Selenium Biofortification Impacts the Nutritive Value, Polyphenolic Content, and Bioactive Constitution of Variable Microgreens Genotypes

 ,
,  ,
,  ,
,  , , , ,
, , , ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Chamber Conditions
2.2. Plant Material and Growth Chamber Conditions
2.3. Dry Matter and Nitrate Content Analysis
2.4. Mineral Analysis by ICP-OES and Consumer Safety of Se-Enriched Microgreens
2.5. Separation and Quantification of Carotenoids by HPLC-DAD
2.6. Extraction and Analysis of Polyphenols by UHPLC-Q-Orbitrap HRMS
2.7. Statistics of Experimental Data
3. Results and Discussion
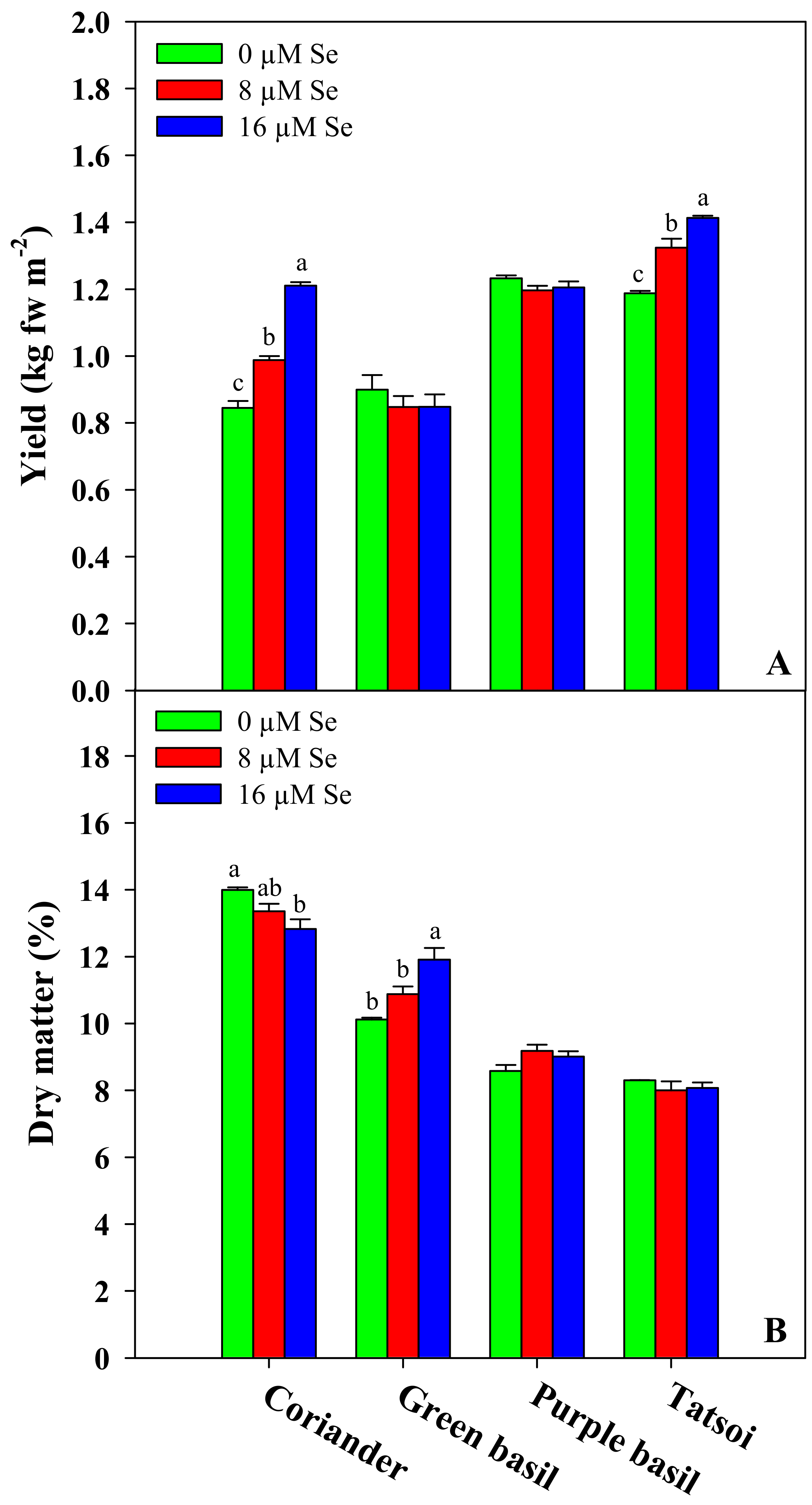
3.1. Fresh Biomass Yield and Dry Matter Content
3.2. Nitrate Content and Mineral Composition
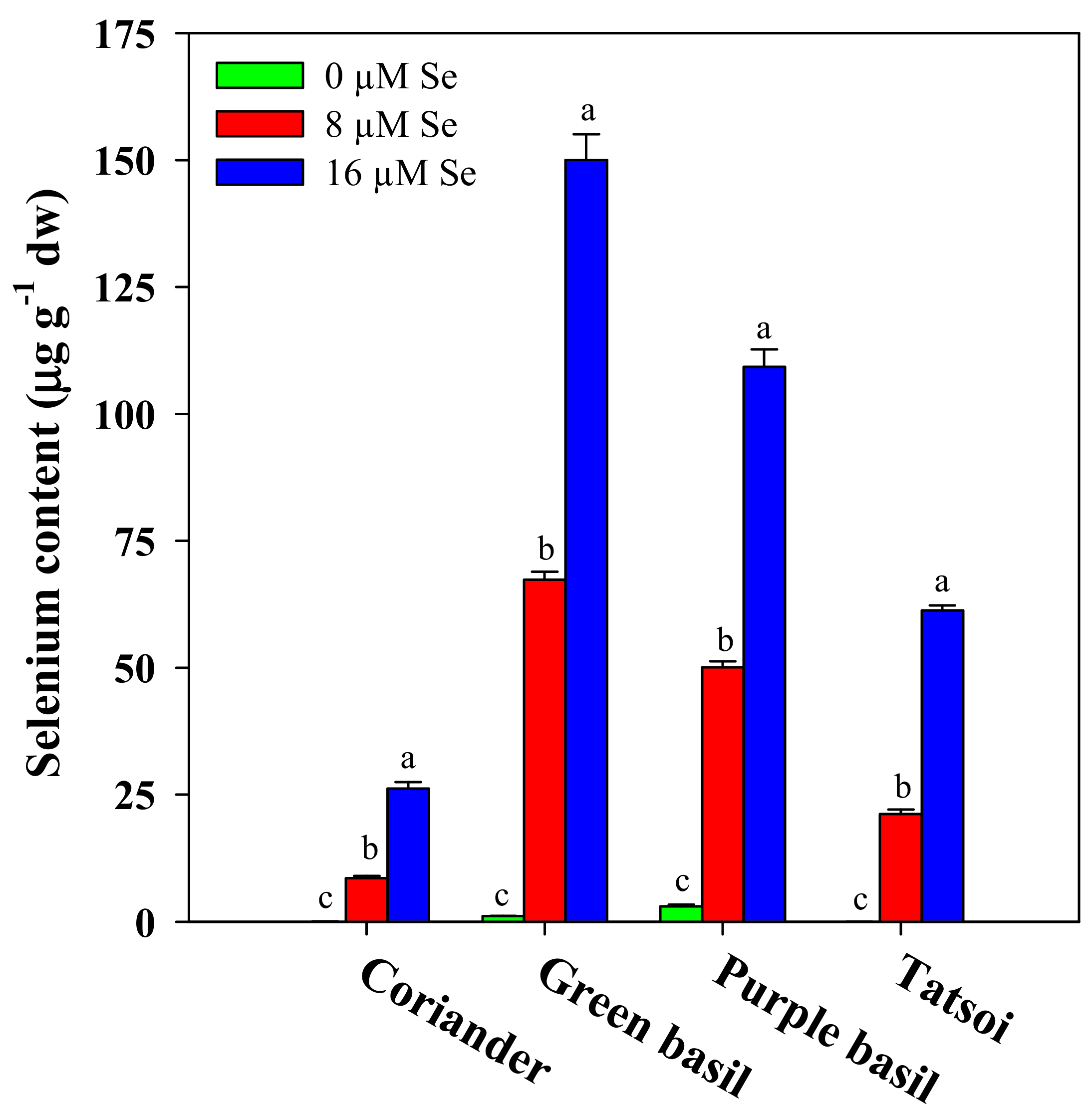
3.3. Selenium Biofortification and Consumer Safety
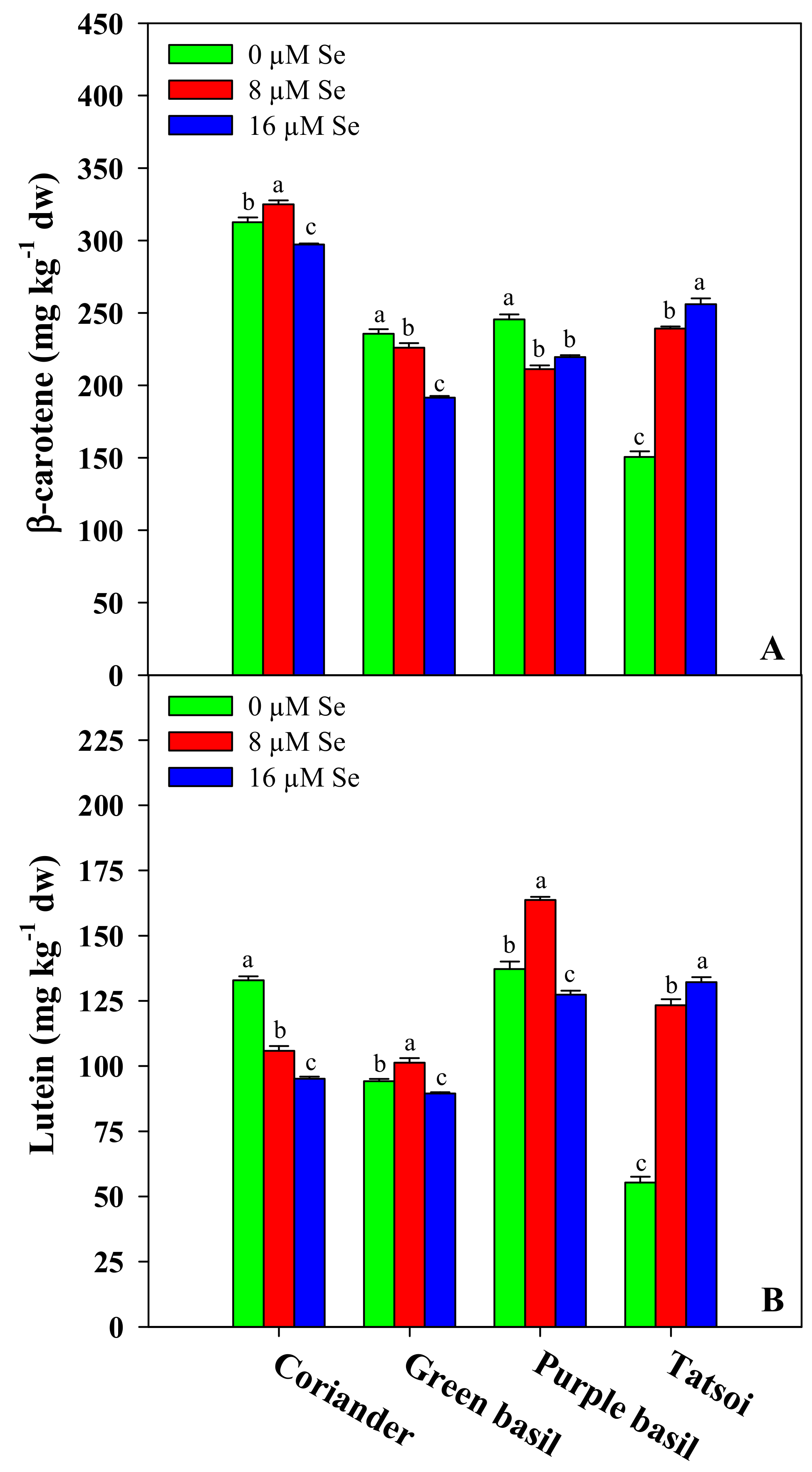
3.4. Target Carotenoids
3.5. Phenolic Compounds Profiling
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Di Gioia, F.; De Bellis, P.; Mininni, C.; Santamaria, P.; Serio, F. Physicochemical, agronomical and microbiological evaluation of alternative growing media for the production of rapini (Brassica rapa L.) microgreens. J. Sci. Food Agric. 2017, 97, 1212–1219. [Google Scholar] [CrossRef] [PubMed]
- Kyriacou, M.C.; El-Nakhel, C.; Graziani, G.; Pannico, A.; Soteriou, G.A.; Giordano, M. Functional quality in novel food sources: Genotypic variation in the nutritive and phytochemical composition of thirteen microgreens species. Food Chem. 2019, 277, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Xiao, Z.; Lin, L.Z.; Lester, G.E.; Wang, Q.; Harnly, J.M.; Chen, P. Profiling polyphenols in five Brassica species microgreens by UHPLC-PDA-ESI/HRMSn. J. Agric. Food Sci. 2013, 61, 10960–10970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyriacou, M.C.; Rouphael, Y.; Di Gioia, F.; Kyratzis, A.; Serio, F.; Renna, M.; De Pascale, S.; Santamaria, P. Micro-scale vegetable production and the rise of microgreens. Trends Food Sci. Technol. 2016, 57, 103–115. [Google Scholar] [CrossRef]
- Yadav, L.P.; Koley, T.K.; Tripathi, A.; Singh, S. Antioxidant potentiality and mineral content of summer season leafy greens: Comparison at mature and microgreen stages using chemometric. Agric. Res. 2019, 8, 165–175. [Google Scholar] [CrossRef]
- Pinto, E.; Almeida, A.A.; Aguiar, A.A.; Ferreira, I.M.P.L.V.O. Comparison between the mineral profile and nitrate content of microgreens and mature lettuces. J. Food Comp. Anal. 2015, 37, 38–43. [Google Scholar] [CrossRef]
- Kopppertcress. Available online: https://www.koppertcress.com/en/nieuws/koppert-cress-leading-the-way-with-new-educational-programme-on-microgreens (accessed on 20 December 2019).
- Xiao, Z.; Lester, G.E.; Park, E.; Saftner, R.A.; Luo, Y.; Wang, Q. Evaluation and correlation of sensory attributes and chemical compositions of emerging fresh produce: Microgreens. Postharvest Biol. Technol. 2015, 110, 140–148. [Google Scholar] [CrossRef]
- Puccinelli, M.; Malorgio, F.; Rosellini, I.; Pezzarossa, B. Production of selenium-biofortified microgreens from selenium-enriched seeds of basil. J. Sci. Food Agric. 2019, 99, 5601–5605. [Google Scholar] [CrossRef]
- Islam, M.Z.; Park, B.J.; Kang, H.M.; Lee, Y.T. Influence of selenium biofortification on the bioactive compounds and antioxidant activity of wheat microgreen extract. Food Chem. 2020, 309, 125763. [Google Scholar] [CrossRef]
- Rayman, M.P. The argument for increasing selenium intake. Proc. Nutr. Soc. 2002, 61, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.D.; Droz, B.; Greve, P.; Gottschalk, P.; Poffet, D.; McGrath, S.P. Selenium deficiency risk predicted to increase under future climate change. Proc. Natl. Acad. Sci. USA 2017, 114, 2848–2853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvalho, S.M.; Vasconcelos, M.W. Producing more with less: Strategies and novel technologies for plant-based food biofortification. Food Res. Int. 2013, 54, 961–971. [Google Scholar] [CrossRef]
- Rayman, M.P. The importance of selenium to human health. Lancet 2000, 356, 233–241. [Google Scholar] [CrossRef] [Green Version]
- Finley, J.W. Proposed criteria for assessing the efficacy of cancer reduction by plant foods enriched in carotenoids, glucosinolates, polyphenols and selenocompounds. Ann. Bot. 2005, 95, 1075–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malagoli, M.; Schiavon, M.; Dall’Acqua, S.; Pilon-Smits, E.A. Effects of selenium biofortification on crop nutritional quality. Front. Plant Sci. 2015, 6, 280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, L.J.; Meacham, S.L.; Kruskall, L.J. The antioxidants-vitamin C, vitamin E, selenium, and carotenoids. J. Agromed. 2003, 9, 65–82. [Google Scholar] [CrossRef]
- Burk, R.F.; Norsworthy, B.K.; Hill, K.E.; Motley, A.K.; Byrne, D.W. Effects of chemical form of selenium on plasma biomarkers in a high-dose human supplementation trial. Cancer Epidemiol. Biomark. 2006, 15, 804–810. [Google Scholar] [CrossRef] [Green Version]
- Finley, J.W. Increased intakes of selenium-enriched foods may benefit human health. J. Sci. Food Agric. 2007, 87, 1620–1629. [Google Scholar] [CrossRef]
- Pilon-Smits, E.A.; Quinn, C.F. Selenium metabolism in plants. In Cell Biology of Metals and Nutrients; Springer: Berlin/Heidelberg, Germany, 2010; pp. 225–241. [Google Scholar] [CrossRef]
- Vatansever, R.; Ozyigit, I.I.; Filiz, E. Essential and beneficial trace elements in plants, and their transport in roots: A review. Appl. Biochem. Biotechnol. 2017, 181, 464–482. [Google Scholar] [CrossRef]
- Chauhan, R.; Awasthi, S.; Srivastava, S.; Dwivedi, S.; Pilon-Smits, E.A.; Dhankher, O.P. Understanding selenium metabolism in plants and its role as a beneficial element. Crit. Rev. Environ. Sci. Technol. 2019, 49, 1937–1958. [Google Scholar] [CrossRef]
- Hartikainen, H. Biogeochemistry of selenium and its impact on food chain quality and human health. J. Trace Elem. Med. Biol. 2005, 18, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Lyons, G.H.; Genc, Y.; Soole, K.M.; Stangoulis, J.C.R.; Liu, F.; Graham, R.D. Selenium increases seed production in Brassica. Plant Soil 2009, 318, 73–80. [Google Scholar] [CrossRef]
- Ardebili, Z.O.; Ardebili, N.O.; Jalili, S.; Safiallah, S. The modified qualities of basil plants by selenium and/or ascorbic acid. Turk. J. Bot. 2015, 39, 401–407. [Google Scholar] [CrossRef]
- Skrypnik, L.; Novikova, A.; Tokupova, E. Improvement of phenolic compounds, essential oil content and antioxidant properties of sweet basil (Ocimum basilicum L.) depending on type and concentration of selenium application. Plants 2019, 8, 458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dall’Acqua, S.; Ertani, A.; Pilon-Smits, E.A.; Fabrega-Prats, M.; Schiavon, M. Selenium biofortification differentially affects sulfur metabolism and accumulation of phytochemicals in two rocket species (Eruca Sativa Mill. and Diplotaxis Tenuifolia) grown in hydroponics. Plants 2019, 8, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.F.; McGrath, S.P.; Zhao, F.J. Selenium uptake, translocation and speciation in wheat supplied with selenate or selenite. New Phytol. 2008, 178, 92–102. [Google Scholar] [CrossRef]
- Lyons, G.H.; Ortiz-Monasterio, I.; Stangoulis, J.; Graham, R. Selenium concentration in wheat grain: Is there sufficient genotypic variation to use in breeding? Plant Soil 2005, 269, 369–380. [Google Scholar] [CrossRef]
- Hawrylak-Nowak, B. Comparative effects of selenite and selenate on growth and selenium accumulation in lettuce plants under hydroponic conditions. Plant Growth Regul. 2013, 70, 149–157. [Google Scholar] [CrossRef] [Green Version]
- White, P.J. Selenium metabolism in plants. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2018, 1862, 2333–2342. [Google Scholar] [CrossRef]
- Puccinelli, M.; Malorgio, F.; Rosellini, I.; Pezzarossa, B. Uptake and partitioning of selenium in basil (Ocimum basilicum L.) plants grown in hydroponics. Sci. Hortic. 2017, 225, 271–276. [Google Scholar] [CrossRef]
- Rouphael, Y.; Kyriacou, M.C. Enhancing quality of fresh vegetables through salinity eustress and biofortification applications facilitated by soilless cultivation. Front. Plant Sci. 2018, 9, 1254. [Google Scholar] [CrossRef] [PubMed]
- El-Nakhel, C.; Pannico, A.; Kyriacou, M.C.; Giordano, M.; De Pascale, S.; Rouphael, Y. Macronutrient deprivation eustress elicits differential secondary metabolites in red and green-pigmented butterhead lettuce grown in a closed soilless system. J. Sci. Food Agric. 2019, 99, 6962–6972. [Google Scholar] [CrossRef] [PubMed]
- Lyons, G.H.; Stangoulis, J.C.; Graham, R.D. Exploiting micronutrient interaction to optimize biofortification programs: The case for inclusion of selenium and iodine in the HarvestPlus program. Nutr. Rev. 2004, 62, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Yang, F.; Xu, J.; Hu, Y.; Hu, Q.; Zhang, Y. Determination of selenium concentration of rice in China and effect of fertilization of selenite and selenate on selenium content of rice. J. Agric. Food Chem. 2002, 50, 5128–5130. [Google Scholar] [CrossRef]
- Hernández-Hernández, H.; Quiterio-Gutiérrez, T.; Cadenas-Pliego, G.; Ortega-Ortiz, H.; Hernández-Fuentes, A.D.; Cabrera de la Fuente, M.; Valdés-Reyna, V.; Juárez-Maldonado, A. Impact of selenium and copper nanoparticles on yield, antioxidant system, and fruit quality of tomato plants. Plants 2019, 8, 355. [Google Scholar] [CrossRef] [Green Version]
- Turakainen, M.; Hartikainen, H.; Seppänen, M.M. Effects of selenium treatments on potato (Solanum tuberosum L.) growth and concentrations of soluble sugars and starch. J. Agric. Food Chem. 2004, 52, 5378–5382. [Google Scholar] [CrossRef]
- Hermosillo-Cereceres, M.A.; Sanchez-Chavez, E.; Balandran-Quintana, R.R.; Mendoza-Wilson, A.M.; Guevara-Aguilar, A.; Munoz-Marquez, E. Ionome variation in bean plant growth under different Se forms and application rates. J. Food Agric. Environ. 2011, 9, 374–378. [Google Scholar] [CrossRef]
- Jerše, A.; Maršić, N.K.; Kroflič, A.; Germ, M.; Šircelj, H.; Stibilj, V. Is foliar enrichment of pea plants with iodine and selenium appropriate for production of functional food? Food Chem. 2018, 267, 368–375. [Google Scholar] [CrossRef]
- Sabatino, L.; Ntatsi, G.; Iapichino, G.; D’Anna, F.; De Pasquale, C. Effect of selenium enrichment and type of application on yield, functional quality and mineral composition of curly endive grown in a hydroponic system. Agronomy 2019, 9, 207. [Google Scholar] [CrossRef] [Green Version]
- Bachiega, P.; Salgado, J.M.; de Carvalho, J.E.; Ruiz, A.L.T.; Schwarz, K.; Tezotto, T.; Morzelle, M.C. Antioxidant and antiproliferative activities in different maturation stages of broccoli (Brassica oleracea Italica) biofortified with selenium. Food Chem. 2016, 190, 771–776. [Google Scholar] [CrossRef] [Green Version]
- Pannico, A.; El Nakhel, C.; Kyriacou, M.C.; Giordano, M.; Stazi, S.R.; De Pascale, S.; Rouphael, Y. Combating micronutrient deficiency and enhancing food functional quality through selenium fortification of select lettuce genotypes grown in a closed soilless system. Front. Plant Sci. 2019, 10, 1495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poldma, P.; Tonutare, T.; Viitak, A.; Luik, A.; Moor, U. Effect of selenium treatment on mineral nutrition, bulb size, and antioxidant properties of garlic (Allium sativum L.). J. Agric. Food Chem. 2011, 59, 5498–5503. [Google Scholar] [CrossRef] [PubMed]
- Golubkina, N.; Zamana, S.; Seredin, T.; Poluboyarinov, P.; Sokolov, S.; Baranova, H.; Pietroantonio, L.; Caruso, G. Effect of selenium biofortification and beneficial microorganism inoculation on yield, quality and antioxidant properties of shallot bulbs. Plants 2019, 8, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Germ, M.; Stibilj, V.; Šircelj, H.; Jerše, A.; Kroflič, A.; Golob, A.; Maršić, N.K. Biofortification of common buckwheat microgreens and seeds with different forms of selenium and iodine. J. Sci. Food Agric. 2019, 99, 4353–4362. [Google Scholar] [CrossRef] [PubMed]
- Volpe, M.G.; Nazzaro, M.; Di Stasio, M.; Siano, F.; Coppola, R.; De Marco, A. Content of micronutrients, mineral and trace elements in some Mediterranean spontaneous edible herbs. Chem. Cent. J. 2015, 9, 57. [Google Scholar] [CrossRef] [Green Version]
- Iris. Integrated Risk Information System—Database; Environmental Protection Agency: Washington, DC, USA, 2011. [Google Scholar]
- D’Amato, R.; Fontanella, M.C.; Falcinelli, B.; Beone, G.M.; Bravi, E.; Marconi, O. Selenium biofortification in rice (Oryza sativa L.) sprouting: Effects on Se yield and nutritional traits with focus on phenolic acid profile. J. Agric. Food Chem. 2018, 66, 4082–4090. [Google Scholar] [CrossRef]
- Singh, M.; Singh, N.; Bhandari, D.K. Interaction of selenium and sulfur on the growth and chemical composition of raya. Soil Sci. 1980, 129, 238–244. [Google Scholar] [CrossRef]
- Hartikainen, H.; Xue, T. The promotive effect of selenium on plant growth as triggered by ultraviolet irradiation. J. Environ. Qual. 1999, 28, 1372–1375. [Google Scholar] [CrossRef]
- Saffaryazdi, A.; Lahouti, M.; Ganjeali, A.; Bayat, H. Impact of selenium supplementation on growth and selenium accumulation on spinach (Spinacia oleracea L.) plants. Not. Sci. Biol. 2012, 4, 95–100. [Google Scholar] [CrossRef] [Green Version]
- Ramos, S.J.; Faquin, V.; Guilherme, L.R.G.; Castro, E.M.; Ávila, F.W.; Carvalho, G.S. Selenium biofortification and antioxidant activity in lettuce plants fed with selenate and selenite. Plant Soil Environ. 2010, 56, 584–588. [Google Scholar] [CrossRef] [Green Version]
- Chauhan, R.; Awasthi, S.; Tripathi, P.; Mishra, S.; Dwivedi, S.; Niranjan, A.; Tripathi, R.D. Selenite modulates the level of phenolics and nutrient element to alleviate the toxicity of arsenite in rice (Oryza sativa L.). Ecotoxicol. Environ. Saf. 2017, 138, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Feng, R.; Wei, C.; Tu, S. The roles of selenium in protecting plants against abiotic stresses. Environ. Exp. Bot. 2013, 87, 58–68. [Google Scholar] [CrossRef]
- Hartikainen, H.; Xue, T.; Piironen, V. Selenium as an anti-oxidant and pro-oxidant in ryegrass. Plant Soil 2000, 225, 193–200. [Google Scholar] [CrossRef]
- Terry, N.; Zayed, A.M.; De Souza, M.P.; Tarun, A.S. Selenium in higher plants. Annu. Rev. Plant Biol. 2000, 51, 401–432. [Google Scholar] [CrossRef] [Green Version]
- Colla, G.; Kim, H.J.; Kyriacou, M.C.; Rouphael, Y. Nitrate in fruits and vegetables. Sci. Hortic. 2018, 237, 221–238. [Google Scholar] [CrossRef]
- Commission Regulation (EU) No 1258/2011. Amending Regulation (EC) No 1881/2006 as regards maximum levels for nitrates in foodstuffs. Off. J. Eur. Union 2011, 320, 15–17. [Google Scholar]
- Rios, J.J.; Blasco, B.; Rosales, M.A.; Sanchez-Rodriguez, E.; Leyva, R.; Cervilla, L.M. Response of nitrogen metabolism in lettuce plants subjected to different doses and forms of selenium. J. Sci. Food Agric. 2010, 90, 1914–1919. [Google Scholar] [CrossRef]
- Nowak, J.; Kaklewski, K.; Ligocki, M. Influence of selenium on oxidoreductive enzymes activity in soil and in plants. Soil Biol. Biochem. 2004, 36, 1553–1558. [Google Scholar] [CrossRef]
- Lee, S.J.; Kang, H.M.; Kim, I.S. Effect of sodium selenate supplied condition by fertigation on the growth and content of minerals, ascorbic acid, nitrate, and selenium of some western vegetables. J. Bio-Environ. Control 2008, 17, 43–50. [Google Scholar]
- Ríos, J.J.; Blasco, B.; Cervilla, L.M.; Rubio-Wilhelmi, M.; Rosales, M.A.; Sanchez-Rodriguez, E. Nitrogen-use efficiency in relation to different forms and application rates of Se in lettuce plants. J. Plant Growth Regul. 2010, 29, 164–170. [Google Scholar] [CrossRef]
- Gupta, U.C.; Gupta, S.C. Sources and deficiency diseases of mineral nutrients in human health and nutrition: A review. Pedosphere 2014, 24, 13–38. [Google Scholar] [CrossRef]
- Xiao, Z.; Codling, E.E.; Luo, Y.; Nou, X.; Lester, G.E.; Wang, Q. Microgreens of Brassicaceae: Mineral composition and content of 30 varieties. J. Food Comp. Anal. 2016, 49, 87–93. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Huang, Z.Z. Selenium assimilation and nutrient element uptake in white clover and tall fescue under the influence of sulphate concentration and selenium tolerance of the plants. J. Exp. Bot. 1992, 43, 549–555. [Google Scholar] [CrossRef]
- Rios, J.J.; Blasco, B.; Leyva, R.; Sanchez-Rodriguez, E.; Rubio-Wilhelmi, M.M.; Romero, L. Nutritional balance changes in lettuce plant grown under different doses and forms of selenium. J. Plant Nutr. 2013, 36, 1344–1354. [Google Scholar] [CrossRef]
- Couto, E.; Boffetta, P.; Lagiou, P.; Ferrari, P.; Buckland, G.; Overvad, K. Mediterranean dietary pattern and cancer risk in the EPIC cohort. Br. J. Cancer 2011, 104, 1493–1499. [Google Scholar] [CrossRef] [Green Version]
- Silva, V.M.; Boleta, E.H.M.; Martins, J.T.; Mendes dos Santos, F.L.; da Rocha Silva, A.C.; Alcock, T.D. Agronomic biofortification of cowpea with selenium: Effects of selenate and selenite applications on selenium and phytate concentrations in seeds. J. Sci. Food Agric. 2019, 29, 371–379. [Google Scholar] [CrossRef]
- Young, A.J.; Lowe, G.M. Antioxidant and prooxidant properties of carotenoids. Arch. Biochem. Biophys. 2001, 385, 20–27. [Google Scholar] [CrossRef]
- O’Neill, M.E.; Carroll, Y.; Corridan, B.; Olmedilla, B.; Granado, F.; Blanco, I.; Van den Berg, H.; Hininger, I.; Rousell, A.M.; Chopra, M.; et al. A European carotenoid database to assess carotenoid intakes and its use in a five-country comparative study. Br. J. Nutr. 2001, 85, 499–507. [Google Scholar] [CrossRef]
- Rapp, L.M.; Maple, S.S.; Choi, J.H. Lutein and zeaxanthin concentrations in rod outer segment membranes from perifoveal and peripheral human retina. Investig. Ophthalmol. Vis. Sci. 2000, 41, 1200–1209. [Google Scholar]
- Naves, M.M.V.; Moreno, F.S. β-carotene and cancer chemoprevention: From epidemiological associations to cellular mechanisms of action. Nutr. Res. 1998, 18, 1807–1824. [Google Scholar] [CrossRef]
- Jiang, C.; Zu, C.; Shen, J.; Shao, F.; Li, T. Effects of selenium on the growth and photosynthetic characteristics of flue-cured tobacco (Nicotiana tabacum L.). Acta Soc. Bot. Pol. 2015, 84, 71–77. [Google Scholar] [CrossRef] [Green Version]
- Akladious, S.A. Influence of different soaking times with selenium on growth, metabolic activities of wheat seedlings under low temperature stress. Afr. J. Biotechnol. 2012, 11, 14792–14804. [Google Scholar]
- Sams, C.E.; Panthee, D.R.; Charron, C.S.; Kopsell, D.A.; Yuan, J.S. Selenium regulates gene expression for glucosinolate and carotenoid biosynthesis in Arabidopsis. J. Am. Soc Hortic. Sci. 2011, 136, 23–34. [Google Scholar] [CrossRef] [Green Version]
- El-Zaeddi, H.; Calín-Sánchez, Á.; Nowicka, P.; Martínez-Tomé, J.; Noguera-Artiaga, L.; Burló, F.; Wojdyło, A.; Carbonell-Barrachina, Á.A. Preharvest treatments with malic, oxalic, and acetylsalicylic acids affect the phenolic composition and antioxidant capacity of coriander, dill and parsley. Food Chem. 2017, 226, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Flanigan, P.M.; Niemeyer, E.D. Effect of cultivar on phenolic levels, anthocyanin composition, and antioxidant properties in purple basil (Ocimum basilicum L.). Food Chem. 2014, 164, 518–526. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Wu, L.; Zhang, H.; Wu, W.; Zhang, M.; Li, X. Regulation of the phenylpropanoid pathway: A mechanism of selenium tolerance in peanut (Arachis hypogaea L.) seedlings. J. Agric. Food Chem. 2016, 64, 3626–3635. [Google Scholar] [CrossRef] [PubMed]
- Ríos, J.J.; Blasco, B.; Cervilla, L.M.; Rubio-Wilhelmi, M.M.; Ruiz, J.M.; Romero, L. Regulation of sulphur assimilation in lettuce plants in the presence of selenium. Plant Growth Regul. 2008, 56, 43. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Variables | Source of Variance | Variables | Source of Variance | ||||
|---|---|---|---|---|---|---|---|
| Genotype (G) | Selenium (S) | G × S | Genotype (G) | Selenium (S) | G × S | ||
| Fresh yield | *** | *** | *** | kaempferol-3-O(caffeoyl)sophoroside-7-O-glucoside | *** | *** | *** |
| Dry matter | *** | ns | *** | isorhamnetin-3-gentiobioside | na | *** | na |
| Nitrate | *** | ** | ns | kaempferol-3-O-(coumaroyl)soph-7-O-glucoside | *** | *** | *** |
| P | *** | *** | *** | luteolin-7-O-glucoside | *** | ns | *** |
| K | *** | *** | *** | apigenin-malonil-glucoside | *** | *** | *** |
| Ca | *** | *** | *** | kaempferol-3-O(feruoyll)sophoroside-7-O-glucoside | *** | *** | *** |
| Mg | *** | *** | *** | coumaroyl quinic acid | *** | *** | *** |
| Na | *** | *** | *** | rutin | *** | *** | *** |
| Fe | *** | *** | *** | apigenin-7-O-rutinoside | *** | *** | *** |
| Zn | *** | *** | *** | quercetin-3-O-glucoside | *** | *** | *** |
| Mn | *** | ** | *** | feruloyl quinic acid | *** | *** | *** |
| Se | *** | *** | *** | rosmarinic acid | *** | * | *** |
| Se intake | *** | *** | *** | cirsiliol | *** | *** | *** |
| RDA-Se | *** | *** | *** | ferulic acid | *** | *** | *** |
| HQgv | *** | *** | *** | luteolin-3-O-rutinoside | *** | *** | *** |
| lutein | *** | *** | *** | kaempferol-7-O-glucoside | *** | *** | *** |
| β-carotene | *** | *** | *** | dicaffeoylquinic acid | *** | *** | *** |
| cichoric acid | *** | *** | *** | kaempferol-3-O-rutinoside | *** | *** | *** |
| chlorogenic acid | *** | ns | ns | quercetin-rhamnoside | *** | ns | ns |
| caffeic acid | *** | *** | *** | luteolin-malonil-hexose | ns | ns | ns |
| kaempferol-3-O-sophoroside-7-O-glucoside | na | *** | na | apigenin-7-rhamnoside-4-rutinoside | na | *** | na |
| quercetin-sophoroside | *** | *** | *** | apigenin-7-O-glucoside | *** | *** | *** |
| caffeic acid hexoside isomers | *** | *** | *** | ||||
| 2-xylosylvitexin | *** | ns | ns | Total polyphenols | *** | ns | *** |
| Genotype | Selenium | Nitrate | P | K | Ca | Mg | Na | Fe | Zn | Mn |
|---|---|---|---|---|---|---|---|---|---|---|
| (mg kg−1 fw) | (mg g−1 dw) | (mg g−1 dw) | (mg g−1 dw) | (mg g−1 dw) | (mg g−1 dw) | (μg g−1 dw) | (μg g−1 dw) | (μg g−1 dw) | ||
| Coriander | 0 µM | 18.5 ± 2.75 a | 3.04 ± 0.08 c | 7.23 ± 0.23 b | 3.15 ± 0.04 b | 2.17 ± 0.08 b | 0.61 ± 0.01 b | 20.45 ± 0.06 c | 23.18 ± 0.39 c | 6.61 ± 0.17 b |
| 8 µM | 10.0 ± 0.43 b | 3.49 ± 0.04 b | 8.09 ± 0.39 ab | 3.41 ± 0.14 b | 2.36 ± 0.03 ab | 0.63 ± 0.03 b | 22.28 ± 0.58 b | 27.22 ± 0.96 b | 7.64 ± 0.59 b | |
| 16 µM | 9.5 ± 0.58 b | 3.72 ± 0.03 a | 9.27 ± 0.43 a | 4.15 ± 0.02 a | 2.58 ± 0.09 a | 0.72 ± 0.01 a | 25.42 ± 0.08 a | 36.79 ± 0.21 a | 9.30 ± 0.05 a | |
| Significance | * | *** | * | *** | * | * | *** | *** | ** | |
| Green basil | 0 µM | 216.1 ± 8.34 a | 1.87 ± 0.12 c | 9.19 ± 0.22 b | 8.16 ± 0.49 b | 2.15 ± 0.03 b | 0.49 ± 0.01 b | 36.73 ± 0.27 a | 64.56 ± 4.52 a | 13.18 ± 0.39 c |
| 8 µM | 159.3 ± 4.05 b | 3.05 ± 0.05 b | 12.80 ± 0.08 a | 9.95 ± 0.42 a | 3.04 ± 0.07 a | 0.54 ± 0.02 b | 35.27 ± 0.25 b | 27.88 ± 0.22 b | 21.27 ± 0.33 b | |
| 16 µM | 84.1 ± 7.63 c | 3.65 ± 0.21 a | 12.90 ± 0.16 a | 10.62 ± 0.54 a | 3.56 ± 0.28 a | 0.56 ± 0.01 a | 36.83 ± 0.51 a | 24.97 ± 1.16 b | 22.85 ± 0.45 a | |
| Significance | *** | *** | *** | * | ** | * | * | *** | *** | |
| Purple basil | 0 µM | 539.9 ± 13.3 a | 3.48 ± 0.19 b | 11.11 ± 0.47 b | 7.30 ± 0.46 b | 2.87 ± 0.13 b | 0.67 ± 0.01 a | 48.71 ± 0.63 c | 35.83 ± 0.31 c | 10.20 ± 0.18 c |
| 8 µM | 451.2 ± 21.9 b | 4.12 ± 0.03 a | 13.03 ± 0.38 a | 8.88 ± 0.34 a | 3.29 ± 0.04 a | 0.46 ± 0.01 b | 51.93 ± 0.90 b | 38.80 ± 0.16 b | 12.41 ± 0.18 b | |
| 16 µM | 453.3 ± 19.8 b | 4.34 ± 0.11 a | 12.63 ± 0.03 a | 9.44 ± 0.08 a | 3.36 ± 0.07 a | 0.48 ± 0.01 b | 57.16 ± 0.68 a | 41.36 ± 0.24 a | 13.32 ± 0.25 a | |
| Significance | * | ** | * | ** | * | *** | *** | *** | *** | |
| Tatsoi | 0 µM | 80.6 ± 2.98 a | 4.48 ± 0.22 a | 4.11 ± 0.19 a | 4.42 ± 0.08 a | 3.33 ± 0.20 a | 0.75 ± 0.02 a | 52.56 ± 3.30 a | 43.22 ± 1.81 a | 46.06 ± 2.40 a |
| 8 µM | 63.7 ± 0.12 b | 3.44 ± 0.06 b | 2.86 ± 0.17 b | 2.85 ± 0.27 b | 2.12 ± 0.08 b | 0.39 ± 0.02 c | 29.98 ± 0.27 c | 34.75 ± 0.16 c | 28.81 ± 0.53 c | |
| 16 µM | 64.7 ± 0.97 b | 3.84 ± 0.04 b | 3.82 ± 0.15 a | 3.90 ± 0.17 a | 2.56 ± 0.10 b | 0.58 ± 0.03 b | 40.79 ± 1.61 b | 38.77 ± 0.05 b | 35.03 ± 1.46 b | |
| Significance | *** | ** | ** | ** | ** | *** | *** | ** | *** |
| Genotype | Selenium | Se Intake with 10 g fw of Microgreens | RDA-Se with 10 g fw of Microgreens | HQgv with 10 g fw of Microgreens |
|---|---|---|---|---|
| (μg day−1) | (%) | |||
| Coriander | 0 µM | 0.07 ± 0.0 c | 0.12 ± 0.0 c | 0.00 ± 0.00 c |
| 8 µM | 11.5 ± 0.8 b | 20.8 ± 1.5 b | 0.03 ± 0.00 b | |
| 16 µM | 33.6 ± 1.6 a | 61.0 ± 2.9 a | 0.08 ± 0.00 a | |
| Significance | *** | *** | *** | |
| Green basil | 0 µM | 1.12 ± 0.1 c | 2.03 ± 0.1 c | 0.00 ± 0.00 c |
| 8 µM | 73.2 ± 1.7 b | 133 ± 3.1 b | 0.18 ± 0.00 b | |
| 16 µM | 178 ± 5.5 a | 325 ± 10 a | 0.45 ± 0.01 a | |
| Significance | *** | *** | *** | |
| Purple basil | 0 µM | 2.60 ± 0.3 c | 4.73 ± 0.6 c | 0.01 ± 0.00 c |
| 8 µM | 46.0 ± 2.0 b | 83.7 ± 3.7 b | 0.12 ± 0.01 b | |
| 16 µM | 98.4 ± 1.5 a | 179 ± 2.6 a | 0.25 ± 0.00 a | |
| Significance | *** | *** | *** | |
| Tatsoi | 0 µM | 0.04 ± 0.0 c | 0.07 ± 0.0 c | 0.00 ± 0.00 c |
| 8 µM | 17.0 ± 1.2 b | 31.0 ± 2.2 b | 0.04 ± 0.00 b | |
| 16 µM | 49.5 ± 1.0 a | 90.0 ± 1.9 a | 0.12 ± 0.00 a | |
| Significance | *** | *** | *** |
| Polyphenol (µg g−1 dw) | Coriander | Green basil | Purple basil | Tatsoi | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 µM Se | 8 µM Se | 16 µM Se | Sig. | 0 µM Se | 8 µM Se | 16 µM Se | Sig. | 0 µM Se | 8 µM Se | 16 µM Se | Sig. | 0 µM Se | 8 µM Se | 16 µM Se | Sig. | |
| cichoric acid | nd | nd | nd | na | 841 ± 63.8 b | 2408 ± 26.6 a | 921 ± 30.0 b | *** | 813 ± 58.2 a | 533 ± 24.3 b | 481 ± 16.8 b | ** | nd | nd | nd | na |
| chlorogenic acid | 4504 ± 409 | 4526 ± 247 | 4519 ± 315 | ns | 208 ± 15.2 b | 272 ± 14.8 a | 171 ± 2.61 b | ** | 7.12 ± 0.18 a | 6.78 ± 0.20 a | 2.83 ± 0.29 b | *** | 12.8 ± 1.36 b | 19.4 ± 1.20 a | 12.4 ± 1.43 b | * |
| caffeic acid | 0.67 ± 0.04 b | 0.99 ± 0.05 a | 0.86 ± 0.04 a | ** | 11.2 ± 0.80 b | 17.1 ± 1.01 a | 8.88 ± 0.24 b | *** | 10.89 ± 0.86 a | 6.33 ± 0.43 b | 5.82 ± 0.16 b | *** | 0.70 ± 0.06 c | 0.87 ± 0.02 b | 1.35 ± 0.05 a | *** |
| kaempferol-3-O-sophoroside-7-O-glucoside | nd | nd | nd | na | nd | nd | nd | na | nd | nd | nd | na | 0.66 ± 0.01 c | 2.47 ± 0.17 b | 7.67 ± 0.46 a | *** |
| quercetin-sophoroside | 1.46 ± 0.02 a | 1.23 ± 0.04 b | 0.85 ± 0.02 c | *** | 0.65 ± 0.03 b | 1.49 ± 0.02 a | 0.42 ± 0.03 c | *** | 0.15 ± 0.00 a | 0.11 ± 0.01 b | 0.08 ± 0.00 c | *** | 12.4 ± 0.65 b | 26.0 ± 3.20 a | 23.9 ± 1.47 a | ** |
| caffeic acid hexoside isomers | 39.9 ± 0.68 a | 12.0 ± 0.19 c | 17.7 ± 1.09 b | *** | 6.26 ± 0.71 c | 17.6 ± 0.73 a | 9.45 ± 0.73 b | *** | 13.0 ± 0.62 a | 7.34 ± 0.71 b | 12.0 ± 0.67 a | ** | 212 ± 8.92 b | 194 ± 20.1 b | 367 ± 5.93 a | *** |
| 2-xylosylvitexin | nd | nd | nd | na | nd | nd | nd | na | 5.04 ± 0.56 | 4.12 ± 0.27 | 4.04 ± 0.14 | ns | 0.94 ± 0.08 b | 0.85 ± 0.04 b | 1.23 ± 0.06 a | * |
| kaempferol-3-O(caffeoyl)sophoroside-7-O-glucoside | nd | nd | nd | na | 0.09 ± 0.00 b | 0.15 ± 0.00 a | 0.07 ± 0.01 c | *** | nd | nd | nd | na | 94.6 ± 6.25 b | 93.2 ± 7.66 b | 190 ± 1.88 a | *** |
| isorhamnetin-3-gentiobioside | nd | nd | nd | na | nd | nd | nd | na | nd | nd | nd | na | 18.9 ± 0.96 b | 18.7 ± 0.50 b | 39.2 ± 1.99 a | *** |
| kaempferol-3-O-(coumaroyl)soph-7-O-glucoside | 0.81 ± 0.04 | 0.11 ± 0.01 | 0.04 ± 0.00 | *** | 0.01 ± 0.00 c | 0.02 ± 0.00 a | 0.02 ± 0.00 b | *** | 18.8 ± 0.62 a | 8.48 ± 0.25 b | 7.42 ± 0.63 b | *** | 7.09 ± 0.46 b | 7.62 ± 0.38 b | 15.3 ± 1.03 a | *** |
| luteolin-7-O-glucoside | 1.48 ± 0.01 b | 1.47 ± 0.01 b | 1.60 ± 0.05 a | * | 14.0 ± 1.00 b | 31.3 ± 1.19 a | 30.2 ± 1.09 a | *** | 39.6 ± 0.39 a | 24.4 ± 0.67 b | 22.7 ± 0.94 b | *** | 2.18 ± 0.26 b | 2.69 ± 0.14 b | 4.92 ± 0.07 a | *** |
| apigenin-malonil-glucoside | 0.04 ± 0.00 b | 0.05 ± 0.00 b | 0.07 ± 0.00 a | *** | 0.34 ± 0.01 c | 0.76 ± 0.03 a | 0.47 ± 0.02 b | *** | 1.10 ± 0.04 | 0.99 ± 0.02 | 1.04 ± 0.03 | ns | 0.05 ± 0.00 a | 0.02 ± 0.00 c | 0.04 ± 0.00 b | *** |
| kaempferol-3-O(feruoyll)sophoroside-7-O-glucoside | 4.65 ± 0.29 a | 0.31 ± 0.01 b | 0.06 ± 0.00 b | *** | 0.03 ± 0.00 a | 0.02 ± 0.00 b | 0.03 ± 0.00 a | *** | 0.59 ± 0.04 a | 0.26 ± 0.02 b | 0.27 ± 0.01 b | *** | 42.1 ± 2.54 b | 58.6 ± 3.29 b | 108 ± 7.91 a | *** |
| coumaroyl quinic acid | 3.21 ± 0.06 a | 0.60 ± 0.06 b | 0.10 ± 0.01 c | *** | 0.02 ± 0.00 c | 0.10 ± 0.01 b | 0.14 ± 0.00 a | *** | 0.05 ± 0.00 a | 0.03 ± 0.00 b | 0.01 ± 0.00 c | *** | 12.3 ± 0.23 b | 15.6 ± 1.21 b | 38.7 ± 1.68 a | *** |
| rutin | 3487 ± 48.9 c | 3857 ± 35.7 b | 4653 ± 86.8 a | *** | 31.2 ± 0.19 a | 13.3 ± 0.57 b | 14.1 ± 0.63 b | *** | 3.42 ± 0.06 | 3.11 ± 0.20 | 3.23 ± 0.05 | ns | 3.16 ± 0.29 b | 1.96 ± 0.05 b | 2.37 ± 0.02 a | ** |
| apigenin-7-O-rutinoside | 0.30 ± 0.02 a | 0.12 ± 0.05 b | 0.01 ± 0.00 c | *** | nd | nd | nd | na | nd | nd | nd | na | 4.55 ± 0.19 c | 6.93 ± 0.31 b | 12.8 ± 0.35 a | *** |
| quercetin-3-O-glucoside | 88.9 ± 5.38 b | 118 ± 2.32 a | 79.8 ± 5.60 b | ** | 4.08 ± 0.03 b | 4.98 ± 0.16 a | 1.61 ± 0.03 c | *** | 39.2 ± 1.02 a | 16.0 ± 0.89 b | 6.83 ± 0.60 c | *** | 0.54 ± 0.00 c | 1.26 ± 0.01 b | 3.30 ± 0.03 a | *** |
| feruloyl quinic acid | 613 ± 11.7 c | 734 ± 34.2 b | 1123 ± 43.3 a | *** | 2.78 ± 0.04 c | 39.6 ± 0.80 a | 9.67 ± 0.17 b | *** | 13.4 ± 1.02 a | 5.99 ± 0.17 b | 5.55 ± 0.15 b | *** | 4.65 ± 0.20 b | 8.48 ± 0.29 a | 4.99 ± 0.40 b | *** |
| rosmarinic acid | nd | nd | nd | na | 10969 ± 765 b | 13025 ± 128 a | 11246 ± 71 b | * | 11532 ± 428 a | 9236 ± 246 b | 9081 ± 176 b | ** | nd | nd | nd | na |
| cirsiliol | nd | nd | nd | na | 44.9 ± 1.44 c | 116 ± 2.17 a | 67.3 ± 0.85 b | *** | 31.9 ± 1.53 a | 18.9 ± 0.49 b | 20.1 ± 0.64 b | *** | nd | nd | nd | na |
| ferulic acid | 32.7 ± 0.97 b | 36.4 ± 0.94 b | 55.8 ± 2.19 a | *** | 25.5 ± 0.54 b | 50.3 ± 2.34 a | 21.6 ± 0.76 b | *** | 36.0 ± 1.29 a | 24.9 ± 0.66 b | 22.0 ± 0.24 b | *** | 0.47 ± 0.01 c | 0.84 ± 0.06 b | 1.02 ± 0.01 a | *** |
| luteolin-3-O-rutinoside | 248 ± 5.90 c | 532 ± 15.5 a | 374 ± 14.6 b | *** | 4.56 ± 0.03 a | 3.99 ± 0.13 b | 2.52 ± 0.06 c | *** | 15.1 ± 0.65 a | 10.4 ± 0.32 b | 10.2 ± 0.59 b | *** | nd | nd | nd | na |
| kaempferol-7-O-glucoside | 3.06 ± 0.13 a | 2.23 ± 0.02 c | 2.65 ± 0.05 b | *** | 2.50 ± 0.13 a | 1.79 ± 0.03 b | 1.74 ± 0.09 b | ** | 13.4 ± 1.15 a | 9.52 ± 0.48 b | 9.23 ± 0.25 b | * | 3.30 ± 0.09 c | 4.02 ± 0.28 b | 8.38 ± 0.15 a | *** |
| dicaffeoylquinic acid | 10.7 ± 1.28 b | 15.6 ± 0.42 a | 9.60 ± 0.35 b | ** | 1.16 ± 0.04 b | 3.09 ± 0.10 a | 2.83 ± 0.13 a | *** | 0.95 ± 0.03 a | 0.67 ± 0.04 b | 0.42 ± 0.04 c | *** | nd | nd | nd | na |
| kaempferol-3-O-rutinoside | 270 ± 9.55 c | 313 ± 4.63 b | 408 ± 9.77 a | *** | 4.48 ± 0.03 b | 3.23 ± 0.14 c | 5.13 ± 0.18 a | *** | 15.1 ± 0.66 a | 10.3 ± 0.34 b | 10.1 ± 0.60 b | *** | 0.10 ± 0.01 c | 0.22 ± 0.01 b | 0.28 ± 0.02 a | *** |
| quercetin-rhamnoside | 0.19 ± 0.02 a | 0.16 ± 0.02 ab | 0.11 ± 0.00 b | * | 52.1 ± 2.81 | 96.7 ± 3.79 | 148 ± 112 | ns | 102 ± 9.49 a | 68.9 ± 1.16 b | 65.0 ± 3.54 b | ** | 0.15 ± 0.01 c | 0.25 ± 0.02 b | 0.33 ± 0.01 a | *** |
| luteolin-malonil-hexose | 0.96 ± 0.08 | 1.04 ± 0.11 | 1.05 ± 0.10 | ns | nd | nd | nd | na | nd | nd | nd | na | 0.83 ± 0.10 b | 0.91 ± 0.05 b | 1.15 ± 0.03 a | * |
| apigenin-7-rhamnoside-4-rutinoside | nd | nd | nd | na | nd | nd | nd | na | nd | nd | nd | na | 7.12 ± 0.36 c | 16.1 ± 0.15 a | 13.2 ± 0.17 b | *** |
| apigenin-7-O-glucoside | 0.05 ± 0.01 | 0.06 ± 0.01 | 0.05 ± 0.00 | ns | 31.6 ± 0.27 b | 46.9 ± 2.36 a | 21.2 ± 0.18 c | *** | 5.71 ± 0.43 a | 3.20 ± 0.12 b | 2.96 ± 0.11 b | *** | 0.10 ± 0.01 | 0.10 ± 0.01 | 0.08 ± 0.01 | ns |
| Total polyphenols | 9311 ± 385 b | 10154 ± 311 ab | 11246 ± 402 a | * | 12255 ± 818 b | 16154 ± 113 a | 12683 ± 199 b | ** | 12717 ± 489 a | 9999 ± 273 b | 9774 ± 201 b | ** | 441 ± 21 b | 481 ± 26 b | 858 ± 8 a | *** |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pannico, A.; El-Nakhel, C.; Graziani, G.; Kyriacou, M.C.; Giordano, M.; Soteriou, G.A.; Zarrelli, A.; Ritieni, A.; De Pascale, S.; Rouphael, Y. Selenium Biofortification Impacts the Nutritive Value, Polyphenolic Content, and Bioactive Constitution of Variable Microgreens Genotypes. Antioxidants 2020, 9, 272. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9040272
Pannico A, El-Nakhel C, Graziani G, Kyriacou MC, Giordano M, Soteriou GA, Zarrelli A, Ritieni A, De Pascale S, Rouphael Y. Selenium Biofortification Impacts the Nutritive Value, Polyphenolic Content, and Bioactive Constitution of Variable Microgreens Genotypes. Antioxidants. 2020; 9(4):272. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9040272
Chicago/Turabian StylePannico, Antonio, Christophe El-Nakhel, Giulia Graziani, Marios C. Kyriacou, Maria Giordano, Georgios A. Soteriou, Armando Zarrelli, Alberto Ritieni, Stefania De Pascale, and Youssef Rouphael. 2020. "Selenium Biofortification Impacts the Nutritive Value, Polyphenolic Content, and Bioactive Constitution of Variable Microgreens Genotypes" Antioxidants 9, no. 4: 272. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9040272