MARGRA Lamb Eating Quality and Human Health-Promoting Omega-3 Long-Chain Polyunsaturated Fatty Acid Profiles of Tattykeel Australian White Sheep: Linebreeding and Gender Effects

 , , ,
, , ,  and
and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Animal Ethics
2.2. Animals and Experimental Design
2.3. Muscle Biopsy Sampling Procedure
2.4. Determination of Intramuscular Fat
2.5. Determination of Fat Melting Point
2.6. Determination of Fatty Acid Composition
2.7. Extraction and Purification of Phenolic Compounds
2.8. Antioxidant Enzyme Activities
2.9. Statistical Analysis
3. Results
3.1. Nutrient Composition of the Grazed Ryegrass Pasture, Muscle Phenolics and Antioxidant Enzyme Activities
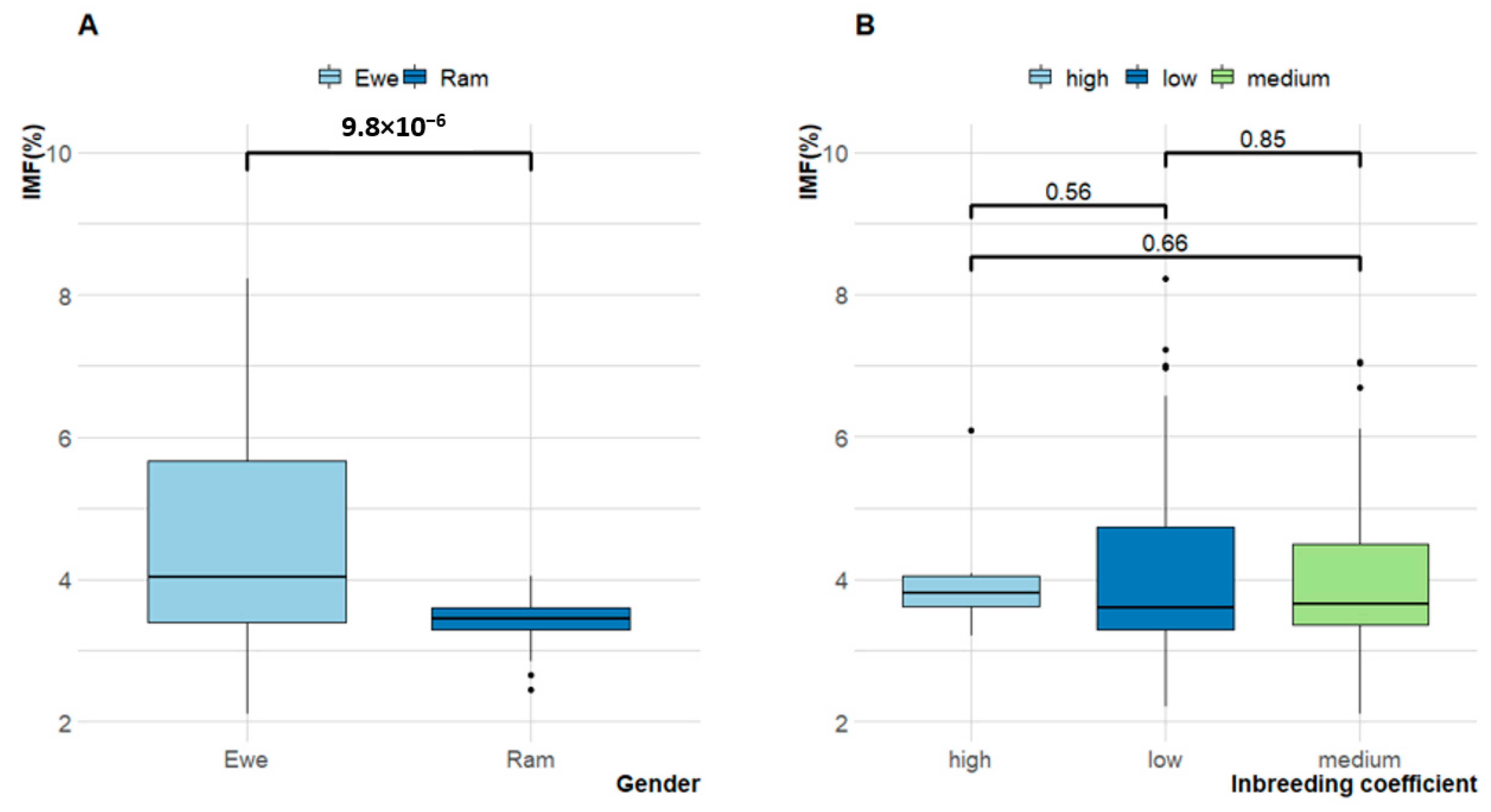
3.2. Intramuscular Fat Content (IMF)
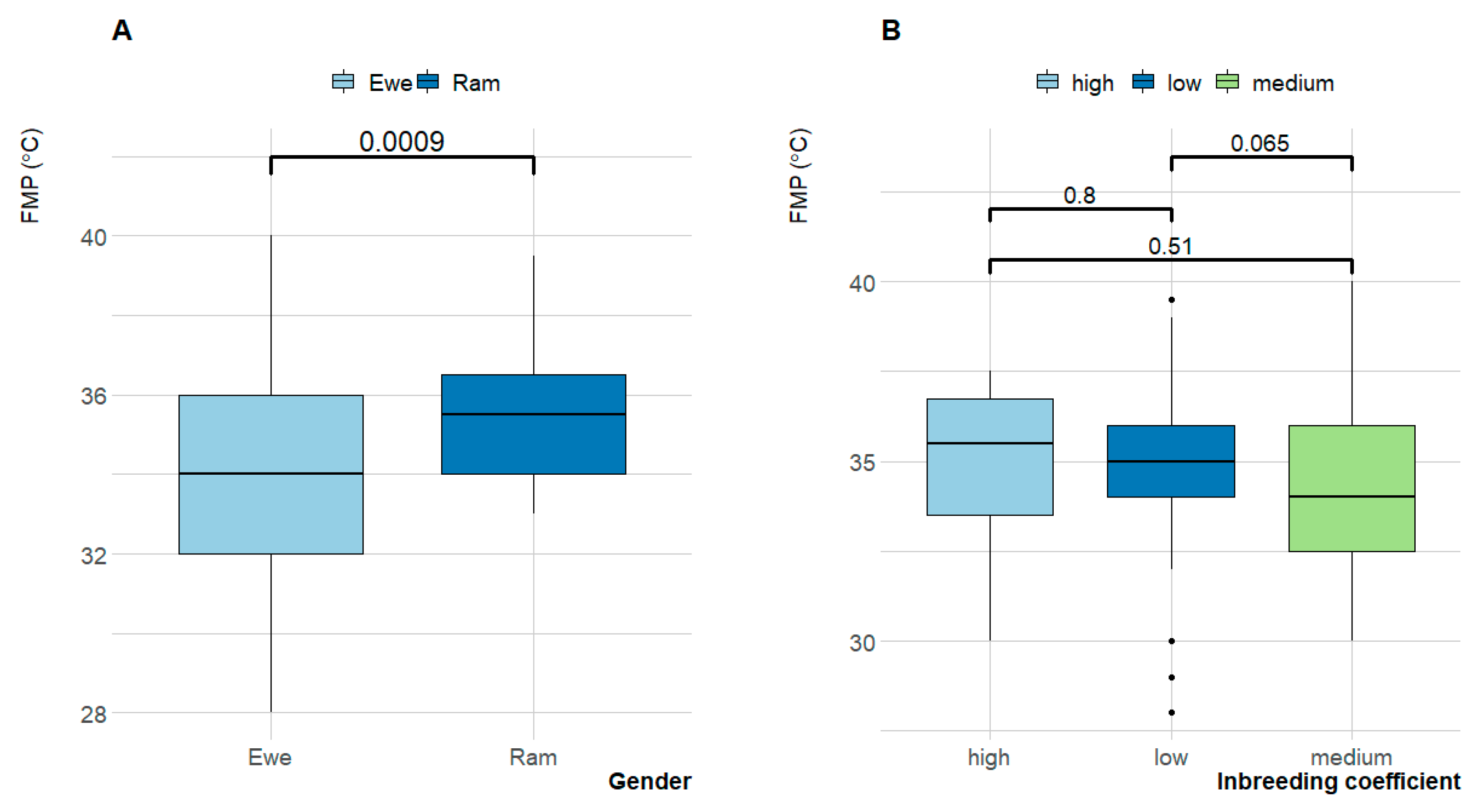
3.3. Fat Melting Point (FMP)
3.4. Faty Acid Composition
4. Discussion
4.1. FMP
4.2. IMF
4.3. Omega-3 Long-Chain Polyunsaturated Fatty Acids
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO. FAO. Food and Agriculture Organization of the United Nations. Meat Quality 2014. Available online: http://www.fao.org/ag/againfo/themes/en/meat/quality_meat (accessed on 3 November 2020).
- MLA. MLA. Meat and Livestock Australia. Eating Quality 2020. Available online: https://www.mla.com.au/research-and-development/feeding-finishing-nutrition/eating-quality (accessed on 3 November 2020).
- Ripoll, G.; Joy, M.; Panea, B. Consumer perception of the quality of lamb and lamb confit. Foods 2018, 7, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elmasry, G.; Barbin, D.F.; Sun, D.-W.; Allen, P. Meat quality evaluation by hyperspectral imaging technique: An overview. Crit. Rev. Food Sci. Nutr. 2012, 52, 689–711. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, A.; Velandia-Valero, M.; Campo, M.M.; Sañudo, C. Some factors that affect ruminant meat quality: From the farm to the fork. Review. Acta Scientiarum. Anim. Sci. 2013, 35, 335–347. [Google Scholar]
- Milewski, S. Health-promoting properties of sheep products. Med. Weter. 2006, 62, 516–519. [Google Scholar]
- Alves, L.G.C.; Osório, J.D.S.; Fernandes, A.; Ricardo, H.D.A.; Cunha, C. Produção de carne ovina com foco no consumidor. Enciclopédia Biosf. 2014, 10, 2399–2415. [Google Scholar]
- Miller, R. Drivers of consumer liking for beef, pork, and lamb: A review. Foods 2020, 9, 428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hocquette, J.-F.; Richardson, R.I.; Prache, S.; Medale, F.; Duffy, G.; Scollan, N.D. The future trends for research on quality and safety of animal products. Ital. J. Anim. Sci. 2005, 4, 49–72. [Google Scholar] [CrossRef]
- Indurain, G.; Carr, T.; Goñi, M.; Insausti, K.; Beriain, M. The relationship of carcass measurements to carcass composition and intramuscular fat in Spanish beef. Meat Sci. 2009, 82, 155–161. [Google Scholar] [CrossRef]
- Hocquette, J.-F.; Minsant, P.; Daudin, J.-D.; Cassar-Malek, I.; Rémond, D.; Doreau, M.; Sans, P.; Bauchart, D.; Verbeke, D.; Picard, B.; et al. Will meat be produced in vitro in the future? INRA Prod. Anim. 2013, 26, 363–374. [Google Scholar] [CrossRef] [Green Version]
- Valdez-Arjona, L.P.; Ramírez-Mella, M. Pumpkin waste as livestock feed: Impact on nutrition and animal health and on quality of meat, milk, and egg. Animals 2019, 9, 769. [Google Scholar] [CrossRef] [Green Version]
- Flakemore, A.R.; McEvoy, P.D.; Balogun, R.O.; Malau-Aduli, B.S.; Nichols, P.; Malau-Aduli, A.E.O. Degummed crude canola oil supplementation affects fat depot melting points in purebred and first-cross Merino sheep. Anim. Vet. Sci. 2014, 2, 75–80. [Google Scholar] [CrossRef] [Green Version]
- Webb, E.C.; O’neill, H. The animal fat paradox and meat quality. Meat Sci. 2008, 80, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.; Enser, M.; Fisher, A.; Nute, G.; Sheard, P.; Richardson, R.; Hughes, S.; Whittington, F. Fat deposition, fatty acid composition and meat quality: A review. Meat Sci. 2008, 78, 343–358. [Google Scholar] [CrossRef] [PubMed]
- McPhee, M.; Hopkins, D.; Pethick, D. Intramuscular fat levels in sheep muscle during growth. Aust. J. Expt. Agric. 2008, 48, 904–909. [Google Scholar] [CrossRef]
- Pannier, L.; Gardner, G.; O′Reilly, R.; Pethick, D. Factors affecting lamb eating quality and the potential for their integration into an MSA sheepmeat grading model. Meat Sci. 2018, 144, 43–52. [Google Scholar] [CrossRef] [Green Version]
- Pannier, L.; Gardner, G.; Pearce, K.; McDonagh, M.; Ball, A.; Jacob, R.; Pethick, D. Associations of sire estimated breeding values and objective meat quality measurements with sensory scores in Australian lamb. Meat Sci. 2014, 96, 1076–1087. [Google Scholar] [CrossRef]
- Thompson, J.M. The effects of marbling on flavour and juiciness scores of cooked beef, after adjusting to a constant tenderness. Aust. J. Expt. Agric. 2004, 44, 645–652. [Google Scholar] [CrossRef]
- Nichols, P.D.; Petrie, J.; Singh, S. Long-chain omega-3 oils–an update on sustainable sources. Nutrients 2010, 2, 572–585. [Google Scholar] [CrossRef] [Green Version]
- Pańka, D.; Piesik, D.; Jeske, M.; Baturo-Cieśniewska, A. Production of phenolics and the emission of volatile organic compounds by perennial ryegrass (Lolium perenne L.)/Neotyphodium lolii association as a response to infection by Fusarium poae. J. Plant Physiol. 2013, 170, 1010–1019. [Google Scholar] [CrossRef]
- Stewart, A.J.; Stewart, R.F. Encyclopedia of Ecology; Sven, E.J., Brian, D.F., Eds.; Elsevier Science: Amsterdam, The Netherlands, 2008; pp. 2682–2689. ISBN 978-0-08-045405-4. [Google Scholar]
- Choi, K.C.; Son, Y.O.; Hwang, J.M.; Kim, B.T.; Chae, M.; Lee, J.C. Antioxidant, anti-inflammatory and anti-septic potential of phenolic acids and flavonoid fractions isolated from Lolium multiflorum. Pharm. Biol. 2017, 55, 611–619. [Google Scholar] [CrossRef] [Green Version]
- Luciano, G.; Monahan, F.J.; Vasta, V.; Biondi, L.; Lanza, M.; Priolo, A. Dietary tannins improve lamb meat colour stability. Meat Sci. 2009, 81, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Luciano, G.; Monahan, F.J.; Vasta, V.; Biondi, L.; Lanza, M.; Priolo, A. Lipid and colour stability of meat from lambs fed fresh herbage or concentrate. Meat Sci. 2009, 82, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Petron, M.J.; Raes, K.; Claeys, E.; Lourenco, M.; Fremaut, D.; De Smet, S. Effect of grazing pastures of different botanical composition on antioxidant enzyme activities and oxidative stability of lamb meat. Meat Sci. 2007, 75, 737–745. [Google Scholar] [CrossRef] [PubMed]
- López-Andrés, P.; Luciano, G.; Vasta, V.; Gibson, T.M.; Scerra, M.; Biondi, L.; Priolo, A.; Mueller-Harvey, I. Antioxidant effects of ryegrass phenolics in lamb liver and plasma. Animal 2014, 8, 51–57. [Google Scholar] [CrossRef] [Green Version]
- O’Reilly, R.A.; Pannier, L.; Gardner, G.E.; Garmyn, A.J.; Luo, H.; Meng, Q.; Miller, M.F.; Pethick, D.W. Influence of demographic factors on sheepmeat sensory scores of American, Australian and Chinese consumers. Foods 2020, 9, 529. [Google Scholar] [CrossRef]
- O’Reilly, R.; Pannier, L.; Gardner, G.; Garmyn, A.; Luo, H.; Meng, Q.; Miller, M.; Pethick, D. Minor differences in perceived sheepmeat eating quality scores of Australian, Chinese and American consumers. Meat Sci. 2020, 164, 108060. [Google Scholar] [CrossRef]
- Pewan, S.B.; Otto, J.R.; Huerlimann, R.; Budd, A.M.; Mwangi, F.W.; Edmunds, R.C.; Holman, B.W.B.; Henry, M.L.E.; Kinobe, R.T.; Adegboye, O.A.; et al. Genetics of omega-3 long-chain polyunsaturated fatty acid metabolism and meat eating quality in Tattykeel Australian White lambs. Genes 2020, 11, 587. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, H.; Liu, M.; Zhao, X.; Luo, H. Effect of breed on the volatile compound precursors and odor profile attributes of lamb meat. Foods 2020, 9, 1178. [Google Scholar] [CrossRef]
- Souza, D.; Selaive-Villarroel, A.; Pereira, E.; Osório, J.; Teixeira, A. Growth performance, feed efficiency and carcass characteristics of lambs produced from Dorper sheep crossed with Santa Inês or Brazilian Somali sheep. Small Rum. Res. 2013, 114, 51–55. [Google Scholar] [CrossRef]
- Monaco, C.A.; Freire, M.T.A.; Melo, L.; Rosa, A.F.; Carrer, C.D.C.; Trindade, M.A. Eating quality of meat from six lamb breed types raised in Brazil. J. Sci. Food Agric. 2015, 95, 1747–1752. [Google Scholar] [CrossRef]
- De Vargas-Junior, F.; Martins, C.F.; Feijó, G.L.D.; Teixeira, A.; Leonardo, A.P.; Ricardo, H.D.A.; Fernandes, A.R.M.; Reis, F.A. Evaluation of genotype on fatty acid profile and sensory of meat of indigenous Pantaneiro sheep and Texel or Santa Inês crossbred finished on feedlot. Small Rum. Res. 2019, 173, 17–22. [Google Scholar] [CrossRef] [Green Version]
- Monteschio, J.D.O.; Burin, P.C.; Leonardo, A.P.; Fausto, D.A.; Silva, A.L.A.; Ricardo, H.A.; Da Silva, M.C.; De Souza, M.R.; Junior, F.M.D.V. Different physiological stages and breeding systems related to the variability of meat quality of indigenous Pantaneiro sheep. PLoS ONE 2018, 13, e0191668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffman, L.C.; Claasen, B.; Van der Merwe, D.A.; Cloete, S.W.P.; Cloete, J.J.E. The effects of production system and sex on the sensory quality characteristics of Dorper lamb. Foods 2020, 9, 725. [Google Scholar] [CrossRef] [PubMed]
- Nuernberg, K.; Fischer, A.; Nuernberg, G.; Ender, K.; Dannenberger, D. Meat quality and fatty acid composition of lipids in muscle and fatty tissue of Skudde lambs fed grass versus concentrate. Small Rum. Res. 2008, 74, 279–283. [Google Scholar] [CrossRef]
- Van der Merwe, D.A.; Brand, T.S.; Hoffman, L.C. Slaughter characteristics of feedlot-finished premium South African lamb: Effects of sex and breed type. Foods 2020, 9, 648. [Google Scholar] [CrossRef] [PubMed]
- Vnučec, I.; Držaić, V.; Mioč, B.; Prpić, Z.; Antunović, Z.; Kegalj, A. Effect of sex on meat chemical composition and fatty acid composition in suckling Pag sheep lambs. Veterinarski. Arhiv. 2016, 86, 217–227. [Google Scholar]
- Bath, D.L.; Marble, V.L. Testing Alfalfa Hay for Its Feeding Value; Leaflet 21437 WREP 109; Division of Agriculture & Natural Resources University of California: Oakland, CA, USA, 1989. [Google Scholar]
- Robinson, P.H.; Givens, D.I.; Getachew, G. Evaluation of NRC, UC Davis and ADAS approaches to estimate the metabolizable energy values of feeds at maintenance energy intake from equations utilizing chemical assays and in vitro determinations. Anim. Feed Sci. Tech. 2004, 114, 75–90. [Google Scholar] [CrossRef]
- Malau-Aduli, A.E.O.; Siebert, B.D.; Bottema, C.D.K.; Pitchford, W.S. Breed comparison of the fatty acid composition of muscle phospholipids in Jersey and Limousin cattle. J. Anim. Sci. 1998, 76, 766–773. [Google Scholar] [CrossRef] [Green Version]
- Holman, B.; Flakemore, A.; Kashan, I.A.; Malau-Aduli, A. Spirulina supplementation, sire breed, sex and basal diet effects on lamb intramuscular fat percentage and fat melting points. Int. J. Vet. Med. 2014, 2014, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Flakemore, A.R.; Balogun, R.O.; McEvoy, P.D.; Malau-Aduli, B.S.; Nichols, P.; Malau-Aduli, A.E.O. Genetic variation in intramuscular fat of prime lambs supplemented with varying concentrations of degummed crude canola oil. Int. J. Nutr. Food Sci. 2014, 3, 203–209. [Google Scholar] [CrossRef]
- Malau-Aduli, A.; Holman, B.; Kashani, A.; Nichols, P. Sire breed and sex effects on the fatty acid composition and content of heart, kidney, liver, adipose and muscle tissues of purebred and first-cross prime lambs. Anim. Prod. Sci. 2016, 56, 2122–2132. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Phys. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, M.R.; Nichols, P.D.; Barnes, J.; Davies, N.W.; Peacock, E.J.; Carter, C.G. Regiospecificity profiles of storage and membrane lipids from the gill and muscle tissue of atlantic salmon (Salmo salar L.) grown at elevated temperature. Lipids 2006, 41, 865–876. [Google Scholar] [CrossRef] [PubMed]
- Clayton, E. Graham Centre Monograph no. 4: Long-Chain Omega-3 Polyunsaturated Fatty Acids in Ruminant Nutrition: Benefits to Animals and Humans; Nugent, T., Nicholls, C., Eds.; Charles Sturt University: Wagga-Wagga, Australia, 2014. [Google Scholar]
- SAS. Statistical Analysis System. SAS/STAT User’s Guide: Statistics; Version 9.4.; SAS Inc.: Cary, NC, USA, 2013. [Google Scholar]
- Font-i-Furnols, M.; Guerrero, L. Consumer preference, behavior and perception about meat and meat products: An overview. Meat Sci. 2014, 98, 361–371. [Google Scholar] [CrossRef]
- de Andrade, J.C.; de Aguiar Sobral, L.; Ares, G.; Deliza, R. Understanding consumers’ perception of lamb meat using free word association. Meat Sci. 2016, 117, 68–74. [Google Scholar] [CrossRef]
- Sañudo, C.; Alfonso, M.; Sanchez, A.; Berge, P.; Dransfield, E.; Zygoyiannis, D.; Stamataris, C.; Thorkelsson, G.; Valdimarsdottir, T.; Piasentier, E.; et al. Meat texture of lambs from different European production systems. Aust. J. Agric. Res. 2003, 54, 551–560. [Google Scholar] [CrossRef]
- Young, O.A.; Lane, G.A.; Priolo, A.; Fraser, K. Pastoral and species flavour in lambs raised on pasture, lucerne or maize. J. Sci. Food Agric. 2003, 83, 93–104. [Google Scholar] [CrossRef]
- Young, O.A.; Berdagué, J.L.; Viallon, C.; Rousset-Akrim, S.; Theriez, M. Fat-borne volatiles and sheepmeat odour. Meat Sci. 1997, 45, 183–200. [Google Scholar] [CrossRef]
- Young, O.A.; Lane, G.A.; Podmore, C.; Fraser, K.; Agnew, M.J.; Cummings, T.L.; Cox, N.R. Changes in composition and quality characteristics of ovine meat and fat from castrates and rams aged to 2 years. N. Z. J. Agric. Res. 2006, 49, 419–430. [Google Scholar] [CrossRef]
- Priolo, A.; Micol, D.; Agabriel, J. Effects of grass feeding systems on ruminant meat colour and flavour. A review. Anim. Res. 2001, 50, 185–200. [Google Scholar] [CrossRef]
- Watkins, P.J.; Frank, D.; Singh, T.K.; Young, O.A.; Warner, R.D. Sheepmeat flavor and the effect of different feeding systems: A review. J. Agric. Food Chem. 2013, 61, 3561–3579. [Google Scholar] [CrossRef] [PubMed]
- Watkins, P.J.; Kearney, G.; Rose, G.; Allen, D.; Ball, A.J.; Pethick, D.W.; Warner, R.D. Effect of branched-chain fatty acids, 3-methylindole and 4-methylphenol on consumer sensory scores of grilled lamb meat. Meat Sci. 2014, 96, 1088–1094. [Google Scholar] [CrossRef] [PubMed]
- Watkins, P.J.; Rose, G.; Salvatore, L.; Allen, D.; Tucman, D.; Warner, R.D.; Dunshea, F.R.; Pethick, D.W. Age and nutrition influence the concentrations of three branched chain fatty acids in sheep fat from Australian abattoirs. Meat Sci. 2010, 86, 594–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bekhit, A.E.D.A.; Hopkins, D.L.; Fahri, F.T.; Ponnampalam, E.N. Oxidative processes in muscle systems and fresh meat: Sources, markers, and remedies. Compr. Rev. Food Sci. Food Saf. 2013, 12, 565–597. [Google Scholar] [CrossRef]
- Daley, C.A.; Abbott, A.; Doyle, P.S.; Nader, G.A.; Larson, S. A review of fatty acid profiles and antioxidant content in grass-fed and grain-fed beef. Nutr. J. 2010, 9, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Elswyk, M.E.; McNeill, S.H. Impact of grass/forage feeding versus grain finishing on beef nutrients and sensory quality: The US experience. Meat Sci. 2014, 96, 535–540. [Google Scholar] [CrossRef]
- Zervas, G.; Tsiplakou, E. The effect of feeding systems on the characteristics of products from small ruminants. Small Rum. Res. 2011, 101, 140–149. [Google Scholar] [CrossRef]
- Baldi, G.; Chauhan, S.S.; Linden, N.; Dunshea, F.R.; Hopkins, D.L.; Sgoifo Rossi, C.A.; Dell’Orto, V.; Ponnampalam, E.N. Comparison of a grain-based diet supplemented with synthetic vitamin E versus a lucerne (alfalfa) hay-based diet fed to lambs in terms of carcass traits, muscle vitamin E, fatty acid content, lipid oxidation, and retail colour of meat. Meat Sci. 2019, 148, 105–112. [Google Scholar] [CrossRef]
- Malau-Aduli, A.E.O.; Kashani, A. Molecular genetics-nutrition interactions in the expression of AANAT, ADRB3, BTG2 and FASN genes in the heart, kidney and liver of Australian lambs supplemented with Spirulina (Arthrospira platensis). Genes Genom. 2015, 37, 633–644. [Google Scholar] [CrossRef]
- Smith, S.B.; Yang, A.; Larsen, T.W.; Tume, R.K. Positional analysis of triacylglycerols from bovine adipose tissue lipids varying in degree of unsaturation. Lipids 1998, 33, 197–207. [Google Scholar] [CrossRef]
- Cafferky, J.; Hamill, R.M.; Allen, P.; O′Doherty, J.V.; Cromie, A.; Sweeney, T. Effect of breed and gender on meat quality of M. longissimus thoracis et lumborum muscle from crossbred beef bulls and steers. Foods 2019, 8, 173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venkata Reddy, B.; Sivakumar, A.S.; Jeong, D.W.; Woo, Y.B.; Park, S.J.; Lee, S.Y.; Hwang, I. Beef quality traits of heifer in comparison with steer, bull and cow at various feeding environments. Anim. Sci. J. 2015, 86, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, D.L.; Mortimer, S.I. Effect of genotype, gender and age on sheep meat quality and a case study illustrating integration of knowledge. Meat Sci. 2014, 98, 544–555. [Google Scholar] [CrossRef] [PubMed]
- Pannier, L.; Pethick, D.; Geesink, G.; Ball, A.; Jacob, R.; Gardner, G. Intramuscular fat in the longissimus muscle is reduced in lambs from sires selected for leanness. Meat Sci. 2014, 96, 1068–1075. [Google Scholar] [CrossRef]
- Pannier, L.; Pethick, D.; Geesink, G.; Ball, A.; Jacob, R.; Gardner, G. Intramuscular fatty acid profile of feedlot lambs fed concentrates with alternative ingredients. Anim. Prod. Sci. 2019, 59, 914–920. [Google Scholar]
- McPhee, M.; Oltjen, J.; Fadel, J.; Mayer, D.; Sainz, R. Parameter estimation and sensitivity analysis of fat deposition models in beef steers using acsIXtreme. Math. Comput. Simul. 2009, 79, 2701–2712. [Google Scholar] [CrossRef]
- Okeudo, N.; Moss, B. Intramuscular lipid and fatty acid profile of sheep comprising four sex-types and seven slaughter weights produced following commercial procedure. Meat Sci. 2007, 76, 195–200. [Google Scholar] [CrossRef]
- Lee, H.K.; Lee, J.K.; Cho, B. The role of androgen in the adipose tissue of males. World J. Mens. Health 2013, 31, 136. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; De Pergola, G.; Björntorp, P. The effects of androgens on the regulation of lipolysis in adipose precursor cells. Endocrinol 1990, 126, 1229–1234. [Google Scholar] [CrossRef]
- Bong, J.J.; Jeong, J.Y.; Rajasekar, P.; Cho, Y.M.; Kwon, E.G.; Kim, H.C.; Baik, M. Differential expression of genes associated with lipid metabolism in longissimus dorsi of Korean bulls and steers. Meat Sci. 2012, 91, 284–293. [Google Scholar] [CrossRef]
- Bong, J.J.; Jeong, J.Y.; Rajasekar, P.; Cho, Y.M.; Kwon, E.G.; Kim, H.C.; Paek, B.H.; Baik, M. Impact of inbreeding on milk fatty acids of a Brazilian Holstein cattle. Anim. Prod. Sci. 2020, 60, 1482–1490. [Google Scholar]
- Zárate, R.; El Jaber-Vazdekis, N.; Tejera, N.; Pérez, J.; Rodriguez, C. Significance of long chain polyunsaturated fatty acids in human health. Clin. Transl. Med. 2017, 6, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gould, J.F.; Smithers, L.G.; Makrides, M. The effect of maternal omega-3 (n-3) LCPUFA supplementation during pregnancy on early childhood cognitive and visual development: A systematic review and meta-analysis of randomized controlled trials. Am. J. Clin. Nutr. 2013, 97, 531–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le, H.V.; Nguyen, D.V.; Nguyen, Q.V.; Malau-Aduli, B.S.; Nichols, P.D.; Malau-Aduli, A.E.O. Fatty acid profiles of muscle, liver, heart and kidney of Australian prime lambs fed different polyunsaturated fatty acids enriched pellets in a feedlot system. Sci. Rep. 2019, 9, 1–11. [Google Scholar]
- Lourenço, M.; Van Ranst, G.; Vlaeminck, B.; De Smet, S.; Fievez, V. Influence of different dietary forages on the fatty acid composition of rumen digesta as well as ruminant meat and milk. Anim. Feed Sci. Technol. 2008, 145, 418–437. [Google Scholar] [CrossRef]
- Ponnampalam, E.N.; Butler, K.L.; Jacob, R.H.; Pethick, D.W.; Ball, A.J.; Hocking-Edward, J.E.; Geesink, G.; Hopkins, D.L. Health beneficial long chain omega-3 fatty acid levels in Australian lamb managed under extensive finishing systems. Meat Sci. 2014, 96, 1104–1110. [Google Scholar] [CrossRef]
- Ponnampalam, E.N.; Dunshea, F.R.; Warner, R.D. Use of lucerne hay in ruminant feeds to improve animal productivity, meat nutritional value and meat preservation under a more variable climate. Meat Sci. 2020, 170, 108235. [Google Scholar] [CrossRef]
- Ponnampalam, E.N.; Pearce, K.M.; Mortimer, S.I.; Pethick, D.W.; Ball, A.J.; Hopkins, D.L. Sources of variation of health claimable long chain omega-3 fatty acids in meat from Australian lamb slaughtered at similar weights. Meat Sci. 2014, 96, 1095–1103. [Google Scholar] [CrossRef]
- Ponnampalam, E.N.; Sinclair, A.J.; Egan, E.R.; Ferrier, G.R.; Leury, B.J. Dietary manipulation of muscle long-chain omega-3 and omega-6 fatty acids and sensory properties of lamb meat. Meat Sci. 2002, 60, 125–132. [Google Scholar] [CrossRef]
- Ponnampalam, E.N.; Trout, G.R.; Sinclair, A.J.; Egan, E.R.; Leury, B.J. Comparison of the color stability and lipid oxidative stability of fresh and vacuum packaged lamb muscle containing elevated omega-3 and omega-6 fatty acid levels from dietary manipulation. Meat Sci. 2001, 58, 151–161. [Google Scholar] [CrossRef]
- Flakemore, A.R.; Malau-Aduli, B.S.; Nichols, P.D.; Malau-Aduli, A.E.O. Omega-3 fatty acids, nutrient retention values, and sensory meat eating quality in cooked and raw Australian lamb. Meat Sci. 2017, 123, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Knight, M.I.; Daetwyler, H.D.; Hayes, B.J.; Hayden, M.J.; Ball, A.J.; Pethick, D.W.; McDonagh, M.B. An independent validation association study of carcass quality, shear force, intramuscular fat percentage and omega-3 polyunsaturated fatty acid content with gene markers in Australian lamb. Meat Sci. 2014, 96, 1025–1033. [Google Scholar] [CrossRef] [PubMed]
- Knight, M.I.; Butler, K.L.; Linden, N.P.; Burnett, V.F.; Ball, A.J.; McDonagh, M.B.; Behrendt, R. Understanding the impact of sire lean meat yield breeding value on carcass composition, meat quality, nutrient and mineral content of Australian lamb. Meat Sci. 2020, 170, 108236. [Google Scholar] [CrossRef] [PubMed]
- De Brito, G.F.; Holman, B.W.B.; McGrath, S.R.; Friend, M.A.; van de Ven, R.; Hopkins, D.L. The effect of forage-types on the fatty acid profile, lipid and protein oxidation and retail colour stability of muscles from White Dorper lambs. Meat Sci. 2017, 130, 81–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fowler, S.M.; Morris, S.; Hopkins, D.L. Nutritional composition of lamb retail cuts from the carcasses of extensively finished lambs. Meat Sci. 2019, 154, 126–132. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutrient | Composition (% DM) |
|---|---|
| DM | 20.7 |
| CP | 19.0 |
| ADF | 26.5 |
| NDF | 30.9 |
| EE | 1.8 |
| Ash | 6.8 |
| %TDN | 62.5 |
| DE (Mcal/kg) | 2.8 |
| ME (MJ/kg) | 9.4 |
| Phenolic Antioxidants: | |
| FCTP (mg GAE/g) | 1.631 |
| FRAP (mmol Fe2+ E/g) | 6.572 |
| Fatty Acid | % Total Fatty Acids |
|---|---|
| 14:0 | 0.6 |
| 15:0 | 0.2 |
| 16:1n-9c | 0.0 |
| 16:1n-7c | 0.2 |
| 16:0 | 15.7 |
| 17:0 | 0.5 |
| 18:2n-6 LA | 14.8 |
| 18:3n-3 ALA | 57.6 |
| 18:1n-9c | 1.0 |
| 18:1n-7c | 0.2 |
| 18:1n-7t | 0.0 |
| 18:0 | 0.1 |
| 20:4n-6 ARA | 0.0 |
| 20:5n-3 EPA | 0.0 |
| 20:3n-6 | 0.1 |
| 20:4n-3 | 0.1 |
| 20:2n-6 | 0.1 |
| 20:0 | 1.6 |
| 22:5n-6 DPA-6 | 0.0 |
| 22:6n-3 DHA | 0.0 |
| 22:5n-3 DPA-3 | 0.0 |
| 22:0 | 1.0 |
| 23:0 | 0.3 |
| 24:0 | 0.9 |
| ∑SFA | 20.9 |
| ∑MUFA | 4.9 |
| ∑PUFA | 73.1 |
| ∑n-3 LC-PUFA | 0.1 |
| ∑n-3 PUFA | 58.0 |
| ∑n-6 PUFA | 15.2 |
| ∑other FA | 1.0 |
| n-6/n-3 | 0.3 |
| Variable | Ram (n = 47) | Ewe (n = 100) | Overall (n = 147) | P-Value |
|---|---|---|---|---|
| Fat Melting Point (°C) | 35.5 ± 1.5 | 34.2 ± 2.4 | 34.6 ± 2.3 | 0.0001 |
| Intramuscular fat (%) | 3.4 ± 0.3 | 4.4 ± 1.4 | 4.1 ± 1.3 | 0.0001 |
| FCTP (mg GAE/g) | 1.142 ± 0.0036 | 1.171 ± 0.0042 | 1.156 ± 0.0039 | 0.4723 |
| FRAP (mmol Fe 2+ E/g) | 5.481 ± 0.0172 | 5.605 ± 0.0198 | 5.543 ± 0.0185 | 0.2982 |
| GSH-Px (U/g) | 0.085 ± 0.0012 | 0.091 ± 0.0024 | 0.088 ± 0.0018 | 0.0921 |
| Cat (U/g) | 39.8 ± 1.3 | 40.1 ± 1.5 | 40.0 ± 1.4 | 0.0882 |
| SOD (U/g) | 63.8 ± 5.7 | 64.9 ± 6.1 | 64.4 ± 5.9 | 0.1566 |
| Fatty Acids (mg /100 g) | ||||
| C12:0 | 0.1 ± 0.5 | 0.0 ± 0.0 | 0.0 ± 0.3 | 0.1453 |
| C13:0 | 3.2 ± 5.4 | 0.8 ± 3.2 | 1.5 ± 4.2 | 0.0009 |
| C14:0 | 438.7 ± 492.2 | 153.8 ± 291.9 | 244.9 ± 389.7 | 0.0001 |
| C14:1 | 11.9 ± 15.7 | 3.0 ± 5.8 | 5.8 ± 10.9 | 0.0001 |
| C15:0 | 168.6 ± 172.2 | 46.1 ± 98.0 | 85.3 ± 138.3 | 0.0001 |
| C16:0 | 3321.2 ± 2631.5 | 1093.0 ± 1457.6 | 1805.4 ± 2170.2 | 0.0001 |
| C16:1 | 260.7 ± 237.0 | 94.6 ± 134.1 | 147.7 ± 189.5 | 0.0001 |
| C17:0 | 321.0 ± 307.3 | 109.1 ± 237.7 | 176.8 ± 279.1 | 0.0001 |
| C17:1 | 233.0 ± 247.8 | 75.3 ± 137.5 | 125.7 ± 194.0 | 0.0001 |
| C18.0 | 2692.0 ± 2283.4 | 1016.9 ± 1649.3 | 1552.5 ± 2025.2 | 0.0001 |
| C18:1 | 4942.6 ± 4041.0 | 1920.3 ± 2489.0 | 2886.6 ± 3368.4 | 0.0001 |
| C18:2 n-6 LA | 423.5 ± 266.6 | 125.4 ± 80.0 | 220.7 ± 214.9 | 0.0001 |
| C18:3 n-3 ALA | 262.6 ± 209.0 | 72.5 ± 81.6 | 133.3 ± 161.9 | 0.0001 |
| C18:3 n-6 | 2.1 ± 4.3 | 2.4 ± 6.5 | 2.3 ± 5.9 | 0.8014 |
| C18:4 n-3 | 3.7 ± 8.6 | 1.6 ± 10.1 | 2.3 ± 9.7 | 0.2262 |
| CLA | 117.9 ± 129.7 | 63.7 ± 190.0 | 81.1 ± 174.4 | 0.0788 |
| C19:1 | 39.1 ± 41.4 | 14.3 ± 26.3 | 22.2 ± 33.8 | 0.0001 |
| C20:0 | 20.3 ± 18.4 | 7.3 ± 10.8 | 11.4 ± 15.0 | 0.0001 |
| C20:1 | 26.3 ± 28.4 | 8.1 ± 12.2 | 13.9 ± 20.7 | 0.0001 |
| C20:2 n-6 | 7.9 ± 8.9 | 2.1 ± 3.1 | 4.0 ± 6.2 | 0.0001 |
| C20:3 | 8.4 ± 4.1 | 11.7 ± 12.7 | 10.6 ± 10.8 | 0.0839 |
| C20:3 n-6 | 8.8 ± 4.9 | 6.1 ± 2.1 | 7.0 ± 3.5 | 0.0001 |
| C20:4 n-3 | 4.8 ± 7.3 | 2.1 ± 1.5 | 3.0 ± 4.5 | 0.0005 |
| C20:4 n-6 | 36.4 ± 14.2 | 33.7 ± 8.0 | 34.6 ± 10.4 | 0.1473 |
| C20:5 n-3 (EPA) | 26.0 ± 8.5 | 24.3 ± 5.2 | 24.9 ± 6.5 | 0.1402 |
| C21:0 | 2.0 ± 2.5 | 0.6 ± 1.4 | 1.0 ± 1.9 | 0.0001 |
| C22:0 | 2.8 ± 3.3 | 2.3 ± 1.3 | 2.5 ± 2.2 | 0.1342 |
| C22:1 | 0.4 ± 1.2 | 0.8 ± 1.6 | 0.7 ± 1.5 | 0.1613 |
| C22:4 n-6 | 0.6 ± 1.2 | 1.4 ± 0.5 | 1.2 ± 0.9 | 0.0001 |
| C22:5 n-3 (DPA) | 22.5 ± 11.6 | 25.2 ± 8.0 | 24.4 ± 9.4 | 0.097 |
| C22:5 n-6 | 0.0 ± 0.1 | 0.2 ± 0.2 | 0.1 ± 0.2 | 0.0001 |
| C22:6 n-3(DHA) | 5.8 ± 3.7 | 8.3 ± 2.7 | 7.5 ± 3.2 | 0.0001 |
| C23:0 | 2.5 ± 2.3 | 2.5 ± 0.7 | 2.5 ± 1.4 | 0.7207 |
| C24:0 | 2.2 ± 2.0 | 2.8 ± 0.9 | 2.6 ± 1.4 | 0.008 |
| C24:1 n-9c | 1.7 ± 2.1 | 3.9 ± 1.8 | 3.2 ± 2.1 | 0.0001 |
| EPA+DHA | 31.9 ± 11.3 | 32.6 ± 7.0 | 32.4 ± 8.5 | 0.6265 |
| EPA+DHA+DPA | 54.4 ± 21.8 | 57.9 ± 13.6 | 56.7 ± 16.7 | 0.2388 |
| SFA | 6971.2 ± 5684.7 | 2434.4 ± 3700.9 | 3884.9 ± 4896.6 | 0.0001 |
| MUFA | 2120.3 ± 2772.9 | 5515.7 ± 4577.1 | 3205.9 ± 3786.7 | 0.0001 |
| PUFA | 380.7 ± 331.9 | 931.1 ± 614.7 | 556.7 ± 510.0 | 0.0001 |
| PUFA/SFA | 0.2 ± 0.1 | 0.2 ± 0.1 | 0.2 ± 0.1 | 0.0036 |
| ∑n-3 PUFA | 134.1 ± 96.5 | 325.5 ± 231.7 | 195.3 ± 176.7 | 0.0001 |
| ∑n-6 PUFA | 479.3 ± 279.5 | 171.3 ± 85.2 | 269.8 ± 224.3 | 0.0001 |
| n-6/ n-3 PUFA | 1.6 ± 0.5 | 1.4 ± 0.3 | 1.5 ± 0.4 | 0.0001 |
| Inbreeding Coefficient (%) | ||||
|---|---|---|---|---|
| Variable | Low (0–5) (n = 49) | Medium(6–10) (n = 49) | High (Above 10) (n = 49) | P-Value |
| Fat Melting Points (°C) | 34.9 ± 2.1 | 34.2 ± 2.4 | 34.8 ± 2.8 | 0.225 |
| Intramuscular fat | 4.1 ± 1.4 | 4.0 ± 1.2 | 4.1 ± 1.0 | 0.9148 |
| FCTP (mg GAE/g) | 1.271 ± 0.0014 | 1.269 ± 0.0019 | 1.290 ± 0.0014 | 0.8524 |
| FRAP (mmol Fe 2+ E/g) | 6.018 ± 0.0027 | 6.083 ± 0.0045 | 6.102 ± 0.0086 | 0.2352 |
| GSH-Px (U/g) | 0.091 ± 0.0041 | 0.086 ± 0.0036 | 0.089 ± 0.0062 | 0.0896 |
| Cat (U/g) | 40.5 ± 1.8 | 40.1 ± 1.6 | 40.7 ± 1.9 | 0.1843 |
| SOD (U/g) | 64.8 ± 5.7 | 65.0 ± 5.9 | 64.5 ± 5.3 | 0.0972 |
| Fatty Acids (mg/100 g) | ||||
| C12:0 | 0.0 ± 0.4 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.6757 |
| C13:0 | 2.3 ± 4.6 | 0.7 ± 3.5 | 0.0 ± 0.0 | 0.0574 |
| C14:0 | 340.3 ± 479.9 | 127.5 ± 176.7 | 94.6 ± 50.8 | 0.0033 |
| C14:1 | 7.7 ± 11.9 | 3.6 ± 9.4 | 2.4 ± 1.7 | 0.0609 |
| C15:0 | 116.6 ± 152.0 | 47.7 ± 112.7 | 27.1 ± 22.9 | 0.0074 |
| C16:0 | 2377.1 ± 2509.5 | 1100.6 ± 1410.2 | 923.7 ± 527.9 | 0.0013 |
| C16:1 | 194.1 ± 215.9 | 90.9 ± 135.4 | 71.8 ± 38.3 | 0.0033 |
| C17:0 | 241.4 ± 322.4 | 98.9 ± 193.4 | 61.0 ± 32.9 | 0.006 |
| C17:1 | 165.6 ± 202.9 | 78.3 ± 178.8 | 46.6 ± 26.2 | 0.0174 |
| C18.0 | 2130.6 ± 2447.3 | 840.2 ± 938.8 | 655.2 ± 288.8 | 0.0004 |
| C18:1 | 3704.7 ± 3831.0 | 1885.3 ± 2423.3 | 1552.9 ± 690.9 | 0.0036 |
| C18:2 n-6 (LA) | 271.1 ± 244.2 | 158.9 ± 156.5 | 139.0 ± 65.3 | 0.0053 |
| C18:3 n-3 (ALA) | 170.3 ± 179.0 | 89.0 ± 129.2 | 63.7 ± 33.0 | 0.0067 |
| C18:3 n-6 | 3.1 ± 7.8 | 1.3 ± 0.9 | 1.5 ± 0.8 | 0.032 |
| C18:4 n-3 | 3.1 ± 11.5 | 1.4 ± 7.0 | 0.4 ± 0.3 | 0.5311 |
| CLA | 114.4 ± 219.4 | 40.4 ± 75.8 | 24.9 ± 13.7 | 0.032 |
| C19:1 | 29.4 ± 37.4 | 13.6 ± 27.5 | 9.1 ± 5.2 | 0.0139 |
| C20:0 | 15.5 ± 17.7 | 6.5 ± 8.4 | 5.2 ± 2.7 | 0.001 |
| C20:1 | 18.4 ± 22.3 | 8.7 ± 17.9 | 5.2 ± 2.2 | 0.0124 |
| C20:2 n-6 | 5.3 ± 7.2 | 2.2 ± 4.3 | 2.0 ± 2.1 | 0.0092 |
| C20:3 | 11.0 ± 14.3 | 10.2 ± 2.7 | 9.4 ± 2.7 | 0.8782 |
| C20:3 n-6 | 7.6 ± 3.9 | 6.2 ± 2.7 | 5.6 ± 1.1 | 0.8782 |
| C20:4 n-3 | 3.6 ± 5.3 | 2.3 ± 3.1 | 1.6 ± 0.6 | 0.171 |
| C20:4 n-6 | 32.8 ± 11.6 | 36.6 ± 8.4 | 38.0 ± 8.3 | 0.0737 |
| C20:5 n-3 (EPA) | 24.3 ± 7.0 | 25.8 ± 5.9 | 23.0 ± 4.8 | 0.3091 |
| C21:0 | 1.6 ± 2.3 | 0.4 ± 0.9 | 0.1 ± 0.2 | 0.0009 |
| C22:0 | 3.0 ± 2.7 | 1.8 ± 1.1 | 1.6 ± 0.8 | 0.0055 |
| C22:1 | 0.8 ± 1.9 | 0.6 ± 0.9 | 0.3 ± 0.3 | 0.5598 |
| C22:4 n-6 | 1.2 ± 1.0 | 1.2 ± 0.7 | 1.3 ± 0.7 | 0.9005 |
| C22:5 n-3 (DPA) | 24.7 ± 11.5 | 23.9 ± 5.7 | 23.3 ± 5.0 | 0.8515 |
| C22:5 n-6 | 0.1 ± 0.2 | 0.2 ± 0.2 | 0.2 ± 0.3 | 0.3705 |
| C22:6 n-3(DHA) | 7.4 ± 3.8 | 7.6 ± 2.4 | 7.5 ± 3.1 | 0.9816 |
| C23:0 | 2.7 ± 1.7 | 2.7 ± 1.0 | 2.0 ± 1.1 | 0.2737 |
| C24:0 | 2.7 ± 1.5 | 2.5 ± 1.2 | 2.3 ± 1.3 | 0.674 |
| C24:1 n-9c | 2.9 ± 2.1 | 3.5 ± 2.1 | 3.7 ± 2.1 | 0.2498 |
| EPA+DHA | 31.8 ± 9.5 | 33.4 ± 7.1 | 30.6 ± 7.6 | 0.4708 |
| EPA+DHA+DPA | 56.5 ± 19.8 | 57.3 ± 11.9 | 53.8 ± 12.4 | 0.8738 |
| SFA | 5231.3 ± 5786.8 | 2228.5 ± 2775.3 | 1772.7 ± 913.9 | 0.0007 |
| MUFA | 4123.6 ± 4281.0 | 2084.4 ± 2782.6 | 1691.9 ± 761.3 | 0.0036 |
| PUFA | 680.1 ± 577.2 | 407.1 ± 373.2 | 341.5 ± 105.3 | 0.0037 |
| PUFA/SFA | 0.2 ± 0.1 | 0.2 ± 0.1 | 0.2 ± 0.0 | 0.0781 |
| ∑n-3 PUFA | 233.4 ± 196.1 | 149.9 ± 141.9 | 119.6 ± 27.5 | 0.0114 |
| ∑n-6 PUFA | 321.2 ± 255.7 | 206.6 ± 162.3 | 187.6 ± 71.6 | 0.0067 |
| n-6/ n-3 PUFA | 1.5 ± 0.4 | 1.5 ± 0.3 | 1.5 ± 0.3 | 0.8487 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pewan, S.B.; Otto, J.R.; Kinobe, R.T.; Adegboye, O.A.; Malau-Aduli, A.E.O. MARGRA Lamb Eating Quality and Human Health-Promoting Omega-3 Long-Chain Polyunsaturated Fatty Acid Profiles of Tattykeel Australian White Sheep: Linebreeding and Gender Effects. Antioxidants 2020, 9, 1118. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9111118
Pewan SB, Otto JR, Kinobe RT, Adegboye OA, Malau-Aduli AEO. MARGRA Lamb Eating Quality and Human Health-Promoting Omega-3 Long-Chain Polyunsaturated Fatty Acid Profiles of Tattykeel Australian White Sheep: Linebreeding and Gender Effects. Antioxidants. 2020; 9(11):1118. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9111118
Chicago/Turabian StylePewan, Shedrach Benjamin, John Roger Otto, Robert Tumwesigye Kinobe, Oyelola Abdulwasiu Adegboye, and Aduli Enoch Othniel Malau-Aduli. 2020. "MARGRA Lamb Eating Quality and Human Health-Promoting Omega-3 Long-Chain Polyunsaturated Fatty Acid Profiles of Tattykeel Australian White Sheep: Linebreeding and Gender Effects" Antioxidants 9, no. 11: 1118. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9111118