Identification of Chemical Profiles and Biological Properties of Rhizophora racemosa G. Mey. Extracts Obtained by Different Methods and Solvents



 , , , ,
, , , , 
 , , ,
, , , 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Plant Material and Preparation of Extracts
2.2. Profile of Bioactive Compounds
2.3. Determination of Antioxidant and Enzyme Inhibitory Effects
2.4. Antibacterial and Anti-fungal Activities
2.5. Bioinformatics and Docking Studies
2.6. Statistical Analysis
3. Results
3.1. Profiling of Bioactive Compounds
3.2. Antioxidant Activities
3.3. Enzymatic Inhibitory Effects
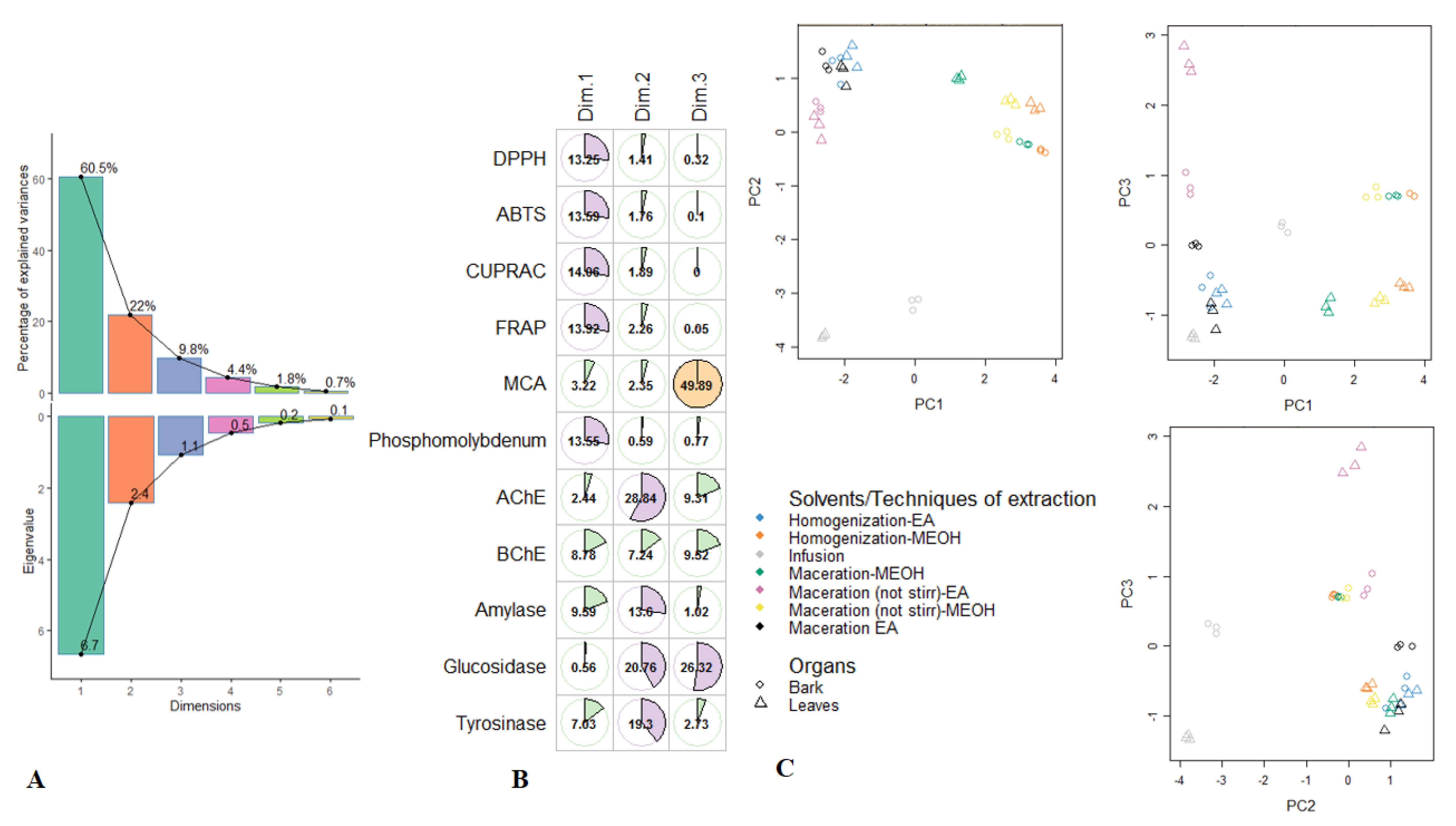
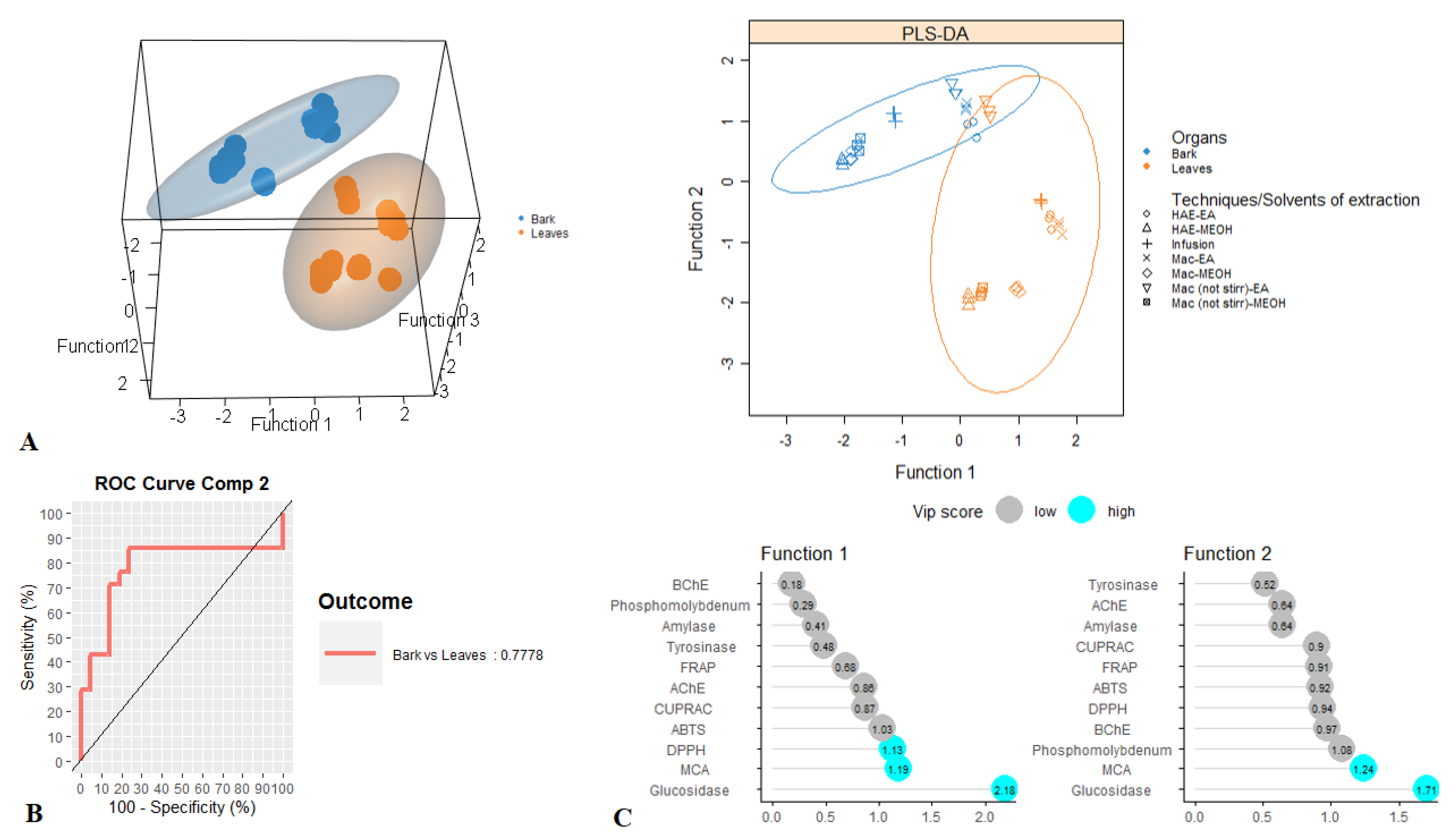
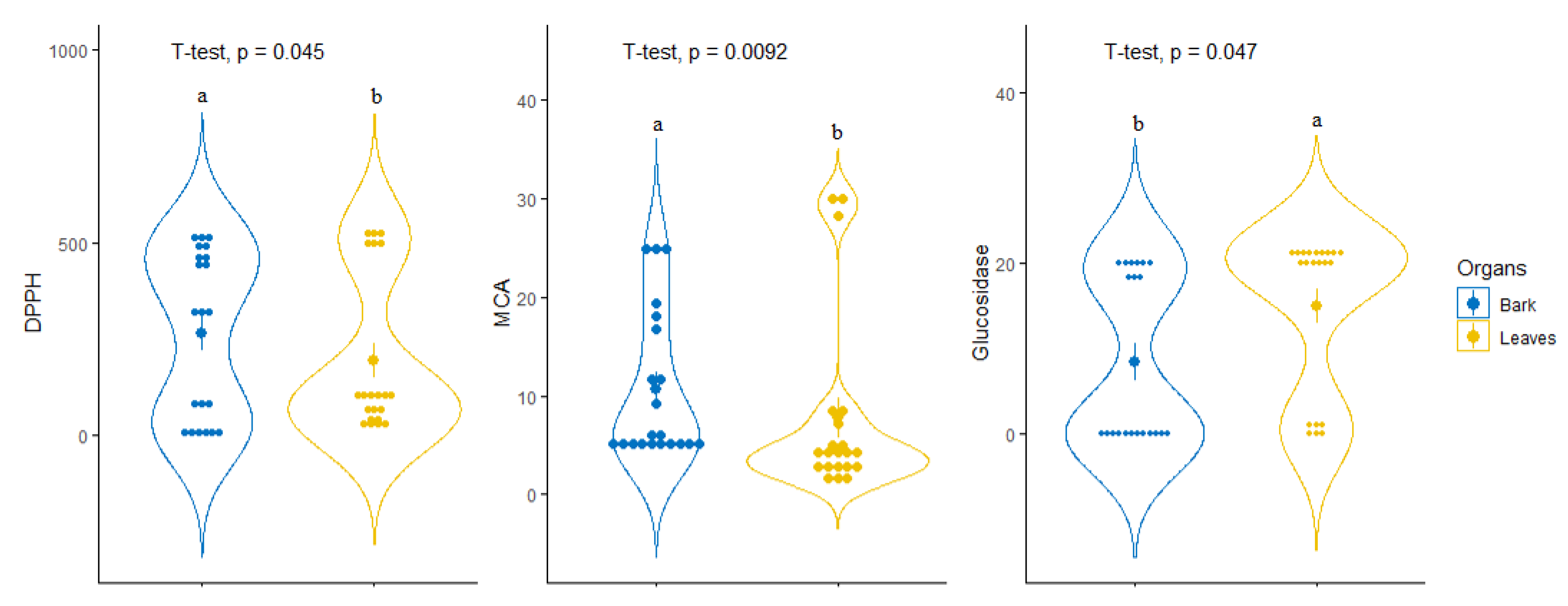
3.4. Exploratory Analysis, Classification and Discriminant Analysis
3.5. Anti-bacterial and Anti-fungal Activities
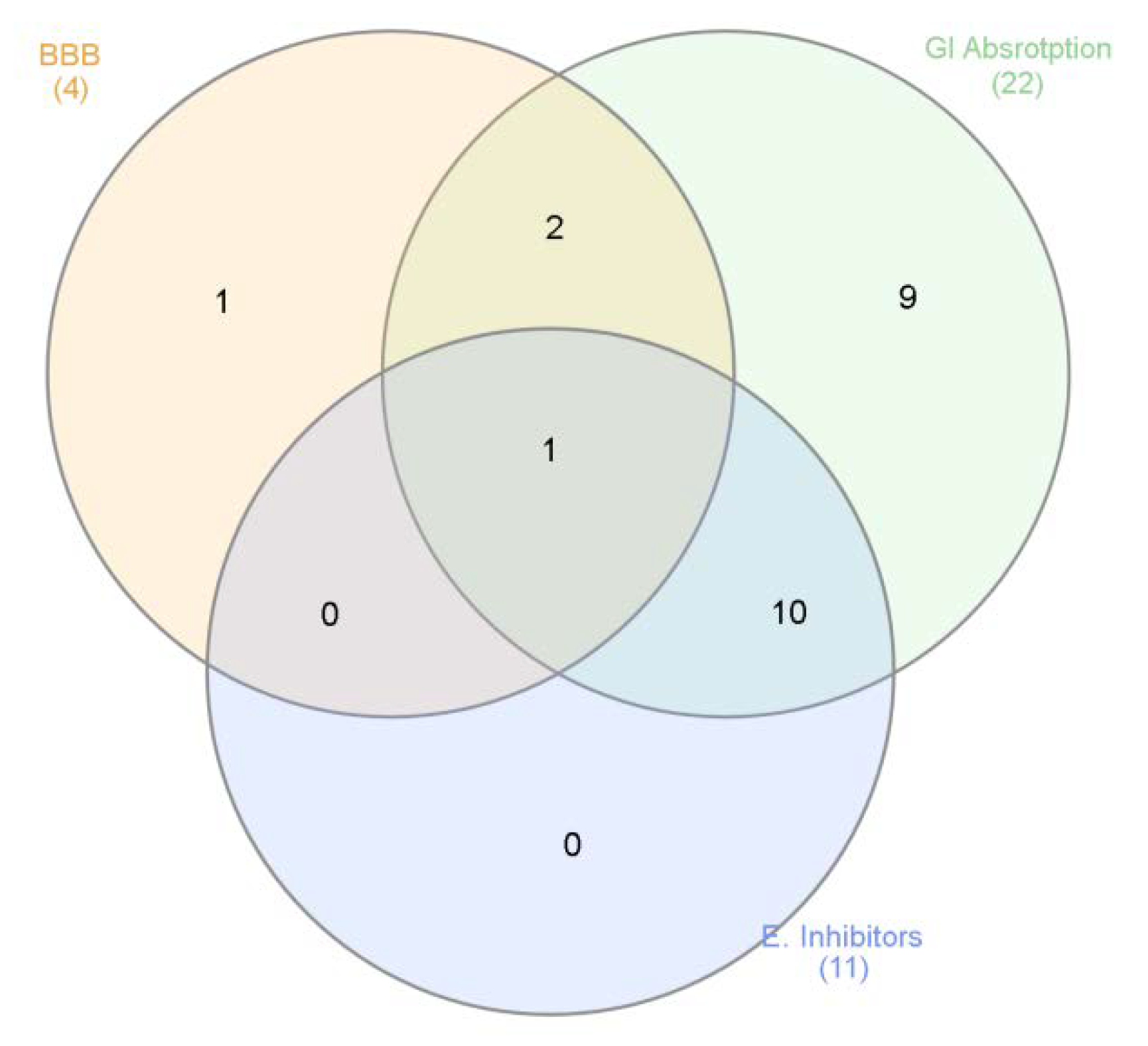
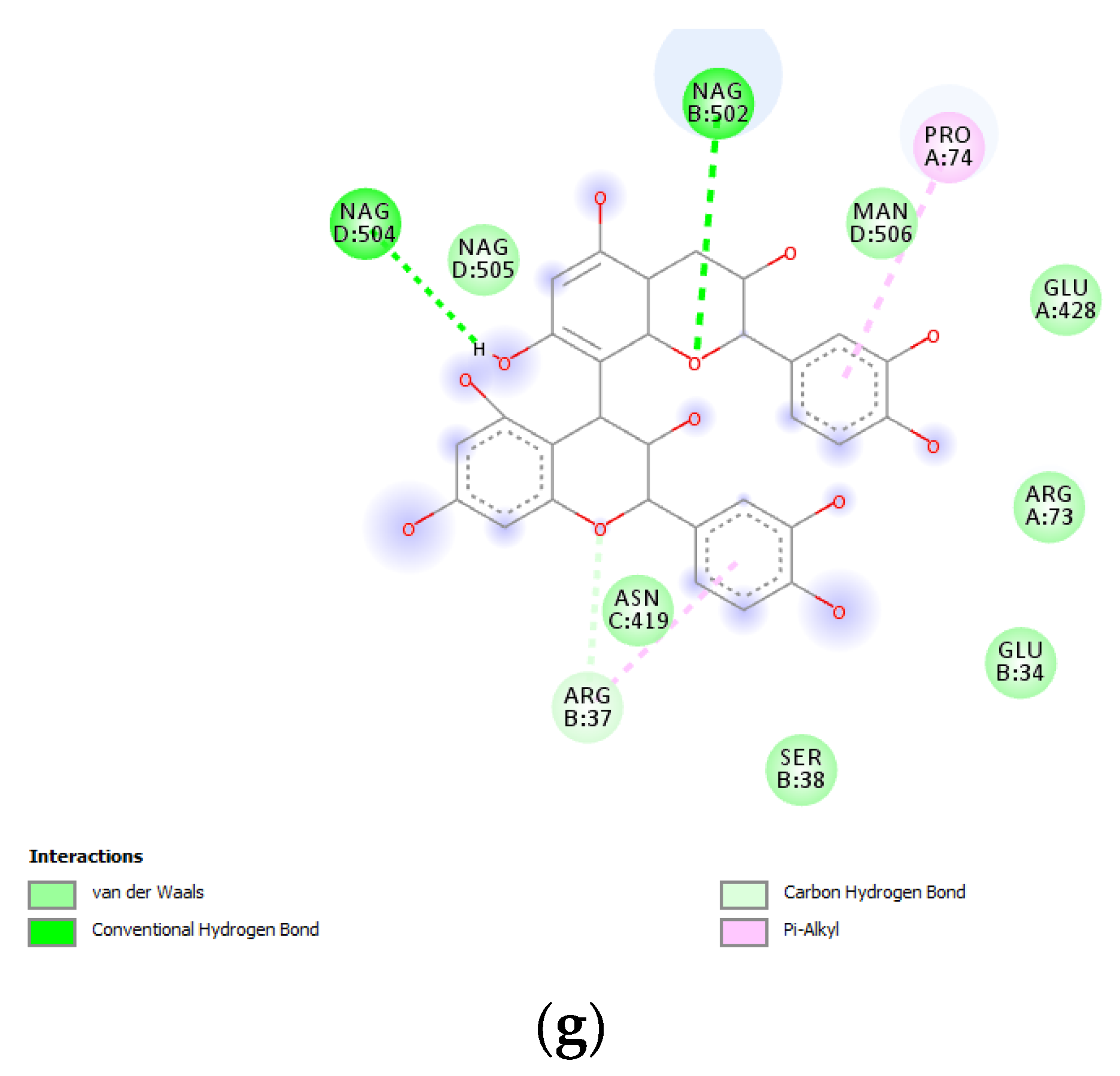
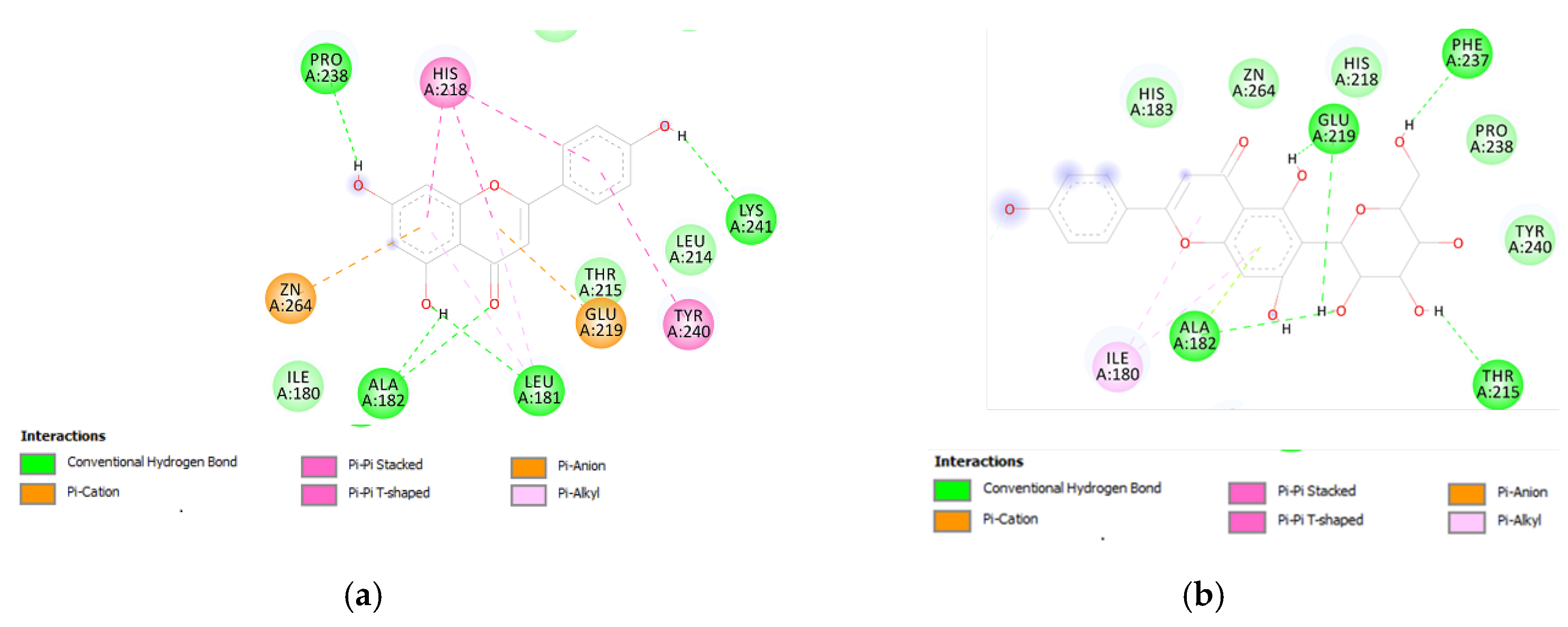
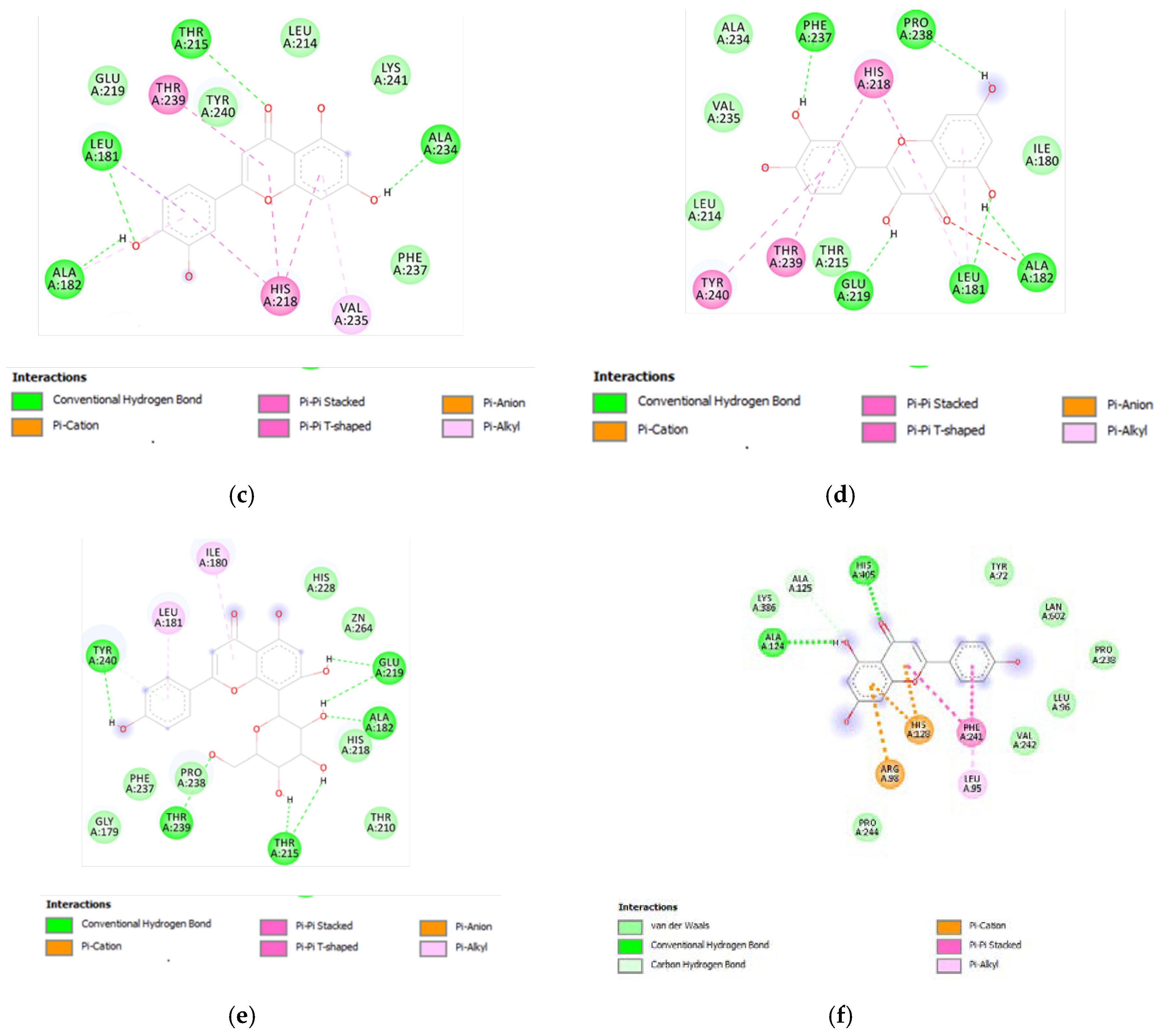
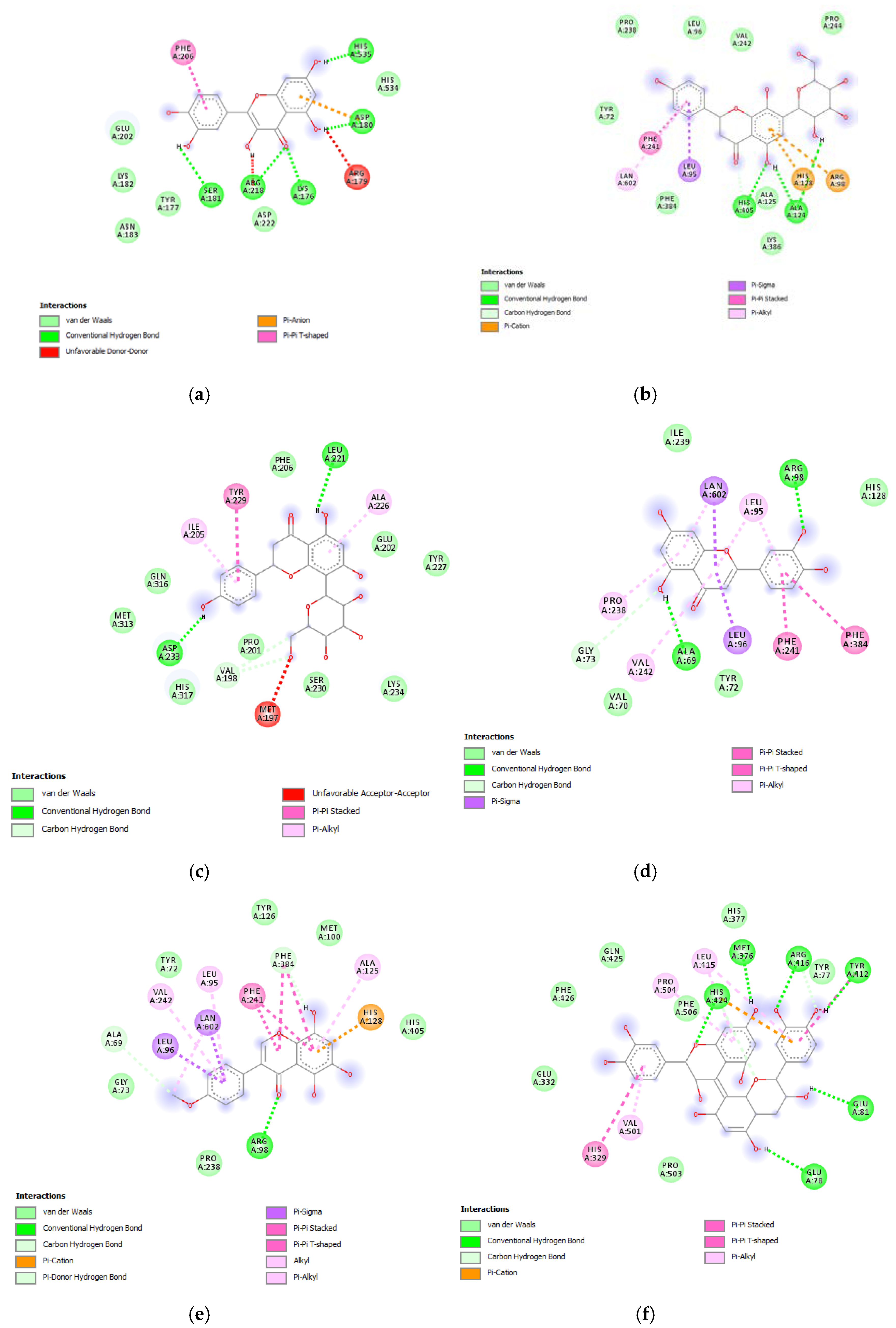
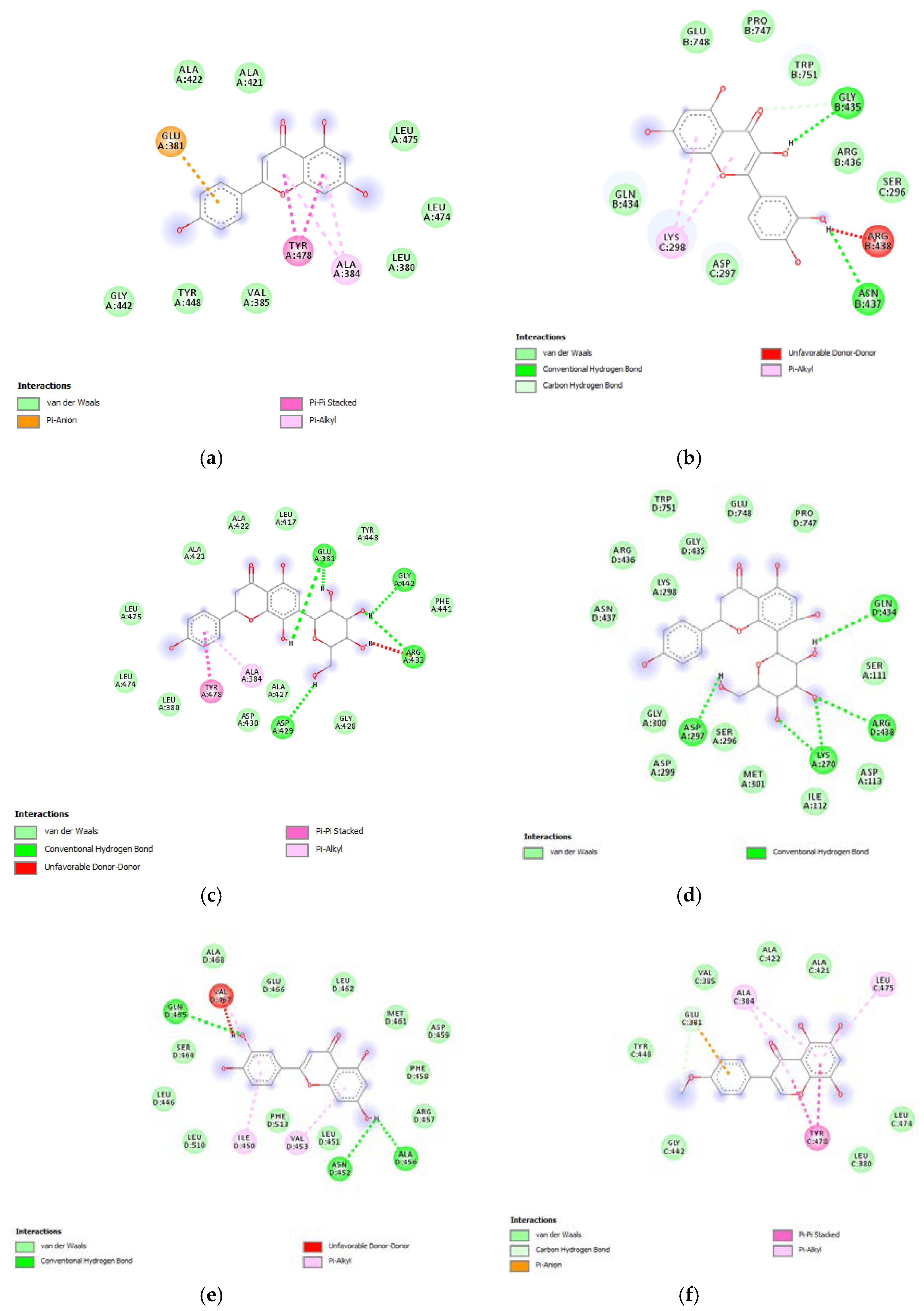
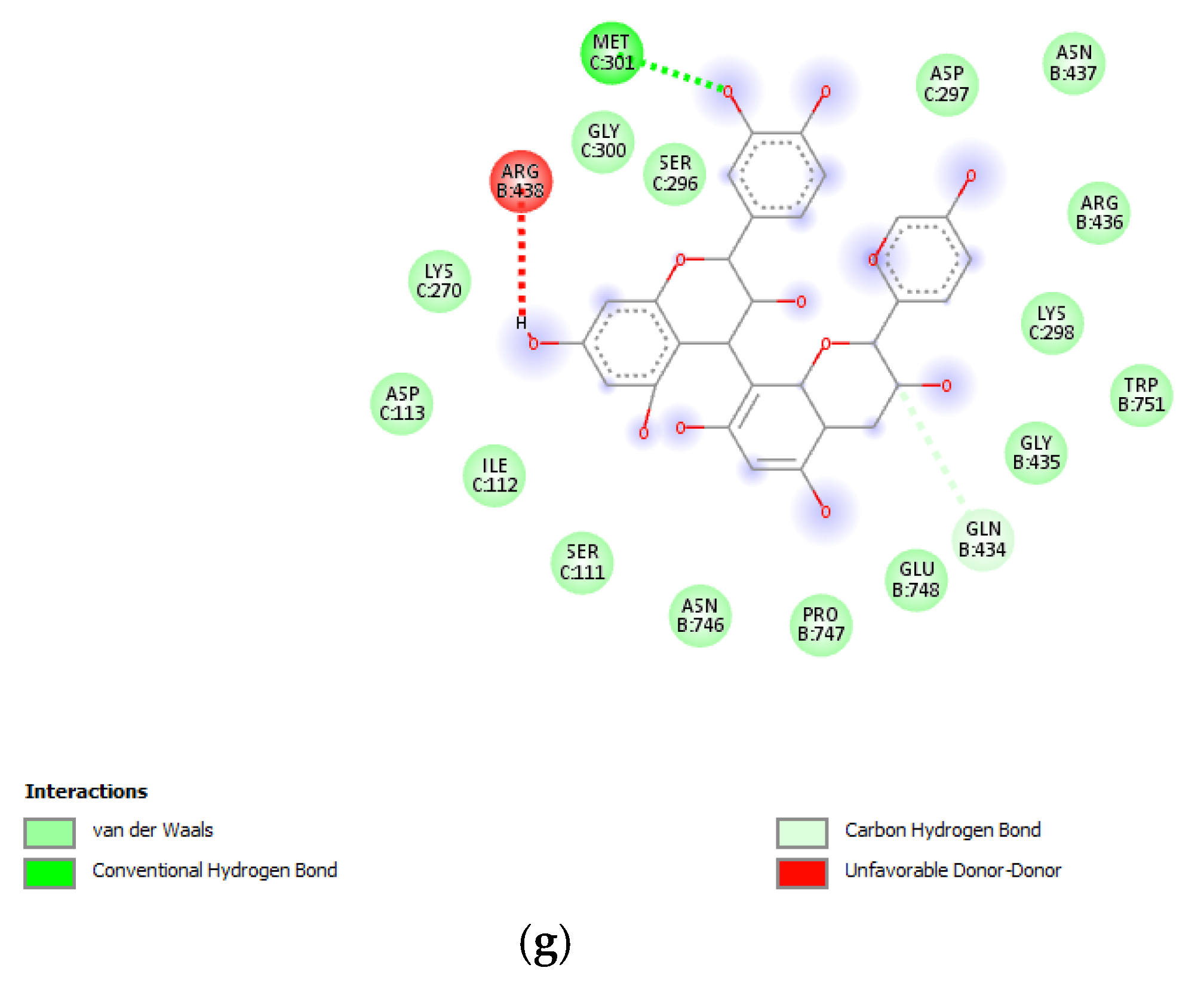
3.6. Bioinformatics and Docking Studies
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Gong, B.; Cao, H.; Peng, C.; Perčulija, V.; Tong, G.; Fang, H.; Wei, X.; Ouyang, S. High-throughput sequencing and analysis of microbial communities in the mangrove swamps along the coast of Beibu Gulf in Guangxi, China. Sci. Rep. 2019, 9, 9377. [Google Scholar] [CrossRef] [PubMed]
- Bandaranayake, W.M. Bioactivities, bioactive compounds and chemical constituents of mangrove plants. Wetl. Ecol. Manag. 2002, 10, 421–452. [Google Scholar] [CrossRef]
- Sur, T.K.; Hazra, A.; Hazra, A.K.; Bhattacharyya, D. Antioxidant and hepatoprotective properties of Indian Sunderban mangrove Bruguiera gymnorrhiza L. leaf. J. Basic Clin. Pharm. 2016, 7, 75–79. [Google Scholar] [CrossRef] [Green Version]
- Bibi, S.N.; Fawzi, M.M.; Gokhan, Z.; Rajesh, J.; Nadeem, N.; Rengasamy, K.R.R.; Albuquerque, R.D.D.G.; Pandian, S.K. Ethnopharmacology, Phytochemistry, and Global Distribution of Mangroves―A Comprehensive Review. Mar. Drugs 2019, 17, 231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomlinson, P.B. Family: Rhizophoraceae. In The Botany of Mangroves, 2nd ed.; Tomlinson, P.B., Ed.; Cambridge University Press: Cambridge, UK, 2016; pp. 315–355. [Google Scholar]
- Angalabiri-Owei, B.; Isirima, J. Evaluation of the lethal dose of the methanol extract of Rhizophora racemosa leaf using karbers method. Afr. J. Cell Pathol 2014, 2, 65–68. [Google Scholar] [CrossRef] [Green Version]
- Dossou-Yovo, H.; Fifanou, G.; Sinsin, B. Ethnobotanical survey of mangrove plant species used as medicine from ouidah to grand-popo districts, southern Benin. Am. J. Ethnomed 2017, 4, 1–6. [Google Scholar]
- Balasubramanian, V.; Rajesh, P.; Rajaram, R.; Kannan, V.R. A review on Rhizophora genus: Therapeutically important perspective phytochemical constituents. Bioactive Phytochemicals: Perspectives for Modern Medicine; Gupta, V.K., Ed.; Daya Publishing House: New Delhi, India, 2015; p. 3. [Google Scholar]
- Ukoima, H.; Ikata, M.; Pepple, G. Control of Lasiodiplodia theobromae (PAT) on Rhizophora racemosa using plants extracts. Am. J. Biotechnol. Mol. Sci. 2013, 3, 1–7. [Google Scholar] [CrossRef]
- Udeozo, I.; Okafor, G.; Ike, O.; Eze, E. The efficacy of Rhizophora racemosa wood: An important study. Fuw Trend. Sci. Technol. J. 2018, 3, 977–980. [Google Scholar]
- List. 2013. Available online: http://www.theplantlist.org/1.1/browse/A/Rhizophoraceae/Rhizophora/ (accessed on 14 April 2020).
- Chiavaroli, A.; Recinella, L.; Ferrante, C.; Locatelli, M.; Macchione, N.; Zengin, G.; Leporini, L.; Leone, S.; Martinotti, S.; Brunetti, L.; et al. Crocus sativus, Serenoa repens and Pinus massoniana extracts modulate inflammatory response in isolated rat prostate challenged with LPS. J. Biol. Regul. Homeost. Agents 2017, 31, 531–541. [Google Scholar]
- Ferrante, C.; Recinella, L.; Ronci, M.; Menghini, L.; Brunetti, L.; Chiavaroli, A.; Leone, S.; Di Iorio, L.; Carradori, S.; Tirillini, B.; et al. Multiple pharmacognostic characterization on hemp commercial cultivars: Focus on inflorescence water extract activity. Food Chem. Toxicol. 2019, 125, 452–461. [Google Scholar] [CrossRef]
- Zengin, G.; Locatelli, M.; Stefanucci, A.; Macedonio, G.; Novellino, E.; Mirzaie, S.; Dvorácskó, S.; Carradori, S.; Brunetti, L.; Orlando, G.; et al. Chemical characterization, antioxidant properties, anti-inflammatory activity, and enzyme inhibition of Ipomoea batatas L. leaf extracts. Int. J. Food Prop. 2017, 20, 1907–1919. [Google Scholar] [CrossRef] [Green Version]
- Mollica, A.; Zengin, Z.; Stefanucci, A.; Ferrante, C.; Menghini, L.; Orlando, G.; Brunetti, L.; Locatelli, M.; Dimmito, M.P.; Novellino, E.; et al. Nutraceutical potential of Corylus avellana daily supplements for obesity and related dysmetabolism. J. Funct. Foods 2018, 47, 562–574. [Google Scholar] [CrossRef]
- Zengin, G.; Aktumsek, A.; Ceylan, R.; Uysal, S.; Mocan, A.; Guler, G.O.; Mahomoodally, M.F.; Glamočlija, J.; Ćirić, A.; Soković, M. Shedding light on the biological and chemical fingerprints of three Achillea species (A. biebersteinii, A. millefolium and A. teretifolia). Food Funct. 2017, 8, 1152–1165. [Google Scholar] [CrossRef]
- Garde-Cerdán, T.; Gonzalo-Diago, A.; Pérez-Álvarez, E.P. Phenolic Compounds: Types, Effects and Research; Nova Science Publishers, Incorporated: Hauppauge, NY, USA, 2017. [Google Scholar]
- Huang, W.-Y.; Cai, Y.-Z.; Zhang, Y. Natural Phenolic Compounds From Medicinal Herbs and Dietary Plants: Potential Use for Cancer Prevention. Nutr. Cancer 2009, 62, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Baba, S.A.; Malik, S.A. Determination of total phenolic and flavonoid content, antimicrobial and antioxidant activity of a root extract of Arisaema jacquemontii Blume. J. Taibah Univ. Sci. 2015, 9, 449–454. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Pandey, A.K. Chemistry and biological activities of flavonoids: An overview. Sci. World J. 2013, 2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, G.A.; Molina, G.; Arruda, H.S.; Pastore, G.M. Optimizing the Homogenizer-Assisted Extraction (HAE) of Total Phenolic Compounds from Banana Peel. J. Food Process Eng. 2017, 40, e12438. [Google Scholar] [CrossRef]
- Santos, J.S.; Alvarenga Brizola, V.R.; Granato, D. High-throughput assay comparison and standardization for metal chelating capacity screening: A proposal and application. Food Chem. 2017, 214, 515–522. [Google Scholar] [CrossRef]
- Zengin, G.; Mahomoodally, M.; Picot-Allain, C.; Cakmak, Y.; Uysal, S.; Aktumsek, A. In vitro tyrosinase inhibitory and antioxidant potential of Consolida orientalis, Onosma isauricum and Spartium junceum from Turkey. S. Afr. J. Bot. 2019, 120, 119–123. [Google Scholar] [CrossRef]
- Targher, G.; Lonardo, A.; Byrne, C.D. Nonalcoholic fatty liver disease and chronic vascular complications of diabetes mellitus. Nat. Rev. Endocrinol. 2018, 14, 99. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Zhu, S.; Yang, Y.; Ma, X.; Guo, S. Matrix metalloproteinase-12 expression is increased in cutaneous melanoma and associated with tumor aggressiveness. Tumor Biol. 2015, 36, 8593–8600. [Google Scholar] [CrossRef]
- Becker, R.; Bubeck, J.W. Staphylococcus aureus and the skin: A longstanding and complex interaction. Skinmed 2015, 13, 111–119. [Google Scholar] [PubMed]
- Babamahmoodi, F.; Shokohi, T.; Ahangarkani, F.; Nabili, M.; Afzalian Ashkezari, E.; Alinezhad, S. Rare case of Aspergillus ochraceus osteomyelitis of calcaneus bone in a patient with diabetic foot ulcers. Case Rep. Med. 2015, 2015, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Bongomin, F.; Batac, C.; Richardson, M.D.; Denning, D.W. A review of onychomycosis due to Aspergillus species. Mycopathologia 2018, 183, 485–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.; Li, D.; Shang, P.; Sun, T.; Xiao, X. Preliminary analyses on bacterial diversity and resistance in infection-related skin disorders. Zhonghua Yi Xue Za Zhi 2015, 95, 1762–1764. [Google Scholar] [PubMed]
- Lambotte, O.; Fihman, V.; Poyart, C.; Buzyn, A.; Berche, P.; Soumelis, V. Listeria monocytogenes skin infection with cerebritis and haemophagocytosis syndrome in a bone marrow transplant recipient. J. Infect. 2005, 50, 356–358. [Google Scholar] [CrossRef] [PubMed]
- Maravić, A.; Rončević, T.; Krce, L.; Ilić, N.; Galić, B.; Čulić Čikeš, V.; Carev, I. Halogenated boroxine dipotassium trioxohydroxytetrafluorotriborate K2 [B3O3F4OH] inhibits emerging multidrug-resistant and β-lactamase-producing opportunistic pathogens. Drug Dev. Ind. Pharm. 2019, 45, 1770–1776. [Google Scholar] [CrossRef]
- Ranjan, A.; Shaik, S.; Nandanwar, N.; Hussain, A.; Tiwari, S.K.; Semmler, T.; Jadhav, S.; Wieler, L.H.; Alam, M.; Colwell, R.R. Comparative genomics of Escherichia coli isolated from skin and soft tissue and other extraintestinal infections. MBio 2017, 8, e01070-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saikia, L.; Gogoi, N.; Das, P.P.; Sarmah, A.; Punam, K.; Mahanta, B.; Bora, S.; Bora, R. Bacillus cereus–Attributable Primary Cutaneous Anthrax-Like Infection in Newborn Infants, India. Emerging Infect. Dis. 2019, 25, 1261. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Liu, X.; Zhang, J.; Cai, L.; Zhou, C. Drug-induced toxic epidermal necrolysis with secondary aspergillus fumigatus infection: A case report. J. Peking Univ. Health Sci. 2019, 51, 977. [Google Scholar]
- Houst, J.; Spizek, J.; Havlicek, V. Antifungal Drugs. Metabolites 2020, 10, 106. [Google Scholar] [CrossRef] [Green Version]
- Khan, T.; Sankhe, K.; Suvarna, V.; Sherje, A.; Patel, K.; Dravyakar, B. DNA gyrase inhibitors: Progress and synthesis of potent compounds as antibacterial agents. Biomed. Pharmacother. 2018, 103, 923–938. [Google Scholar] [CrossRef]
- Smirnoff, N. Antioxidants and Reactive Oxygen Species in Plants; John Wiley & Sons: Hoboken, NJ, USA, 2008. [Google Scholar]
- Davalli, P.; Mitic, T.; Caporali, A.; Lauriola, A.; D’Arca, D. ROS, cell senescence, and novel molecular mechanisms in aging and age-related diseases. Oxidat. Med. Cell. Longev. 2016, 2016, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandrasekaran, A.; Idelchik, M.D.P.S.; Melendez, J.A. Redox control of senescence and age-related disease. Redox Biol. 2017, 11, 91–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segura Campos, M.R.; Ruiz Ruiz, J.; Chel-Guerrero, L.; Betancur Ancona, D. Coccoloba uvifera (L.)(Polygonaceae) fruit: Phytochemical screening and potential antioxidant activity. J. Chem. 2015, 2015, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Litalien, C.; Beaulieu, P. Chapter 117-Molecular Mechanisms of Drug Actions: From Receptors to Effectors. In Pediatric Critical Care, 4th ed.; Fuhrman, B.P., Zimmerman, J.J., Eds.; Mosby: Saint Louis, MO, USA, 2011; pp. 1553–1568. [Google Scholar]
- Rauf, A.; Jehan, N. Natural products as a potential enzyme inhibitors from medicinal plants. In Enzyme Inhibitors and Activators; InTech: Rijeka, Croatia, 2017; pp. 165–177. [Google Scholar]
- Nascimento, D.K.; Souza, I.A.; Oliveira, A.F.D.; Barbosa, M.O.; Santana, M.A.; Pereira, D.F., Jr.; Lira, E.C.; Vieira, J.R. Phytochemical screening and acute toxicity of aqueous extract of leaves of Conocarpus erectus Linnaeus in swiss albino mice. Anais da Academia Brasileira de Ciências 2016, 88, 1431–1437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mannina, L.; Segre, A.L.; Ritieni, A.; Fogliano, V.; Vinale, F.; Randazzo, G.; Maddau, L.; Bottalico, A. A new fungal growth inhibitor from Trichoderma viride. Tetrahedron 1997, 53, 3135–3144. [Google Scholar] [CrossRef]
- Meng, D.; Garba, B.; Ren, Y.; Yao, M.; Xia, X.; Li, M.; Wang, Y. Antifungal activity of chitosan against Aspergillus ochraceus and its possible mechanisms of action. Int. J. Biol. Macromol. 2020. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parts | Methods-Solvents | TPC (mg GAE/g) | TFC (mg RE/g) | TPAC (mg CAE/g) | TFlv (mg CE/g) |
|---|---|---|---|---|---|
| Leaves | infusion | 83.85 ± 1.12 f | 17.22 ± 0.30 e | 26.40 ± 0.77 d | 1.66 ± 0.01 ij |
| HAE-EA | 50.17 ± 0.57 g | 24.06 ± 2.09 cd | nd | 8.28 ± 0.03 f | |
| HAE-MEOH | 217.21 ± 0.69 a | 33.10 ± 1.78 b | 58.50 ± 2.03 a | 62.43 ± 0.41 b | |
| Maceration EA | 35.44 ±0.32 h | 25.57 ± 0.75 c | nd | 2.92 ± 0.06 hi | |
| Maceration-MEOH | 198.57 ± 5.51 c | 38.45 ± 0.75 a | 49.69 ± 2.35 b | 60.42 ± 0.36 c | |
| Maceration (not stirred)-EA | 24.27 ± 0.40 j | 22.20 ± 0.35 d | nd | 1.46 ± 0.02 ij | |
| Maceration (not stirred)-MEOH | 191.74 ± 0.64 d | 38.07 ± 0.60 a | 40.39 ± 7.11 c | 58.11 ± 0.98 d | |
| Bark | infusion | 145.15 ± 0.39 e | 15.01 ± 1.76 ef | 61.50 ± 1.65 a | 5.27 ± 0.08 g |
| HAE-EA | 30.34 ± 0.33 i | 7.96 ± 0.10 i | nd | 3.81 ± 0.02 gh | |
| HAE-MEOH | 210.00 ± 1.28 b | 9.00 ± 0.52 hi | 47.73 ± 0.93 b | 66.17 ± 1.40 a | |
| Maceration EA | 30.84 ± 0.18 hi | 8.81 ± 0.20 hi | nd | 2.29 ± 0.06 hij | |
| Maceration-MEOH | 201.73 ± 0.43 c | 13.64 ± 0.79 fg | 34.04 ± 0.41 c | 63.21 ± 1.20 b | |
| Maceration (not stirred)-EA | 28.51 ± 0.26 ij | 10.96 ± 0.45 gh | nd | 0.73 ± 0.01 j | |
| Maceration (not stirred)-MEOH | 192.92 ± 0.74 d | 12.52 ± 0.15 fg | 26.70 ± 2.74 d | 11.59 ± 0.09 e |
| No. | Name | Formula | Leaves Infusion | Leaves HAE-EA | Leaves HAE-MeOH | Bark Infusion | Bark HAE-EA | Bark HAE-MeOH |
|---|---|---|---|---|---|---|---|---|
| 1 | Gallic acid (3,4,5-Trihydroxybenzoic acid) | C7H6O5 | + | − | − | + | − | + |
| 2 | Protocatechuic acid (3,4-Dihydroxybenzoic acid) | C7H6O4 | + | + | + | + | + | + |
| 3 | Pantothenic acid | C9H17NO5 | − | − | − | + | − | + |
| 4 | Neochlorogenic acid (5-O-Caffeoylquinic acid) | C16H18O9 | + | + | + | + | + | + |
| 5 | Prodelphinidin C | C30H26O13 | − | − | − | − | − | + |
| 6 | Hydroxybenzaldehyde | C7H6O2 | + | + | + | − | − | − |
| 7 | 3-O-(4-Coumaroyl)quinic acid | C16H18O8 | + | + | + | + | + | + |
| 8 1 | Catechin | C15H14O6 | + | − | + | + | + | + |
| 9 | Esculetin (6,7-Dihydroxycoumarin) | C9H6O4 | + | + | + | + | + | + |
| 10 1 | Chlorogenic acid (3-O-Caffeoylquinic acid) | C16H18O9 | + | + | + | + | + | + |
| 11 | 3-O-Feruloylquinic acid | C17H20O9 | + | − | + | + | − | + |
| 12 | Caffeic acid | C9H8O4 | + | + | + | + | + | + |
| 13 | Procyanidin B | C30H26O12 | + | + | + | + | + | + |
| 14 | Cinchonain II isomer 1 | C39H32O15 | + | − | + | + | − | + |
| 15 | Chryptochlorogenic acid (4-O-Caffeoylquinic acid) | C16H18O9 | + | + | + | + | + | + |
| 16 | Cinchonain II isomer 2 | C39H32O15 | + | − | + | + | − | + |
| 17 | 2-Oxindole | C8H7NO | + | + | + | + | + | + |
| 18 | Procyanidin C | C45H38O18 | + | + | + | + | + | + |
| 19 | 5-O-(4-Coumaroyl)quinic acid | C16H18O8 | + | + | + | + | − | + |
| 20 1 | Epicatechin | C15H14O6 | + | + | + | + | + | + |
| 21 | Cinchonain II isomer 3 | C39H32O15 | + | − | + | − | − | − |
| 22 | 4-O-(4-Coumaroyl)quinic acid | C16H18O8 | + | − | + | + | − | + |
| 23 | Cinchonain II isomer 4 | C39H32O15 | − | − | + | − | − | − |
| 24 | Antiarol (3,4,5-Trimethoxyphenol) | C9H12O4 | + | + | + | + | + | + |
| 25 | 5-O-Feruloylquinic acid | C17H20O9 | + | − | + | + | − | − |
| 26 1 | 4-Coumaric acid | C9H8O3 | + | + | + | + | + | + |
| 27 | Tuberonic acid or 12-Hydroxyjasmonic acid | C12H18O4 | + | + | + | + | + | + |
| 28 | 4-O-Feruloylquinic acid | C17H20O9 | + | − | + | + | - | + |
| 29 | Riboflavin | C17H20N4O6 | + | − | + | + | − | + |
| 30 | Cinchonain I isomer 1 | C24H20O9 | − | − | + | − | + | + |
| 31 | Naringenin-C-hexoside isomer 1 | C21H22O10 | + | − | + | + | − | + |
| 32 1 | Taxifolin (Dihydroquercetin) | C15H12O7 | + | + | + | + | + | + |
| 33 | Quercetin-O-rhamnosyldihexoside | C33H40O21 | + | + | + | + | + | + |
| 34 | Naringenin-C-hexoside isomer 2 | C21H22O10 | + | − | + | + | − | + |
| 35 | Quercetin-O-hexosylhexoside | C27H30O17 | + | + | + | + | + | + |
| 36 | Naringenin-C-hexoside isomer 3 | C21H22O10 | + | − | + | + | − | + |
| 37 | Quercetin-O-dirhamnosylhexoside | C33H40O20 | + | + | + | + | − | + |
| 38 | Hexahydroxy(iso)flavone-O-hexoside | C21H20O13 | + | − | + | + | − | + |
| 39 | Hexahydroxy(iso)flavone-O-rhamnosylhexoside | C27H30O17 | + | + | + | + | + | + |
| 40 | Cinchonain I isomer 2 | C24H20O9 | − | − | + | − | + | + |
| 41 | Cinchonain I isomer 3 | C24H20O9 | − | − | + | − | + | + |
| 42 1 | Vitexin (Apigenin-8-C-glucoside) | C21H20O10 | + | + | + | + | + | + |
| 43 | Kaempferol-O-dirhamnosylhexoside | C33H40O19 | + | + | + | − | − | − |
| 44 | Isovitexin (Apigenin-6-C-glucoside) | C21H20O10 | + | + | + | + | + | + |
| 45 1 | Naringin (Naringenin-7-O-neohesperidoside) | C27H32O14 | + | − | + | + | + | + |
| 46 1 | Isoquercitrin (Quercetin-3-O-glucoside) | C21H20O12 | − | − | + | + | + | + |
| 47 1 | Rutin (Quercetin-3-O-rutinoside) | C27H30O16 | + | + | + | + | + | + |
| 48 | Quercetin-O-pentoside | C20H18O11 | + | + | + | − | − | − |
| 49 | Astragalin (Kaempferol-3-O-glucoside) | C21H20O11 | + | + | + | − | − | − |
| 50 | Kaempferol-3-O-rutinoside (Nicotiflorin) | C27H30O15 | + | + | + | + | + | + |
| 51 | Cinchonain I isomer 4 | C24H20O9 | − | − | + | − | + | + |
| 52 1 | Eriodictyol (3′,4′,5,7-Tetrahydroxyflavanone) | C15H12O6 | + | + | + | + | + | + |
| 53 1 | Isorhamnetin-3-O-glucoside | C22H22O12 | − | − | + | + | − | − |
| 54 | Tetrahydroxy(iso)flavone-O-rhamnosylhexoside | C28H32O16 | + | + | + | + | + | + |
| 55 | Abscisic acid | C15H20O4 | + | + | + | + | + | + |
| 56 1 | Quercetin (3,3’,4’,5,7-Pentahydroxyflavone) | C15H10O7 | + | + | + | + | + | + |
| 57 1 | Naringenin (4’,5,7-Trihydroxyflavanone) | C15H12O5 | + | + | + | + | + | + |
| 58 1 | Luteolin (3’,4’,5,7-Tetrahydroxyflavone) | C15H10O6 | + | + | + | + | + | + |
| 59 1 | Kaempferol (3,4’,5,7-Tetrahydroxyflavone) | C15H10O6 | + | + | + | − | − | − |
| 60 1 | Apigenin (4’,5,7-Trihydroxyflavone) | C15H10O5 | − | + | + | + | + | + |
| 61 1 | Isorhamnetin (3’-Methoxy-3,4’,5,7-tetrahydroxyflavone) | C16H12O7 | − | + | + | − | + | − |
| 62 | Methoxy-trihydroxy(iso)flavone | C16H12O6 | − | + | + | − | + | + |
| 63 | Pinocembrin (5,7-Dihydroxyflavanone) | C15H12O4 | − | + | + | − | − | − |
| 64 | Tetrahydroxyxanthone | C13H8O6 | − | − | − | + | − | − |
| 65 | Tetrahydroxyxanthone isomer 1 | C13H8O6 | − | − | − | − | − | + |
| 66 | Tetrahydroxyxanthone isomer 2 | C13H8O6 | − | − | − | − | − | + |
| 67 | Luteolin-7-O-glucoside (Cynaroside) | C21H20O11 | − | − | − | + | − | + |
| 68 | Luteolin-7-O-rutinoside (Scolymoside) | C27H30O15 | − | − | − | + | − | + |
| 69 | Methoxy-tetrahydroxy(iso)flavone | C15H10O5 | − | − | − | + | − | − |
| 70 | Methoxy-trihydroxy(iso)flavone isomer 1 | C16H12O6 | − | − | − | + | − | − |
| 71 | Methoxy-trihydroxy(iso)flavone isomer 2 | C16H12O6 | − | − | − | + | − | − |
| Parts | Methods-Solvents | DPPH (mg TE/g) | ABTS (mg TE/g) | CUPRAC (mg TE/g) | FRAP (mg TE/g) | Metal chelating (mg EDTAE/g) | Phosphomolybdenum (mmol TE/g) |
|---|---|---|---|---|---|---|---|
| Leaves | infusion | 101.26 ± 0.24 fg | 144.25 ± 3.10 e | 315.43 ± 5.58 g | 200.02 ± 4.73 f | 8.08 ± 0.76 e | 2.15 ± 0.11 d |
| HAE-EA | 67.69 ± 5.49 h | 89.09 ± 1.99 f | 193.20 ± 3.83 h | 77.29 ± 1.16 g | 1.95 ± 0.49 h | 2.12 ± 0.29 d | |
| HAE-MEOH | 525.84 ± 1.37 a | 600.84 ± 13.60 a | 1047.10 ± 9.45 b | 544.76 ± 11.89 b | 4.50 ± 0.53 fg | 4.82 ± 0.42 a | |
| Maceration EA | 20.97 ± 0.28 ij | 32.41 ± 6.38 g | 124.15 ± 2.58 i | 50.25 ± 1.91 hi | 2.32 ± 0.50 gh | 2.17 ± 0.08 d | |
| Maceration-MEOH | 105.56 ± 0.06 f | 159.42 ± 0.12 e | 753.22 ± 4.81 f | 401.16 ± 5.03 e | 4.31 ± 0.95 fg | 4.29 ± 0.20 abc | |
| Maceration (not stirred)-EA | 39.06 ± 2.14 i | 12.31 ± 0.29 hi | 84.92 ± 0.74 j | 40.54 ± 0.63 hi | 29.39 ± 1.03 a | 2.40 ± 0.13 d | |
| Maceration (not stirred)-MEOH | 496.94 ± 3.30 bc | 519.71 ± 6.78 c | 825.71 ± 25.99 e | 519.69 ± 2.96 c | 3.64 ± 0.31 fgh | 3.95 ± 0.44 bc | |
| Bark | infusion | 318.53 ± 8.82 e | 353.91 ± 11.81 d | 766.83 ± 7.23 f | 398.10 ± 6.46 e | 18.06 ± 1.29 c | 3.63 ± 0.14 c |
| HAE-EA | 81.08 ± 5.23 gh | 33.45 ± 1.66 g | 94.03 ± 2.12 j | 49.72 ± 1.12 hi | 5.39 ± 0.81 f | 1.69 ± 0.21 d | |
| HAE-MEOH | 512.37 ± 7.15 ab | 581.39 ± 6.42 a | 1129.33 ± 8.62 a | 633.53 ± 8.97 a | 5.42 ± 0.06 f | 4.74 ± 0.17 ab | |
| Maceration EA | 15.86 ± 1.83 ij | 29.36 ± 1.37 gh | 126.72 ± 4.57 j | 55.45 ± 0.32 h | 10.88 ± 1.41 d | 1.68 ± 0.15 d | |
| Maceration-MEOH | 478.39 ± 21.72 c | 555.13 ± 8.47 b | 931.13 ± 18.04 d | 559.27 ± 2.63 b | 5.61 ± 0.16 f | 4.51 ± 0.36 ab | |
| Maceration (not stirred)-EA | na | 7.26 ± 0.10 i | 74.82 ± 0.34 j | 37.67 ± 1.60 i | 24.93 ± 0.24 b | 2.13 ± 0.17 d | |
| Maceration (not stirred)-MEOH | 450.51 ± 15.42 d | 513.29 ± 8.26 c | 961.26 ± 1.49 c | 422.30 ± 12.50 d | 4.89 ± 0.18 f | 4.25 ± 0.53 abc |
| Parts | Methods-Solvents | AChE Inhibition (mg GALAE/g) | BChE Inhibition (mg GALAE/g) | Tyrosinase Inhibition (mg KAE/g) | Amylase Inhibition (mmol ACAE/g) | Glucosidase Inhibition (mmol ACAE/g) |
|---|---|---|---|---|---|---|
| Leaves | infusion | 5.18 ± 0.08 d | 0.91 ± 0.07 d | na | 0.15 ± 0.01 g | na |
| HAE-EA | 8.48 ± 0.21 ab | 2.97 ± 0.29 abc | 100.94 ± 1.41 e | 0.86 ± 0.04 c | 20.01 ± 0.08 b | |
| HAE-MEOH | 8.78 ± 0.07 a | na | 142.59 ± 1.14 d | 1.24 ± 0.01 a | 21.17 ± 0.02 a | |
| Maceration EA | 8.35 ± 0.20 ab | 2.19 ± 0.48 c | 94.19 ± 0.88 fg | 0.76 ± 0.02 de | 20.01 ± 0.24 b | |
| Maceration-MEOH | 8.62 ± 0.04 a | na | 146.76 ± 2.41 bc | 1.14 ± 0.01 b | 21.17 ± 0.11 a | |
| Maceration (not stirred)-EA | 8.56 ± 0.23 a | 3.50 ± 0.43 a | 96.38 ± 1.56 fg | 0.71 ± 0.02 e | 0.96 ± 0.19 d | |
| Maceration (not stirred)-MEOH | 8.56 ± 0.08 a | na | 146.06 ± 1.34 cd | 1.22 ± 0.03 a | 20.98 ± 0.06 a | |
| Bark | infusion | 7.07 ± 0.11 c | na | 25.64 ± 1.13 h | 0.42 ± 0.04 f | na |
| HAE-EA | 8.34 ± 0.41 ab | 2.52 ± 0.41 bc | 96.74 ± 0.95 f | 0.82 ± 0.01 cd | 20.04 ± 0.15 b | |
| HAE-MEOH | 8.76 ± 0.05 a | 0.59 ± 0.10 de | 150.75 ± 2.26 ab | 1.27 ± 0.02 a | na | |
| Maceration EA | 8.61 ± 0.16 a | 3.03 ± 0.38 ab | 104.13 ± 0.59 e | 0.69 ± 0.02 e | 19.87 ± 0.02 b | |
| Maceration-MEOH | 8.76 ± 0.04 a | 0.52 ± 0.03 de | 154.33 ± 1.10 a | 1.23 ± 0.01 a | na | |
| Maceration (not stirred)-EA | 8.04 ± 0.14 b | 2.19 ± 0.42 c | 92.22 ± 0.61 g | 0.75 ± 0.04 de | 18.48 ± 0.40 c | |
| Maceration (not stirred)-MEOH | 8.67 ± 0.01 a | 1.19 ± 0.27 d | 149.60 ± 1.98 bc | 1.19 ± 0.03 ab | na |
| Samples | MIC/MBC | S.a. | B.c. | L.m. | P.a. | E.c. | S.t. | En.cl. |
|---|---|---|---|---|---|---|---|---|
| Leaves- infusion | MIC | 0.75 | 0.37 | 0.37 | 0.30 | 0.43 | 0.37 | 0.56 |
| MBC | 1.50 | 0.75 | 0.75 | 0.43 | 0.87 | 0.75 | 0.75 | |
| Leaves- HAE-EA | MIC | 0.75 | 0.75 | 1.12 | 0.43 | 0.30 | 0.56 | 0.75 |
| MBC | 1.50 | 1.50 | 1.50 | 0.87 | 0.43 | 0.75 | 1.50 | |
| Leaves- HAE-MEOH | MIC | 0.37 | 0.13 | 0.27 | 0.27 | 0.27 | 0.27 | 0.37 |
| MBC | 0.75 | 0.18 | 0.37 | 0.37 | 0.37 | 0.37 | 0.75 | |
| Leaves- Maceration EA | MIC | 1.12 | 1.12 | 1.12 | 0.30 | 0.43 | 1.12 | 1.12 |
| MBC | 1.50 | 1.50 | 1.50 | 0.43 | 0.87 | 1.50 | 1.50 | |
| Leaves- Maceration-MEOH | MIC | 0.37 | 0.27 | 0.56 | 0.27 | 0.18 | 0.56 | 0.56 |
| MBC | 0.75 | 0.37 | 0.75 | 0.37 | 0.37 | 0.75 | 0.75 | |
| Leaves- Maceration (not stirred)-EA | MIC | 1.50 | 0.75 | 1.12 | 0.18 | 0.09 | 0.75 | 1.12 |
| MBC | 3.00 | 1.50 | 1.50 | 0.37 | 0.18 | 1.50 | 1.50 | |
| Leaves- Maceration (not stirred)-MEOH | MIC | 0.37 | 0.27 | 0.56 | 0.37 | 0.09 | 0.56 | 0.56 |
| MBC | 0.75 | 0.37 | 0.75 | 0.75 | 0.18 | 0.75 | 0.75 | |
| Bark- infusion | MIC | 0.56 | 0.27 | 0.75 | 0.75 | 0.18 | 0.37 | 0.37 |
| MBC | 0.75 | 0.37 | 1.50 | 1.50 | 0.37 | 0.75 | 0.75 | |
| Bark- HAE-EA | MIC | 0.75 | 0.56 | 0.75 | 0.65 | 0.75 | 0.75 | 1.50 |
| MBC | 1.50 | 0.75 | 1.50 | 0.87 | 1.50 | 1.50 | 3.00 | |
| Bark- HAE-MEOH | MIC | 0.56 | 0.18 | 0.37 | 0.37 | 0.56 | 0.37 | 0.37 |
| MBC | 0.75 | 0.37 | 0.75 | 0.75 | 0.75 | 0.75 | 0.75 | |
| Bark- Maceration EA | MIC | 0.56 | 0.18 | 0.75 | 0.75 | 0.56 | 1.12 | 0.75 |
| MBC | 0.75 | 0.37 | 1.5 | 1.50 | 0.75 | 1.50 | 1.50 | |
| Bark- Maceration-MEOH | MIC | 0.37 | 0.27 | 0.75 | 0.75 | 0.27 | 1.12 | 1.12 |
| MBC | 0.75 | 0.37 | 1.50 | 1.50 | 0.37 | 1.50 | 1.50 | |
| Bark- Maceration (not stirred)-EA | MIC | 2.25 | 2.25 | 1.50 | 1.50 | 2.25 | 1.12 | 1.12 |
| MBC | 3.00 | 3.00 | 3.00 | 3.00 | 3.00 | 1.50 | 1.50 | |
| Bark- Maceration (not stirred)-MEOH | MIC | 0.27 | 0.27 | 0.56 | 0.09 | 0.56 | 0.56 | 0.37 |
| MBC | 0.37 | 0.37 | 0.75 | 0.18 | 0.75 | 0.75 | 0.75 | |
| Streptomycin | MIC | 0.10 | 0.025 | 0.15 | 0.025 | 0.10 | 0.10 | 0.025 |
| MBC | 0.20 | 0.05 | 0.30 | 0.05 | 0.20 | 0.20 | 0.05 | |
| Ampicillin | MIC | 0.10 | 0.10 | 0.15 | 0.05 | 0.15 | 0.10 | 0.10 |
| MBC | 0.15 | 0.15 | 0.30 | 0.10 | 0.20 | 0.20 | 0.15 |
| Samples | MIC/MFC | A.f. | A.o. | A.n. | A.v. | P.o. | P.f. | T.v. | P.v.c. |
|---|---|---|---|---|---|---|---|---|---|
| Leaves- infusion | MIC | 2.25 | 0.37 | 0.37 | 0.37 | 0.09 | 0.18 | 0.27 | 0.56 |
| MFC | 3.00 | 0.75 | 0.75 | 0.75 | 0.18 | 0.37 | 0.37 | 0.75 | |
| Leaves- HAE-EA | MIC | 1.12 | 0.75 | 1.50 | 0.75 | 0.37 | 0.75 | 0.75 | 1.50 |
| MFC | 1.50 | 1.50 | 3.00 | 1.50 | 0.75 | 1.50 | 1.50 | 3.00 | |
| Leaves- HAE-MEOH | MIC | 0.56 | 0.75 | 0.75 | 0.27 | 1.12 | 0.37 | 0.22 | 0.75 |
| MFC | 1.12 | 1.50 | 1.50 | 0.37 | 1.50 | 0.75 | 0.44 | 1.50 | |
| Leaves- Maceration EA | MIC | 1.50 | 0.37 | 1.50 | 0.75 | 0.37 | 1.12 | 0.37 | 1.50 |
| MFC | 3.00 | 0.75 | 3.00 | 1.50 | 0.75 | 1.50 | 0.75 | 3.00 | |
| Leaves- Maceration-MEOH | MIC | 0.75 | 0.18 | 0.75 | 0.37 | 0.75 | 0.37 | 0.22 | 0.75 |
| MFC | 1.50 | 0.37 | 1.50 | 0.75 | 1.50 | 0.75 | 0.44 | 1.50 | |
| Leaves- Maceration (not stirred)-EA | MIC | 1.50 | 0.75 | 1.50 | 0.37 | 1.12 | 0.75 | 0.37 | 1.12 |
| MFC | 3.00 | 1.50 | 3.00 | 0.75 | 1.50 | 1.50 | 0.75 | 1.50 | |
| Leaves- Maceration (not stirred)-MEOH | MIC | 0.75 | 0.18 | 0.27 | 0.37 | 0.13 | 0.27 | 0.37 | 0.37 |
| MFC | 1.50 | 0.37 | 0.37 | 0.75 | 0.18 | 0.37 | 0.75 | 0.75 | |
| Bark- infusion | MIC | 0.75 | 0.37 | 0.75 | 1.12 | 1.12 | 0.75 | 0.27 | 0.84 |
| MFC | 1.50 | 0.75 | 1.50 | 1.50 | 1.15 | 1.50 | 0.37 | 1.12 | |
| Bark- HAE-EA | MIC | 3.00 | 3.00 | 3.00 | 1.50 | 2.25 | 3.00 | 0.56 | 3.00 |
| MFC | 6.00 | 6.00 | 6.00 | 3.00 | 3.00 | 6.00 | 0.75 | 6.00 | |
| Bark- HAE-MEOH | MIC | 0.75 | 0.37 | 0.75 | 1.50 | 0.75 | 0.37 | 0.004 | 0.56 |
| MFC | 1.50 | 0.75 | 1.50 | 3.00 | 1.50 | 0.75 | 0.008 | 0.75 | |
| Bark- Maceration EA | MIC | 1.12 | 0.37 | 0.75 | 0.75 | 0.75 | 1.12 | 0.27 | 0.75 |
| MFC | 1.50 | 0.75 | 1.50 | 1.50 | 1.50 | 1.50 | 0.37 | 1.50 | |
| Bark- Maceration-MEOH | MIC | 1.12 | 0.37 | 0.75 | 0.75 | 0.27 | 0.27 | 0.017 | 1.12 |
| MFC | 1.50 | 0.75 | 1.50 | 1.50 | 0.37 | 0.37 | 0.025 | 1.15 | |
| Bark- Maceration (not stirred)-EA | MIC | 0.75 | 0.37 | 0.56 | 0.37 | 0.56 | 0.56 | 0.27 | 0.75 |
| MFC | 1.50 | 0.75 | 0.75 | 0.75 | 0.75 | 0.75 | 0.37 | 1.50 | |
| Bark- Maceration (not stirred)-MEOH | MIC | 0.75 | 0.37 | 0.56 | 0.37 | 0.56 | 0.37 | 0.18 | 1.12 |
| MFC | 1.50 | 0.75 | 0.75 | 0.75 | 0.75 | 1.50 | 0.37 | 1.15 | |
| Bifonazole | MIC | 0.15 | 0.15 | 0.15 | 0.1 | 0.20 | 0.2 | 0.15 | 0.1 |
| MFC | 0.2 | 0.20 | 0.2 | 0.2 | 0.25 | 0.25 | 0.2 | 0.2 | |
| Ketoconazole | MIC | 0.2 | 0.15 | 0.2 | 0.2 | 1.00 | 0.2 | 1 | 0.2 |
| MFC | 0.5 | 0.20 | 0.5 | 0.5 | 1.50 | 0.5 | 1.5 | 0.3 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiavaroli, A.; Sinan, K.I.; Zengin, G.; Mahomoodally, M.F.; Bibi Sadeer, N.; Etienne, O.K.; Cziáky, Z.; Jekő, J.; Glamočlija, J.; Soković, M.; et al. Identification of Chemical Profiles and Biological Properties of Rhizophora racemosa G. Mey. Extracts Obtained by Different Methods and Solvents. Antioxidants 2020, 9, 533. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9060533
Chiavaroli A, Sinan KI, Zengin G, Mahomoodally MF, Bibi Sadeer N, Etienne OK, Cziáky Z, Jekő J, Glamočlija J, Soković M, et al. Identification of Chemical Profiles and Biological Properties of Rhizophora racemosa G. Mey. Extracts Obtained by Different Methods and Solvents. Antioxidants. 2020; 9(6):533. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9060533
Chicago/Turabian StyleChiavaroli, Annalisa, Koaudio Ibrahime Sinan, Gokhan Zengin, Mohamad Fawzi Mahomoodally, Nabeelah Bibi Sadeer, Ouattara Katinan Etienne, Zoltán Cziáky, József Jekő, Jasmina Glamočlija, Marina Soković, and et al. 2020. "Identification of Chemical Profiles and Biological Properties of Rhizophora racemosa G. Mey. Extracts Obtained by Different Methods and Solvents" Antioxidants 9, no. 6: 533. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9060533