Highlighting the Biological Potential of the Brown Seaweed Fucus spiralis for Skin Applications

 , , , , ,
, , , , ,  , ,
, ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Solvents and Reagents
2.2. Seaweed Collection and Preparation
2.3. Seaweed Extraction and Fractionation
2.4. Evaluation of Fucus spiralis Biological Properties
2.4.1. Quantification of Total Phenolic Content (TPC)
2.4.2. Antioxidant Capacity
1,1-Diphenyl-2-picrylhydrazyl (DPPH) Radical Scavenging Activity
Oxygen Radical Absorbance Capacity (ORAC)
Ferric Reducing Antioxidant Power (FRAP)
2.4.3. Enzymatic Activities
Hyaluronidase Activity
Collagenase Activity
Elastase Activity
2.4.4. Antimicrobial Activity
2.4.5. Evaluation of Biological Activities of F. spiralis Fractions on In Vitro Cellular Models
Maintenance of Cell Culture
Evaluation of Cytotoxicity
Quantification of Nitric Oxide (NO) on RAW 264.7 Cells
Reactive Oxygen Species (ROS) Production on HaCaT Cells
2.4.6. Chemical Characterization
UV–Visible Absorption Spectra
LC-MS Analysis
2.4.7. Data and Statistical Analysis
3. Results
3.1. Antioxidant Capacity
3.2. Enzymatic Activity
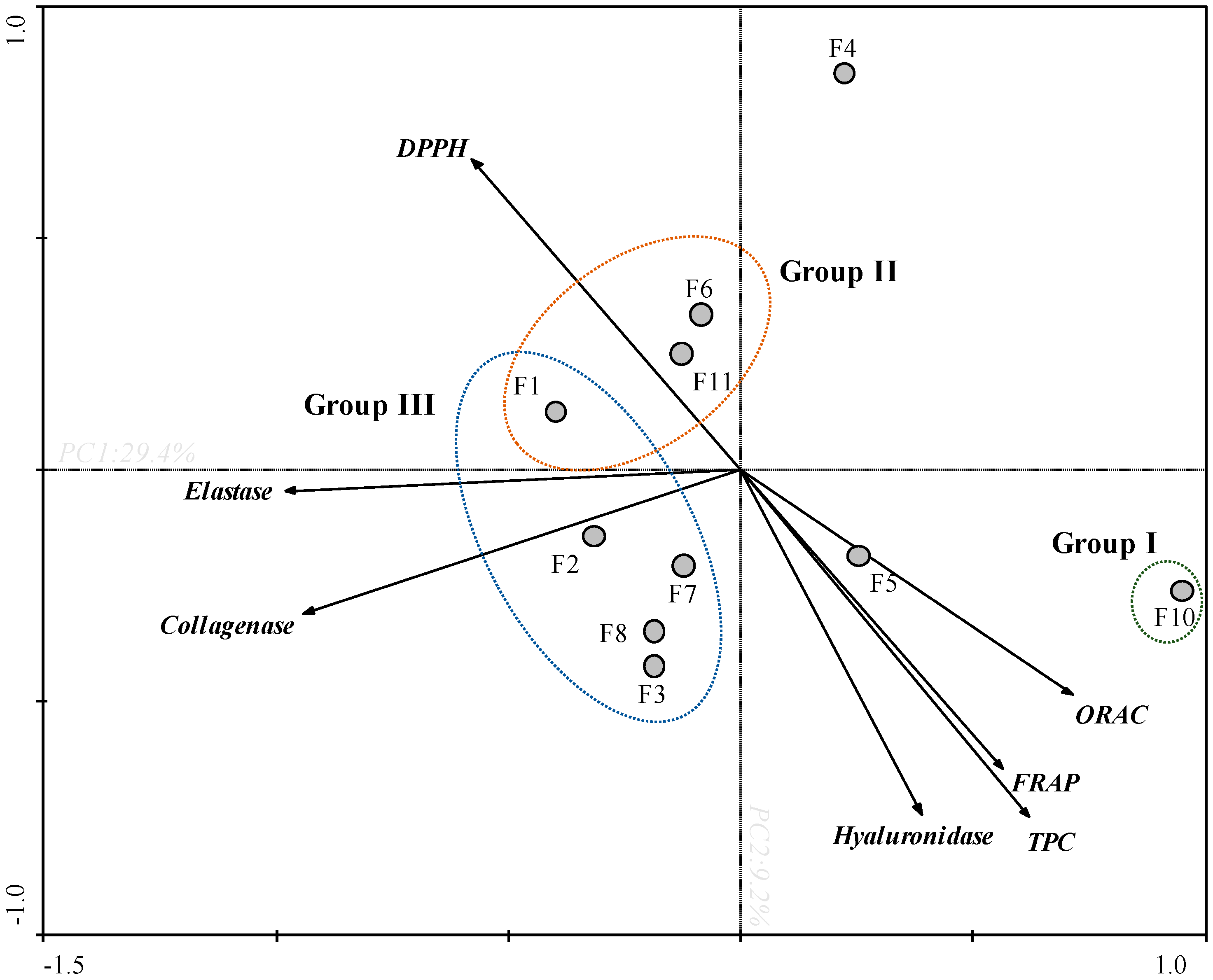
3.3. Principal Component Analysis (PCA)
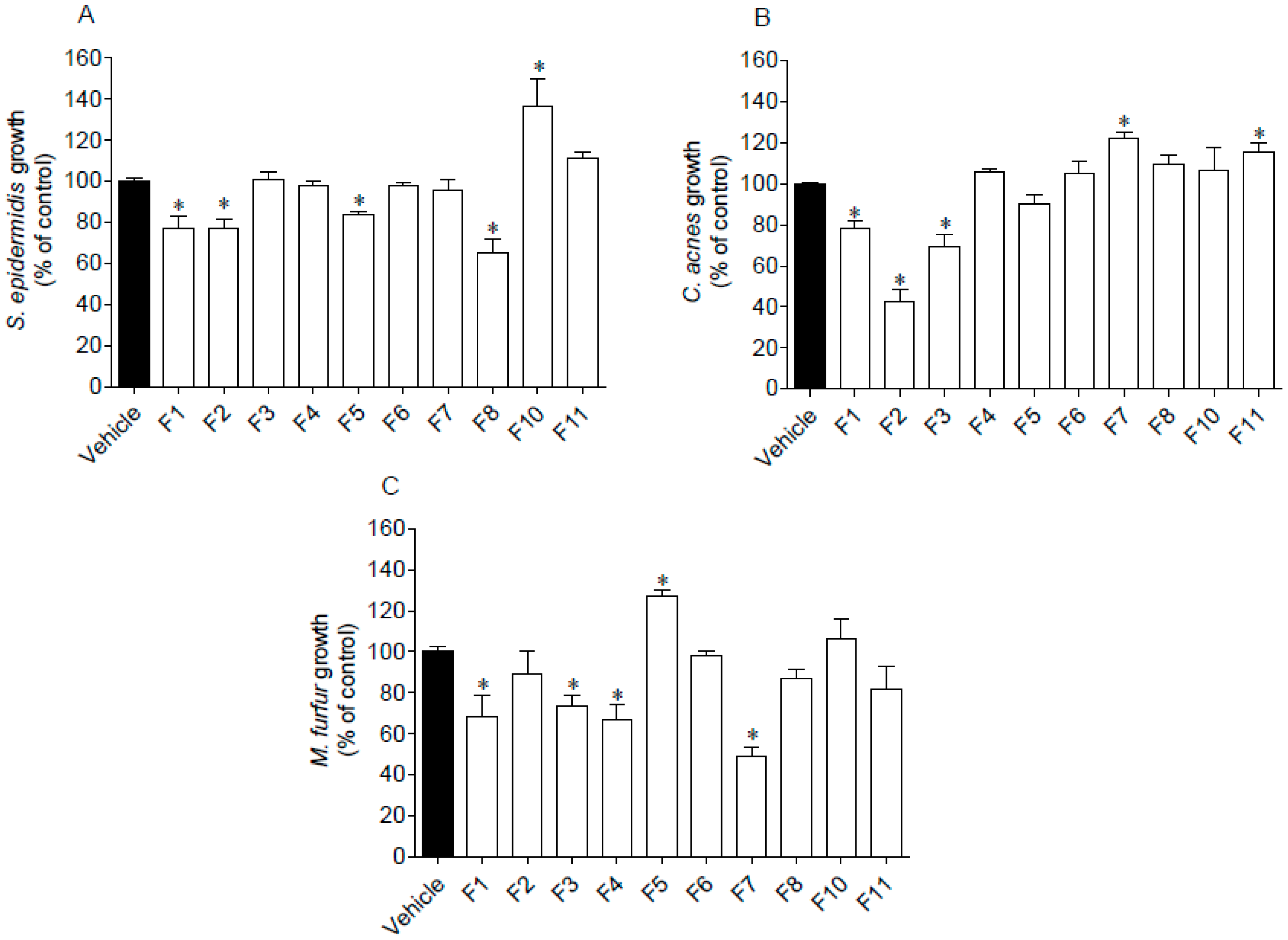
3.4. Antimicrobial Activity
3.5. Biological Activities of F. spiralis Fractions on In Vitro Cellular Models
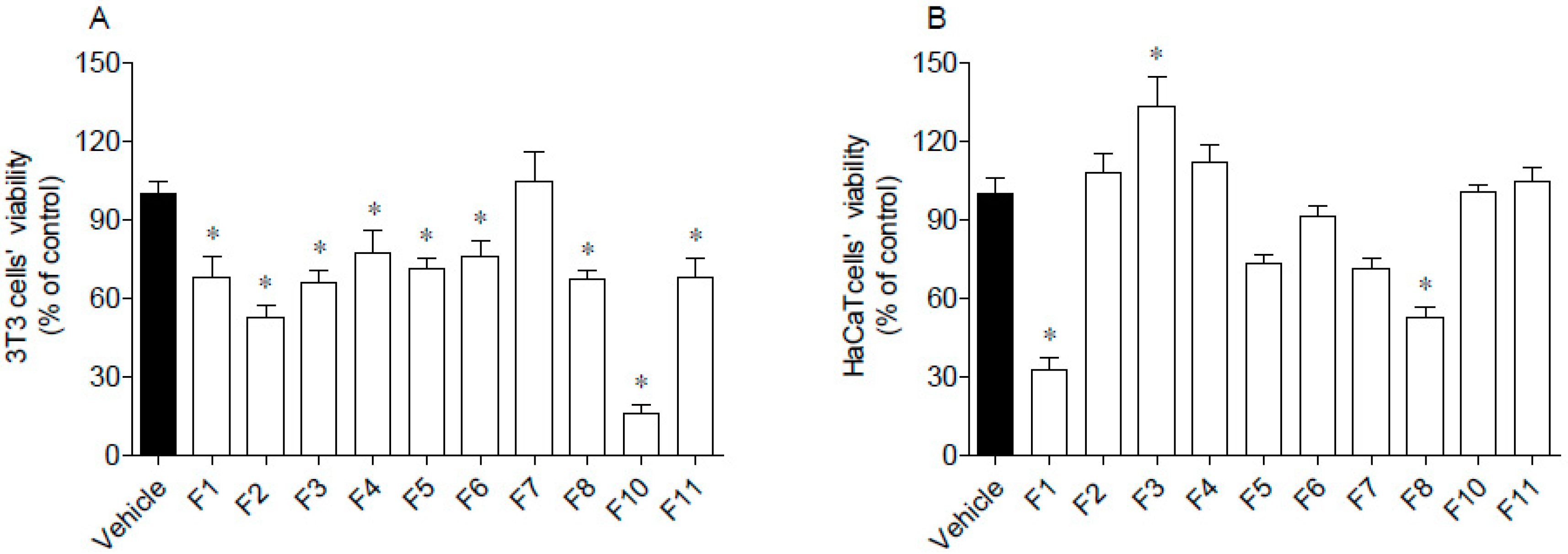
3.5.1. Cytotoxicity on 3T3 and HaCaT Cells
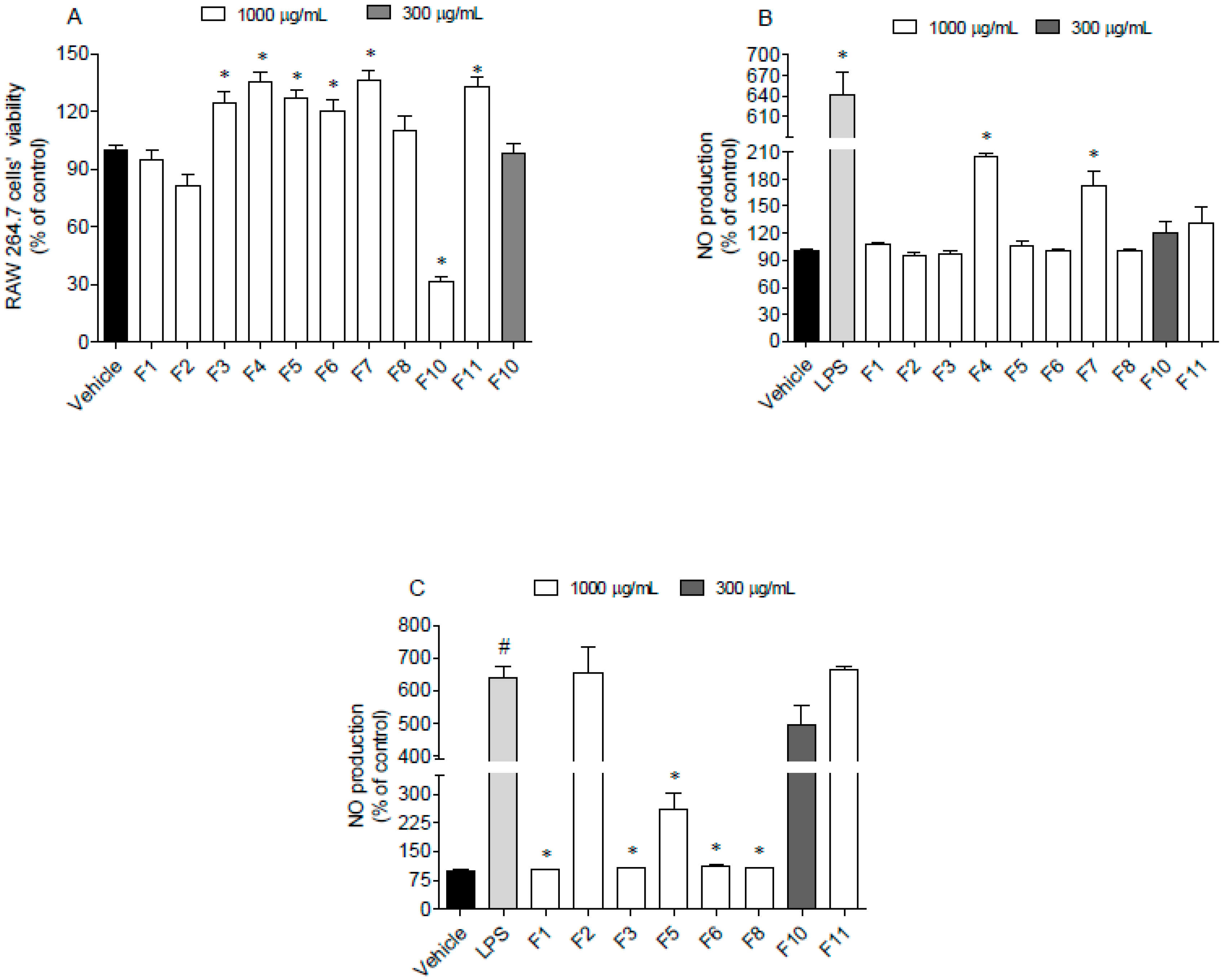
3.5.2. Nitric Oxide Production on RAW264.7 Cells
3.5.3. Reactive Oxygen Species (ROS) Production on HaCaT Cells after Exposure to H2O2 or UVB Light
3.6. UV–Visible Absorption Spectra
3.7. Chemical Caracterization by LC-MS
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Espinosa-Leal, C.A.; García-Lara, S. Current methods for the discovery of new active ingredients from natural products for cosmeceutical applications. Planta Med. 2019, 85, 535–551. [Google Scholar] [CrossRef] [Green Version]
- Fernando, I.P.S.; Kim, K.-N.; Kim, D.; Jeon, Y.-J. Algal polysaccharides: Potential bioactive substances for cosmeceutical applications. Crit. Rev. Biotechnol. 2018, 39, 99–113. [Google Scholar] [CrossRef] [PubMed]
- Ganceviciene, R.; Liakou, A.I.; Theodoridis, A.; Makrantonaki, E.; Zouboulis, C. Skin anti-aging strategies. Dermato-Endocrinology 2012, 4, 308–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masaki, H. Role of antioxidants in the skin: Anti-aging effects. J. Dermatol. Sci. 2010, 58, 85–90. [Google Scholar] [CrossRef]
- Dunaway, S.; Odin, R.; Zhou, L.; Ji, L.; Zhang, Y.; Kadekaro, A.L. Natural antioxidants: Multiple mechanisms to protect skin from solar radiation. Front. Pharmacol. 2018, 9, 392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahal, A.; Kumar, A.; Singh, V.; Yadav, B.; Tiwari, R.; Chakraborty, S.; Dhama, K. Oxidative stress, prooxidants, and antioxidants: The interplay. BioMed Res. Int. 2014, 2014, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imokawa, G.; Ishida, K. Biological mechanisms underlying the ultraviolet radiation-induced formation of skin wrinkling and sagging I: Reduced skin elasticity, highly associated with enhanced dermal elastase activity, triggers wrinkling and sagging. Int. J. Mol. Sci. 2015, 16, 7753–7775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papakonstantinou, E.; Roth, M.; Karakiulakis, G. Hyaluronic acid: A key molecule in skin aging. Dermato-Endocrinology 2012, 4, 253–258. [Google Scholar] [CrossRef] [Green Version]
- Aguilar-Toalá, J.; Hernández-Mendoza, A.; González-Córdova, A.; Vallejo-Cordoba, B.; Liceaga, A.M. Potential role of natural bioactive peptides for development of cosmeceutical skin products. Peptides 2019, 122, 170170. [Google Scholar] [CrossRef]
- Pereira, L. Seaweeds as source of bioactive substances and skin care therapy—Cosmeceuticals, algotheraphy, and thalassotherapy. Cosmetics 2018, 5, 68. [Google Scholar] [CrossRef] [Green Version]
- Jesumani, V.; Du, H.; Aslam, M.; Pei, P.; Huang, N. Potential use of seaweed bioactive compounds in skincare—A review. Mar. Drugs 2019, 17, 688. [Google Scholar] [CrossRef] [Green Version]
- Murray, P.M.; Moane, S.; Collins, C.; Beletskaya, T.; Thomas, O.P.; Duarte, A.W.; Nobre, F.S.; Owoyemi, I.O.; Pagnocca, F.C.; Sette, L.D.; et al. Sustainable production of biologically active molecules of marine based origin. New Biotechnol. 2013, 30, 839–850. [Google Scholar] [CrossRef]
- Alves, C.; Silva, J.; Pinteus, S.; Gaspar, H.; Alpoim, M.C.M.D.C.; Botana, L.M.; Pedrosa, R. From marine origin to therapeutics: The antitumor potential of marine algae-derived compounds. Front. Pharmacol. 2018, 9, 777. [Google Scholar] [CrossRef] [Green Version]
- Pinteus, S.; Lemos, M.F.; Alves, C.; Neugebauer, A.; Silva, J.; Thomas, O.P.; Botana, L.M.; Gaspar, H.; Pedrosa, R. Marine invasive macroalgae: Turning a real threat into a major opportunity - the biotechnological potential of Sargassum muticum and Asparagopsis armata. Algal Res. 2018, 34, 217–234. [Google Scholar] [CrossRef]
- Silva, J.; Alves, C.; Freitas, R.; Martins, A.; Pinteus, S.; Ribeiro, J.; Gaspar, H.; Alfonso, A.; Pedrosa, R. Antioxidant and neuroprotective potential of the brown seaweed Bifurcaria bifurcata in an in vitro Parkinson’s disease model. Mar. Drugs 2019, 17, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinteus, S.; Lemos, M.F.; Freitas, R.; Duarte, I.M.; Alves, C.; Silva, J.; Marques, S.C.; Pedrosa, R. Medusa polyps adherence inhibition: A novel experimental model for antifouling assays. Sci. Total Environ. 2020, 715, 136796. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Sharifi-Rad, J.; Seca, A.M.L.; Pinto, D.C.G.A.; Michalak, I.; Trincone, A.; Mishra, A.P.; Nigam, M.; Zam, W.; Martins, N.; et al. Current trends on seaweeds: Looking at chemical composition, phytopharmacology, and cosmetic applications. Molecules 2019, 24, 4182. [Google Scholar] [CrossRef] [Green Version]
- Catarino, M.D.; Silva, A.M.S.; Cardoso, S.M. Phycochemical Constituents and Biological Activities of Fucus spp. Mar. Drugs 2018, 16, 249. [Google Scholar] [CrossRef] [Green Version]
- Fitton, J.H.; Dell’Acqua, G.; Gardiner, V.-A.; Karpiniec, S.S.; Stringer, D.N.; Davis, E. Topical benefits of two fucoidan-rich extracts from marine macroalgae. Cosmetics 2015, 2, 66–81. [Google Scholar] [CrossRef] [Green Version]
- Fujimura, T.; Tsukahara, K.; Moriwaki, S.; Kitahara, T.; Sano, T.; Takema, Y. Treatment of human skin with an extract of Fucus vesiculosus changes its thickness and mechanical properties. J. Cosmet. Sci. 2002, 53, 1–9. [Google Scholar] [PubMed]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144. [Google Scholar]
- Pinteus, S.; Silva, J.; Alves, C.; Horta, A.; Fino, N.; Rodrigues, A.I.; Mendes, S.; Pedrosa, R. Cytoprotective effect of seaweeds with high antioxidant activity from the Peniche coast (Portugal). Food Chem. 2017, 218, 591–599. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Davalos, A.; Gómez-Cordovés, C.; Bartolomé, B. Extending applicability of the Oxygen Radical Absorbance Capacity (ORAC−Fluorescein) Assay. J. Agric. Food Chem. 2004, 52, 48–54. [Google Scholar] [CrossRef]
- Benzie, I.; Strain, J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of “Antioxidant Power”: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Yahaya, Y.A.; Don, M.M. Evaluation of Trametes lactinea extracts on the inhibition of hyaluronidase, lipoxygenase and xanthine oxidase activities in vitro. J. Phys. Sci. 2012, 23, 1–15. [Google Scholar]
- Bailey, L.C.; Levine, N.A. Optimization of the USP assay for hyaluronidase. J. Pharm. Biomed. Anal. 1993, 11, 285–292. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Yang, E.-J.; Yim, E.-Y.; Song, G.; Kim, G.-O.; Hyun, C.-G. Inhibition of nitric oxide production in lipopolysaccharide-activated RAW 264.7 macrophages by Jeju plant extracts. Interdiscip. Toxicol. 2009, 2, 245–249. [Google Scholar] [CrossRef] [Green Version]
- Marto, J.; Neves, Â.; Gonçalves, L.M.; Pinto, P.; Almeida, C.M.M.; Simões, S. Rice water: A traditional ingredient with anti-aging efficacy. Cosmetics 2018, 5, 26. [Google Scholar] [CrossRef] [Green Version]
- Pan, H. A non-covalent dimer formed in electrospray ionisation mass spectrometry behaving as a precursor for fragmentations. Rapid Commun. Mass Spectrom. 2008, 22, 3555–3560. [Google Scholar] [CrossRef]
- Ferreres, F.; Lopes, G.; Gil-Izquierdo, A.; Andrade, P.B.; Sousa, C.; Mouga, T.; Valentão, P. Phlorotannin extracts from Fucales characterized by HPLC-DAD-ESI-MSn: Approaches to hyaluronidase inhibitory capacity and antioxidant properties. Mar. Drugs 2012, 10, 2766–2781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tierney, M.S.; Soler-Vila, A.; Rai, D.K.; Croft, A.K.; Brunton, N.; Smyth, T. UPLC-MS profiling of low molecular weight phlorotannin polymers in Ascophyllum nodosum, Pelvetia canaliculata and Fucus spiralis. Metabolomics 2013, 10, 524–535. [Google Scholar] [CrossRef]
- Lopes, G.; Barbosa, M.; Vallejo, F.; Gil-Izquierdo, Á.; Andrade, P.B.; Valentão, P.; Pereira, D.M.; Ferreres, F. Profiling phlorotannins from Fucus spp. of the Northern Portuguese coastline: Chemical approach by HPLC-DAD-ESI/MS and UPLC-ESI-QTOF/MS. Algal Res. 2018, 29, 113–120. [Google Scholar] [CrossRef]
- Hermund, D.B.; Plaza, M.; Turner, C.; Jónsdóttir, R.; Kristinsson, H.G.; Jacobsen, C.; Nielsen, K.F. Structure dependent antioxidant capacity of phlorotannins from Icelandic Fucus vesiculosus by UHPLC-DAD-ECD-QTOFMS. Food Chem. 2018, 240, 904–909. [Google Scholar] [CrossRef] [Green Version]
- Catarino, M.D.; Silva, A.M.S.; Mateus, N.; Cardoso, S.M. Optimization of phlorotannins extraction from Fucus vesiculosus and evaluation of their potential to prevent metabolic disorders. Mar. Drugs 2019, 17, 162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catarino, M.D.; Silva, A.M.S.; Cardoso, S.M. Fucaceae: A source of bioactive phlorotannins. Int. J. Mol. Sci. 2017, 18, 1327. [Google Scholar] [CrossRef] [Green Version]
- Paiva, L.; Lima, E.; Neto, A.I.; Baptista, J. Seasonal variability of the biochemical composition and antioxidant properties of Fucus spiralis at two Azorean islands. Mar. Drugs 2018, 16, 248. [Google Scholar] [CrossRef] [Green Version]
- Rosa, G.P.; Barreto, M.C.; Seca, A. Pharmacological effects of Fucus spiralis extracts and phycochemicals: A comprehensive review. Bot. Mar. 2019, 62, 167–178. [Google Scholar] [CrossRef]
- Li, Y.-X.; Wijesekara, I.; Kim, S.-K.; Li, Y. Phlorotannins as bioactive agents from brown algae. Process. Biochem. 2011, 46, 2219–2224. [Google Scholar] [CrossRef]
- Pimentel, F.B.; Alves, R.C.; Rodrigues, F.; Oliveira, M.B.P. Macroalgae-derived ingredients for cosmetic industry—An update. Cosmetics 2017, 5, 2. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.Y.; Kim, J.-Y.; Shin, W.-S.; Lee, S.J.; Chi, S.-G.; Lee, J.-Y.; Park, M.-J. Saccharina japonica extract suppresses stemness of glioma stem cells by degrading epidermal growth factor receptor/epidermal growth factor receptor variant III. J. Med. Food 2018, 21, 496–505. [Google Scholar] [CrossRef] [PubMed]
- Cikoš, A.-M.; Jerković, I.; Molnar, M.; Šubarić, D.; Jokić, S. New trends for macroalgal natural products applications. Nat. Prod. Res. 2019, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Lee, J.-E.; Kim, K.H.; Kang, N.J. Beneficial effects of marine algae-derived carbohydrates for skin health. Mar. Drugs 2018, 16, 459. [Google Scholar] [CrossRef] [Green Version]
- Dreno, B.; Pécastaings, S.; Corvec, S.; Veraldi, S.; Khammari, A.; Roques, C. Cutibacterium acnes (Propionibacterium acnes) and acne vulgaris: A brief look at the latest updates. J. Eur. Acad. Dermatol. Venereol. 2018, 32, 5–14. [Google Scholar] [CrossRef] [Green Version]
- Gaitanis, G.; Magiatis, P.; Hantschke, M.; Bassukas, I.D.; Velegraki, A. The Malassezia Genus in Skin and Systemic Diseases. Clin. Microbiol. Rev. 2012, 25, 106–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theelen, B.; Cafarchia, C.; Gaitanis, G.; Bassukas, I.D.; Boekhout, T.; Dawson, T. Malassezia ecology, pathophysiology, and treatment. Med Mycol. 2018, 56, S10–S25. [Google Scholar] [CrossRef] [Green Version]
- Lopes, G.; Sousa, C.; Silva, L.R.; Pinto, E.; Andrade, P.B.; Bernardo, J.; Mouga, T.; Valentão, P. Can phlorotannins purified extracts constitute a novel pharmacological alternative for microbial infections with associated inflammatory conditions? PLoS ONE 2012, 7, e31145. [Google Scholar] [CrossRef]
- Barbalace, M.C.; Malaguti, M.; Giusti, L.; Lucacchini, A.; Hrelia, S.; Angeloni, C. Anti-inflammatory activities of marine algae in neurodegenerative diseases. Int. J. Mol. Sci. 2019, 20, 3061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopes, G.; Daletos, G.; Proksch, P.; Andrade, P.B.; Valentão, P. Anti-inflammatory potential of monogalactosyl diacylglycerols and a monoacylglycerol from the edible brown seaweed Fucus spiralis Linnaeus. Mar. Drugs 2014, 12, 1406–1418. [Google Scholar] [CrossRef]
- Seca, A.M.L.; Gouveia, V.L.M.; Barreto, M.D.C.; Silva, A.M.S.; Pinto, D.C.G.A. Comparative study by GC-MS and chemometrics on the chemical and nutritional profile of Fucus spiralis L. juvenile and mature life-cycle phases. Environ. Boil. Fishes 2018, 30, 2539–2548. [Google Scholar] [CrossRef]
- Prasedya, E.S.; Syafitri, S.M.; Geraldine, B.A.F.D.; Hamdin, C.D.; Frediansyah, A.; Miyake, M.; Kobayashi, D.; Hazama, A.; Sunarpi, H. UVA photoprotective activity of brown macroalgae Sargassum cristafolium. Biomedicines 2019, 7, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pangestuti, R.; Siahaan, E.A.; Kim, S.-K. Photoprotective substances derived from marine algae. Mar. Drugs 2018, 16, 399. [Google Scholar] [CrossRef] [Green Version]
- Pozharitskaya, O.N.; Shikov, A.N.; Obluchinskaya, E.D.; Vuorela, H. The pharmacokinetics of fucoidan after topical application to rats. Mar. Drugs 2019, 17, 687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pozharitskaya, O.N.; Obluchinskaya, E.D.; Shikov, A.N. Mechanisms of bioactivities of fucoidan from the brown seaweed Fucus vesiculosus L. of the Barents Sea. Mar. Drugs 2020, 18, 275. [Google Scholar] [CrossRef] [PubMed]
- Obluchinsksya, E.D.; Makarova, M.N.; Pozharitskaya, O.N.; Shikov, A.N. Composition and anticoagulant properties of dry Fucus extract. Pharm. Chem. J. 2015, 49, 183–186. [Google Scholar] [CrossRef]
- Jacobsen, C.; Sørensen, A.-D.M.; Holdt, S.; Akoh, C.C.; Hermund, D.B. Source, extraction, characterization, and applications of novel antioxidants from seaweed. Annu. Rev. Food Sci. Technol. 2019, 10, 541–568. [Google Scholar] [CrossRef]
- Załuski, D.; Olech, M.; Kuźniewski, R.; Verpoorte, R.; Nowak, R.; Smolarz, H.D. LC-ESI-MS/MS profiling of phenolics from Eleutherococcus spp. inflorescences, structure-activity relationship as antioxidants, inhibitors of hyaluronidase and acetylcholinesterase. Saudi Pharm. J. 2017, 25, 734–743. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fraction | Yield (%) | TPC a | DPPH b | ORAC c | FRAP d |
|---|---|---|---|---|---|
| F1 | 1.43 | 62.7 ± 6.0 | 645.7 (588.3–708.6) | 1319.6 ± 162.5 | 333.8 ± 47.1 |
| F2 | 1.24 | 230.9 ± 11.2 | 235.1 (260.4–267.6) | 3214.4 ± 237.9 | 700.9 ± 118.9 |
| F3 | 10.31 | 362.1 ± 9.7 | 157.6 (113.1–219.8) | 3557.9 ± 243.1 | 1821.8 ± 253.7 |
| F4 | 1.94 | 48.6 ± 7.9 | >1000 | 1755.9 ± 275.3 | 504.0 ± 56.3 |
| F5 | 6.99 | 309.5 ± 12.7 | 244.7 (208.9–286.8) | 4238.1 ± 134.6 | 1209.4 ± 114.4 |
| F6 | 6.61 | 95.1 ± 7.3 | >1000 | 2794.1 ± 269.1 | 505.2 ± 69.3 |
| F7 | 0.73 | 272.0 ± 26.7 | 631.5 (522.0–764.0) | 3518.7 ± 376.8 | 1602.1 ± 95.4 |
| F8 | 1.83 | 197.7 ± 8.4 | 257.2 (231.4–285.8 | 3319.4 ± 138.3 | 674.3 ± 98.2 |
| F9 | 0.01 | - | - | - | - |
| F10 | 0.65 | 1679.8 ± 34.0 | 38.5 (29.51–50.36) | 16464.5 ± 1223.4 | 2378.2 ± 93.5 |
| F11 | 11.34 | 121.5 ± 4.8 | 324.5 (289.4–363.8) | 2353.6 ± 88.9 | 735.1 ± 102.7 |
| BHT | - | - | 164.5 (142.7–189.7) | 142.9 ± 9.1 | 2821.5 ± 51.5 |
| Fraction | Collagenase | Elastase | Hyaluronidase |
|---|---|---|---|
| F1 | >1000 | >1000 | 148.9 (122.2–181.3) |
| F2 | >1000 | >1000 | 79.5 (66.1–95.6) |
| F3 | 512.9 (442.8–594.1) | >1000 | 348.8 (290.5–418.7) |
| F4 | 40.4 (26.2–62.5) | 67.8 (47.6–96.6) | 61.1 (47.3–78.9) |
| F5 | 89.9 (69.1–117.1) | 409.0 (293.9–569.2) | >1000 |
| F6 | 156.7 (123.6–198.5) | >1000 | 128.8 (103.0–161.1) |
| F7 | 4.3 (3.5–5.3) | 123.8 (103.0–148.7) | >1000 |
| F8 | 391.6 (328.7–466.6) | >1000 | >1000 |
| F10 | 0.037 (0.009–0.142) | 3.0 (2.5–3.6) | >1000 |
| F11 | 31.3 (28.4–34.5) | 586.5 (445.8–771.6) | 110.1 (89.1–136.1) |
| EGCG | 4.8 (4.1–5.5) | 113.9 (80.7–160.0) | 119.1 (126.1–320.4) |
| M (m/z) | [M−H]− (m/z) | [M+H]+ (m/z) | Other Ions | Tentative Assignment | References |
|---|---|---|---|---|---|
| 374 | 373 | 375 | - | Trifucol | [34,35,36] |
| 498 | 497 | 499 | - | Tetrafucol or Fucodiphloroethol or their isomers | [32,33,35,36] |
| 498 | 497 | 499 | - | ||
| 622 | 621 | 623 | - | Trifucophlorethol or its isomers | [34,35,36] |
| 746 | 745 | 747 | 1493 [2M+H]+ | Hexafucol or its isomers | [31,32,33,34,35,36] |
| 746 | 745 | 747 | 1493 [2M+H]+ | ||
| 870 | 869 | 871 | - | Difucotetraphloroethol or Trifucotriphloethol or their isomers | [32,33,35,36] |
| 870 | 869 | 871 | - | ||
| 994 | 993 | 995 | 496 [M−2H]2− | Tetrafucotetraphlorethol, or Pentafucodiphlorethol or Hexafucophlorethol or their isomers | [32,33,36] |
| 994 | 993 | 995 | 496 [M−2H]2− | ||
| 994 | 993 | 995 | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Freitas, R.; Martins, A.; Silva, J.; Alves, C.; Pinteus, S.; Alves, J.; Teodoro, F.; Ribeiro, H.M.; Gonçalves, L.; Petrovski, Ž.; et al. Highlighting the Biological Potential of the Brown Seaweed Fucus spiralis for Skin Applications. Antioxidants 2020, 9, 611. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9070611
Freitas R, Martins A, Silva J, Alves C, Pinteus S, Alves J, Teodoro F, Ribeiro HM, Gonçalves L, Petrovski Ž, et al. Highlighting the Biological Potential of the Brown Seaweed Fucus spiralis for Skin Applications. Antioxidants. 2020; 9(7):611. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9070611
Chicago/Turabian StyleFreitas, Rafaela, Alice Martins, Joana Silva, Celso Alves, Susete Pinteus, Joana Alves, Fernando Teodoro, Helena Margarida Ribeiro, Lídia Gonçalves, Željko Petrovski, and et al. 2020. "Highlighting the Biological Potential of the Brown Seaweed Fucus spiralis for Skin Applications" Antioxidants 9, no. 7: 611. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9070611