpotato storage roots and their effect on the differences in DPPH radical-scavenging activity. Sci. Hortic. (Amst.) 2011, 127, 469–474. [Google Scholar] [CrossRef]
- Assous, M.T.M.; Abdel-Hady, M.M.; Medany, G.M. Evaluation of red pigment extracted from purple carrots and its utilization as antioxidant and natural food colorants. Ann. Agric. Sci. 2014, 59, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Chhikara, N.; Kushwaha, K.; Sharma, P.; Gat, Y.; Panghal, A. Bioactive compounds of beetroot and utilization in food processing industry: A critical review. Food Chem. 2019, 272, 192–200. [Google Scholar] [CrossRef]
- Stintzing, F.C.; Carle, R. Betalains-emerging prospects for food scientists. Trends Food Sci. Technol. 2007, 18, 514–525. [Google Scholar] [CrossRef]
- Stokstad, E. The new potato. Science 2019, 363, 574–577. [Google Scholar] [CrossRef]
- Tsang, C.; Smail, N.F.; Almoosawi, S.; McDougall, G.J.M.; Al-Dujaili, E.A.S. Antioxidant Rich Potato Improves Arterial Stiffness in Healthy Adults. Plant Foods Hum. Nutr. 2018, 73, 203–208. [Google Scholar] [CrossRef] [Green Version]
- Tian, J.; Chen, J.; Ye, X.; Chen, S. Health benefits of the potato affected by domestic cooking: A review. Food Chem. 2016, 202, 165–175. [Google Scholar] [CrossRef]
- Camire, M.E.; Kubow, S.; Donelly, D.J. Potatoes and human health. Crit. Rev. Food Sci. Nutr. 2009, 49, 823–840. [Google Scholar] [CrossRef]
- Burlingame, B.; Mouillé, B.; Charrondière, R. Nutrients, bioactive non-nutrients and anti-nutrients in potatoes. J. Food Compos. Anal. 2009, 22, 494–502. [Google Scholar] [CrossRef]
- Nemś, A.; Pęksa, A. Polyphenols of coloured-flesh potatoes as native antioxidants in stored fried snacks. LWT Food Sci. Technol. 2018, 97, 597–602. [Google Scholar] [CrossRef]
- Ombra, M.N.; Fratianni, F.; Granese, T.; Cardinale, F.; Cozzolino, A.; Nazzaro, F. In vitro antioxidant, antimicrobial and anti-proliferative activities of purple potato extracts (Solanum tuberosum cv Vitelotte noire) following simulated gastro-intestinal digestion. Nat. Prod. Res. 2015, 29, 1087–1091. [Google Scholar] [CrossRef] [PubMed]
- Shock, C.C.; Brown, C.R.; Sathuvalli, V.; Charlton, B.A.; Yilma, S.; Hane, D.C.; Quick, R.; Rykbost, K.A. TerraRossa: A mid-season specialty potato with red flesh and skin and resistance to common scab and golden cyst nematode. Am. J. Potato Res. 2018, 95, 597–605. [Google Scholar] [CrossRef]
- Ayoub, H.M.; McDonald, M.R.; Sullivan, J.A.; Tsao, R.; Meckling, K.A. Proteomic profiles of adipose and liver tissues from an animal model of metabolic syndrome fed purple vegetables. Nutrients 2018, 10, 456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayoub, H.M.; McDonald, M.R.; Sullivan, J.A.; Tsao, R.; Platt, M.; Simpson, J.; Meckling, K.A. The effect of anthocyanin-rich purple vegetable diets on metabolic syndrome in obese zucker rats. J. Med. Food 2017, 20, 1240–1249. [Google Scholar] [CrossRef]
- Sun, X.; Du, M.; Navarre, D.A.; Zhu, M.J. Purple potato extract promotes intestinal epithelial differentiation and barrier function by activating AMP-activated protein kinase. Mol. Nutr. Food Res. 2018, 62, 1700536. [Google Scholar] [CrossRef]
- Madiwale, G.P.; Reddivari, L.; Holm, D.G.; Vanamala, J. Storage elevates phenolic content and antioxidant activity but suppresses antiproliferative and pro-apoptotic properties of colored-flesh potatoes against human colon cancer cell lines. J. Agric. Food Chem. 2011, 59, 8155–8166. [Google Scholar] [CrossRef]
- Kim, D.H.; Kim, M.; Oh, S.B.; Lee, K.M.; Kim, S.M.; Nho, C.W.; Yoon, W.B.; Kang, K.; Pan, C.H. The Protective Effect of Antioxidant Enriched Fractions from Colored Potatoes Against Hepatotoxic Oxidative Stress in Cultured Hepatocytes and Mice. J. Food Biochem. 2017, 41, e12315. [Google Scholar] [CrossRef] [Green Version]
- Tian, J.; Chen, J.; Lv, F.; Chen, S.; Chen, J.; Liu, D.; Ye, X. Domestic cooking methods affect the phytochemical composition and antioxidant activity of purple-fleshed potatoes. Food Chem. 2016, 197, 1264–1270. [Google Scholar] [CrossRef]
- Tierno, R.; Hornero-Méndez, D.; Gallardo-Guerrero, L.; López-Pardo, R.; de Galarreta, J.I.R. Effect of boiling on the total phenolic, anthocyanin and carotenoid concentrations of potato tubers from selected cultivars and introgressed breeding lines from native potato species. J. Food Compos. Anal. 2015, 41, 58–65. [Google Scholar] [CrossRef] [Green Version]
- Lachman, J.; Hamouz, K.; Orsák, M.; Pivec, V.; Hejtmánková, K.; Pazderů, K.; Dvořák, P.; Čepl, J. Impact of selected factors-Cultivar, storage, cooking and baking on the content of anthocyanins in coloured-flesh potatoes. Food Chem. 2012, 133, 1107–1116. [Google Scholar] [CrossRef]
- Bellumori, M.; Innocenti, M.; Michelozzi, M.; Cerretani, L.; Mulinacci, N. Coloured-fleshed potatoes after boiling: Promising sources of known antioxidant compounds. J. Food Compos. Anal. 2017, 59, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Perla, V.; Holm, D.G.; Jayanty, S.S. Effects of cooking methods on polyphenols, pigments and antioxidant activity in potato tubers. LWT Food Sci. Technol. 2012, 45, 161–171. [Google Scholar] [CrossRef]
- Qiu, G.; Wang, D.; Song, X.; Deng, Y.; Zhao, Y. Degradation kinetics and antioxidant capacity of anthocyanins in air-impingement jet dried purple potato slices. Food Res. Int. 2018, 105, 121–128. [Google Scholar] [CrossRef]
- Rytel, E.; Tajner-Czopek, A.; Kita, A.; Kucharska, A.Z.; Sokół-Łętowska, A.; Hamouz, K. Content of anthocyanins and glycoalkaloids in blue-fleshed potatoes and changes in the content of α-solanine and α-chaconine during manufacture of fried and dried products. Int. J. Food Sci. Technol. 2018, 53, 719–727. [Google Scholar] [CrossRef]
- Zhang, L.; Zhao, L.; Bian, X.; Guo, K.; Zhou, L.; Wei, C. Characterization and comparative study of starches from seven purple sweet potatoes. Food Hydrocoll. 2018, 80, 168–176. [Google Scholar] [CrossRef]
- Li, A.; Xiao, R.; He, S.; An, X.; He, Y.; Wang, C.; Yin, S.; Wang, B.; Shi, X.; He, J. Research advances of purple sweet potato anthocyanins: extraction, identification, stability, bioactivity, application, and biotransformation. Molecules 2019, 24, 3816. [Google Scholar] [CrossRef] [Green Version]
- Suda, I.; Oki, T.; Masuda, M.; Kobayashi, M.; Nishiba, Y.; Furuta, S. Physiological functionality of purple-fleshed sweet potatoes containing anthocyanins and their utilization in foods. Jpn. Agric. Res. Q. 2003, 37, 167–173. [Google Scholar] [CrossRef]
- Bassa, I.A.; Francis, F.J. Stability of anthocyanins from sweet potatoes in a model beverage. J. Food Sci. 1987, 52, 1753–1754. [Google Scholar] [CrossRef]
- Drapal, M.; Rossel, G.; Heider, B.; Fraser, P.D. Metabolic diversity in sweet potato (Ipomoea batatas, L.) leaves and storage roots. Hortic. Res. 2019, 6, 2. [Google Scholar] [CrossRef] [Green Version]
- Rosero, A.; Granda, L.; Pérez, J.L.; Rosero, D.; Burgos-Paz, W.; Martínez, R.; Morelo, J.; Pastrana, I.; Burbano, E.; Morales, A. Morphometric and colourimetric tools to dissect morphological diversity: An application in sweet potato [Ipomoea batatas (L.) Lam.]. Genet. Resour. Crop Evol. 2019, 66, 1257–1278. [Google Scholar] [CrossRef] [Green Version]
- FAO. Statistics Division Production and Trade Statistics; FAO: Rome, Italy, 2017. [Google Scholar]
- Cui, R.; Zhu, F. Physicochemical properties and bioactive compounds of different varieties of sweetpotato flour treated with high hydrostatic pressure. Food Chem. 2019, 299, 125129. [Google Scholar] [CrossRef] [PubMed]
- De Albuquerque, T.M.R.; Sampaio, K.B.; de Souza, E.L. Sweet potato roots: Unrevealing an old food as a source of health promoting bioactive compounds–A review. Trends Food Sci. Technol. 2019, 85, 277–286. [Google Scholar] [CrossRef]
- Guo, K.; Liu, T.; Xu, A.; Zhang, L.; Bian, X.; Wei, C. Structural and functional properties of starches from root tubers of white, yellow, and purple sweet potatoes. Food Hydrocoll. 2019, 89, 829–836. [Google Scholar] [CrossRef]
- Wang, S.; Pan, D.; Lv, X.; Song, X.; Qiu, Z.; Huang, C.; Huang, R.; Chen, W. Proteomic approach reveals that starch degradation contributes to anthocyanin accumulation in tuberous root of purple sweet potato. J. Proteom. 2016, 143, 298–305. [Google Scholar] [CrossRef]
- Zhao, J.G.; Yan, Q.Q.; Xue, R.Y.; Zhang, J.; Zhang, Y.Q. Isolation and identification of colourless caffeoyl compounds in purple sweet potato by HPLC-DAD-ESI/MS and their antioxidant activities. Food Chem. 2014, 161, 22–26. [Google Scholar] [CrossRef]
- Kim, M.Y.; Lee, B.W.; Lee, H.; Lee, Y.Y.; Kim, M.H.; Lee, J.Y.; Lee, B.K.; Woo, K.S.; Kim, H. Phenolic compounds and antioxidant activity of sweet potato after heat treatment. J. Sci. Food Agric. 2019. [Google Scholar] [CrossRef]
- Moumouni Koala, A.H.; Somé, K.; Palé, E.; Sérémé, A.; Belem, J.; Nacro, M. Evaluation of eight orange fleshed sweetpotato (OFSP) varieties for their total antioxidant, total carotenoid and polyphenolic contents. Evaluation 2013, 3, 67–73. [Google Scholar]
- Neela, S.; Fanta, S.W. Review on nutritional composition of orange-fleshed sweet potato and its role in management of vitamin A deficiency. Food Sci. Nutr. 2019, 7, 1920–1945. [Google Scholar] [CrossRef] [Green Version]
- Lebot, V.; Michalet, S.; Legendre, L. Identification and quantification of phenolic compounds responsible for the antioxidant activity of sweet potatoes with different flesh colours using high performance thin layer chromatography (HPTLC). J. Food Compos. Anal. 2016, 49, 94–101. [Google Scholar] [CrossRef]
- Kubow, S.; Iskandar, M.M.; Sabally, K.; Azadi, B.; Sadeghi, S.; Kumarathasan, P.; Dhar, D.; Prakash, S.; Burgos, G. Biotransformation of anthocyanins from two purple-fleshed sweet potato accessions in a dynamic gastrointestinal system. Food Chem. 2016, 192, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.J.; Tan, C.; Feng, Y. Solvent extraction and in vitro simulated gastrointestinal digestion of phenolic compounds from purple sweet potato. Int. J. Food Sci. Technol. 2019, 2887–2896. [Google Scholar] [CrossRef]
- Oliveira, H.; Perez-Gregório, R.; de Freitas, V.; Mateus, N.; Fernandes, I. Comparison of the in vitro gastrointestinal bioavailability of acylated and non-acylated anthocyanins: Purple-fleshed sweet potato vs red wine. Food Chem. 2019, 276, 410–418. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.W.; Tang, C.E.; Zhang, J.L.; Zhou, Q.; Zhang, Z.C. Stability and antioxidant activity of anthocyanins from purple sweet potato (Ipomoea batatas L. cultivar Eshu No. 8) subjected to simulated in vitro gastrointestinal digestion. Int. J. Food Sci. Technol. 2019, 2604–2614. [Google Scholar] [CrossRef]
- Sun, H.; Zhang, P.; Zhu, Y.; Lou, Q.; He, S. Antioxidant and prebiotic activity of five peonidin-based anthocyanins extracted from purple sweet potato (Ipomoea batatas (L.) Lam.). Sci. Rep. 2018, 8, 5018. [Google Scholar] [CrossRef] [PubMed]
- Esatbeyoglu, T.; Rodríguez-werner, M.; Schlösser, A.; Winterhalter, P.; Rimbach, G. Fractionation, enzyme inhibitory and cellular antioxidant activity of bioactives from purple sweet potato (Ipomoea batatas). Food Chem. 2017, 221, 447–456. [Google Scholar] [CrossRef]
- Gérard, V.; Ay, E.; Morlet-Savary, F.; Graff, B.; Galopin, C.; Ogren, T.; Mutilangi, W.; Lalevée, J. Thermal and photochemical stability of anthocyanins from black carrot, grape juice, and purple sweet potato in model beverages in the presence of ascorbic acid. J. Agric. Food Chem. 2019, 67, 5647–5660. [Google Scholar] [CrossRef]
- Gras, C.C.; Nemetz, N.; Carle, R.; Schweiggert, R.M. Anthocyanins from purple sweet potato (Ipomoea batatas (L.) Lam.) and their color modulation by the addition of phenolic acids and food-grade phenolic plant extracts. Food Chem. 2017, 235, 265–274. [Google Scholar] [CrossRef]
- He, W.; Zeng, M.; Chen, J.; Jiao, Y.; Niu, F.; Tao, G.; Zhang, S.; Qin, F.; He, Z. Identification and quantitation of anthocyanins in purple-fleshed sweet potatoes cultivated in China by UPLC-PDA and UPLC-QTOF-MS/MS. J. Agric. Food Chem. 2016, 64, 171–177. [Google Scholar] [CrossRef]
- Truong, V.D.; Nigel, D.; Thompson, R.T.; Mcfeeters, R.F.; Dean, L.O.; Pecota, K.V.; Yencho, G.C. Characterization of anthocyanins and anthocyanidins in purple-fleshed sweetpotatoes by HPLC-DAD/ESI-MS/MS. J. Agric. Food Chem. 2010, 58, 404–410. [Google Scholar] [CrossRef]
- Li, J.; Li, X.D.; Zhang, Y.; Zheng, Z.D.; Qu, Z.Y.; Liu, M.; Zhu, S.H.; Liu, S.; Wang, M.; Qu, L. Identification and thermal stability of purple-fleshed sweet potato anthocyanins in aqueous solutions with various pH values and fruit juices. Food Chem. 2013, 136, 1429–1434. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.; Song, L.; Qian, B.; Xu, W.; Ren, J.; Jing, P.; Oey, I. Understanding the effect of anthocyanins extracted from purple sweet potatoes on alcohol-induced liver injury in mice. Food Chem. 2018, 245, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.H.; Kim, H.W.; Kim, S.Y.; Kim, S.M.; Kim, J.B.; Lee, Y.M. In vitro and in vivo hypoglycemic effects of cyanidin 3-caffeoyl-p-hydroxybenzoylsophoroside-5-glucoside, an anthocyanin isolated from purple-fleshed sweet potato. Food Chem. 2019, 272, 688–693. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhao, Y.; Zhou, Q.; Luo, C.L.; Deng, A.P.; Zhang, Z.C.; Zhang, J.L. Characterization and hepatoprotective activity of anthocyanins from purple sweet potato (Ipomoea batatas L. cultivar Eshu No. 8). J. Food Drug Anal. 2017, 25, 607–618. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Su, X.; Lim, S.; Griffin, J.; Carey, E.; Katz, B.; Tomich, J.; Smith, J.S.; Wang, W. Characterisation and stability of anthocyanins in purple-fleshed sweet potato P40. Food Chem. 2015, 186, 90–96. [Google Scholar] [CrossRef] [Green Version]
- Laksmiani, N.P.L.; Widiastari, M.I.; Reynaldi, K.R. The inhibitory activity of peonidin purple sweet potato in human epidermal receptor-2 receptor (her-2) expression by in silico study. J. Phys. Conf. Ser. 2018, 1040, 012010. [Google Scholar] [CrossRef] [Green Version]
- Luo, C.; Zhou, Q.; Yang, Z.; Wang, R.; Zhang, J. Evaluation of structure and bioprotective activity of key high molecular weight acylated anthocyanin compounds isolated from the purple sweet potato (Ipomoea batatas L. cultivar Eshu No. 8 ). Food Chem. 2018, 241, 23–31. [Google Scholar] [CrossRef]
- Lu, J.; Wu, D.M.; Zheng, Y.L.; Hu, B.; Cheng, W.; Zhang, Z.F. Purple sweet potato color attenuates domoic acid-induced cognitive deficits by promoting estrogen receptor-α-mediated mitochondrial biogenesis signaling in mice. Free Radic. Biol. Med. 2012, 52, 646–659. [Google Scholar] [CrossRef]
- Zhuang, J.; Lu, J.; Wang, X.; Wang, X.; Hu, W.; Hong, F.; Zhao, X.X.; Zheng, Y. lin Purple sweet potato color protects against high-fat diet-induced cognitive deficits through AMPK-mediated autophagy in mouse hippocampus. J. Nutr. Biochem. 2019, 65, 35–45. [Google Scholar] [CrossRef]
- Li, J.; Shi, Z.; Mi, Y. Purple sweet potato color attenuates high fat-induced neuroinflammation in mouse brain by inhibiting mapk and NF-κB activation. Mol. Med. Rep. 2018, 17, 4823–4831. [Google Scholar] [CrossRef]
- Li, W.L.; Yu, H.Y.; Zhang, X.J.; Ke, M.; Hong, T. Purple sweet potato anthocyanin exerts antitumor effect in bladder cancer. Oncol. Rep. 2018, 40, 73–82. [Google Scholar] [CrossRef]
- Schmitt, E.; Stopper, H. Estrogenic activity of naturally occurring anthocyanidins. Nutr. Cancer 2001, 41, 145–149. [Google Scholar] [CrossRef]
- Yoshimoto, M.; Okuno, S.; Yamaguchi, M.; Yamakawa, O. Antimutagenicity of deacylated anthocyanins in purple-fleshed sweetpotato. Biosci. Biotechnol. Biochem. 2001, 65, 1652–1655. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.G.; Yan, Q.Q.; Lu, L.Z.; Zhang, Y.Q. In vivo antioxidant, hypoglycemic, and anti-tumor activities of anthocyanin extracts from purple sweet potato. Nutr. Res. Pract. 2013, 7, 359–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Zhou, Q.; Yang, Y.; Wang, Y.; Zhang, J. Highly Acylated Anthocyanins from purple sweet potato (Ipomoea batatas L.) alleviate hyperuricemia and kidney inflammation in hyperuricemic mice: Possible attenuation effects on allopurinol. J. Agric. Food Chem. 2019, 67, 6202–6211. [Google Scholar] [CrossRef] [PubMed]
- Zheng, G.H.; Shan, Q.; Mu, J.J.; Wang, Y.J.; Zhang, Z.F.; Fan, S.H.; Hu, B.; Li, M.Q.; Xie, J.; Chen, P.; et al. Purple sweet potato color attenuates kidney damage by blocking VEGFR2/ROS/NLRP3 signaling in high-fat diet-treated mice. Oxid. Med. Cell. Longev. 2019, 2019. [Google Scholar] [CrossRef]
- Jiang, Z.; Chen, C.; Xie, W.; Wang, M.; Wang, J.; Zhang, X. Anthocyanins attenuate alcohol-induced hepatic injury by inhibiting pro-inflammation signalling. Nat. Prod. Res. 2016, 30, 469–473. [Google Scholar] [CrossRef]
- Valenti, L.; Riso, P.; Mazzocchi, A.; Porrini, M.; Fargion, S.; Agostoni, C. Dietary anthocyanins as nutritional therapy for nonalcoholic fatty liver disease. Oxid. Med. Cell. Longev. 2013, 2013. [Google Scholar] [CrossRef] [Green Version]
- Del Rio, D.; Rodriguez-Mateos, A.; Spencer, J.P.E.; Tognolini, M.; Borges, G.; Crozier, A. Dietary (poly)phenolics in human health: structures, bioavailability, and evidence of protective effects against chronic diseases. Antioxid. Redox Signal. 2012, 18, 1818–1892. [Google Scholar] [CrossRef] [Green Version]
- Han, K.H.; Shimada, K.I.; Sekikawa, M.; Fukushima, M. Anthocyanin-rich red potato flakes affect serum lipid peroxidation and hepatic SOD mRNA level in rats. Biosci. Biotechnol. Biochem. 2007, 71, 1356–1359. [Google Scholar] [CrossRef]
- Hwang, Y.P.; Choi, J.H.; Choi, J.M.; Chung, Y.C.; Jeong, H.G. Protective mechanisms of anthocyanins from purple sweet potato against tert-butyl hydroperoxide-induced hepatotoxicity. Food Chem. Toxicol. 2011, 49, 2081–2089. [Google Scholar] [CrossRef] [PubMed]
- Suda, I.; Ishikawa, F.; Hatakeyama, M.; Miyawaki, M.; Kudo, T.; Hirano, K.; Ito, A.; Yamakawa, O.; Horiuchi, S. Intake of purple sweet potato beverage affects on serum hepatic biomarker levels of healthy adult men with borderline hepatitis. Eur. J. Clin. Nutr. 2008, 62, 60–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.F.; Lu, J.; Zheng, Y.L.; Wu, D.M.; Hu, B.; Shan, Q.; Cheng, W.; Li, M.Q.; Sun, Y.Y. Purple sweet potato color attenuates hepatic insulin resistance via blocking oxidative stress and endoplasmic reticulum stress in high-fat-diet-treated mice. J. Nutr. Biochem. 2013, 24, 1008–1018. [Google Scholar] [CrossRef] [PubMed]
- Qin, S.; Sun, D.; Mu, J.; Ma, D.; Tang, R.; Zheng, Y. Purple sweet potato color improves hippocampal insulin resistance via down-regulating SOCS3 and galectin-3 in high-fat diet mice. Behav. Brain Res. 2019, 359, 370–377. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, Z.F.; Zheng, G.H.; Wang, A.M.; Sun, C.H.; Qin, S.P.; Zhuang, J.; Lu, J.; Ma, D.F.; Zheng, Y.L. The inhibitory effects of purple sweet potato color on hepatic inflammation is associated with restoration of NAD+ levels and attenuation of nlrp3 inflammasome activation in high-fat-diet-treated mice. Molecules 2017, 22, 1315. [Google Scholar] [CrossRef] [PubMed]
- Gofur, A.; Witjoro, A.; Ningtiyas, E.W.; Setyowati, E.; Mukharromah, S.A.; Athoillah, M.F.; Lestari, S.R. The evaluation of dietary black soybean and purple sweet potato on insulin sensitivity in streptozotocin-induced diabetic rats. Pharmacogn. J. 2019, 11, 639–646. [Google Scholar] [CrossRef] [Green Version]
- Park, S.-J.; Lee, D.; Lee, M.; Kwon, H.-O.; Kim, H.; Park, J.; Jeon, W.; Cha, M.; Jun, S.; Park, K.; et al. The Effects of Curcuma longa L., Purple sweet potato, and mixtures of the two on immunomodulation in C57BL/6J mice infected with LP-BM5 murine leukemia retrovirus. J. Med. Food 2018, 21, 689–700. [Google Scholar] [CrossRef]
- Ju, R.; Zheng, S.; Luo, H.; Wang, C.; Duan, L.; Sheng, Y.; Zhao, C.; Xu, W.; Huang, K. Purple Sweet Potato Attenuate Weight Gain in High Fat Diet Induced Obese Mice. J. Food Sci. 2017, 82, 787–793. [Google Scholar] [CrossRef]
- Chen, H.; Sun, J.; Liu, J.; Gou, Y.; Zhang, X.; Wu, X.; Sun, R.; Tang, S.; Kan, J.; Qian, C.; et al. Structural characterization and anti-inflammatory activity of alkali-soluble polysaccharides from purple sweet potato. Int. J. Biol. Macromol. 2019, 131, 484–494. [Google Scholar] [CrossRef]
- Gou, Y.; Sun, J.; Liu, J.; Chen, H.; Kan, J.; Qian, C.; Zhang, N.; Jin, C. Structural characterization of a water-soluble purple sweet potato polysaccharide and its effect on intestinal inflammation in mice. J. Funct. Foods 2019, 61, 103502. [Google Scholar] [CrossRef]
- Sun, J.; Zhou, B.; Tang, C.; Gou, Y.; Chen, H.; Wang, Y.; Jin, C.; Liu, J.; Niu, F.; Kan, J.; et al. Characterization, antioxidant activity and hepatoprotective effect of purple sweetpotato polysaccharides. Int. J. Biol. Macromol. 2018, 115, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.; Sun, J.; Liu, J.; Jin, C.; Wu, X.; Zhang, X.; Chen, H.; Gou, Y.; Kan, J.; Qian, C.; et al. Immune-enhancing effects of polysaccharides from purple sweet potato. Int. J. Biol. Macromol. 2019, 123, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.; Sun, J.; Zhou, B.; Jin, C.; Liu, J.; Gou, Y.; Chen, H.; Kan, J.; Qian, C.; Zhang, N. Immunomodulatory effects of polysaccharides from purple sweet potato on lipopolysaccharide treated RAW 264.7 macrophages. J. Food Biochem. 2018, 42, e12535. [Google Scholar] [CrossRef]
- Tang, C.; Sun, J.; Zhou, B.; Jin, C.; Liu, J.; Kan, J.; Qian, C.; Zhang, N. Effects of polysaccharides from purple sweet potatoes on immune response and gut microbiota composition in normal and cyclophosphamide treated mice. Food Funct. 2018, 9, 937–950. [Google Scholar] [CrossRef]
- Jiang, T.; Mao, Y.; Sui, L.; Yang, N.; Li, S.; Zhu, Z.; Wang, C.; Yin, S.; He, J.; He, Y. Degradation of anthocyanins and polymeric color formation during heat treatment of purple sweet potato extract at different pH. Food Chem. 2019, 274, 460–470. [Google Scholar] [CrossRef]
- Quan, W.; He, W.; Lu, M.; Yuan, B.; Zeng, M.; Gao, D.; Qin, F.; Chen, J.; He, Z. Anthocyanin composition and storage degradation kinetics of anthocyanins-based natural food colourant from purple-fleshed sweet potato. Int. J. Food Sci. Technol. 2019, 54, 2529–2539. [Google Scholar] [CrossRef]
- Thi Lan Khanh, P.; Chittrakorn, S.; Rutnakornpituk, B.; Phan Tai, H.; Ruttarattanamongkol, K. Processing effects on anthocyanins, phenolic acids, antioxidant activity, and physical characteristics of Vietnamese purple-fleshed sweet potato flours. J. Food Process. Preserv. 2018, 42, e13722. [Google Scholar] [CrossRef]
- Yea, C.S.; Nevara, G.A.; Muhammad, K.; Ghazali, H.M.; Karim, R. Physical properties, resistant starch content and antioxidant profile of purple sweet potato powder after 12 months of storage. Int. J. Food Prop. 2019, 22, 974–984. [Google Scholar] [CrossRef] [Green Version]
- Lagnika, C.; Jiang, N.; Song, J.; Li, D.; Liu, C.; Huang, J.; Wei, Q.; Zhang, M. Effects of pretreatments on properties of microwave-vacuum drying of sweet potato slices. Dry. Technol. 2018, 37, 1901–1914. [Google Scholar] [CrossRef]
- Phan, K.T.L.; Chittrakorn, S.; Tai, H.P.; Ruttarattanamongkol, K. Effects of cooking methods on the changes of total anthocyanins, phenolics content and physical characteristics of purple-fleshed sweet potato (Ipomoea batatas L.) grown in Vietnam. Int. J. Adv. Sci. Eng. Inf. Technol. 2018, 8, 227–233. [Google Scholar] [CrossRef]
- Kamiloglu, S.; Van Camp, J.; Capanoglu, E. Black carrot polyphenols: Effect of processing, storage and digestion—An overview. Phytochem. Rev. 2018, 17, 379–395. [Google Scholar] [CrossRef]
- Arscott, S.A.; Tanumihardjo, S.A. Carrots of many colors provide basic nutrition and bioavailable phytochemicals acting as a functional food. Compr. Rev. Food Sci. Food Saf. 2010, 9, 223–239. [Google Scholar] [CrossRef]
- Kamiloglu, S.; Capanoglu, E.; Bilen, F.D.; Gonzales, G.B.; Grootaert, C.; Van de Wiele, T.; Van Camp, J. Bioaccessibility of polyphenols from plant-processing byproducts of black carrot (Daucus carota L.). J. Agric. Food Chem. 2016, 64, 2450–2458. [Google Scholar] [CrossRef] [PubMed]
- Kamiloglu, S.; Pasli, A.A.; Ozcelik, B.; Van Camp, J.; Capanoglu, E. Influence of different processing and storage conditions on in vitro bioaccessibility of polyphenols in black carrot jams and marmalades. Food Chem. 2015, 186, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Dahuja, A.; Sachdev, A.; Kaur, C.; Varghese, E.; Saha, S.; Sairam, K.V.S.S. Black Carrot (Daucus carota ssp.) and Black Soybean (Glycine max (L.) Merr.) Anthocyanin Extract: A Remedy to Enhance Stability and Functionality of Fruit Juices by Copigmentation, Waste and Biomass Valorization; Springer: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
- Horvitz, M.A.; Simon, P.W.; Tanumihardjo, S.A. Lycopene and β-carotene are bioavailable from lycopene “red” carrots in humans. Eur. J. Clin. Nutr. 2004, 58, 803–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kammerer, D.; Carle, R.; Schieber, A. Quantification of anthocyanins in black carrot extracts (Daucus carota ssp. sativus var. atrorubens Alef.) and evaluation of their color properties. Eur. Food Res. Technol. 2004, 219, 479–486. [Google Scholar] [CrossRef]
- Smeriglio, A.; Denaro, M.; Barreca, D.; D’Angelo, V.; Germanò, M.P.; Trombetta, D. Polyphenolic profile and biological activities of black carrot crude extract (Daucus carota L. ssp. sativus var. atrorubens Alef.). Fitoterapia 2018, 124, 49–57. [Google Scholar] [CrossRef]
- Olejnik, A.; Rychlik, J.; Kidoń, M.; Czapski, J.; Kowalska, K.; Juzwa, W.; Olkowicz, M.; Dembczyński, R.; Moyer, M.P. Antioxidant effects of gastrointestinal digested purple carrot extract on the human cells of colonic mucosa. Food Chem. 2016, 190, 1069–1077. [Google Scholar] [CrossRef]
- Blando, F.; Calabriso, N.; Berland, H.; Maiorano, G.; Gerardi, C.; Carluccio, M.A.; Andersen, Ø.M. Radical scavenging and anti-inflammatory activities of representative anthocyanin groupings from pigment-rich fruits and vegetables. Int. J. Mol. Sci. 2018, 19, 169. [Google Scholar] [CrossRef] [Green Version]
- Kammerer, D.; Carle, R.; Schieber, A. Characterization of phenolic acids in black carrots (Daucus carota ssp. sativus var. atrorubens Alef.) by high-performance liquid chromatography/ electrospray ionization mass spectrometry. Rapid Commun. Mass Spectrom. 2004, 18, 1331–1340. [Google Scholar] [CrossRef]
- Khandare, V.; Walia, S.; Singh, M.; Kaur, C. Black carrot (Daucus carota ssp. sativus) juice: Processing effects on antioxidant composition and color. Food Bioprod. Process. 2011, 89, 482–486. [Google Scholar] [CrossRef]
- Iliopoulou, I.; Thaeron, D.; Baker, A.; Jones, A.; Robertson, N. Analysis of the thermal degradation of the individual anthocyanin compounds of black carrot (Daucus carota L.): A new approach using high-resolution proton nuclear magnetic resonance spectroscopy. J. Agric. Food Chem. 2015, 63, 7066–7073. [Google Scholar] [CrossRef] [PubMed]
- Santana-Gálvez, J.; Pérez-Carrillo, E.; Velázquez-Reyes, H.H.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. Application of wounding stress to produce a nutraceutical-rich carrot powder ingredient and its incorporation to nixtamalized corn flour tortillas. J. Funct. Foods 2016, 27, 655–666. [Google Scholar] [CrossRef]
- Wang, M.; Lopez-Nieves, S.; Goldman, I.L.; Maeda, H.A. Limited tyrosine utilization explains lower betalain contents in yellow than in red table beet genotypes. J. Agric. Food Chem. 2017, 65, 4305–4313. [Google Scholar] [CrossRef]
- Herbach, K.; Stintzing, F.; Carle, R. Impact of thermal treatment on color and pigment pattern of red beet (Beta vulgaris L.) preparations. J. Food Sci. 2004, 69, 419–498. [Google Scholar] [CrossRef]
- Montes-Lora, S.; Rodríguez-Pulido, F.J.; Cejudo-Bastante, M.J.; Heredia, F.J. Implications of the red beet ripening on the colour and betalain composition relationships. Plant Foods Hum. Nutr. 2018, 73, 216–221. [Google Scholar] [CrossRef]
- Wettasinghe, M.; Bolling, B.; Plhak, L.; Xiao, H.; Parkin, K. Phase II enzyme-inducing and antioxidant activities of beetroot (Beta vulgaris L.) extracts from phenotypes of different pigmentation. J. Agric. Food Chem. 2002, 50, 6704–6709. [Google Scholar] [CrossRef]
- Vulić, J.J.; Ćebović, T.N.; Čanadanović-Brunet, J.M.; Ćetković, G.S.; Čanadanović, V.M.; Djilas, S.M.; Šaponjac, V.T.T. In vivo and in vitro antioxidant effects of beetroot pomace extracts. J. Funct. Foods 2014, 6, 168–175. [Google Scholar] [CrossRef]
- Leng, M.S.; Tobit, P.; Demasse, A.M.; Wolf, K.; Gouado, I.; Ndjouenkeu, R.; Rawel, H.M.; Schweigert, F.J. Nutritional and anti-oxidant properties of yam (Dioscorea schimperiana) based complementary food formulation. Sci. African 2019, 5, e00132. [Google Scholar] [CrossRef]
- Champagne, A.; Hilbert, G.; Legendre, L.; Lebot, V. Diversity of anthocyanins and other phenolic compounds among tropical root crops from Vanuatu, South Pacific. J. Food Compos. Anal. 2011, 24, 315–325. [Google Scholar] [CrossRef]
- Li, T.; Teng, H.; An, F.; Huang, Q.; Chen, L.; Song, H. The beneficial effects of purple yam (Dioscorea alata L.) resistant starch on hyperlipidemia in high-fat-fed hamsters. Food Funct. 2019, 10, 2642–2650. [Google Scholar] [CrossRef]
- Patel, K.S.; Karmakar, N.; Desai, K.D.; Narwade, A.V.; Chakravarty, G.; Debnath, M.K. Exploring of greater yam (Dioscorea alata L.) genotypes through biochemical screening for better cultivation in south Gujarat zone of India. Physiol. Mol. Biol. Plants 2019, 25, 1235–1249. [Google Scholar] [CrossRef]
- Li, T.; Chen, L.; Xiao, J.; An, F.; Wan, C.; Song, H. Prebiotic effects of resistant starch from purple yam (Dioscorea alata L.) on the tolerance and proliferation ability of Bifidobacterium adolescentis in vitro. Food Funct. 2018, 9, 2416–2425. [Google Scholar] [CrossRef]
- Estiasih, T.; Umaro, D.; Harijono, H. Hypoglycemic effect of crude water soluble polysaccharide extracted from tubers of purple and yellow water yam (Dioscorea alata L.) on alloxan-induced hyperglycemia Wistar rats. Prog. Nutr. 2018, 20, 59–67. [Google Scholar]
- Rosida; Purnawati, A.; Susiloningsih, E.K. Hypocholesterolemic and hypoglycemic effects of autoclaved-cooled water yam (Dioscorea alata) on hypercholesterolemia rats. Int. Food Res. J. 2018, 25, S181–S184. [Google Scholar]
- Chen, C.T.; Wang, Z.H.; Hsu, C.C.; Lin, H.H.; Chen, J.H. Taiwanese and Japanese yam (Dioscorea spp.) extracts attenuate doxorubicin-induced cardiotoxicity in mice. J. Food Drug Anal. 2017, 25, 872–880. [Google Scholar] [CrossRef] [Green Version]
- Khan, R.; Upaganlawar, A.B.; Upasani, C. Protective effects of Dioscorea alata L. in aniline exposure-induced spleen toxicity in rats: A biochemical study. Toxicol. Int. 2014, 21, 294–299. [Google Scholar]
- Chiu, C.S.; Deng, J.S.; Chang, H.Y.; Chen, Y.C.; Lee, M.M.; Hou, W.C.; Lee, C.Y.; Huang, S.S.; Huang, G.J. Antioxidant and anti-inflammatory properties of taiwanese yam (Dioscorea japonica Thunb. var. pseudojaponica (Hayata) Yamam.) and its reference compounds. Food Chem. 2013, 141, 1087–1096. [Google Scholar] [CrossRef]
- Chang, W.T.; Chen, H.M.; Yin, S.Y.; Chen, Y.H.; Wen, C.C.; Wei, W.C.; Lai, P.; Wang, C.H.; Yang, N.S. Specific dioscorea phytoextracts enhance potency of TCL-loaded DC-based cancer vaccines. Evidence-Based Complement. Altern. Med. 2013, 2013. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.F.; Chang, S.Y.; Lee, T.C.; Chuang, L.Y.; Guh, J.Y.; Hung, C.Y.; Hung, T.J.; Hung, Y.J.; Chen, P.Y.; Hsieh, P.F.; et al. Dioscorea alata attenuates renal interstitial cellular fibrosis by regulating smad- and epithelial-mesenchymal transition signaling pathways. PLoS ONE 2012, 7, e47482. [Google Scholar] [CrossRef] [Green Version]
- Chan, Y.C.; Chang, S.C.; Liu, S.Y.; Yang, H.L.; Hseu, Y.C.; Liao, J.W. Beneficial effects of yam on carbon tetrachloride-induced hepatic fibrosis in rats. J. Sci. Food Agric. 2010, 90, 161–167. [Google Scholar] [CrossRef]
- Wang, C.-H.; Tsai, C.-H.; Lin, H.-J.; Wang, T.-C.; Chen, H.-L. Uncooked Taiwanese yam (Dioscorea alata L. cv. Tainung No. 2) beneficially modulated the large bowel function and faecal microflora in BALB/c mice. J. Sci. Food Agric. 2006, 87, 1374–1380. [Google Scholar] [CrossRef]
- Hsu, C.C.; Huang, Y.C.; Yin, M.C.; Lin, S.J. Effect of yam (Dioscorea alata compared to Dioscorea japonica) on gastrointestinal function and antioxidant activity in mice. J. Food Sci. 2006, 71, 5–8. [Google Scholar] [CrossRef]
- Chan, Y.-C.; Hsu, C.-K.; Wang, M.-F.; Liao, J.-W.; Su, T.-Y. Beneficial effect of yam on the amyloid β-protein, monoamine oxidase B and cognitive deficit in mice with accelerated senescence. J. Sci. Food Agric. 2006, 86, 1517–1525. [Google Scholar] [CrossRef]
- Chan, Y.C.; Hsu, C.K.; Wang, M.F.; Su, T.Y. A diet containing yam reduces the cognitive deterioration and brain lipid peroxidation in mice with senescence accelerated. Int. J. Food Sci. Technol. 2004, 39, 99–107. [Google Scholar] [CrossRef]
- Fu, S.L.; Hsu, Y.H.; Lee, P.Y.; Hou, W.C.; Hung, L.C.; Lin, C.H.; Chen, C.M.; Huang, Y.J. Dioscorin isolated from Dioscorea alata activates TLR4-signaling pathways and induces cytokine expression in macrophages. Biochem. Biophys. Res. Commun. 2006, 339, 137–144. [Google Scholar] [CrossRef]
- Mahmad, N.; Taha, R.M.; Othman, R.; Abdullah, S.; Anuar, N.; Elias, H.; Rawi, N. Anthocyanin as potential source for antimicrobial activity in Clitoria ternatea L. and Dioscorea alata L. Pigment Resin Technol. 2018, 47, 490–495. [Google Scholar] [CrossRef]
- Chen, T.; Hu, S.; Zhang, H.; Guan, Q.; Yang, Y.; Wang, X. Anti-inflammatory effects of Dioscorea alata L. anthocyanins in a TNBS-induced colitis model. Food Funct. 2017, 8, 659–669. [Google Scholar] [CrossRef]
- Guo, X.; Sha, X.; Cai, S.; Wang, O.; Ji, B. Antiglycative and antioxidative properties of ethyl acetate fraction of Chinese purple yam (Dioscorea alata L.) extracts. Food Sci. Technol. Res. 2015, 21, 563–571. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.X.; Sha, X.H.; Liu, J.; Cai, S.B.; Wang, Y.; Ji, B.P. Chinese purple yam (Dioscorea alata L.) extracts inhibit diabetes-related enzymes and protect HepG2 cells against oxidative stress and insulin resistance induced by FFA. Food Sci. Technol. Res. 2015, 21, 677–683. [Google Scholar] [CrossRef] [Green Version]
- Mahmoudian-Sani, M.R.; Asadi-Samani, M.; Luther, T.; Saeedi-Boroujeni, A.; Gholamian, N. A new approach for treatment of type 1 diabetes: Phytotherapy and phytopharmacology of regulatory T cells. J. Ren. Inj. Prev. 2017, 6, 158–163. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Tian, H.; Zhan, P.; Du, F.; Zong, A.; Xu, T. Isolation and identification of phenolic compounds in Chinese purple yam and evaluation of antioxidant activity. LWT Food Sci. Technol. 2018, 96, 161–165. [Google Scholar] [CrossRef]
- Dey, P.; Ray, S.; Chaudhuri, T.K. Immunomodulatory activities and phytochemical characterisation of the methanolic extract of Dioscorea alata aerial tuber. J. Funct. Foods 2016, 23, 315–328. [Google Scholar] [CrossRef]
- Lu, J.; Wong, R.N.S.; Zhang, L.; Wong, R.Y.L.; Ng, T.B.; Lee, K.F.; Zhang, Y.B.; Lao, L.X.; Liu, J.Y.; Sze, S.C.W. Comparative Analysis of Proteins with Stimulating Activity on Ovarian Estradiol Biosynthesis from Four Different Dioscorea Species in vitro Using Both Phenotypic and Target-based Approaches: Implication for Treating Menopause. Appl. Biochem. Biotechnol. 2016, 180, 79–93. [Google Scholar] [CrossRef]
- Wu, Z.G.; Jiang, W.; Nitin, M.; Bao, X.Q.; Chen, S.L.; Tao, Z.M. Characterizing diversity based on nutritional and bioactive compositions of yam germplasm (Dioscorea spp.) commonly cultivated in China. J. Food Drug Anal. 2016, 24, 367–375. [Google Scholar] [CrossRef] [Green Version]
- Han, C.H.; Liu, J.C.; Fang, S.U.; Hou, W.C. Antioxidant activities of the synthesized thiol-contained peptides derived from computer-aided pepsin hydrolysis of yam tuber storage protein, dioscorin. Food Chem. 2013, 138, 923–930. [Google Scholar] [CrossRef]
- Koo, H.J.; Park, H.J.; Byeon, H.E.; Kwak, J.H.; Um, S.H.; Kwon, S.T.; Rhee, D.K.; Pyo, S. Chinese yam extracts containing β-sitosterol and ethyl linoleate protect against atherosclerosis in apolipoprotein e-deficient mice and inhibit muscular expression of VCAM-1 in vitro. J. Food Sci. 2014, 79, H719–H729. [Google Scholar] [CrossRef]
- Harijono, T.E.; Ariestiningsih, A.D.; Wardani, N.A.K. The effect of crude diosgenin extract from purple and yellow greater yams (Dioscorea alata) on the lipid profile of dyslipidemia rats. Emirates J. Food Agric. 2016, 28, 506–512. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.C.; Lin, J.T.; Wang, C.K.; Chen, C.H.; Yang, D.J. Antiproliferative effects of fractions of furostanol and spirostanol glycosides from yam (Dioscorea pseudojaponica Yamamoto) and diosgenin on cancer and normal cells and their apoptotic effects for MCF-7 cells. J. Food Biochem. 2012, 36, 75–85. [Google Scholar] [CrossRef]
- Jailani, F.; Kusumawardani, S.; Puspitasari, C.; Maula, A.; Purwandari, U. Annealled purple yam (Dioscorea alata var. purpurea) flour improved gelatinisation profile, but increased glycemic index of substituted bread. Int. Food Res. J. 2013, 20, 865–871. [Google Scholar]
- Hsu, C.L.; Hurang, S.L.; Chen, W.; Weng, Y.M.; Tseng, C.Y. Qualities and antioxidant properties of bread as affected by the incorporation of yam flour in the formulation. Int. J. Food Sci. Technol. 2004, 39, 231–238. [Google Scholar] [CrossRef]
- Li, P.H.; Huang, C.C.; Yang, M.Y.; Wang, C.C.R. Textural and sensory properties of salted noodles containing purple yam flour. Food Res. Int. 2012, 47, 223–228. [Google Scholar] [CrossRef]
- Seguchi, M.; Ozawa, M.; Nakamura, C.; Tabara, A. Development of gluten-free bread baked with yam flour. Food Sci. Technol. Res. 2012, 18, 543–548. [Google Scholar] [CrossRef] [Green Version]
- Brewster, J.L. Onions and Other Vegetable Alliums, 2nd ed.; CABI: Wallingford, UK, 2008; ISBN 9781845933999. [Google Scholar]
- Slimestad, R.; Fossen, T.; Vågen, I.M. Onions: A source of unique dietary flavonoids. J. Agric. Food Chem. 2007, 55, 10067–10080. [Google Scholar] [CrossRef]
- Benkeblia, N. Free-Radical Scavenging capacity and antioxidant properties of some selected onions (Allium cepa L.) and garlic ( Allium sativum L.) extracts. Brazilian Arch. Biol. Technol. 2005, 48, 753–759. [Google Scholar] [CrossRef]
- Zhang, S.; Deng, P.; Xu, Y.C.; Lü, S.W.; Wang, J.J. Quantification and analysis of anthocyanin and flavonoids compositions, and antioxidant activities in onions with three different colors. J. Integr. Agric. 2016, 15, 2175–2181. [Google Scholar] [CrossRef] [Green Version]
- Oboh, G.; Ademiluyi, A.O.; Agunloye, O.M.; Ademosun, A.O.; Ogunsakin, B.G. Inhibitory Effect of garlic, purple onion, and white onion on key enzymes linked with type 2 diabetes and hypertension. J. Diet. Suppl. 2019, 16, 105–118. [Google Scholar] [CrossRef]
- Albishi, T.; John, J.A.; Al-Khalifa, A.S.; Shahidi, F. Antioxidant, anti-inflammatory and DNA scission inhibitory activities of phenolic compounds in selected onion and potato varieties. J. Funct. Foods 2013, 5, 930–939. [Google Scholar] [CrossRef]
- Ou, B.; Huang, D.; Hampsch-Woodill, M.; Flanagan, J.A.; Deemer, E.K. Analysis of antioxidant activities of common vegetables employing oxygen radical absorbance capacity (ORAC) and ferric reducing antioxidant power (FRAP) assays: A comparative study. J. Agric. Food Chem. 2002, 50, 3122–3128. [Google Scholar] [CrossRef]
- Khiari, Z.; Makris, D.P.; Kefalas, P. An Investigation on the Recovery of Antioxidant Phenolics from Onion Solid Wastes Employing Water/Ethanol-Based Solvent Systems. Food Process. Technol. 2009, 2, 337–343. [Google Scholar] [CrossRef]
- Hanlon, P.R.; Barnes, D.M. Phytochemical composition and biological activity of 8 varieties of radish (Raphanus sativus L.) sprouts and mature taproots. J. Food Sci. 2011, 76, 185–192. [Google Scholar] [CrossRef]
- Masukawa, T.; Kadowaki, M.; Matsumoto, T.; Nakatsuka, A.; Cheon, K.S.; Kato, K.; Tatsuzawa, F.; Kobayashi, N. Enhancement of food functionality of a local pungent radish “izumo orochi daikon” ‘susanoo’ by introduction of a colored root character. Hortic. J. 2018, 87, 356–363. [Google Scholar] [CrossRef]
- Tatsuzawa, F.; Saito, N.; Toki, K.; Shinoda, K.; Shigihara, A.; Honda, T. Acylated cyanidin 3-sophoroside-5-glucosides from the purple roots of red radish (Raphanus sativus L.) “Benikanmi.”. J. Jpn. Soc. Hortic. Sci. 2010, 79, 103–107. [Google Scholar] [CrossRef]
- Ahn, M.; Koh, R.; Kim, G.O.; Shin, T. Aqueous extract of purple Bordeaux radish, Raphanus sativus L. ameliorates ethanol-induced gastric injury in rats. Orient. Pharm. Exp. Med. 2013, 13, 247–252. [Google Scholar] [CrossRef]
- Pak, W.M.; Kim, K.B.W.R.; Kim, M.J.; Kang, B.K.; Bark, S.W.; Kim, B.R.; Ahn, N.K.; Choi, Y.U.; Yoon, S.R.; Ahn, D.H. Antioxidative effect of extracts from different parts of Kohlrabi. J. Appl. Biol. Chem. 2014, 57, 353–358. [Google Scholar] [CrossRef]
- Jung, H.A.; Karki, S.; Ehom, N.; Yoon, M.; Kim, E.J.; Choi, J.S. Anti-diabetic and anti-inflammatory effects of green and red kohlrabi cultivars (Brassica oleracea var. gongylodes). 2014, 19, 281–290. [Google Scholar] [CrossRef] [Green Version]
- Paull, R.E.; Uruu, G.; Arakaki, A. Variation in the cooked and chipping quality of taro. Horttechnology 2000, 10, 823–829. [Google Scholar] [CrossRef] [Green Version]
- Beyene, T.M. Morpho−Agronomical Characterization of Taro (Colocasia esculenta) Accessions in Ethiopia. Plant 2013, 1, 1–9. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Edible Part | Color | Class of Compounds | Compounds | References |
|---|---|---|---|---|---|
| Potato (Solanum tuberosum L.) | Tuber (stem tuber) | Purple | Petunidin derivatives | Petunidin-3-p-coumaroylrutinoside-5-glucoside, petunidin-2-p-coumarylrutinoside-5-glucoside | [21,22,23,24] |
| Red | Pelargonidin, delphinidin, cyanidin, peonidin, and malvidin acyl-glycoside derivatives | Pelargonidin-3-p-coumaroylrutinoside-5-glucoside | [21,23] | ||
| Purple/red | Carotenoids | Neoxanthin, violaxanthin | [25] | ||
| Yellow | Carotenoids | Antheraxanthin | [25] | ||
| Sweet potato (Ipomoea batatas L. Lam.) | Tuberous root (root tuber) | Purple | Acylated anthocyanins | Cyanidin, peonidin, and pelargonidin derivatives | [26,27,28,29,30] |
| Carrot (Daucus carota L. ssp. sativus Hoffm.) | Taproot (swollen hypocotyl and root) | Purple or black | Cyanidin derivatives | Acylated cyanidin 3-xylosyl(glucosyl)galactosides with sinapic acid, ferulic acid, and coumaric acid; | [31,32,33,34,35] |
| Vinylphenol and vinylguaiacol adducts of cyanidin derivatives | [36] | ||||
| Red and yellow | Carotenoids | Lycopene and β-carotene | [37,38] | ||
| Beet root (Beta vulgaris L.) | Root (swollen hypocotyl and root) | Purple | Betalains | Betacyanins | [39,40] |
| Yellow | Betalains | Betaxanthins | [41] | ||
| Vulgaxanthin I and betanin | [42] | ||||
| Yam (Dioscurea sp. L.) | Tuber (stem tuber) | Purple | Cyanidin, pelargonidin, and peonidin-type compounds; alatanins A–C | Cyanidin 3-hexoside acylated with two hydroxycinnamic acids, cyanidin 3-glycoside acylated with one hydroxycinnamic acid, cyanidin 3-glycoside acylated with one hydroxycinnamic acid, peonidin 3-glycoside acylated with one hydroxycinnamic acid, alatanin-C | [43,44,45,46] |
| Yellow | Carotenoids | β-Carotene | [47] | ||
| Onion (Allium cepa L.) | Bulb (swollen basis of leaves) | Purple | Flavonols and acylated and non-acylated cyanidin glucosides | Dihydroflavonol taxifolin and its 3-, 7-, and 4′-glucosides | [48,49] |
| Radish (Raphanus sativus L.) | Taproot (swollen root and hypocotyl) | Purple | Cyanidin glucosides | Cyanidin 3-(glucosylacyl)acylsophoroside-5-diglucosides, cyanidin 3-sophoroside-5-diglucosides, cyanidin 3-sophoroside-5-glucosides, cyanidin 3-O-[2-O-(β-glucopyranosyl)-6-O-(trans-feruloyl)-β-glucopyranoside]-5-O-[6-O-(malonyl)-β-glucopyranoside] cyanidin 3-[2-(glucosyl)-6-(cis-p-coumaroyl)-glucoside]-5-[6-(malonyl)-glucoside] | [50,51] |
| Red | Anthocyanins | Pelargonidin 3-sophoroside-5-glucoside, pelargonidin 3-[2-(glucosyl)-6-(trans-p-coumaroyl)-glucoside]-5-glucoside, pelargonidin 3-[2-(glucosyl)-6-(trans-feruloyl)-glucoside]-5-glucoside, pelargonidin 3-[2-(glucosyl)-6-(trans-p-coumaroyl)-glucoside]-5-(6-malonylglucoside), pelargonidin 3-[2-(glucosyl)-6-(trans-feruloyl)-glucoside]-5-(6-malonylglucoside), 3-O-[2-O-(b-d-glucopyranosyl)-6-O-(trans-caffeoyl)-b-d-glucopyr-anoside]-5-O-(6-O-malonyl-b-d-glucopyranoside), pelargonidin 3-O-[2-O-(b-d-glucopyranosyl)-6-O-(cis-p-cou-maroyl)-b-d-glucopyranoside]-5-O-(6-O-malonyl-b-d-glucopyranoside | [52,53] | ||
| Kohlrabi (Brassica oleracea var. gongylodes) | Swollen epicotyl | Purple | Cyanidin and cyanidin glucoside | Cyanidin-3-diglucoside-5-glucoside, cyanidin-3-(sinapoyl)-diglucoside-5-glucoside, cyanidin 3-(feruloyl) (sinapoyl) diglucoside-5-glucoside | [54,55,56,57] |
| Taro (Colocasia esculenta) | Corm | Purple | Cyanidin and pelargonidin glucosides | [58] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petropoulos, S.A.; Sampaio, S.L.; Di Gioia, F.; Tzortzakis, N.; Rouphael, Y.; Kyriacou, M.C.; Ferreira, I. Grown to be Blue—Antioxidant Properties and Health Effects of Colored Vegetables. Part I: Root Vegetables. Antioxidants 2019, 8, 617. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox8120617
Petropoulos SA, Sampaio SL, Di Gioia F, Tzortzakis N, Rouphael Y, Kyriacou MC, Ferreira I. Grown to be Blue—Antioxidant Properties and Health Effects of Colored Vegetables. Part I: Root Vegetables. Antioxidants. 2019; 8(12):617. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox8120617
Chicago/Turabian StylePetropoulos, Spyridon A., Shirley L. Sampaio, Francesco Di Gioia, Nikos Tzortzakis, Youssef Rouphael, Marios C. Kyriacou, and Isabel Ferreira. 2019. "Grown to be Blue—Antioxidant Properties and Health Effects of Colored Vegetables. Part I: Root Vegetables" Antioxidants 8, no. 12: 617. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox8120617