Biopotential of Bersama abyssinica Fresen Stem Bark Extracts: UHPLC Profiles, Antioxidant, Enzyme Inhibitory, and Antiproliferative Propensities



 , , , ,
, , , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Preparation of Extracts
2.2. Profile of Bioactive Compounds
2.3. Determination of Antioxidant and Enzyme Inhibitory Effects
2.4. Artemia Salina Lethality Test
2.5. Human Colon Cancer HCT116 Cell Culture
2.6. Statistical Analysis
3. Results and Discussion
3.1. Phytochemical Profile
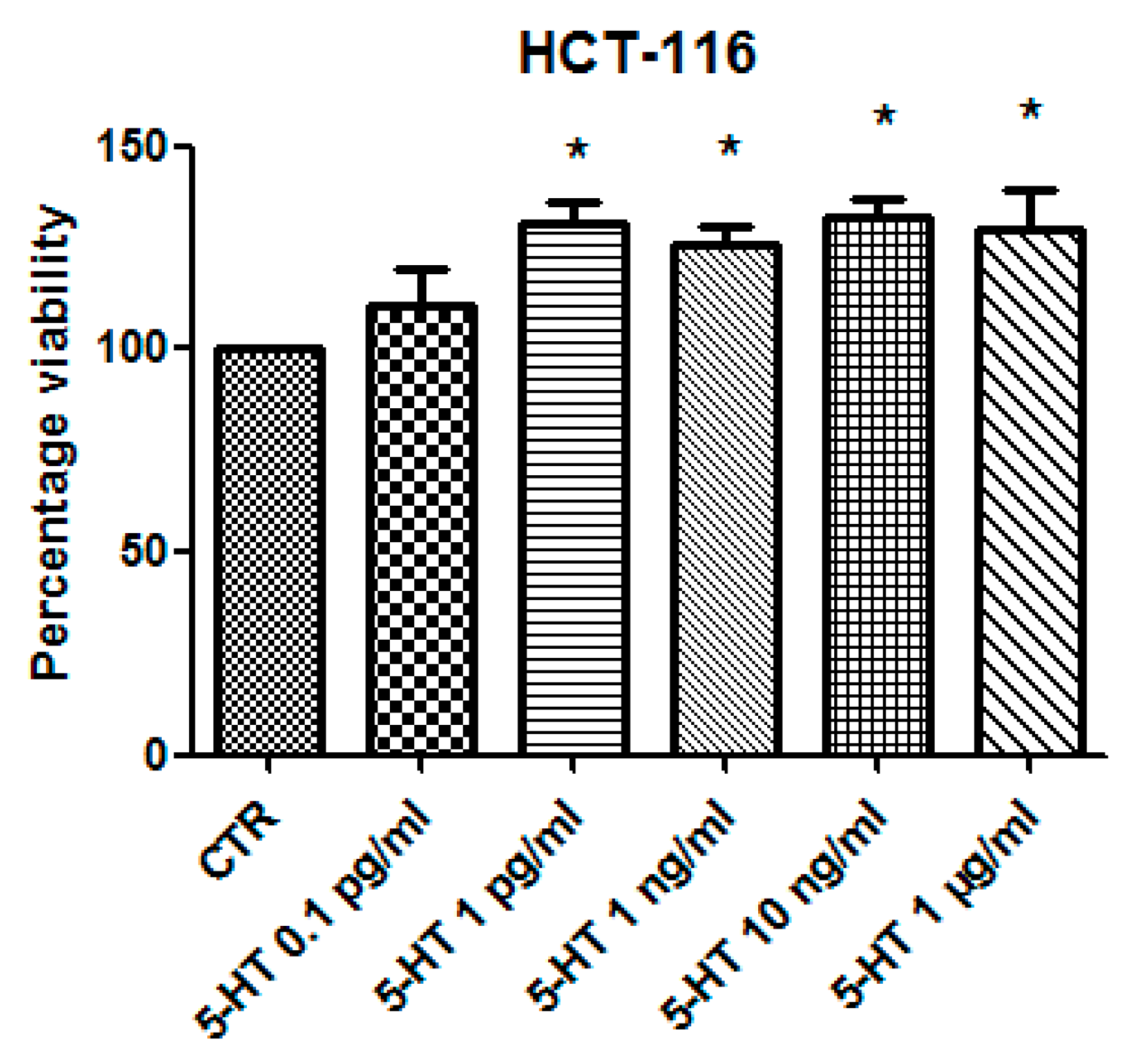
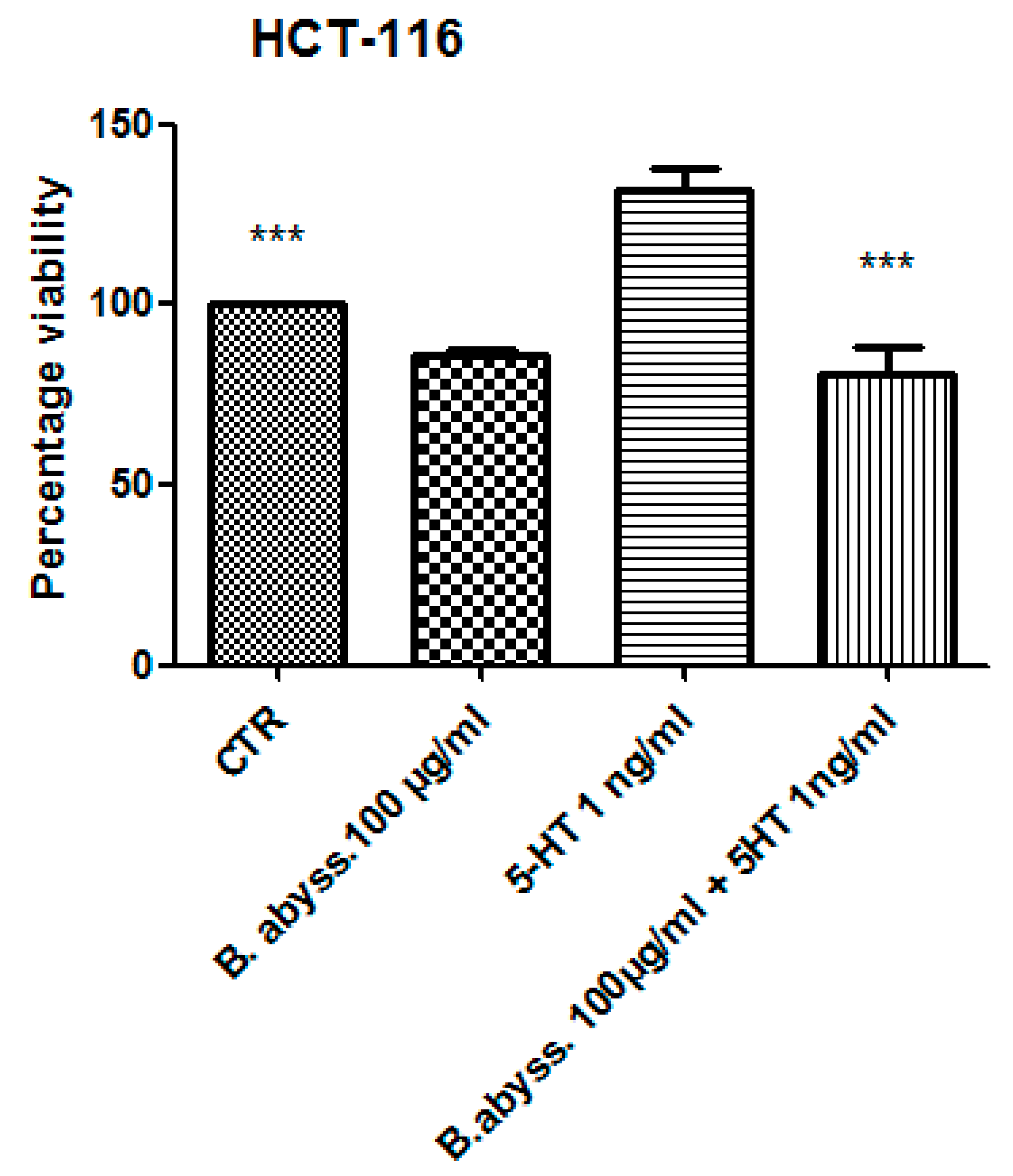
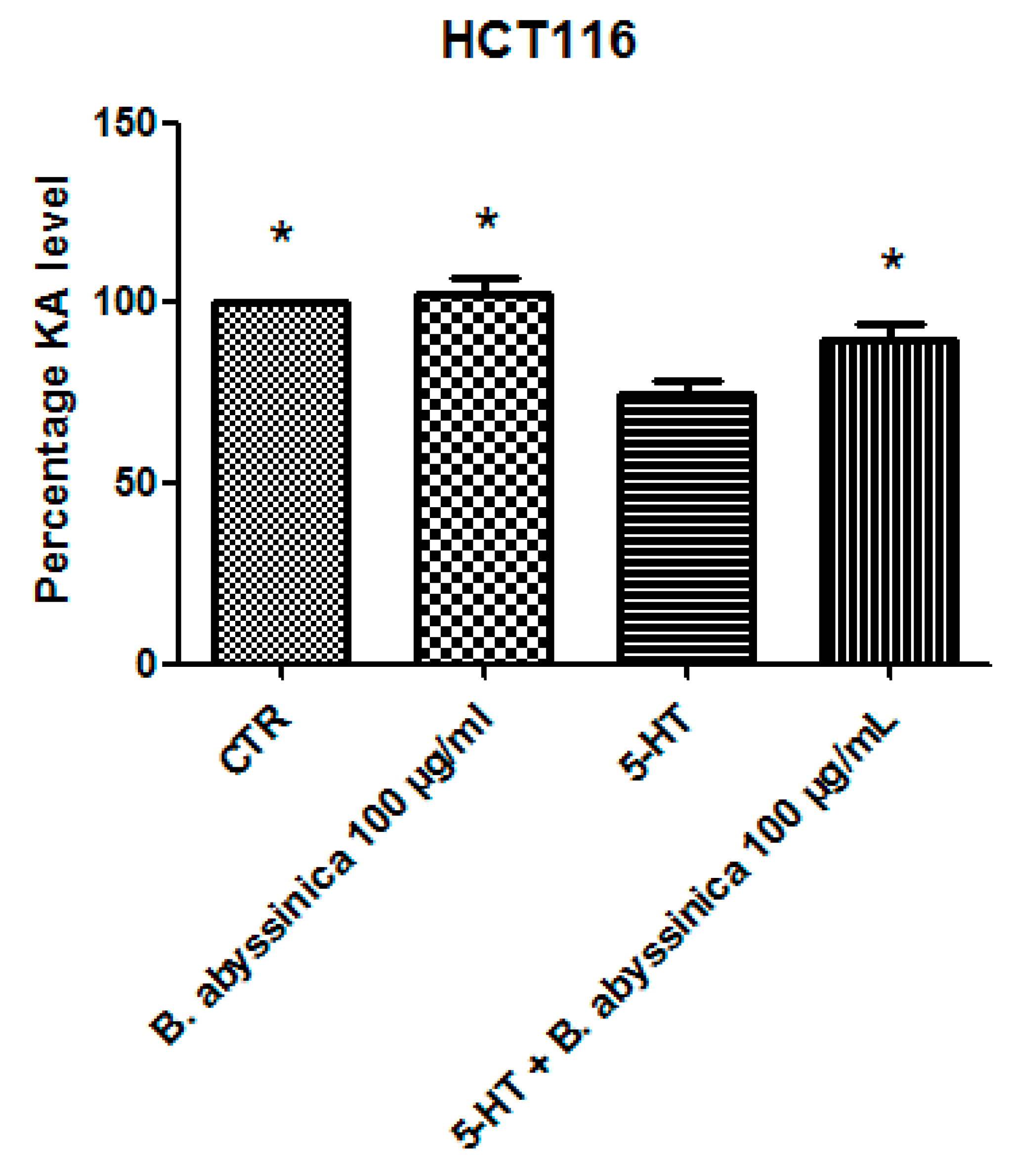
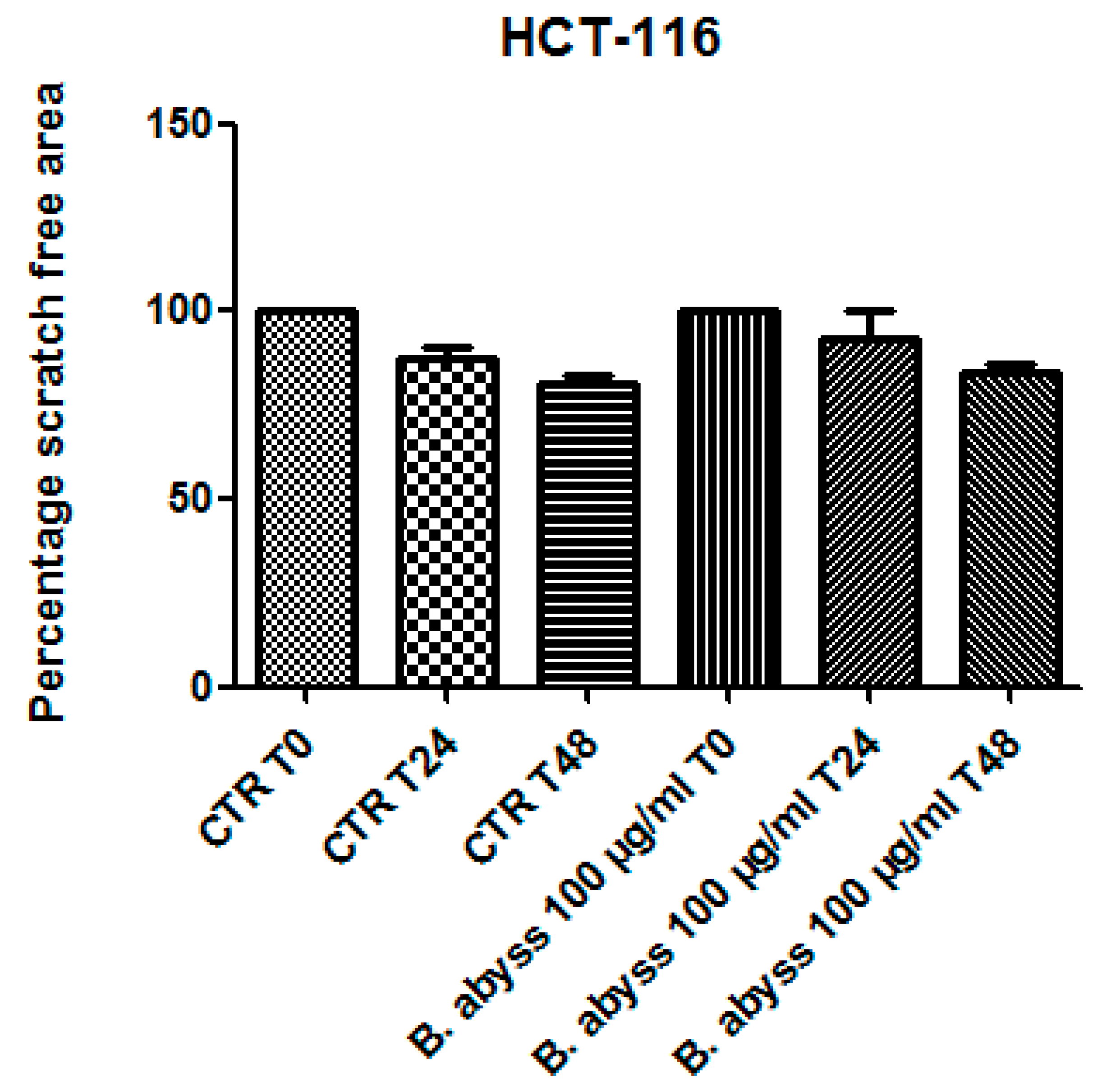
3.2. Pharmacological Study
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Amit, L.; Vikas, G.; Vaibhav, T.; Vikash, K.; Siddhartha, G.; Lather, A. Phytochemistry and pharmacological activities of Bersama engleriana Guerke-An overview. Sex. Transm. Dis. 2010, 11, 12. [Google Scholar]
- Kidane, B.; van Andel, T.; van der Maesen, L.J.G.; Asfaw, Z. Use and management of traditional medicinal plants by Maale and Ari ethnic communities in southern Ethiopia. J. Ethnobiol. Ethnomed. 2014, 10, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zekeya, N.; Chacha, M.; Shahada, F.; Kidukuli, A. Analysis of phytochemical composition of Bersama abyssinica by gas chromatography-mass spectrometry. J. Pharmacogn. Phytochem. 2014, 3, 246–252. [Google Scholar]
- Tuasha, N.; Petros, B.; Asfaw, Z. Medicinal plants used by traditional healers to treat malignancies and other human ailments in Dalle District, Sidama Zone, Ethiopia. J. Ethnobiol. Ethnomedicine 2018, 14, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kupchan, S.M.; Ognyanov, I.; Moniot, J.L. Tumor inhibitors. LXIV. Isolation and structural elucidation of novel bufadienolides, the cytotoxic principles of Bersama abyssinica. Bioorg. Chem. 1971, 1, 13–31. [Google Scholar] [CrossRef]
- Kupchan, S.M.; Hemingway, R.J.; Hemingway, J.C. The isolation and characterization of hellebrigenin 3-acetate and hellebrigenin 3,5-diacetate, bufadienolide tumor inhibitors from Bersama abyssinica. Tetrahedron Lett. 1968, 9, 149–152. [Google Scholar] [CrossRef]
- Ameya, G.; Manilal, A.; Idhayadhulla, A. Phytochemical Analysis and Antimicrobial Activity of Bersama abyssinica Fresen against Multidrug-Resistant Bacterial Uropathogens: Picolinyl Hydrazide Is a Major Compound. J. HerbsSpices Med. Plants 2019, 25, 389–400. [Google Scholar] [CrossRef]
- Ataee, R.; Ajdary, S.; Rezayat, M.; Shokrgozar, M.A.; Shahriari, S.; Zarrindast, M.R. Study of 5HT3 and HT4 receptor expression in HT29 cell line and human colon adenocarcinoma tissues. Arch. Iran. Med. 2010, 13, 120–125. [Google Scholar]
- Ballou, Y.; Rivas, A.; Belmont, A.; Patel, L.; Amaya, C.N.; Lipson, S.; Khayou, T.; Dickerson, E.B.; Nahleh, Z.; Bryan, B.A. 5-HT serotonin receptors modulate mitogenic signaling and impact tumor cell viability. Mol. Clin. Oncol. 2018, 9, 243–254. [Google Scholar] [CrossRef] [Green Version]
- Regmi, S.C.; Park, S.-Y.; Ku, S.K.; Kim, J.-A. Serotonin regulates innate immune responses of colon epithelial cells through Nox2-derived reactive oxygen species. Free Radic. Biol. Med. 2014, 69, 377–389. [Google Scholar] [CrossRef]
- Uysal, S.; Aktumsek, A. A phytochemical study on Potentilla anatolica: An endemic Turkish plant. Ind. Crop. Prod. 2015, 76, 1001–1007. [Google Scholar] [CrossRef]
- Vladimir-Knezevic, S.; Blazekovic, B.; Stefan, M.B.; Alegro, A.; Koszegi, T.; Petrik, J. Antioxidant activities and polyphenolic contents of three selected Micromeria species from Croatia. Molecules 2011, 16, 1454–1470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zengin, G.; Aktumsek, A. Investigation of antioxidant potentials of solvent extracts from different anatomical parts of Asphodeline anatolica E. Tuzlaci: An endemic plant to Turkey. Afr. J. Tradit. Complement. Altern. Med. 2014, 11, 481–488. [Google Scholar] [CrossRef] [Green Version]
- Djemgou, P.C.; Hussien, T.A.; Hegazy, M.E.F.; Ngandeu, F.; Neguim, G.; Tane, P.; Mohamed, A.E.H.H. C-Glucoside xanthone from the stem bark extract of Bersama engleriana. Pharmacogn. Res. 2010, 2, 229. [Google Scholar]
- Kupchan, S.M.; Moniot, J.; Sigel, C.; Hemingway, R.J. Tumor inhibitors. LXV. Bersenogenin, berscillogenin, and 3-epiberscillogenin, three new cytotoxic bufadienolides from Bersama abyssinica. J. Org. Chem. 1971, 36, 2611–2616. [Google Scholar] [CrossRef]
- Di Giacomo, V.; Chiavaroli, C.; Orlando, G.; Cataldi, A.; Rapino, M.; Di Valerio, V.; Leone, S.; Brunetti, L.; Menghini, L.; Recinella, L.; et al. Neuroprotective and Neuromodulatory Effects Induced by Cannabidiol and Cannabigerol in Rat Hypo-E22 cells and Isolated Hypothalamus. Antioxidants 2020, 9, 71. [Google Scholar] [CrossRef] [Green Version]
- Aktumsek, A.; Zengin, G.; Guler, G.O.; Cakmak, Y.S.; Duran, A. Antioxidant potentials and anticholinesterase activities of methanolic and aqueous extracts of three endemic Centaurea L. species. Food Chem. Toxicol. 2013, 55, 290–296. [Google Scholar] [CrossRef]
- Grochowski, D.M.; Uysal, S.; Aktumsek, A.; Granica, S.; Zengin, G.; Ceylan, R.; Locatelli, M.; Tomczyk, M. In vitro enzyme inhibitory properties, antioxidant activities, and phytochemical profile of Potentilla thuringiaca. Phytochem. Lett. 2017, 20, 365–372. [Google Scholar] [CrossRef]
- Ferrante, C.; Recinella, L.; Ronci, M.; Menghini, L.; Brunetti, L.; Chiavaroli, A.; Leone, S.; Di Iorio, L.; Carradori, S.; Tirillini, B. Multiple pharmacognostic characterization on hemp commercial cultivars: Focus on inflorescence water extract activity. Food Chem. Toxicol. 2019, 125, 452–461. [Google Scholar] [CrossRef]
- Delgoda, R.; Murray, J.E. Chapter 7—Evolutionary Perspectives on the Role of Plant Secondary Metabolites. In Pharmacognosy; Badal, S., Delgoda, R., Eds.; Academic Press: Boston, MA, USA, 2017; pp. 93–100. [Google Scholar]
- Sinan, K.I.; Bene, K.; Zengin, G.; Diuzheva, A.; Jekő, J.; Cziáky, Z.; Picot-Allain, C.M.N.; Mollica, A.; Rengasamy, K.R.; Mahomoodally, M.F. A comparative study of the HPLC-MS profiles and biological efficiency of different solvent leaf extracts of two African plants: Bersama abyssinica and Scoparia dulcis. Int. J. Environ. Health Res. 2019, 1–13. [Google Scholar] [CrossRef]
- Ramos-Chávez, L.A.; Lugo Huitrón, R.; González Esquivel, D.; Pineda, B.; Ríos, C.; Silva-Adaya, D.; Sánchez-Chapul, L.; Roldán-Roldán, G.; Pérez de la Cruz, V. Relevance of Alternative Routes of Kynurenic Acid Production in the Brain. Oxidative Med. Cell. Longev. 2018, 2018, 14. [Google Scholar] [CrossRef] [Green Version]
- Blanco, A.; Blanco, G. Chapter 8—Enzymes. In Medical Biochemistry; Blanco, A., Blanco, G., Eds.; Academic Press: London, UK, 2017; Volume 8, pp. 153–175. [Google Scholar]
- Curtis, J.J.; Seymour, C.B.; Mothersill, C.E. Cell Line-Specific Direct Irradiation and Bystander Responses are Influenced by Fetal Bovine Serum Serotonin Concentrations. Radiat. Res. 2018, 190, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Tsai, F.-M.; Wu, C.-C.; Shyu, R.-Y.; Wang, C.-H.; Jiang, S.-Y. Tazarotene-induced gene 1 inhibits prostaglandin E2-stimulated HCT116 colon cancer cell growth. J. Biomed. Sci. 2011, 18, 88. [Google Scholar] [CrossRef] [Green Version]
- Notarangelo, F.M.; Beggiato, S.; Schwarcz, R. Assessment of Prenatal Kynurenine Metabolism Using Tissue Slices: Focus on the Neosynthesis of Kynurenic Acid in Mice. Dev. Neurosci. 2019, 41, 1–10. [Google Scholar] [CrossRef]
- Fukui, S.; Schwarcz, R.; Rapoport, S.I.; Takada, Y.; Smith, Q.R. Blood–brain barrier transport of kynurenines: Implications for brain synthesis and metabolism. J. Neurochem. 1991, 56, 2007–2017. [Google Scholar] [CrossRef]
- Ferrante, C.; Recinella, L.; Locatelli, M.; Guglielmi, P.; Secci, D.; Leporini, L.; Chiavaroli, A.; Leone, S.; Martinotti, S.; Brunetti, L.; et al. Protective Effects Induced by Microwave-Assisted Aqueous Harpagophytum Extract on Rat Cortex Synaptosomes Challenged with Amyloid β-Peptide. Phytother. Res. 2017, 31, 1257–1264. [Google Scholar] [CrossRef] [PubMed]
- Oláh, G.; Herédi, J.; Menyhárt, Á.; Czinege, Z.; Nagy, D.; Fuzik, J.; Kocsis, K.; Knapp, L.; Krucsó, E.; Gellért, L. Unexpected effects of peripherally administered kynurenic acid on cortical spreading depression and related blood–brain barrier permeability. Drug Des. Dev. Ther. 2013, 7, 981. [Google Scholar]
- Marciniak, S.; Wnorowski, A.; Smolińska, K.; Walczyna, B.; Turski, W.; Kocki, T.; Paluszkiewicz, P.; Parada-Turska, J. Kynurenic Acid Protects against Thioacetamide-Induced Liver Injury in Rats. Anal. Cell. Pathol. 2018, 2018, 1270483. [Google Scholar] [CrossRef] [Green Version]
- Walczak, K.; Turski, W.A.; Rajtar, G. Kynurenic acid inhibits colon cancer proliferation in vitro: Effects on signaling pathways. Amino Acids 2014, 46, 2393–2401. [Google Scholar] [CrossRef] [Green Version]
- Walczak, K.; Turski, W.A.; Rzeski, W. Kynurenic acid enhances expression of p21 Waf1/Cip1 in colon cancer HT-29 cells. Pharmacol. Rep. 2012, 64, 745–750. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extracts | Total Phenolic Content (mg GAE/g Extract) | Total Flavonoid Content (mg RE/g Extract) | Total Phenolic Acid Content (mg CAE/g) | Total Flavanol Content (mg CE/g) | Total Tannin Content (mg CE/g) | Total Saponin Content (mg QE/g) |
|---|---|---|---|---|---|---|
| EA | 100.57 ± 0.67 c | 56.25 ± 0.52 b | nd | 2.79 ± 0.01 a | 2.67 ± 0.16 a | 97.37 ± 10.22 a |
| MeOH | 216.79 ± 1.11 b | 75.82 ± 0.50 a | 2.90 ± 0.46 a | 1.23 ± 0.02 b | 1.25 ± 0.02 b | 76.44 ± 10.70 a |
| Water | 230.83 ± 1.64 a | 40.32 ± 0.26 c | 3.46 ± 0.04 a | 1.23 ± 0.01 b | 1.12 ± 0.19 b | 57.59 ± 7.44 b |
| Extracts | Gallic Acid (mg/g Extract) | Rutin (mg/g Extract) |
|---|---|---|
| EA | 1.63 ± 0.15 | 0.60 ± 0.07 |
| MeOH | 3.64 ± 0.33 | 0.14 ± 0.02 |
| Water | 3.26 ± 0.13 | 0.12 ± 0.01 |
| Extracts | DPPH (mg TE/g Extract) | ABTS (mg TE/g Extract) | CUPRAC (mgTE/g Extract) | FRAP (mgTE/g Extract) | Phosphomolybdenum (mmol TE/g) | Metal Chelating Ability (mg EDTAE/g) |
|---|---|---|---|---|---|---|
| EA | 392.04 ± 1.83 b | 232.90 ± 5.46 b | 508.51 ± 2.04 b | 363.82 ± 2.00 c | 2.48 ± 0.14 c | 30.68 ± 1.43 b |
| MeOH | 1092.46 ± 7.38 a | 698.86 ± 32.24 a | 1464.08 ± 7.46 a | 983.86 ± 7.86 b | 2.90 ± 0.09 b | 32.07 ± 0.55 b |
| Water | 1073.20 ± 22.99 a | 560.32 ± 28.57 a | 1449.46 ± 6.15 a | 1063.70 ± 16.20 a | 3.18 ± 0.07 a | 49.74 ± 0.11 a |
| No. | Name | Formula | Rt | [M+H]+ | [M−H]− | Fragment 1 | Fragment 2 | Fragment 3 | Fragment 4 | Fragment 5 |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Galloylhexose isomer 1 | C13H16O10 | 1.26 | 331.07 | 271.05 | 241.03 | 211.02 | 169.01 | 125.02 | |
| 2 | Citric acid | C6H8O7 | 1.56 | 191.02 | 173.01 | 129.02 | 111.01 | 87.01 | 85.03 | |
| 3 | Galloylhexose isomer 2 | C13H16O10 | 1.70 | 331.07 | 271.05 | 241.04 | 211.02 | 169.01 | 125.02 | |
| 4 | Galloylhexose isomer 3 | C13H16O10 | 2.19 | 331.07 | 271.05 | 241.03 | 211.02 | 169.01 | 125.02 | |
| 5 1 | Gallic acid (3,4,5-trihydroxybenzoic acid) | C7H6O5 | 2.30 | 169.01 | 125.02 | 97.03 | 81.03 | 79.02 | 69.03 | |
| 6 | Galloylhexose isomer 4 | C13H16O10 | 2.82 | 331.07 | 271.05 | 241.04 | 211.02 | 169.01 | 125.02 | |
| 7 | Protocatechuic acid (3,4-dihydroxybenzoic acid) | C7H6O4 | 4.65 | 153.02 | 109.03 | 108.02 | 91.02 | 81.03 | ||
| 8 | Syringic acid-O-hexoside isomer 1 | C15H20O10 | 10.17 | 359.10 | 197.05 | 182.02 | 167.00 | 153.05 | 138.03 | |
| 9 | Syringic acid-O-hexoside isomer 2 | C15H20O10 | 10.60 | 359.10 | 197.05 | 182.02 | 167.00 | 153.05 | 138.03 | |
| 10 | Caffeic acid | C9H8O4 | 14.28 | 179.03 | 135.04 | 107.05 | ||||
| 11 | Unidentified tannin | C34H26O23 | 14.38 | 801.08 | 649.07 | 561.09 | 499.07 | 347.06 | 301.00 | |
| 12 | Trigalloylhexose isomer 1 | C27H24O18 | 14.46 | 635.09 | 483.08 | 465.07 | 313.06 | 169.01 | 125.02 | |
| 13 | Digalloylhexose | C20H20O14 | 15.18 | 483.08 | 331.07 | 313.06 | 271.05 | 169.01 | 125.02 | |
| 14 | Trigalloylhexose isomer 2 | C27H24O18 | 15.37 | 635.09 | 483.08 | 465.07 | 313.06 | 169.01 | 125.02 | |
| 15 | Trigalloylhexose isomer 3 | C27H24O18 | 16.56 | 635.09 | 483.07 | 465.07 | 313.06 | 169.01 | 125.02 | |
| 16 | Ethyl syringate | C11H14O5 | 16.68 | 227.09 | 181.05 | 155.07 | 140.05 | 123.04 | 95.05 | |
| 17 | Feruloylhexose isomer 1 | C16H20O9 | 16.73 | 355.10 | 295.08 | 235.06 | 193.05 | 175.04 | 134.04 | |
| 18 | Trigalloylhexose isomer 4 | C27H24O18 | 17.31 | 635.09 | 483.08 | 465.07 | 313.06 | 169.01 | 125.02 | |
| 19 | Synapoylhexose isomer 1 | C17H22O10 | 17.56 | 385.11 | 325.09 | 265.07 | 223.06 | 205.05 | 190.03 | |
| 201 | 4-Coumaric acid | C9H8O3 | 17.70 | 163.04 | 119.05 | 93.03 | ||||
| 21 | Feruloylhexose isomer 2 | C16H20O9 | 17.84 | 355.10 | 295.08 | 235.06 | 193.05 | 175.04 | 134.04 | |
| 22 | Myrciaphenone B | C21H22O13 | 17.87 | 481.10 | 313.06 | 169.01 | 125.02 | |||
| 23 | Mangiferin (Aphloiol, Chinonin) | C19H18O11 | 18.38 | 421.08 | 343.05 | 331.05 | 301.04 | 272.03 | 259.03 | |
| 24 | Synapoylhexose isomer 2 | C17H22O10 | 18.53 | 385.11 | 325.09 | 265.07 | 223.06 | 205.05 | 190.03 | |
| 25 | Ferulic acid | C10H10O4 | 19.24 | 193.05 | 178.03 | 149.06 | 137.02 | 134.04 | ||
| 26 | Tetragalloylhexose | C34H28O22 | 19.36 | 787.10 | 635.09 | 617.08 | 465.07 | 313.06 | 169.01 | |
| 27 | Loliolide | C11H16O3 | 19.45 | 197.12 | 179.11 | 161.10 | 135.12 | 133.10 | 107.09 | |
| 28 | Ellagic acid-4-O-glucoside | C20H16O13 | 19.89 | 463.05 | 301.00 | 299.99 | 298.98 | |||
| 29 | Feruloyl-galloylhexose | C23H24O13 | 20.98 | 507.11 | 313.06 | 193.05 | 179.03 | 169.01 | 125.02 | |
| 30 | Quercetin-O-galloylhexoside isomer 1 | C28H24O16 | 21.31 | 615.10 | 463.09 | 301.03 | 300.03 | 255.03 | 169.01 | |
| 311 | Vitexin (Apigenin-8-C-glucoside) | C21H20O10 | 21.34 | 433.11 | 415.10 | 397.09 | 367.08 | 313.07 | 283.06 | |
| 32 | Quercetin-O-galloylhexoside isomer 2 | C28H24O16 | 21.56 | 615.10 | 463.09 | 301.04 | 300.03 | 271.02 | 169.01 | |
| 33 | Isovitexin (Apigenin-6-C-glucoside) | C21H20O10 | 22.24 | 433.11 | 415.10 | 397.09 | 367.08 | 313.07 | 283.06 | |
| 34 | Sinapoyl-galloylhexose | C24H26O14 | 22.36 | 537.12 | 325.09 | 265.07 | 223.06 | 169.01 | 125.02 | |
| 35 | Hyperoside (Quercetin-3-O-galactoside) | C21H20O12 | 22.64 | 463.09 | 301.04 | 300.03 | 271.02 | 255.03 | 151.00 | |
| 36 | Ellagic acid-O-pentoside | C19H14O12 | 22.75 | 433.04 | 301.00 | 299.99 | 298.99 | |||
| 37 | Acetylmangiferin | C21H20O12 | 23.11 | 463.09 | 331.05 | 301.04 | 271.03 | 259.03 | ||
| 38 | Ellagic acid | C14H6O8 | 23.37 | 301.00 | 284.00 | 257.01 | 229.01 | 201.02 | 185.02 | |
| 39 | Avicularin (Quercetin-3-O-arabinofuranoside) | C20H18O11 | 23.45 | 433.08 | 301.04 | 300.03 | 271.03 | 255.03 | 179.00 | |
| 40 | Unidentified terpenoid | C26H34O8 | 24.67 | 475.23 | 379.19 | 361.18 | 351.20 | 333.19 | 315.18 | |
| 41 | Isorhamnetin-O-hexoside | C22H22O12 | 24.70 | 477.10 | 315.05 | 314.04 | 285.04 | 271.03 | 243.03 | |
| 42 | Kaempferol-O-pentoside | C20H18O10 | 24.83 | 417.08 | 285.04 | 284.03 | 255.03 | 227.03 | ||
| 431 | Isorhamnetin-3-O-glucoside | C22H22O12 | 24.88 | 477.10 | 315.05 | 314.04 | 285.04 | 271.03 | 243.03 | |
| 44 | 4-Methoxycinnamic acid | C10H10O3 | 25.31 | 179.07 | 161.06 | 133.07 | 105.07 | 79.05 | ||
| 45 | 3-O-Methylellagic acid | C15H8O8 | 25.66 | 315.01 | 299.99 | 244.00 | ||||
| 46 | Tetrahydroxyxanthone | C13H8O6 | 26.12 | 259.02 | 231.03 | 215.03 | 203.03 | 187.04 | ||
| 47 | Dihydroactinidiolide | C11H16O2 | 26.57 | 181.12 | 163.11 | 145.10 | 135.12 | 107.09 | 93.07 | |
| 481 | Naringenin (4’,5,7-trihydroxyflavanone) | C15H12O5 | 27.15 | 271.06 | 177.02 | 151.00 | 119.05 | 107.01 | 93.03 | |
| 49 | 3,3’-Di-O-methylellagic acid | C16H10O8 | 27.81 | 329.03 | 314.01 | 298.98 | 270.99 | |||
| 50 | 3,3’,4-Tri-O-methylellagic acid | C17H12O8 | 30.17 | 343.05 | 328.02 | 313.00 | 297.98 | 285.00 | ||
| 51 | 3,3’,4-Tri-O-methylflavellagic acid | C17H12O9 | 31.21 | 359.04 | 344.02 | 328.99 | 313.97 | 301.00 | ||
| 52 | 3,3’,4,4’-Tetra-O-methylellagic acid | C18H14O8 | 31.98 | 359.08 | 344.05 | 343.05 | 329.03 | 313.03 | ||
| 53 | Bruguierol A | C12H14O2 | 36.05 | 191.11 | 173.10 | 161.10 | 147.08 | 135.08 | 107.05 | |
| 54 | Ginsenoside Ro or isomer | C48H76O19 | 36.23 | 955.49 | 793.44 | 731.44 | 613.37 | 569.38 | 551.38 | |
| 55 | Cynarasaponin C or isomer | C42H66O14 | 37.20 | 793.44 | 631.39 | 587.40 | 569.38 | 497.37 | 455.35 | |
| 56 | Ginsenoside Ro or isomer | C48H76O19 | 39.75 | 955.49 | 793.44 | 731.44 | 569.38 | 551.37 | 455.35 | |
| 57 | Unidentified saponin | C48H76O19 | 40.25 | 955.49 | 893.48 | 793.44 | 731.44 | 551.37 | 455.35 | |
| 58 | Hexadecanedioic acid | C16H30O4 | 40.30 | 285.21 | 267.20 | 223.21 | 57.03 |
| No. | Name | Formula | Rt | [M+H]+ | [M−H]− | Fragment 1 | Fragment 2 | Fragment 3 | Fragment 4 | Fragment 5 |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Galloylhexose isomer 1 | C13H16O10 | 1.22 | 331.07 | 271.05 | 241.03 | 211.02 | 169.01 | 125.02 | |
| 2 | Citric acid | C6H8O7 | 1.56 | 191.02 | 173.01 | 129.02 | 111.01 | 87.01 | 85.03 | |
| 3 | Galloylhexose isomer 2 | C13H16O10 | 1.71 | 331.07 | 271.05 | 241.04 | 211.02 | 169.01 | 125.02 | |
| 4 | Galloylhexose isomer 3 | C13H16O10 | 2.18 | 331.07 | 271.05 | 241.03 | 211.02 | 169.01 | 125.02 | |
| 5 1 | Gallic acid (3,4,5-trihydroxybenzoic acid) | C7H6O5 | 2.31 | 169.01 | 125.02 | 97.03 | 81.03 | 79.02 | 69.03 | |
| 6 | Galloylhexose isomer 4 | C13H16O10 | 2.81 | 331.07 | 271.05 | 241.03 | 211.02 | 169.01 | 125.02 | |
| 7 | Protocatechuic acid (3,4-dihydroxybenzoic acid) | C7H6O4 | 4.65 | 153.02 | 109.03 | 108.02 | 91.02 | 81.03 | ||
| 8 | Syringic acid-O-hexoside isomer 1 | C15H20O10 | 10.19 | 359.10 | 197.05 | 182.02 | 167.00 | 153.05 | 138.03 | |
| 9 | Syringic acid-O-hexoside isomer 2 | C15H20O10 | 10.61 | 359.10 | 197.05 | 182.02 | 167.00 | 153.05 | 138.03 | |
| 10 | Kynurenic acid | C10H7NO3 | 13.07 | 190.05 | 162.06 | 144.04 | 116.05 | |||
| 11 | Caffeic acid | C9H8O4 | 14.28 | 179.03 | 135.04 | 107.05 | ||||
| 12 | Unidentified tannin | C34H26O23 | 14.37 | 801.08 | 649.07 | 561.09 | 499.07 | 347.06 | 301.00 | |
| 13 | Trigalloylhexose isomer 1 | C27H24O18 | 14.46 | 635.09 | 483.08 | 465.07 | 313.06 | 169.01 | 125.02 | |
| 14 | Digalloylhexose | C20H20O14 | 15.19 | 483.08 | 331.07 | 313.06 | 271.05 | 169.01 | 125.02 | |
| 15 | Trigalloylhexose isomer 2 | C27H24O18 | 15.37 | 635.09 | 483.08 | 465.07 | 313.06 | 169.01 | 125.02 | |
| 16 | Castalin or vescalin | C27H20O18 | 15.65 | 631.06 | 451.00 | 301.00 | 299.99 | 298.98 | 270.99 | |
| 17 | Methylcoumarin | C10H8O2 | 15.93 | 161.06 | 133.07 | 105.07 | 91.05 | |||
| 18 | Castalin or vescalin | C27H20O18 | 16.38 | 631.06 | 451.00 | 301.00 | 299.99 | 298.98 | 270.99 | |
| 19 | Trigalloylhexose isomer 3 | C27H24O18 | 16.57 | 635.09 | 483.08 | 465.07 | 313.06 | 169.01 | 125.02 | |
| 20 | Ethyl syringate | C11H14O5 | 16.67 | 227.09 | 181.05 | 155.07 | 140.05 | 123.04 | 95.05 | |
| 21 | Feruloylhexose isomer 1 | C16H20O9 | 16.73 | 355.10 | 295.08 | 235.06 | 193.05 | 175.04 | 134.04 | |
| 22 | Trigalloylhexose isomer 4 | C27H24O18 | 17.31 | 635.09 | 483.08 | 465.07 | 313.06 | 169.01 | 125.02 | |
| 23 | Synapoylhexose isomer 1 | C17H22O10 | 17.56 | 385.11 | 325.09 | 265.07 | 223.06 | 205.05 | 190.03 | |
| 241 | 4-Coumaric acid | C9H8O3 | 17.70 | 163.04 | 119.05 | 93.03 | ||||
| 25 | Feruloylhexose isomer 2 | C16H20O9 | 17.84 | 355.10 | 295.08 | 235.06 | 193.05 | 175.04 | 134.04 | |
| 26 | Myrciaphenone B | C21H22O13 | 17.86 | 481.10 | 313.06 | 169.01 | 125.02 | |||
| 27 | Mangiferin (Aphloiol, Chinonin) | C19H18O11 | 18.33 | 421.08 | 343.05 | 331.05 | 301.04 | 272.03 | 259.02 | |
| 28 | Synapoylhexose isomer 2 | C17H22O10 | 18.52 | 385.11 | 325.09 | 265.07 | 223.06 | 205.05 | 190.03 | |
| 29 | Ferulic acid | C10H10O4 | 19.24 | 193.05 | 178.03 | 149.06 | 137.02 | 134.04 | ||
| 30 | Loliolide | C11H16O3 | 19.46 | 197.12 | 179.11 | 161.10 | 135.12 | 133.10 | 107.09 | |
| 31 | Tetragalloylhexose | C34H28O22 | 19.47 | 787.10 | 635.09 | 617.08 | 465.07 | 313.06 | 169.01 | |
| 32 | Ellagic acid-4-O-glucoside | C20H16O13 | 19.90 | 463.05 | 301.00 | 299.99 | 298.98 | |||
| 33 | Feruloyl-galloylhexose | C23H24O13 | 20.99 | 507.11 | 313.06 | 193.05 | 179.03 | 169.01 | 125.02 | |
| 34 | Berscillogenin or 3-epiberscillogenin | C24H30O6 | 21.30 | 415.21 | 397.20 | 379.19 | 367.19 | 361.18 | 351.20 | |
| 35 | Quercetin-O-galloylhexoside isomer 1 | C28H24O16 | 21.33 | 615.10 | 463.09 | 301.04 | 300.03 | 255.03 | 169.01 | |
| 361 | Vitexin (Apigenin-8-C-glucoside) | C21H20O10 | 21.37 | 433.11 | 415.10 | 397.09 | 367.08 | 313.07 | 283.06 | |
| 37 | Quercetin-O-galloylhexoside isomer 2 | C28H24O16 | 21.55 | 615.10 | 463.09 | 301.04 | 300.03 | 271.02 | 169.01 | |
| 38 | Berscillogenin or 3-epiberscillogenin | C24H30O6 | 22.02 | 415.21 | 397.21 | 379.19 | 367.19 | 361.18 | 351.19 | |
| 39 | Isovitexin (Apigenin-6-C-glucoside) | C21H20O10 | 22.24 | 433.11 | 415.10 | 397.09 | 367.08 | 313.07 | 283.06 | |
| 40 | Sinapoyl-galloylhexose | C24H26O14 | 22.37 | 537.12 | 325.09 | 265.07 | 223.06 | 169.01 | 125.02 | |
| 41 | Hyperoside (Quercetin-3-O-galactoside) | C21H20O12 | 22.65 | 463.09 | 301.04 | 300.03 | 271.03 | 255.03 | 151.00 | |
| 42 | Ellagic acid-O-pentoside | C19H14O12 | 22.75 | 433.04 | 301.00 | 299.99 | 298.99 | |||
| 431 | Isoquercitrin (Quercetin-3-O-glucoside) | C21H20O12 | 22.89 | 463.09 | 301.04 | 300.03 | 271.03 | 255.03 | 151.00 | |
| 44 | Acetylmangiferin | C21H20O12 | 23.11 | 463.09 | 331.05 | 301.04 | 271.02 | 259.02 | ||
| 45 | Eschweilenol A | C20H10O11 | 23.27 | 425.01 | 301.00 | 299.99 | 298.98 | |||
| 46 | Ellagic acid | C14H6O8 | 23.35 | 301.00 | 284.00 | 257.01 | 229.01 | 201.02 | 185.02 | |
| 47 | Avicularin (Quercetin-3-O-arabinofuranoside) | C20H18O11 | 23.47 | 433.08 | 301.04 | 300.03 | 271.03 | 255.03 | 179.00 | |
| 48 | Unidentified terpenoid | C26H34O8 | 24.66 | 475.23 | 379.19 | 361.18 | 351.20 | 333.19 | 315.17 | |
| 49 | Isorhamnetin-O-hexoside | C22H22O12 | 24.70 | 477.10 | 315.05 | 314.04 | 285.04 | 271.02 | 243.03 | |
| 50 | Kaempferol-O-pentoside | C20H18O10 | 24.84 | 417.08 | 285.04 | 284.03 | 255.03 | 227.03 | ||
| 511 | Isorhamnetin-3-O-glucoside | C22H22O12 | 24.90 | 477.10 | 315.05 | 314.04 | 285.04 | 271.03 | 243.03 | |
| 52 | 4-Methoxycinnamic acid | C10H10O3 | 25.32 | 179.07 | 161.06 | 133.07 | 105.07 | 79.05 | ||
| 53 | 3-O-Methylellagic acid | C15H8O8 | 25.66 | 315.01 | 299.99 | 244.00 | ||||
| 54 | Tetrahydroxyxanthone | C13H8O6 | 26.13 | 259.02 | 231.03 | 215.03 | 203.03 | 187.04 | ||
| 55 | Dihydroactinidiolide | C11H16O2 | 26.58 | 181.12 | 163.11 | 145.10 | 135.12 | 107.09 | 93.07 | |
| 561 | Naringenin (4’,5,7-trihydroxyflavanone) | C15H12O5 | 27.17 | 271.06 | 177.02 | 151.00 | 119.05 | 107.01 | 93.03 | |
| 57 | 3,3’-Di-O-methylellagic acid | C16H10O8 | 27.82 | 329.03 | 314.01 | 298.98 | 270.99 | |||
| 58 | Scilliglaucosidin | C24H30O5 | 27.87 | 399.22 | 381.21 | 363.19 | 345.19 | 335.20 | ||
| 59 | 3,3’,4-Tri-O-methylellagic acid | C17H12O8 | 30.18 | 343.05 | 328.02 | 313.00 | 297.98 | 285.00 | ||
| 60 | 3,3’,4-Tri-O-methylflavellagic acid | C17H12O9 | 31.21 | 359.04 | 344.02 | 328.99 | 313.97 | 301.00 | ||
| 61 | 3,3’,4,4’-Tetra-O-methylellagic acid | C18H14O8 | 31.98 | 359.08 | 344.05 | 343.05 | 329.03 | 313.03 | ||
| 62 | Bruguierol A | C12H14O2 | 36.05 | 191.11 | 173.10 | 161.10 | 147.08 | 135.08 | 107.05 | |
| 63 | Ginsenoside Ro or isomer | C48H76O19 | 36.24 | 955.49 | 793.44 | 731.44 | 613.37 | 569.38 | 551.37 | |
| 64 | Cynarasaponin C or isomer | C42H66O14 | 37.18 | 793.44 | 631.39 | 587.40 | 569.39 | 497.37 | 455.35 | |
| 65 | Ginsenoside Ro or isomer | C48H76O19 | 39.71 | 955.49 | 793.44 | 731.44 | 569.38 | 551.37 | 455.35 | |
| 66 | Unidentified saponin 1 | C48H76O19 | 40.28 | 955.49 | 893.49 | 793.44 | 731.44 | 551.37 | 455.35 | |
| 67 | Hexadecanedioic acid | C16H30O4 | 40.29 | 285.21 | 267.20 | 223.21 | 57.03 | |||
| 68 | Unidentified saponin 2 | C49H78O19 | 41.70 | 969.51 | 951.50 | 585.38 | 537.36 | 455.35 | 453.33 |
| No. | Name | Formula | Rt | [M+H]+ | [M−H]− | Fragment 1 | Fragment 2 | Fragment 3 | Fragment 4 | Fragment 5 |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Galloylhexose isomer 1 | C13H16O10 | 1.31 | 331.07 | 271.05 | 241.04 | 211.02 | 169.01 | 125.02 | |
| 2 | Citric acid | C6H8O7 | 1.55 | 191.02 | 173.01 | 129.02 | 111.01 | 87.01 | 85.03 | |
| 3 | Galloylhexose isomer 2 | C13H16O10 | 1.76 | 331.07 | 271.05 | 241.03 | 211.02 | 169.01 | 125.02 | |
| 4 | Galloylhexose isomer 3 | C13H16O10 | 2.12 | 331.07 | 271.05 | 241.03 | 211.02 | 169.01 | 125.02 | |
| 5 1 | Gallic acid (3,4,5-trihydroxybenzoic acid) | C7H6O5 | 2.28 | 169.01 | 125.02 | 97.03 | 81.03 | 79.02 | 69.03 | |
| 6 | Galloylhexose isomer 4 | C13H16O10 | 2.79 | 331.07 | 271.05 | 241.04 | 211.02 | 169.01 | 125.02 | |
| 7 | Protocatechuic acid (3,4-dihydroxybenzoic acid) | C7H6O4 | 4.69 | 153.02 | 109.03 | 108.02 | 91.02 | 81.03 | ||
| 8 | Piscidic acid (4-hydroxybenzyltartaric acid) | C11H12O7 | 5.40 | 255.05 | 193.05 | 179.03 | 165.05 | 149.06 | 72.99 | |
| 9 | Syringic acid-O-hexoside | C15H20O10 | 10.58 | 359.10 | 197.05 | 182.02 | 167.00 | 153.05 | 138.03 | |
| 10 | Kynurenic acid | C10H7NO3 | 13.07 | 190.05 | 162.06 | 144.04 | 116.05 | |||
| 11 | Caffeic acid | C9H8O4 | 14.32 | 179.03 | 135.04 | 107.05 | ||||
| 12 | Unidentified tannin | C34H26O23 | 14.40 | 801.08 | 649.07 | 561.09 | 499.07 | 347.06 | 301.00 | |
| 13 | Trigalloylhexose isomer 1 | C27H24O18 | 14.48 | 635.09 | 483.08 | 465.07 | 313.06 | 169.01 | 125.02 | |
| 14 | Digalloylhexose | C20H20O14 | 15.21 | 483.08 | 331.07 | 313.06 | 271.05 | 169.01 | 125.02 | |
| 15 | Trigalloylhexose isomer 2 | C27H24O18 | 15.40 | 635.09 | 483.08 | 465.07 | 313.06 | 169.01 | 125.02 | |
| 16 | Castalin or vescalin | C27H20O18 | 15.69 | 631.06 | 450.99 | 301.00 | 299.99 | 298.98 | 270.99 | |
| 17 | Methylcoumarin | C10H8O2 | 15.92 | 161.06 | 133.07 | 105.07 | 91.05 | |||
| 18 | Castalin or vescalin | C27H20O18 | 16.38 | 631.06 | 451.00 | 301.00 | 299.98 | 298.98 | 270.99 | |
| 19 | Trigalloylhexose isomer 3 | C27H24O18 | 16.57 | 635.09 | 483.08 | 465.07 | 313.06 | 169.01 | 125.02 | |
| 20 | Ethyl syringate | C11H14O5 | 16.69 | 227.09 | 181.05 | 155.07 | 140.05 | 123.04 | 95.05 | |
| 21 | Feruloylhexose isomer 1 | C16H20O9 | 16.75 | 355.10 | 295.08 | 235.06 | 193.05 | 175.04 | 134.04 | |
| 22 | Trigalloylhexose isomer 4 | C27H24O18 | 17.32 | 635.09 | 483.08 | 465.07 | 313.06 | 169.01 | 125.02 | |
| 23 | Synapoylhexose isomer 1 | C17H22O10 | 17.57 | 385.11 | 325.09 | 265.07 | 223.06 | 205.05 | 190.03 | |
| 241 | 4-Coumaric acid | C9H8O3 | 17.71 | 163.04 | 119.05 | 93.03 | ||||
| 25 | Feruloylhexose isomer 2 | C16H20O9 | 17.85 | 355.10 | 295.08 | 235.06 | 193.05 | 175.04 | 134.04 | |
| 26 | Myrciaphenone B | C21H22O13 | 17.86 | 481.10 | 313.06 | 169.01 | 125.02 | |||
| 27 | Mangiferin (Aphloiol, Chinonin) | C19H18O11 | 18.33 | 421.08 | 343.05 | 331.05 | 301.04 | 272.03 | 259.03 | |
| 28 | Synapoylhexose isomer 2 | C17H22O10 | 18.53 | 385.11 | 325.09 | 265.07 | 223.06 | 205.05 | 190.03 | |
| 29 | Loliolide | C11H16O3 | 19.46 | 197.12 | 179.11 | 161.10 | 135.12 | 133.10 | 107.09 | |
| 30 | Tetragalloylhexose | C34H28O22 | 19.49 | 787.10 | 635.09 | 617.08 | 465.07 | 313.06 | 169.01 | |
| 31 | Ellagic acid-4-O-glucoside | C20H16O13 | 19.90 | 463.05 | 301.00 | 299.99 | 298.98 | |||
| 32 | Feruloyl-galloylhexose | C23H24O13 | 20.97 | 507.11 | 313.06 | 193.05 | 179.03 | 169.01 | 125.02 | |
| 33 | Quercetin-O-galloylhexoside isomer 1 | C28H24O16 | 21.32 | 615.10 | 463.09 | 301.04 | 300.03 | 255.03 | 169.01 | |
| 341 | Vitexin (Apigenin-8-C-glucoside) | C21H20O10 | 21.34 | 433.11 | 415.10 | 397.09 | 367.08 | 313.07 | 283.06 | |
| 35 | Quercetin-O-galloylhexoside isomer 2 | C28H24O16 | 21.56 | 615.10 | 463.09 | 301.04 | 300.03 | 271.02 | 169.01 | |
| 36 | Isovitexin (Apigenin-6-C-glucoside) | C21H20O10 | 22.26 | 433.11 | 415.10 | 397.09 | 367.08 | 313.07 | 283.06 | |
| 37 | Sinapoyl-galloylhexose | C24H26O14 | 22.37 | 537.12 | 325.09 | 265.07 | 223.06 | 169.01 | 125.02 | |
| 38 | Hyperoside (Quercetin-3-O-galactoside) | C21H20O12 | 22.64 | 463.09 | 301.04 | 300.03 | 271.03 | 255.03 | 151.00 | |
| 39 | Ellagic acid-O-pentoside | C19H14O12 | 22.75 | 433.04 | 301.00 | 299.99 | 298.98 | |||
| 401 | Isoquercitrin (Quercetin-3-O-glucoside) | C21H20O12 | 22.88 | 463.09 | 301.04 | 300.03 | 271.03 | 255.03 | 151.00 | |
| 41 | Acetylmangiferin | C21H20O12 | 23.10 | 463.09 | 331.05 | 301.04 | 271.03 | 259.03 | ||
| 42 | Ellagic acid | C14H6O8 | 23.36 | 301.00 | 284.00 | 257.01 | 229.01 | 201.02 | 185.02 | |
| 43 | Avicularin (Quercetin-3-O-arabinofuranoside) | C20H18O11 | 23.48 | 433.08 | 301.04 | 300.03 | 271.03 | 255.03 | 179.00 | |
| 44 | Unidentified terpenoid | C26H34O8 | 24.67 | 475.23 | 379.19 | 361.18 | 351.20 | 333.19 | 315.17 | |
| 45 | Isorhamnetin-O-hexoside | C22H22O12 | 24.70 | 477.10 | 315.05 | 314.04 | 285.04 | 271.03 | 243.03 | |
| 46 | Kaempferol-O-pentoside | C20H18O10 | 24.85 | 417.08 | 285.04 | 284.03 | 255.03 | 227.03 | ||
| 471 | Isorhamnetin-3-O-glucoside | C22H22O12 | 24.90 | 477.10 | 315.05 | 314.04 | 285.04 | 271.03 | 243.03 | |
| 48 | 4-Methoxycinnamic acid | C10H10O3 | 25.32 | 179.07 | 161.06 | 133.07 | 105.07 | 79.05 | ||
| 49 | 3-O-Methylellagic acid | C15H8O8 | 25.66 | 315.01 | 299.99 | 244.00 | ||||
| 50 | Tetrahydroxyxanthone | C13H8O6 | 26.12 | 259.02 | 231.03 | 215.03 | 203.03 | 187.04 | ||
| 51 | Dihydroactinidiolide | C11H16O2 | 26.57 | 181.12 | 163.11 | 145.10 | 135.12 | 107.09 | 93.07 | |
| 521 | Naringenin (4’,5,7-trihydroxyflavanone) | C15H12O5 | 27.17 | 271.06 | 177.02 | 151.00 | 119.05 | 107.01 | 93.03 | |
| 53 | Di-O-methylellagic acid isomer 1 | C16H10O8 | 27.41 | 329.03 | 314.01 | 298.98 | 270.99 | |||
| 54 | 3,3’-Di-O-methylellagic acid | C16H10O8 | 27.81 | 329.03 | 314.01 | 298.98 | 270.99 | |||
| 55 | Scilliglaucosidin | C24H30O5 | 27.88 | 399.22 | 381.21 | 363.20 | 345.18 | 335.20 | ||
| 56 | Di-O-methylellagic acid isomer 2 | C16H10O8 | 28.22 | 329.03 | 314.01 | 298.98 | 270.99 | |||
| 57 | 3,3’,4-Tri-O-methylellagic acid | C17H12O8 | 30.16 | 343.05 | 328.02 | 313.00 | 297.98 | 285.00 | ||
| 58 | 3,3’,4,4’-Tetra-O-methylellagic acid | C18H14O8 | 31.99 | 359.08 | 344.05 | 343.05 | 329.03 | 313.04 | ||
| 59 | Ginsenoside Ro or isomer | C48H76O19 | 36.24 | 955.49 | 793.44 | 731.44 | 613.37 | 569.39 | 551.38 | |
| 60 | Cynarasaponin C or isomer | C42H66O14 | 37.18 | 793.44 | 631.38 | 587.40 | 569.39 | 497.36 | 455.35 | |
| 61 | Ginsenoside Ro or isomer | C48H76O19 | 39.73 | 955.49 | 793.45 | 731.44 | 569.38 | 551.38 | 455.35 | |
| 62 | Unidentified saponin | C48H76O19 | 40.27 | 955.49 | 893.48 | 793.44 | 731.44 | 551.38 | 455.35 | |
| 63 | Hexadecanedioic acid | C16H30O4 | 40.29 | 285.21 | 267.20 | 223.21 | 57.03 |
| Extracts | AChE (mg GALAE/g Extract) | BChE (mg GALAE/g Extract) | Tyrosinase (mg KAE/g Extract) | α-Amylase (mmol ACAE/g Extract) | α-Glucosidase (mmol ACAE/g Extract) |
|---|---|---|---|---|---|
| EA | 4.43 ± 0.15 a | 1.27 ± 0.06 | 127.36 ± 0.98 b | 0.97 ± 0.06 a | 15.22 ± 0.11 |
| MeOH | 4.00 ± 0.05 a | na | 136.51 ± 0.70 a | 0.68 ± 0.03 b | na |
| Water | 3.15 ± 0.02 b | na | 81.63 ± 2.10 c | 0.21 ± 0.01 c | na |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sinan, K.I.; Chiavaroli, A.; Orlando, G.; Bene, K.; Zengin, G.; Cziáky, Z.; Jekő, J.; Fawzi Mahomoodally, M.; Picot-Allain, M.C.N.; Menghini, L.; et al. Biopotential of Bersama abyssinica Fresen Stem Bark Extracts: UHPLC Profiles, Antioxidant, Enzyme Inhibitory, and Antiproliferative Propensities. Antioxidants 2020, 9, 163. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9020163
Sinan KI, Chiavaroli A, Orlando G, Bene K, Zengin G, Cziáky Z, Jekő J, Fawzi Mahomoodally M, Picot-Allain MCN, Menghini L, et al. Biopotential of Bersama abyssinica Fresen Stem Bark Extracts: UHPLC Profiles, Antioxidant, Enzyme Inhibitory, and Antiproliferative Propensities. Antioxidants. 2020; 9(2):163. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9020163
Chicago/Turabian StyleSinan, Kouadio Ibrahime, Annalisa Chiavaroli, Giustino Orlando, Kouadio Bene, Gokhan Zengin, Zoltán Cziáky, József Jekő, Mohamad Fawzi Mahomoodally, Marie Carene Nancy Picot-Allain, Luigi Menghini, and et al. 2020. "Biopotential of Bersama abyssinica Fresen Stem Bark Extracts: UHPLC Profiles, Antioxidant, Enzyme Inhibitory, and Antiproliferative Propensities" Antioxidants 9, no. 2: 163. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9020163