Beneficial Regulatory Effects of Polymethoxyflavone—Rich Fraction from Ougan (Citrus reticulata cv. Suavissima) Fruit on Gut Microbiota and Identification of Its Intestinal Metabolites in Mice


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Animals
2.2. Extraction of PMF-Rich Fraction from Ougan Fruit
2.3. Collection of the Intestinal Contents
2.4. Extraction of Flavonoid Components from the Intestinal Contents
2.5. Identification of Flavonoids
2.6. DNA Extraction and 16S ribosomal DNA Sequencing of Intestinal Content from Mice
2.7. Sequencing Data Processing
2.8. Statistical Analysis
3. Results
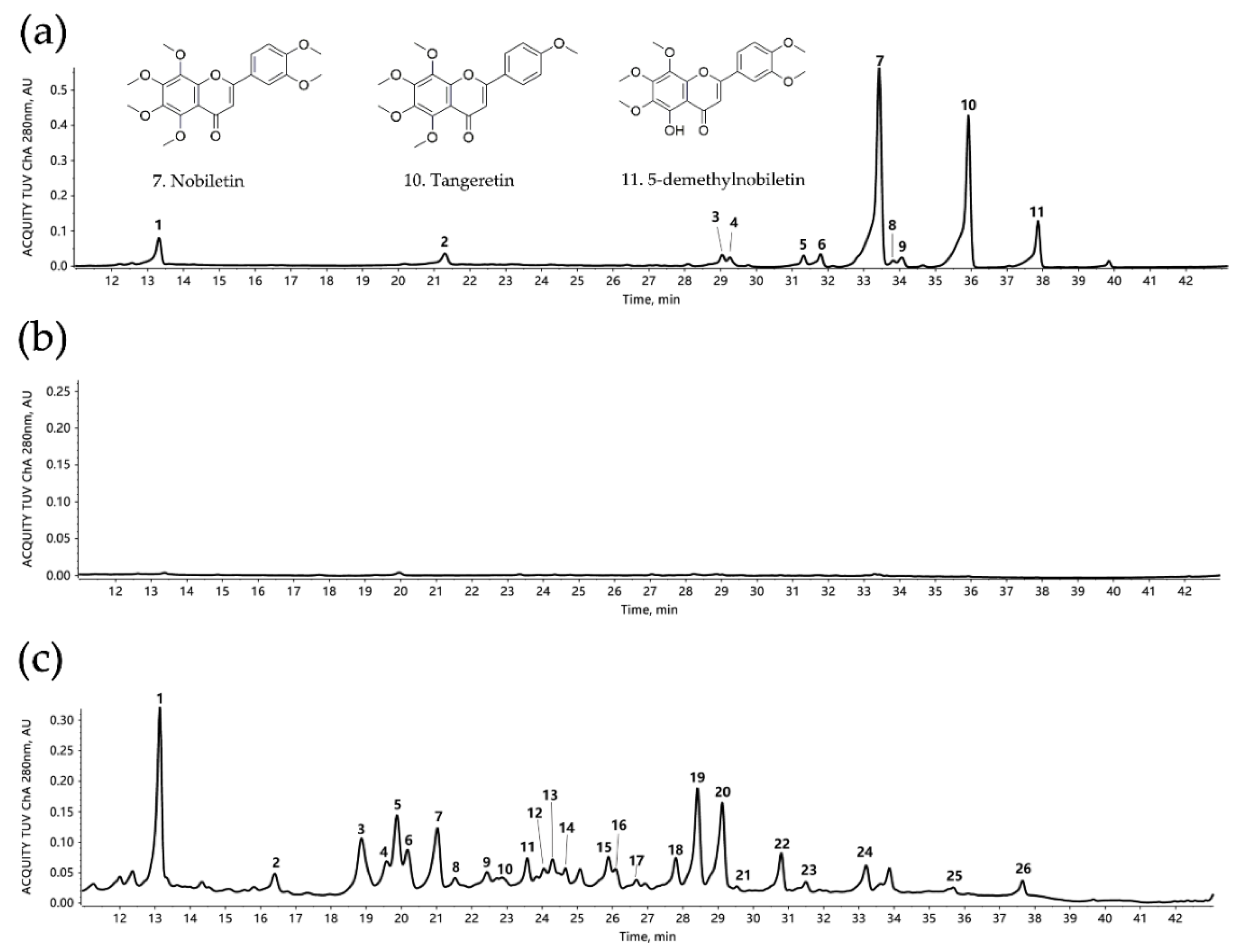
3.1. Identification of PMF-Rich Fraction
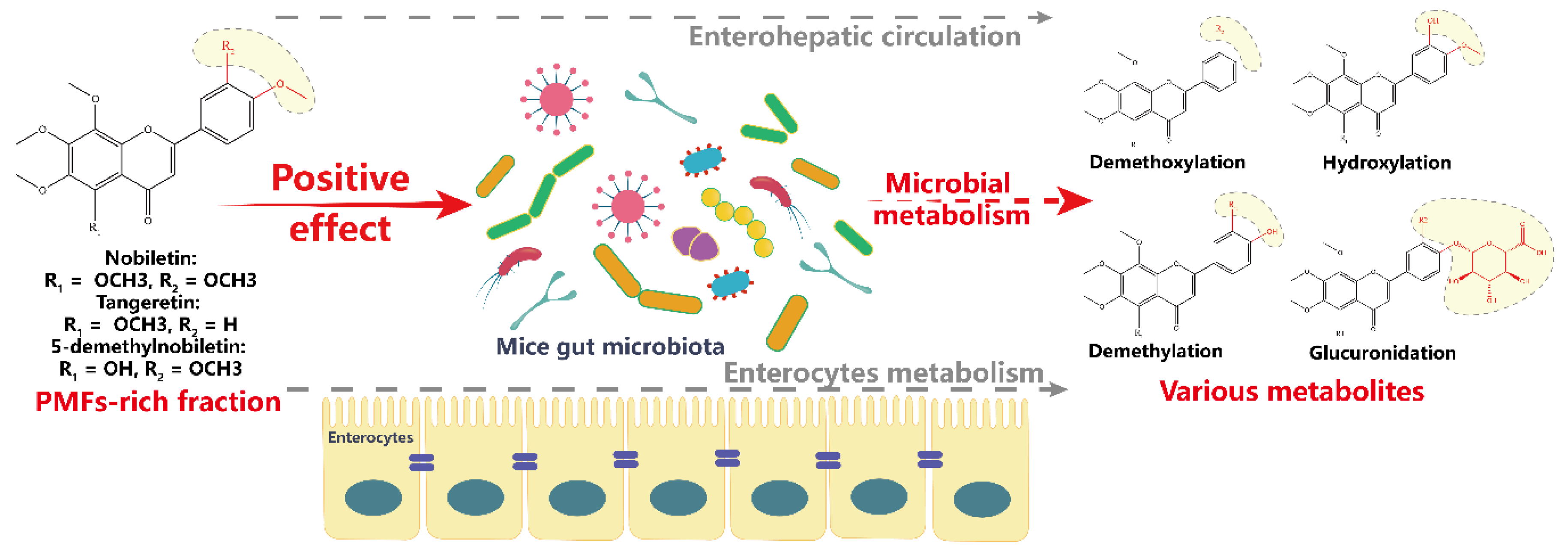
3.2. Identification of Metabolites of PMFs in Intestinal Content
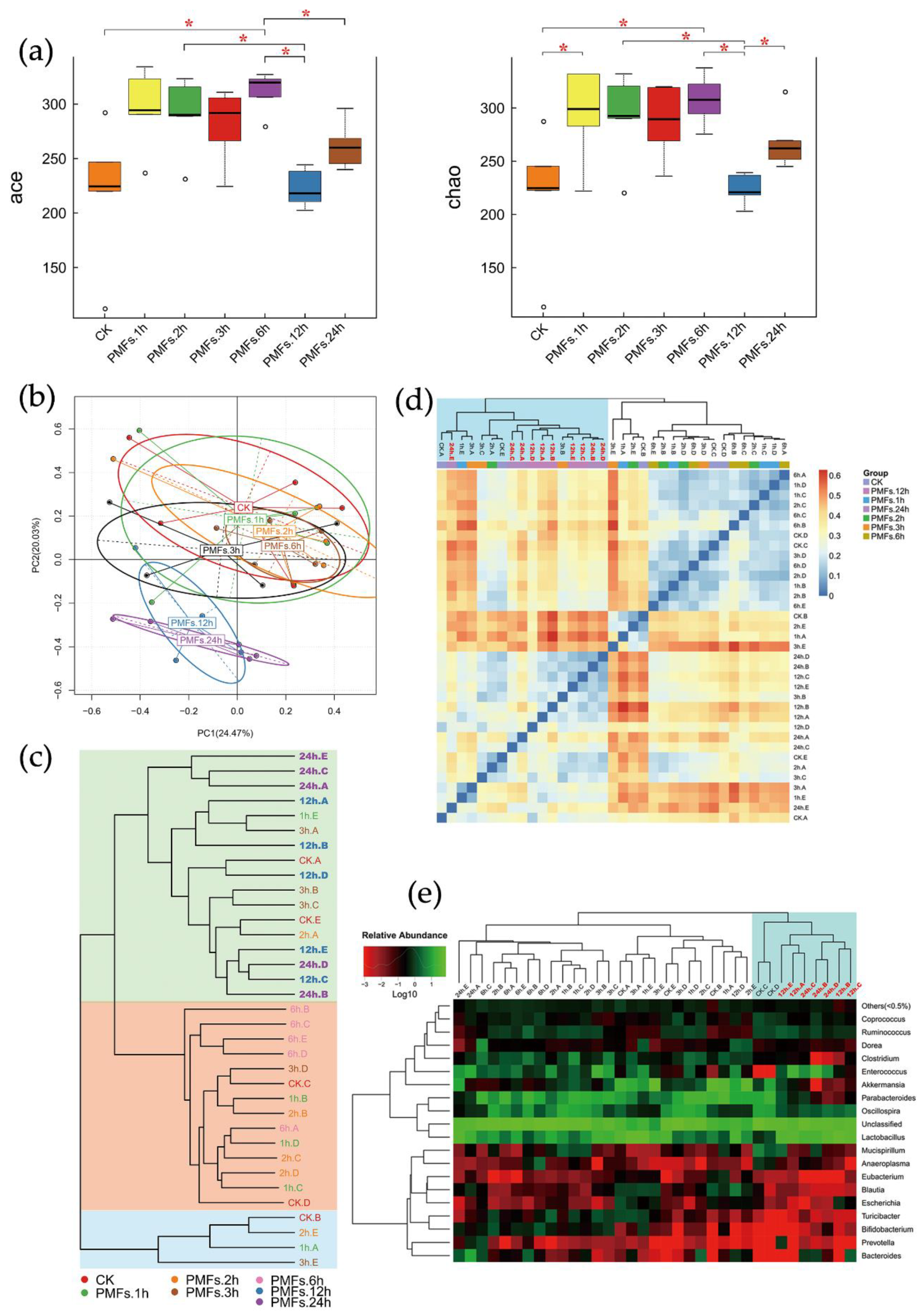
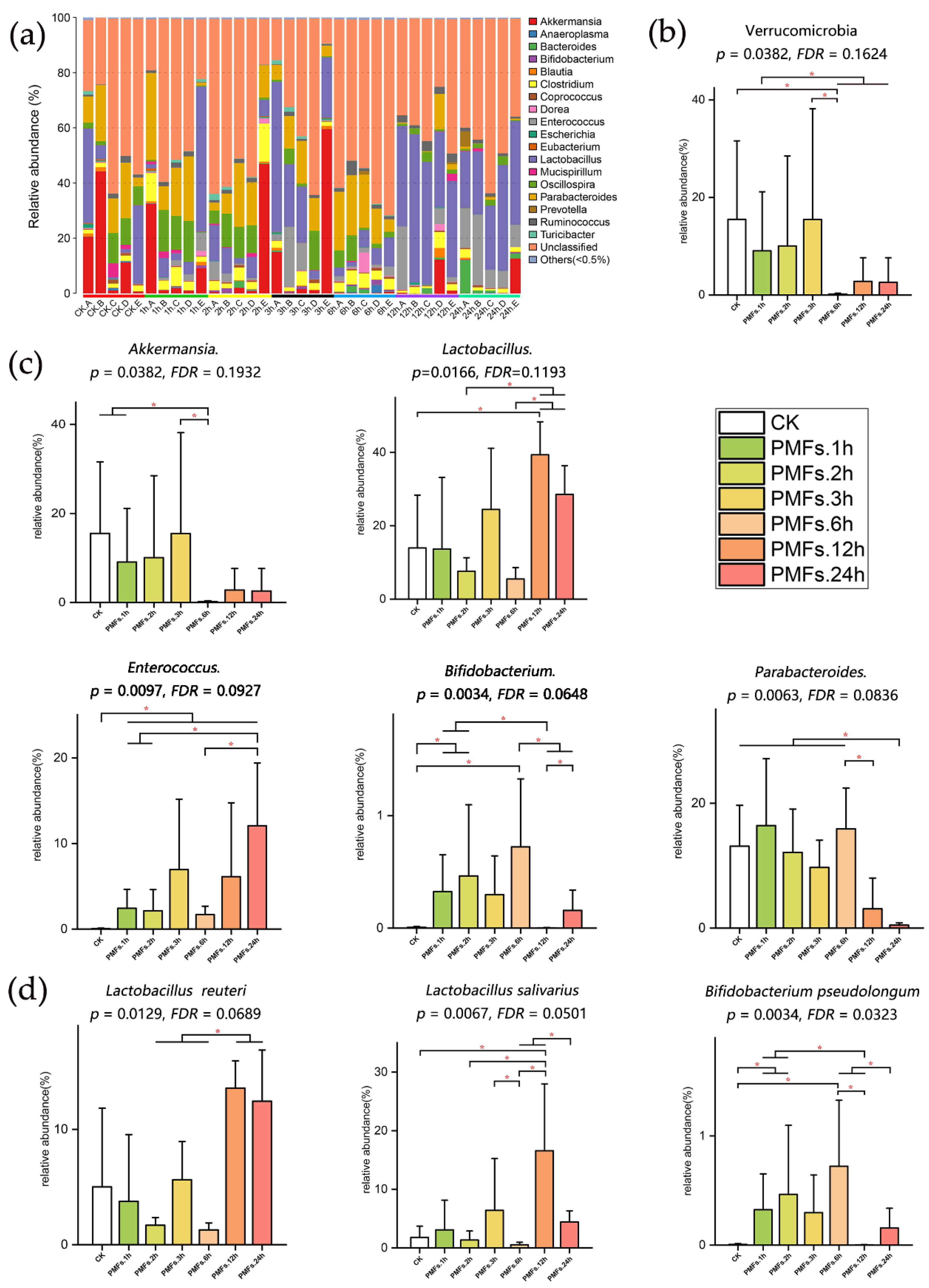
3.3. Effects of PMF-Rich Fraction on Gut Microbiota
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hansen, N.W.; Sams, A. The Microbiotic Highway to Health-New Perspective on Food Structure, Gut Microbiota, and Host Inflammation. Nutrients 2018, 10, 1590. [Google Scholar] [CrossRef] [Green Version]
- Ganesan, K.; Chung, S.K.; Vanamala, J.; Xu, B. Causal Relationship between Diet-Induced Gut Microbiota Changes and Diabetes: A Novel Strategy to Transplant Faecalibacterium prausnitzii in Preventing Diabetes. Int. J. Mol. Sci. 2018, 19, 3720. [Google Scholar] [CrossRef] [Green Version]
- Kristek, A.; Schar, M.Y.; Soycan, G.; Alsharif, S.; Kuhnle, G.G.C.; Walton, G.; Spencer, J.P.E. The gut microbiota and cardiovascular health benefits: A focus on wholegrain oats. Nutr. Bull. 2018, 43, 358–373. [Google Scholar] [CrossRef] [Green Version]
- Zopf, Y.; Reljic, D.; Dieterich, W. Dietary Effects on Microbiota-New Trends with Gluten-Free or Paleo Diet. Med. Sci. 2018, 6, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, C.; Zhao, C.; Guven, E.C.; Paoli, P.; Simal--Gandara, J.; Ramkumar, K.M.; Wang, S.; Buleu, F.; Pah, A.; Turi, V. Dietary polyphenols as antidiabetic agents: Advances and opportunities. Food Front. 2020, 1, 18–44. [Google Scholar] [CrossRef] [Green Version]
- Anhe, F.F.; Roy, D.; Pilon, G.; Dudonne, S.; Matamoros, S.; Varin, T.V.; Garofalo, C.; Moine, Q.; Desjardins, Y.; Levy, E.; et al. A polyphenol-rich cranberry extract protects from diet-induced obesity, insulin resistance and intestinal inflammation in association with increased Akkermansia spp. population in the gut microbiota of mice. Gut 2015, 64, 872–883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neyrinck, A.M.; Van Hee, V.F.; Bindels, L.B.; De Backer, F.; Cani, P.D.; Delzenne, N.M. Polyphenol-rich extract of pomegranate peel alleviates tissue inflammation and hypercholesterolaemia in high-fat diet-induced obese mice: Potential implication of the gut microbiota. Br. J. Nutr. 2013, 109, 802–809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molan, A.-L.; Liu, Z.; Kruger, M. The ability of blackcurrant extracts to positively modulate key markers of gastrointestinal function in rats. World J. Microbiol. Biotechnol. 2010, 26, 1735–1743. [Google Scholar] [CrossRef]
- Sembries, S.; Dongowski, G.; Mehrlaender, K.; Will, F.; Dietrich, H. Physiological effects of extraction juices from apple, grape, and red beet pomaces in rats. J. Agric. Food. Chem. 2006, 54, 10269–10280. [Google Scholar] [CrossRef]
- Molan, A.L.; Lila, M.A.; Mawson, J.; De, S. In vitro and in vivo evaluation of the prebiotic activity of water-soluble blueberry extracts. World J. Microbiol. Biotechnol. 2009, 25, 1243–1249. [Google Scholar] [CrossRef]
- Yanhui, H.; Hang, X. Whole food-based approaches to modulating gut microbiota and associated diseases. Annu. Rev. Food Sci. Technol. 2020, 11, 119–143. [Google Scholar]
- Peterson, J.; Dwyer, J. Flavonoids: Dietary occurrence and biochemical activity. Nutr. Res. 1998, 18, 1995–2018. [Google Scholar] [CrossRef]
- Najmanová, I.; Vopršalová, M.; Saso, L.; Mladěnka, P. The pharmacokinetics of flavanones. Crit. Rev. Food Sci. Nutr. 2019, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Courts, F.L.; Williamson, G. The Occurrence, Fate and Biological Activities of C-glycosyl Flavonoids in the Human Diet. Crit. Rev. Food Sci. Nutr. 2015, 55, 1352–1367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benavente-García, O.; Castillo, J.; Marin, F.R.; Ortuño, A.; Del Río, J.A. Uses and Properties of Citrus Flavonoids. J. Agric. Food. Chem. 1997, 45, 4505–4515. [Google Scholar] [CrossRef]
- Wang, Y.; Qian, J.; Cao, J.; Wang, D.; Liu, C.; Yang, R.; Li, X.; Sun, C. Antioxidant Capacity, Anticancer Ability and Flavonoids Composition of 35 Citrus (Citrus reticulata Blanco) Varieties. Molecules 2017, 22, 1114. [Google Scholar] [CrossRef] [PubMed]
- Stevens, Y.; Van Rymenant, E.; Grootaert, C.; Van Camp, J.; Possemiers, S.; Masclee, A.; Jonkers, D. The Intestinal Fate of Citrus Flavanones and Their Effects on Gastrointestinal Health. Nutrients 2019, 11, 1464. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Wang, H.; Guo, L.; Zhao, H.; Ho, C.-T. Chemistry and bioactivity of nobiletin and its metabolites. J. Funct. Foods 2014, 6, 2–10. [Google Scholar] [CrossRef]
- Choi, S.-Y.; Ko, H.-C.; Ko, S.-Y.; Hwang, J.-H.; Park, J.-G.; Kang, S.-H.; Han, S.-H.; Yun, S.-H.; Kim, S.-J. Correlation between flavonoid content and the NO production inhibitory activity of peel extracts from various citrus fruits. Biol. Pharm. Bull. 2007, 30, 772–778. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.; Cao, A.; Shi, J.; Yin, P.; Wang, L.; Ji, G.; Xie, J.; Wu, D. Tangeretin, a citrus polymethoxyflavonoid, induces apoptosis of human gastric cancer AGS cells through extrinsic and intrinsic signaling pathways. Oncol. Rep. 2014, 31, 1788–1794. [Google Scholar] [CrossRef] [Green Version]
- Shu, Z.; Yang, B.; Zhao, H.; Xu, B.; Jiao, W.; Wang, Q.; Wang, Z.; Kuang, H. Tangeretin exerts anti-neuroinflammatory effects via NF-kappa B modulation in lipopolysaccharide-stimulated microglial cells. Int. Immunopharmacol. 2014, 19, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Mulvihill, E.E.; Assini, J.M.; Lee, J.K.; Allister, E.M.; Sutherland, B.G.; Koppes, J.B.; Sawyez, C.G.; Edwards, J.Y.; Telford, D.E.; Charbonneau, A.; et al. Nobiletin Attenuates VLDL Overproduction, Dyslipidemia, and Atherosclerosis in Mice With Diet-Induced Insulin Resistance. Diabetes 2011, 60, 1446–1457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Z.; Gao, W.; Zeng, S.-L.; Li, P.; Liu, E.H. Chemical structures, bioactivities and molecular mechanisms of citrus polymethoxyflavones. J. Funct. Foods 2018, 40, 498–509. [Google Scholar] [CrossRef]
- Li, S.; Sang, S.; Pan, M.-H.; Lai, C.-S.; Lo, C.-Y.; Yang, C.S.; Ho, C.-T. Anti-inflammatory property of the urinary metabolites of nobiletin in mouse. Bioorg. Med. Chem. Lett. 2007, 17, 5177–5181. [Google Scholar] [CrossRef]
- Kim, M.; Kim, N.; Han, J. Metabolism of Kaempferia parviflora Polymethoxyflavones by Human Intestinal Bacterium Bautia sp MRG-PMF1. J. Agric. Food. Chem. 2014, 62, 12377–12383. [Google Scholar] [CrossRef]
- Wang, Y.; Zang, W.; Ji, S.; Cao, J.; Sun, C. Three Polymethoxyflavones Purified from Ougan (Citrus reticulata cv. Suavissima) Inhibited LPS-Induced NO Elevation in the Neuroglia BV-2 Cell Line via the JAK2/STAT3 Pathway. Nutrients 2019, 11, 791. [Google Scholar] [CrossRef] [Green Version]
- Goncalves, D.R.; Manthey, J.A.; da Costa, P.I.; Rodrigues, M.C.M.; Cesar, T.B. Analysis of Fluorescence Spectra of Citrus Polymethoxylated Flavones and Their Incorporation into Mammalian Cells. J. Agric. Food. Chem. 2018, 66, 7531–7541. [Google Scholar] [CrossRef]
- Zheng, J.; Bi, J.; Johnson, D.; Sun, Y.; Song, M.; Qiu, P.; Dong, P.; Decker, E.; Xiao, H. Analysis of 10 Metabolites of Polymethoxyflavones with High Sensitivity by Electrochemical Detection in High-Performance Liquid Chromatography. J. Agric. Food. Chem. 2015, 63, 509–516. [Google Scholar] [CrossRef]
- Li, S.M.; Wang, Z.Y.; Sang, S.M.; Huang, M.T.; Ho, C.T. Identification of nobiletin metabolites in mouse urine. Mol. Nutr. Food Res. 2006, 50, 291–299. [Google Scholar] [CrossRef]
- Zeng, S.-L.; Duan, L.; Chen, B.-Z.; Li, P.; Liu, E.H. Chemicalome and metabolome profiling of polymethoxylated flavonoids in Citri Reticulatae Pericarpium based on an integrated strategy combining background subtraction and modified mass defect filter in a Microsoft Excel Platform. J. Chromatogr. A 2017, 1508, 106–120. [Google Scholar] [CrossRef]
- Tung, Y.-C.; Chang, W.-T.; Li, S.; Wu, J.-C.; Badmeav, V.; Ho, C.-T.; Pan, M.-H. Citrus peel extracts attenuated obesity and modulated gut microbiota in mice with high-fat diet-induced obesity. Food Funct. 2018, 9, 3363–3373. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhu, J.; Zhang, X.; Zhao, D.G.; Ma, Y.Y.; Li, D.; Ho, C.T.; Huang, Q. Aged citrus peel (chenpi) extract causes dynamic alteration of colonic microbiota in high-fat diet induced obese mice. Food Funct. 2020, 11, 2667–2678. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.-S.; Song, E.-J.; Nam, Y.-D.; Lee, S.-Y. Probiotics in human health and disease: From nutribiotics to pharmabiotics. J. Microbiol. 2018, 56, 773–782. [Google Scholar] [CrossRef]
- Robles Alonso, V.; Guarner, F. Linking the gut microbiota to human health. Br. J. Nutr. 2013, 109, S21–S26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukuda, S.; Toh, H.; Hase, K.; Oshima, K.; Nakanishi, Y.; Yoshimura, K.; Tobe, T.; Clarke, J.M.; Topping, D.L.; Suzuki, T.; et al. Bifidobacteria can protect from enteropathogenic infection through production of acetate. Nature 2011, 469, U543–U791. [Google Scholar] [CrossRef] [PubMed]
- Levy, M.; Thaiss, C.A.; Elinav, E. Metabolites: Messengers between the microbiota and the immune system. Genes Dev. 2016, 30, 1589–1597. [Google Scholar] [CrossRef]
- An, H.M.; Park, S.Y.; Lee, D.K.; Kim, J.R.; Cha, M.K.; Lee, S.W.; Lim, H.T.; Kim, K.J.; Ha, N.J. Antiobesity and lipid-lowering effects of Bifidobacterium spp. in high fat diet-induced obese rats. Lipids Health Dis. 2011, 10, 116. [Google Scholar] [CrossRef] [Green Version]
- Tuohy, K.M.; Fava, F.; Viola, R. The way to a man’s heart is through his gut microbiota-dietary pro- and prebiotics for the management of cardiovascular risk. Proc. Nutr. Soc. 2014, 73, 172–185. [Google Scholar] [CrossRef]
- Yunes, R.A.; Poluektova, E.U.; Dyachkova, M.S.; Klimina, K.M.; Kovtun, A.S.; Averina, O.V.; Orlova, V.S.; Danilenko, V.N. GABA production and structure of gadB/gadC genes in Lactobacillus and Bifidobacterium strains from human microbiota. Anaerobe 2016, 42, 197–204. [Google Scholar] [CrossRef]
- Hsu, T.-C.; Huang, C.-Y.; Liu, C.-H.; Hsu, K.-C.; Chen, Y.-H.; Tzang, B.-S. Lactobacillus paracasei GMNL-32, Lactobacillus reuteri GMNL-89 and L-reuteri GMNL-263 ameliorate hepatic injuries in lupus-prone mice. Br. J. Nutr. 2017, 117, 1066–1074. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak No. | Retention Time (min) | [M + H]+ or [M − H]− (m/z) | Error (ppm) | Formula | Fragment Ions (m/z) | Tentative Compounds |
|---|---|---|---|---|---|---|
| 1 | 13.4438 | 609.1827 (-) | 0.5 | C28H34O15 | 489, 343, 301, 286, 242 | Neohesperidin |
| 2 | 21.2998 | 593.1876 (-) | 0.0 | C28H34O14 | 285 | Poncirin |
| 3 | 29.1432 | 373.1291 | 2.2 | C20H20O7 | 358, 343, 327, 315, 299, 181, 163, 153 | Isosinensetin |
| 4 | 29.3402 | 389.1231 | 2.6 | C20H20O8 | 374, 359, 344, 331, 313, 298, 287, 211, 183 | Monohydroxy-pentamethoxyflavone |
| 5 | 31.3811 | 373.1282 | 0.9 | C20H20O7 | 357, 343, 329, 312, 297, 153 | Sinensetin |
| 6 | 31.8459 | 343.1176 | 0.8 | C19H18O6 | 328, 313, 285, 257, 181, 153 | Tetramethyl-O-isoscutellarein |
| 7 | 33.4704 | 403.1387 | 0.6 | C21H22O8 | 388, 373, 358, 355, 327, 211, 183 | Nobiletin |
| 8 | 33.853 | 375.1074 | 1.7 | C19H18O8 | 360, 345, 330, 327, 317, 302, 197, 169, 149 | 5,4′-Dihydroxyl-3,7,8,3′-tetramethoxyflavone |
| 9 | 34.0789 | 343.1176 | 2.0 | C19H18O6 | 328, 313, 285, 257, 181, 153 | Tetramethyl-O-scutellarein |
| 10 | 35.9401 | 373.1282 | 0.3 | C20H20O7 | 358, 343, 328, 297, 211, 183 | Tangeretin |
| 11 | 37.8647 | 389.1231 | 0.5 | C20H20O8 | 374, 359, 341, 331, 197 | 5-demethylnobiletin |
| Peak No. | Retention Time (min) | [M + H]+ or [M − H]− (m/z) | Formula | Fragment Ions (m/z) | Tentative Compounds |
|---|---|---|---|---|---|
| 1 | 13.1284 | 609.1825 (-) | C28H34O15 | 489, 343, 325, 301, 286, 257, 242, 164 | Neohesperidin |
| 2 | 16.3972 | 563.1460 (-) | C26H28O14 | 387, 372, 357, 342, 314, 299, 175, 113 | Nobelitin-O-glucuronide (1) |
| 3 | 18.8731 | 535.1449 | C25H26O13 | 359, 344, 329, 314, 311, 301, 298, 286, 283, 257 | Tangeretin-O-glucuronide (1) |
| 4 | 19.5825 | 565.1555 | C26H28O14 | 389, 374, 359, 345, 344, 343, 341, 331, 328, 327, 316, 315, 313 | Nobelitin-O-glucuronide (2) |
| 5 | 19.8712 | 565.1551 | C26H28O14 | 389, 374, 359, 345, 344, 343, 341, 339, 331, 329, 328, 327, 316, 315, 313, 310, 301, 298 | Nobelitin-O-glucuronide (3) |
| 6 | 20.1621 | 375.1078 | C19H18O8 | 360, 345, 330, 327, 317, 302, 299, 197, 169, 149 | Dihydroxy-tetramethoxyflavone (1) |
| 7 | 21.0102 | 287.0917 | C16H14O5 | 161, 153, 135, 133, 125, 121, 118, 111, 103, 97, 69, 67 | Dihydroxy-monomethoxyflavone |
| 8 | 21.5526 | 345.0974 | C18H16O7 | 330, 315, 297, 287, 272, 197, 169 | Dihydroxy-trimethoxyflavone |
| 9 | 22.4321 | 375.1081 | C19H18O8 | 360, 345, 330, 327, 317, 302, 197, 169 | Dihydroxy-tetramethoxyflavone (2) |
| 10 | 22.6898 | 327.0874 (-) | C18H16O6 | 327, 312, 297, 282, 269, 254, 226, 182, 177, 117 | Monohydroxy-trimethoxyflavone |
| 11 | 23.5766 | 535.1449 | C25H26O13 | 359, 344, 329, 311 | Tangeretin-O-glucuronide(2) |
| 12 | 24.0618 | 375.1081 | C19H18O8 | 360, 345, 331, 330, 327, 317, 314, 302, 301, 299, 287, 285, 274, 273, 271, 211, 183, 168, 165, 147, 139, 137, 135, 134, 127 | Dihydroxy-tetramethoxyflavone (3) |
| 13 | 24.2942 | 375.1084 | C19H18O8 | 360, 345, 331, 330, 327, 325, 317, 314, 313, 302, 299, 287, 285, 274, 273, 271, 230, 211, 183, 168, 137, 135, 127 | Dihydroxy-tetramethoxyflavone (4) |
| 14 | 24.6514 | 565.1555 | C26H28O14 | 389, 374, 359, 356, 341 | Nobelitin-O-glucuronide (4) |
| 15 | 25.8876 | 389.1237 | C20H20O8 | 374, 359, 356, 341, 331, 316, 285, 244, 197, 169, 163, 148, 113 | Monohydroxy-pentamethoxyflavone (1) |
| 16 | 26.0809 | 271.0607 | C15H10O5 | 253, 243, 215, 197, 169, 153, 149, 115, 91 | Trihydroxy-flavone |
| 17 | 26.6764 | 359.1135 | C19H18O7 | 344, 343, 341, 329, 327, 325, 315, 301, 300, 298, 297, 283, 272, 255, 227, 181, 153 | Monohydroxy-tetramethoxyflavone (1) |
| 18 | 27.7937 | 389.1238 | C20H20O8 | 374, 359, 344, 343, 341, 331, 316, 197, 169, 165, 163 | Monohydroxy-pentamethoxyflavone (2) |
| 19 | 28.4142 | 359.1128 | C19H18O7 | 344, 329, 315, 314, 311, 309, 301, 298, 297, 286, 285, 283, 271, 268, 258, 257, 255, 240, 230, 228, 215, 214, 212, 211, 200, 193, 187, 183, 168, 165, 139, 131, 127, 121 | Monohydroxy-tetramethoxyflavone (2) |
| 20 | 29.1096 | 389.1234 | C20H20O8 | 374, 359, 345, 344, 343, 341, 339, 331, 328, 326, 316, 315, 313, 310, 301, 299, 288, 287, 285, 270, 260, 245, 230, 217, 211, 193, 183, 168, 151, 127 | Monohydroxy-pentamethoxyflavone (3) |
| 21 | 29.5447 | 359.0772 | C17H14O6 | 344, 329, 314, 311, 301, 286, 283, 258, 242, 230, 214, 202, 193, 177, 174, 133 | Dihydroxy-dimethoxyflavone |
| 22 | 30.7912 | 359.1135 | C19H18O7 | 344, 329, 314, 311, 301, 286, 283, 197, 169, 133 | Monohydroxy-tetramethoxyflavone (3) |
| 23 | 31.4917 | 343.1182 | C19H18O6 | 328, 313, 285, 181, 153, 133 | Tetramethoxyflavone |
| 24 | 33.2004 | 403.1395 | C21H22O8 | 388, 387, 373, 359, 358, 357, 355, 353, 345, 343, 341, 339, 330, 327, 325, 315, 313, 311, 301, 299, 259, 257, 244, 231, 211, 193, 183, 175, 168, 163, 162 | Nobiletin |
| 25 | 35.6699 | 373.1291 | C20H20O7 | 358, 343, 325, 315, 300, 297, 283, 271, 269, 229, 211, 193, 183, 168, 135 | Tangeretin |
| 26 | 37.6439 | 389.1241 | C20H20O8 | 374, 359, 356, 341, 343, 328, 316, 313, 197, 169, 163 | 5-demethylnobiletin |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.; Wang, Y.; Zhu, T.; Yang, S.; Cao, J.; Li, X.; Wang, L.-S.; Sun, C. Beneficial Regulatory Effects of Polymethoxyflavone—Rich Fraction from Ougan (Citrus reticulata cv. Suavissima) Fruit on Gut Microbiota and Identification of Its Intestinal Metabolites in Mice. Antioxidants 2020, 9, 831. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9090831
Chen J, Wang Y, Zhu T, Yang S, Cao J, Li X, Wang L-S, Sun C. Beneficial Regulatory Effects of Polymethoxyflavone—Rich Fraction from Ougan (Citrus reticulata cv. Suavissima) Fruit on Gut Microbiota and Identification of Its Intestinal Metabolites in Mice. Antioxidants. 2020; 9(9):831. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9090831
Chicago/Turabian StyleChen, Jiebiao, Yue Wang, Tailin Zhu, Sijia Yang, Jinping Cao, Xian Li, Li-Shu Wang, and Chongde Sun. 2020. "Beneficial Regulatory Effects of Polymethoxyflavone—Rich Fraction from Ougan (Citrus reticulata cv. Suavissima) Fruit on Gut Microbiota and Identification of Its Intestinal Metabolites in Mice" Antioxidants 9, no. 9: 831. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9090831