Control strategies of gecko’s toe in response to reduced gravity



1
Special Robot Research Center, Hefei CSG Smart Robot Technology Co., Ltd., Hefei 230088, China
2
Special Robot Research Center, CSG Smart Science & Technology Co., Ltd., Shanghai 201619, China
3
Institute of Bio-inspired Structure and Surface Engineering, Nanjing University of Aeronautics and Astronautics, Nanjing 210016, China
4
Department Functional Morphology and Biomechanics, Zoological Institute of the University of Kiel, Kiel 24118, Germany
*
Author to whom correspondence should be addressed.
Appl. Sci. 2020, 10(7), 2257; https://0-doi-org.brum.beds.ac.uk/10.3390/app10072257
Submission received: 7 March 2020
/
Revised: 24 March 2020
/
Accepted: 24 March 2020
/
Published: 26 March 2020
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Shear-induced adhesion is one of the key properties for the gecko moving safely and quickly in a three-dimensional environment. The control strategies of such locomotion strongly relying on adhesion are still not well understood. In this study, we measured foot alignment and three-dimensional reaction forces of the single toes of the Tokay gecko running on the ground freely (gravity condition) and running in a situation where the gravity force was counterbalanced (reduced gravity condition). The forelimb rotated from the outward position to the front-facing position and the hindlimb rotated from the outward position to the rear-facing position, when running with balanced force, which indicated that the adhesive system was employed behaviorally through the modulation of the foot alignment. The toe was compressed and pulled in the gravity condition, but it was tensed and pulled in the reduced gravity condition. There was an approximately linear relationship between peak normal forces and the corresponding shear forces in both the reduced gravity condition (FN = −0.40FS − 0.008) and the gravity condition (FN = 2.70FS − 0.12). The footpad was compressed and pushed in the gravity condition, whereas it was tensed and pulled in the reduced gravity condition. There was an approximately linear relationship between peak normal forces and the corresponding shear forces in both the reduced gravity condition (FN = −0.39FS − 0.001) and in the gravity condition (FN = −2.80FS − 0.08). The shear-induced adhesion of the gecko footpad is controlled by the coupling of the normal force and shear forces: that is why in this system adhesion was shear-sensitive and friction was load-sensitive. Our measurements of single toe reaction forces also show that geckos control their footpad attachment using ‘toe rolling-in and gripping’ motion in both gravity and reduced gravity conditions.
1. Introduction
Geckos are able to run on vertical and even inverted surfaces as fast as other terrestrial animals can run on normal ground. It is widely believed that the secret of such extraordinary locomotory ability lies in the adhesive structures found on the toes. [1,2,3]. One fundamental property of their adhesive structures that helps to achieve a controlled, energy-efficient adhesion is the directional dependence of their adhesion. Patterns of ground reaction forces show that geckos can control attachment and detachment via shear forces, by pulling their legs towards or pushing their legs away from the body [1,4]. Separate parts of this control system seem to be established at different levels of the hierarchical organization of this adhesive system [5,6,7,8,9,10,11,12,13]. For example, the animal can control adhesion by shifting the position of the center of gravity, by changing each footfall position (e.g., with different joint movement), by varying gait characteristics (e.g., velocity, stride length, stride frequency and duty factor) or by employing specific features of the adhesive system itself (e.g., viscoelasticity of the underlying layers or hierarchy of setal organization).
Although the basic functional mechanisms of adhesive pads are rather well understood, the control principles of attachment and detachment are still largely unknown. Control strategies that have been investigated in terrestrial animals with perturbation suggest that both active (neuro-muscular) control and passive (mechanical) reactions play important roles in the control of rapid locomotion [14,15,16,17,18]. Patterns of single leg ground reaction forces suggest dynamic stability control in gecko locomotion [4,5,7]. Differences in locomotor performances [4,5,7,19] suggest that a trade-off between successful adhesion and the ability to respond dynamically to locomotor perturbations may exist.
Multiple toes differing in orientation appear to play an important role during gecko locomotion, as the animal can avoid detachment by aligning some of their toes opposite to the force vector. It is likely that employment of the adhesive system is associated with modulation of inter-digital angle and foot alignment [20,21]. However, much less is known about adhesion force of single toes during locomotion.
In the present study, we aimed to obtain measurements of ground reaction forces generated by the single toes of geckos running on level substrate. We used helium-filled balloons to counterbalance the gecko’s body weight and to observe how the adhesive pads were applied to the ground. We addressed the following questions: what are the single leg and single toe ground reaction forces, and how do geckos employ their adhesive system in response to the reduced gravity?
2. Materials and Methods
2.1. Study Animals
Six individual Tokay geckos (Gekko gecko; body mass 57.3 ± 7.8 g; snout-vent length 133.5 ± 16.9 cm; mean ± s.d.) were obtained from a supplier in Guangxi Province, China. They were housed in pairs in separate terraria, provided with fresh water and live insects ad libitum, and were kept on a natural light cycle at a temperature of 24–27 °C and at a relative humidity of 60%–70%.
2.2. Force Measurement Set-up
To measure ground reaction forces of both single legs and single toes in all three directions, a force measurement array (FMA) was used [22]. The FMA consisted of 24 separate, custom-built three dimensional force sensors, arranged in three rows and eight columns with each sensor having a glass cover-slip (ca 3 cm × 3 cm × 0.1 cm with ca 0.1 cm clearance gap) on top, resulting in a tiled strip of ca 25 cm × 9 cm measurement area (average roughness Ra = 4.7 ± 2.1 μm, N = 24, mean ± s.d.).
The x-axis is defined as the left-right axis in the plane of the platform. The x-axis points to the locomotion direction of the animal. The forces along the x-axis are termed as fore-aft forces. The y-axis is defined as being perpendicular to the x-axis and is situated in the plane of the platform. The y-axis forces are termed as lateral forces or shear forces. The z-axis is defined as being perpendicular to the xy-plane and the forces are termed as normal forces.
The sensor was made up of T-shaped aluminium alloy [22] and forces were measured from the deflection of the cantilever beams by 350 Ω foil strain gauges glued to the beams. The resolution for each direction was similar (ca 1–2 mN); the fundamental resonance frequency for the x- and y-axes was lower (ca 252 and ca 125 Hz) than for the z-axis (ca 355 Hz). Signals from each sensor were amplified and collected by a multiple channel strain signal processing device (National Instruments, Austin, TX, USA) at a sampling frequency of 500 Hz. Data were filtered using a Butterworth filter at a cut-off frequency of 100 Hz.
2.3. Video Recording
Two high speed digital video cameras (Olympus i-SPEED 3, 1280 × 1024 pixels) were synchronized with the force recordings from top and side views at a sampling frequency of 500 Hz. A mirror was mounted at an angle of 60° next to the FMA, giving the other side view of the gecko. The camera ran in synchrony with the data acquisition, using a common starting pulse from a manual switch. Multiple cold light sources (light-emitting diodes) placed above the platform were used to supply adequate illumination for high speed video recording.
2.4. Force Measurements
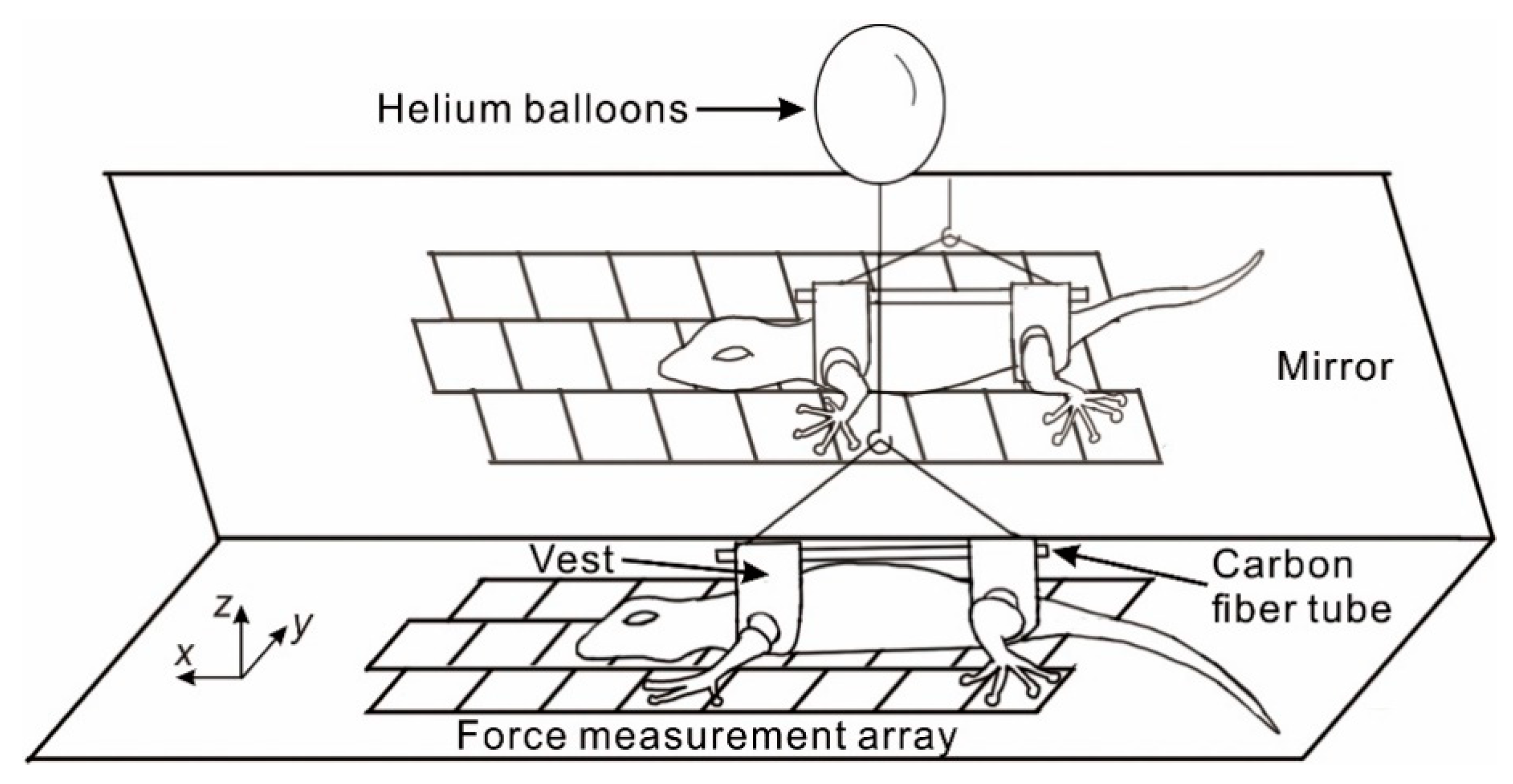
To evaluate the influence of balanced force on gecko locomotion, helium-filled balloons were fixed on a carbon fiber tube (inner diameter: 2 mm, outer diameter: 3 mm, mass: 1 g) which went through two custom-built vests that wrapped around the front and hind legs, respectively, so that the balloons were pulling from close to the center of mass of the animal (Figure 1). The gecko’s effective weight was reduced by counterbalancing the normal force arising from its body weight. The force generated by the balloons was about 0.6 N. We acquired locomotion with zero balanced force and with about 100% body weight balanced force.
The animal was placed on one side of the force platform, and the balloons were initially held by hand. Then, the hand was removed slightly, to gradually let the animal run on the FMA. Simultaneously, the data acquisition and video recording were initiated by a manual switch. Prior to each trial, we measured the body mass with balanced force (Figure 1).
We carefully checked the recorded videos, and selected only those recordings which met the following conditions: (i) the animal ran continuously straight along the FMA and through the field of view; (ii) no slip of the feet took place over the course of the experiment; (iii) no sudden acceleration or deceleration occurred during the trial..
2.5. Analysis of Forces and Video Recordings
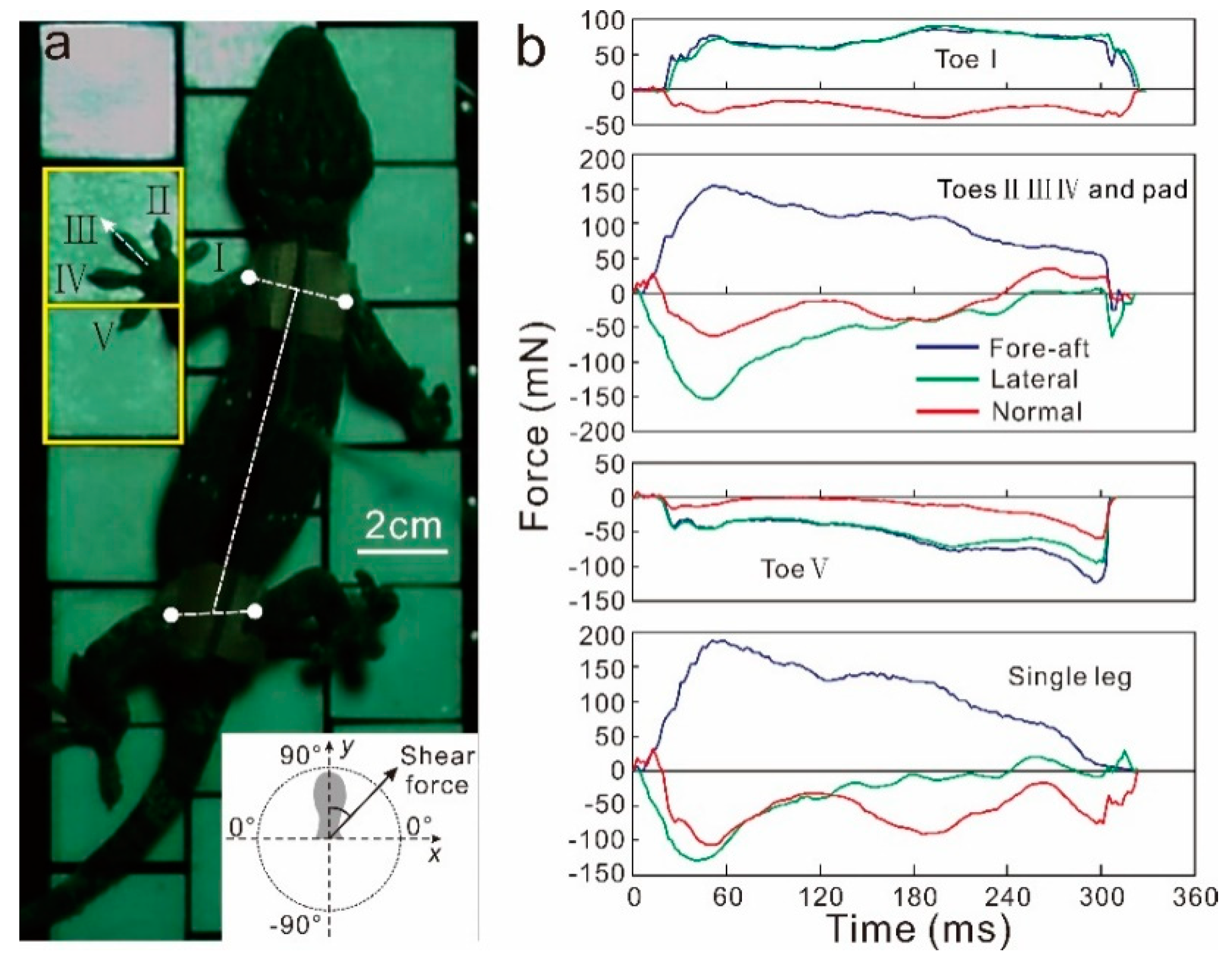
The forces and video analysis were both performed with custom-built MATLAB scripts (v2015b, Mathworks Corporation, USA). Joints were carefully digitized using DLT_dv5 [23]. The line between the mid-points of the shoulder and hips was defined as the body axis. The angle between the digit III and the body axis was defined as the limb orientation angle, and the angle between the digit I and digit V was defined as the limb spread angle (Figure 2a).
For the forces analysis, normal forces were divided into load (+) or adhesion (−) components, fore-aft forces represented propulsive (+) or braking (−) effort, and lateral forces indicated whether a limb was pushed away (+) or pulled towards the body (−). For cases where different toes of the same limb touch multiple sensors, the forces of individual toes and multiple toes were analysed, respectively, and the force components for each direction were summed up in order to obtain 3 resultant force vectors of each leg (Figure 2b). For single toe force orientation, we defined the toe orientation as y-axis and point to 90°. The x-axis was defined as being perpendicular to the y-axis and point to 0°. We defined the toe III force orientation as the footpad force orientation. If the angle of ground reaction force vector ranged from 0° to approximately 90°, we noted the toe/footpad was pulling; if the angle of ground reaction force vector ranged from 0° to approximately −90°, we noted the toe/footpad was pushing.
3. Statistics
All statistical tests were carried out using the statistical toolbox in Matlab (v2015b, Mathworks Corporation, USA). To evaluate the influence of balanced force on gecko locomotion, the data were divided into two categories: gravity and reduced gravity. The Mann–Whitney U-test was used to test between two categories. For the plots, we have indicated a significant difference between two samples using the ‘*’-symbol, if P < 0.05, ‘**’, if P < 0.01 and ‘***’, if P < 0.001. If a test just failed to reach the significance level, we stated the computed P-value.
Results were plotted using ‘box-whisker’ plots. The median and the 25 percent and 75 percent quartiles (‘box’) are indicated. The plotted ‘whiskers’ extend to the most extreme data values that are not outliers. Points are drawn as outliers, if they are larger than q1-w(q3-q1) or smaller than q1-w(q3-q1), with w = 1.5 being the whisker length and q1 and q3 the 25th and 75th percentiles, respectively. All tested data are presented as means ± standard deviation unless otherwise indicated.
4. Results
4.1. Foot Alignment
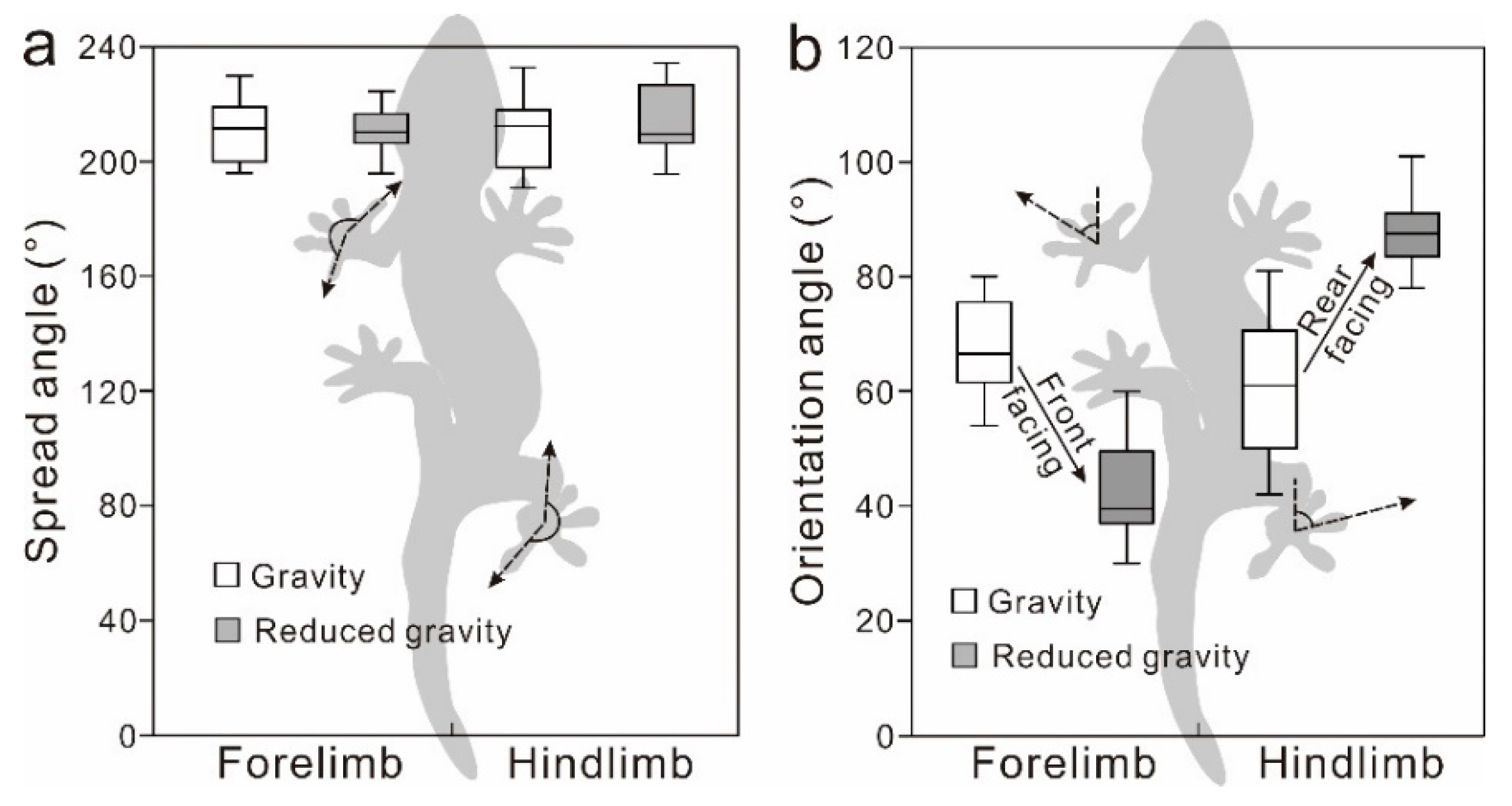
The spread angle had no significant difference between gravity condition and reduced gravity condition in both forelimbs (Mann–Whitney U-test: W = 198.5, Z = −0.21, P = 0.84) and hindlimbs (Mann–Whitney U-test: W = 182, Z = −0.97, P = 0.33). The orientation angle of the forelimb was significantly lower in the reduced gravity condition than in the gravity condition (Mann–Whitney U-test: W = 140.5, Z = −4.66, R = 528, P < 0.001): this indicates the rotation of the forelimb from the outward to the front-facing position. The orientation angle of the hindlimb was significantly higher in the reduced gravity condition than in the gravity condition (Mann–Whitney U-test: W = 139, Z = 4.71, R = 528, P < 0.001): this indicates the increased rotation of the hindlimb from the outward position to the rear-facing position (Figure 3).
4.2. Ground Reaction Forces
In both gravity and reduced gravity conditions, gecko toes were pulling. The angle of toe force orientation shows that the toe approximately aligned with the force vector in the reduced gravity condition (ranged from 82° to 90°), but deviated from the force vector in the gravity condition (ranged from 13° to 83°).
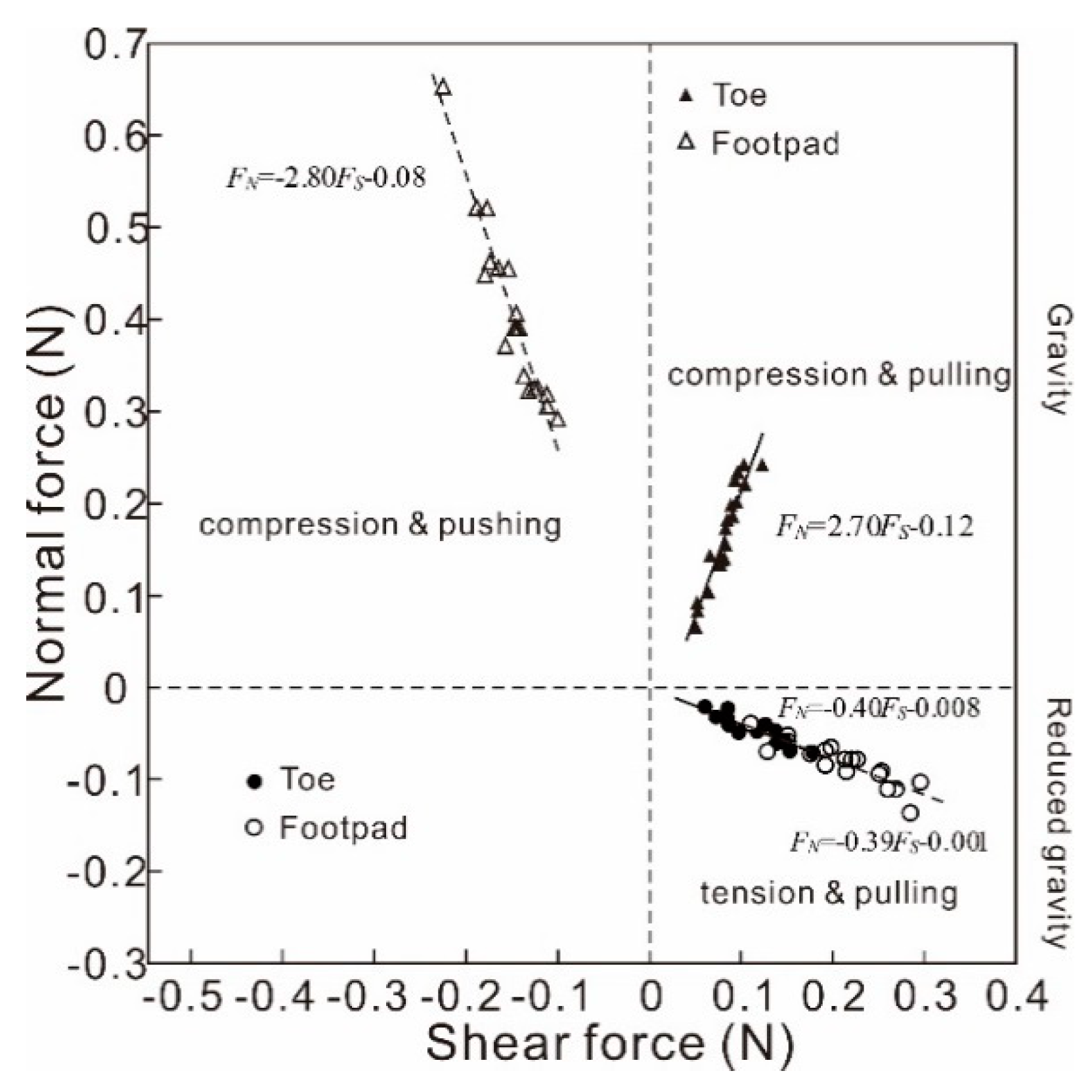
The toe was compressed and pulled in the gravity condition, but it was tensed and pulled in the reduced gravity condition. There was an approximately linear relationship between peak normal forces and the corresponding shear forces in both the reduced gravity condition (FN = − 0.40FS − 0.008) and in the gravity condition (FN = 2.70FS − 0.12). The footpad was compressed and pushed in the gravity condition, whereas it was tensed and pulled in the reduced gravity condition. There was an approximately linear relationship between peak normal forces and the corresponding shear forces in both the reduced gravity condition (FN = − 0.39FS − 0.001) and in the gravity condition (FN = − 2.80FS − 0.08), as shown in Figure 4
5. Discussion
Our results show that geckos make dynamic use of their attachment foot pads with ‘toe rolling-in and gripping’ motion. As a toe approaches the surface, a small loading force perpendicular to the contact surface and a short drag parallel to the contact surface is applied to orient and preload setae as well as spatulae [1,3,24], and thereby may generate a large contact area of the toes. Then, the toe rolls in and grips for further attachment enhancement. It has been previously shown for the Gekko gecko that initial preload and lateral shear are both necessary to generate high adhesion [1,3,24]. However, the adhesion force turns into a repulsive force when the load is above some critical value [25]. This is consistent with our result of single toe force, where the force vector shows pulling and tension, while the toe acts as an adhesive system (reduced gravity condition). However, where the force vector shows pulling and compression, the toe has a supporting role (gravity condition). This trend can be explained by a Lennard–Jones potential function [26].
where Ez and Fvdw are the normal potential and the normal force between two surfaces. EA is the attractive energy and ER is the repulsive energy.
Ez = −EA(z/z0)−n + −ER(z/z0)−m (m > n),
Fvdw = −dEz/dz,
When Ez reaches its minimum value, the normal force Fvdw is zero with a surface gap distance D0. When the setae and the surface are pressed together by a loading force with D > D0, the force Fvdw is attractive, reaching its maximum value of max at a certain surface gap distance. When the setae and the surface are pressed together by a loading force with D < D0, the force Fvdw is repulsive.
The synergy between normal force and shear force, known as shear-induced adhesion, is one of the most important mechanical properties of the gecko seta. When the setal arrays are dragged against the gripping direction, the shear force is proportional to the normal force with a coefficient of 0.31 [2]. When the toe is dragged along the griping direction with a small loading force, the shearing behavior is totally different and could not be described in terms of the Amontons’ law like the friction coefficient. Surprisingly, although forces are not equally shared among different individual toes, the shear force is proportional to the normal force with a friction coefficient of 0.37, when the toe is dragged along the griping direction in the gravity condition. This value is in the same order of magnitude as the value of 0.36 found for the setal arrays of Gekko gecko [25]. Our findings here show that the shear-induced adhesion is controlled by the coupling of the normal force and shear force. It means that in this system, the adhesion is shear-sensitive and the friction is load-sensitive. It has been reported that positive normal force (preload) is necessary for adhesion generation. However, it is still unclear how the normal force affects adhesion in detail. Adhesion force increases linearly with the shear force and approaches steady-state values only after the setal tip begins to slide and reorient in the drag step [1,2,27]. Such a slide contact is achieved by a combined normal and shear force [28,29,30]. The body weight, which contributes to an increase in the normal force, and the shear force, which depends on the toe orientation, may induce a transition from tip to slide contact. According to the Lennard–Jones potential function [26,31], the normal force turns into a repulsive force depending on the contact state of the setal tip. Previous studies have shown that adhesion force was influenced by the manner how the setae and spatula contact the substrate [27,32,33]. We thus conclude that adhesion is not solely controlled by direction-dependence and morphological anisotropy, but also can be switched on and off by applying the load in a different manner. Interestingly, the force vector shows that the footpad pushes away from the body and the toe grips in (pull toward) during level running. The heel, without any adhesive structures, may play an important role in the footpad pushing away from the body.
The performance of the adhesive pads does not solely depend on the adhesive setae layer itself, but also on the way it is used for locomotion at the toe, footpad and whole animal level. Geckos alter foot orientation to facilitate adhesion in response to changes in habitat structure [20,21]. Here, we provide evidence that the adhesive system is engaged and disengaged behaviorally through the modulation of the foot alignment. Movements of joints show that geckos are likely to alter proximal joints to provide extra work, rather than altering distal joints that could affect the success of their adhesive capabilities [20]. Such an adjustment in joints leads to the minimization of torques [34]. Electromyography analysis in geckos shows that there are few significant changes in muscle activity patterns in response to the degree of incline [35]. This in turn suggests that the motion to engage and disengage the adhesive/gripping system is very stereotyped by using a passive “toe-gripping” actuation, which has been previously studied in insects [36,37]. Geckos may use the “toe-gripping” actuation as a control strategy and a safety mechanism for unpredictable and fast perturbations, preventing detachment from the substrate. The passive control of toe use represents a striking example of a mechanical system that is not only capable of stabilizing locomotion against rapid perturbations [17,38], but also to react to an environmental condition by adopting an ‘‘adaptive’’ state without any neuronal feedback. Controlling attachment using smart, mechanical constructions instead of additional muscles has a series of advantages [39,40,41]. First, a passive control is always faster than an active one, as a sensor-input system requires extra time for muscle contraction. Second, less musculature involved in control is a cost-effective way which can conserve metabolic energy.
Separate use of friction and adhesion appears to be wide-spread in natural organisms. It has been reported that arthropods have different pads on the same foot for fundamentally different functions; load-sensitive “heel’’ pads for friction and shear-sensitive ‘‘toe’’ pads for adhesion, such as in cockroaches [42], stick insects [43,44], spiders [45] and weaver ants [46]. Furthermore, the modulation of foot orientation is also an important biological principle that promises to be very useful for technical application [33]. The characteristic structure of the pes and manus of the gecko, allowing the feet to rotate freely and the digits to be spread widely [47], leads to the animal’s ability to adhere in the head up and head down orientation as well as on vertical surfaces [48]. Observation in tree frogs showed that they are unable to adhere to smooth surfaces in the head-down orientation [49], probably due to the constraint in digit spread and foot alignment. The sophisticated mechanical systems involved in gecko locomotion are particularly promising as inspiration for new robot designs.
Author Contributions
Z.W. and Z.D. conceived and designed the experiments; Z.W. performed the experiments, analyzed the data; Z.W., S.N.G. and Z.D. wrote the paper. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China grant number [51435008] No. to Zhendong Dai and partially by Jiangsu Province Friendship Award to Stanislav N. Gorb.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Autumn, K.; Liang, Y.A.; Hsieh, S.T.; Zesch, W.; Chan, W.P.; Kenny, T.W.; Fearing, R.; Full, R.J. Adhesive force of a single gecko foot-hair. Nature 2000, 405, 681–685. [Google Scholar] [CrossRef] [PubMed]
- Autumn, K.; Dittmore, A.; Santos, D.; Spenko, M.; Cutkosky, M. Frictional adhesion: A new angle ongecko attachment. J. Exp. Biol. 2006, 209, 3569–3579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, H.; Wang, X.; Yao, H.; Gorb, S.; Arzt, E. Mechanics of hierarchical adhesion structures of geckos. Mech. Mater. 2005, 37, 275–285. [Google Scholar] [CrossRef]
- Wang, Z.; Dai, Z.; Ji, A.; Ren, L.; Xing, Q.; Dai, L. Biomechanics of gecko locomotion: The patterns of reaction forces on inverted, vertical and horizontal substrates. Bioinsp. Biomim. 2015, 10, 016019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Autumn, K.; Hsieh, S.T.; Dudek, D.M.; Chen, J.; Chitaphan, C.; Full, R.J. Dynamics of geckos running vertically. J. Exp. Biol. 2006, 209, 260–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birn-Jeffery, A.V.; Higham, T.E. Geckos decouple fore-and hind limb kinematics in response to changes in incline. Front. Zool. 2016, 13, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.J.; Peattie, A.M.; Autumn, K.; Full, R.J. Differential leg function in a sprawled-posture quadrupedal trotter. J. Exp. Biol. 2006, 209, 249–259. [Google Scholar] [CrossRef] [Green Version]
- Higham, T.E.; Birn-Jeffery, A.V.; Collins, C.E.; Hulsey, C.D.; Russell, A.P. Adaptive simplification and the evolution of gecko locomotion: Morphological and biomechanical consequences of losing adhesion. Proc. Natl. Acad. Sci. USA 2015, 112, 809–814. [Google Scholar] [CrossRef] [Green Version]
- Irschick, D.J.; Jayne, B.C. Effects of incline on speed, acceleration, body posture and hindlimb kinematics in two species of lizard Callisaurus draconoides and Uma scoparia. J. Exp. Biol. 1998, 201, 273–287. [Google Scholar]
- Irschick, D.J.; Jayne, B.C. Comparative three-dimensional kinematics of the hindlimb for high-speed bipedal and quadrupedal locomotion of lizards. J. Exp. Biol. 1999, 202, 1047–1065. [Google Scholar]
- Irschick, D.J.; Vanhooydonck, B.; Herrel, A.; Andronescu, A. Effects of loading and size on maximum power output and gait characteristics in geckos. J. Exp. Biol. 2003, 206, 3923–3934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaaf, A.; Van Damme, R.; Herrel, A.; Aerts, P. Spatio-temporal gait characteristics of level and vertical locomotion in a ground-dwelling and a climbing gecko. J. Exp. Biol. 2001, 204, 1233–1246. [Google Scholar] [PubMed]
- Filippov, A.E.; Gorb, S.N. Spatial model of the gecko foot hair: Functional significance of highly specialized non-uniform geometry. Interface Focus 2015, 5, 20140065. [Google Scholar] [CrossRef] [PubMed]
- Daley, M.A.; Biewener, A.A. Running over rough terrain reveals limb control for intrinsic stability. Proc. Natl. Acad. Sci. USA 2006, 103, 15681–15686. [Google Scholar] [CrossRef] [Green Version]
- Dickinson, M.H.; Farley, C.T.; Full, R.J.; Koehl, M.A.R.; Kram, R.; Lehman, S. How animals move: An integrative view. Science 2000, 288, 100–106. [Google Scholar] [CrossRef] [Green Version]
- Full, R.J.; Koditschek, D.E. Templates and anchors: Neuromechanical hypotheses of legged locomotion on land. J. Exp. Biol. 1999, 202, 3325–3332. [Google Scholar]
- Jindrich, D.L.; Full, R.J. Dynamic stabilization of rapid hexapedallocomotion. J. Exp. Biol. 2002, 205, 2803–2823. [Google Scholar]
- Sponberg, S.; Full, R.J. Neuromechanical response of musculo-skeletal structures in cockroaches during rapid running on rough terrain. J. Exp. Biol. 2008, 211, 433–446. [Google Scholar] [CrossRef] [Green Version]
- Russell, A.P.; Higham, T.E. A new angle on clinging in geckos: Incline, not substrate, triggers the deployment of the adhesive system. Proc. R. Soc. B 2009, 276, 3705–3709. [Google Scholar] [CrossRef] [Green Version]
- Birn-Jeffery, A.V.; Higham, T.E. Geckos significantly alter foot orientation to facilitate adhesion during downhill locomotion. Biol. Lett. 2014, 10, 20140456. [Google Scholar] [CrossRef]
- Zhuang, M.V.; Higham, T.E. Arboreal day geckos (Phelsuma madagascariensis) Differentially modulate fore- and hind limb kinematics in response to changes in habitat structure. PLoS ONE 2016, 11, e0153520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, Z.; Wang, Z.; Ji, A. Dynamics of gecko locomotion: A force-measuring array to measure 3D reaction forces. J. Exp. Biol. 2011, 214, 703–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedrick, T.L. Software techniques for two- and three-dimensional kinematic measurements of biological and biomimetic systems. Bioinsp. Biomim. 2008, 3, 034001. [Google Scholar] [CrossRef] [PubMed]
- Huber, G.; Gorb, S.N.; Spolenak, R.; Arzt, E. Resolving the nanoscale adhesion of individual gecko spatulae by atomic force microscopy. Biol. Lett. 2005, 1, 2–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, J.; Tian, Y.; Zhou, M.; Zhang, X.J.; Meng, Y.G. Experimental research of load effect on the anisotropic friction behaviors of gecko seta array. Acta Phys. Sin. 2012, 61, 016202. [Google Scholar]
- Tian, Y.; Pesika, N.; Zeng, H.; Rosenberg, K.; Zhao, B.; McGuiggan, P.; Autumn, K. Adhesion and friction in gecko toe attachment and detachment. Proc. Natl. Acad. Sci. USA 2006, 103, 19320–19325. [Google Scholar] [CrossRef] [Green Version]
- Zhao, B.; Pesika, N.; Rosenberg, K.; Tian, Y.; Zeng, H.; McGuiggan, P.; Autumn, K.; Israelachvili, J. Adhesion and friction force coupling of gecko setal arrays: Implications for structured adhesive surfaces. Langmuir 2008, 24, 1517–1524. [Google Scholar] [CrossRef]
- Jeong, H.E.; Lee, J.K.; Kwak, M.K.; Moon, S.H.; Suh, K.Y. Effect of leaning angle of gecko-inspired slantedpolymer nanohairs on dry adhesion. Appl. Phys. Lett. 2010, 96, 043704. [Google Scholar] [CrossRef] [Green Version]
- Piccardo, M.; Chateauminois, A.; Fretigny, C.; Pugno, N.M.; Sitti, M. Contact compliance effects in the frictional response of bioinspired fibrillar adhesives. J. R. Soc. Interface 2013, 10, 20130182. [Google Scholar] [CrossRef]
- Shen, L.; Jagota, A.; Hui, C.Y. Mechanism of slidingfriction on a film-terminated fibrillar interface. Langmuir 2009, 25, 2772–2780. [Google Scholar] [CrossRef]
- Filippov, A.; Gorb, S.N. Frictional-anisotropy-based systems in biology: Structural diversity and numerical model. Sci. Rep. 2013, 3, 1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, G.C.; Soto, D.R.; Peattie, A.M.; Full, R.J.; Kenny, T.W. Orientation angle and the adhesion of single gecko setae. J. R. Soc. Interface 2011, 8, 926–933. [Google Scholar] [CrossRef] [PubMed]
- Filippov, A.; Popov, V.L.; Gorb, S.N. Shear induced adhesion: Contact mechanics of biological spatula-like attachment devices. J. Theor. Biol. 2011, 276, 126–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Full, R.J.; Blickhan, R.; Ting, L. Leg design in hexapedal runners. J. Exp. Biol. 1991, 158, 369–390. [Google Scholar]
- Foster, K.L.; Higham, T.E. Context-dependent changes in motor control and kinematics during locomotion: Modulation and decoupling. Proc. R. Soc. B 2014, 281, 20133331. [Google Scholar] [CrossRef] [Green Version]
- Bußhardt, P.; Gorb, S.N. Walking on smooth and rough ground: Activity and timing of the claw retractor muscle in the beetle Pachnoda marginata peregrina (Coleoptera, Scarabaeidae). J. Exp. Biol. 2013, 216, 319–328. [Google Scholar] [CrossRef] [Green Version]
- Bußhardt, P.; Gorb, S.N. Ground reaction forces in vertically ascending beetles and corresponding activity of the claw retractor muscle on smooth and rough substrates. J. Comp. Physiol. A 2014, 200, 385–398. [Google Scholar] [CrossRef]
- Kubow, T.M.; Full, R.J. The role of the mechanical system in control: A hypothesis of self-stabilization in hexapedal runners. Philos. Trans. R. Soc. Lond. B 1999, 354, 849–861. [Google Scholar] [CrossRef] [Green Version]
- Frantsevich, L.; Gorb, S.N. Arcus as a tensegrity structure in the arolium of wasps (Hymenoptera: Vespidae). Zoology 2002, 105, 225–237. [Google Scholar] [CrossRef]
- Frantsevich, L.; Gorb, S.N. Structure and mechanics of the tarsal chain in the hornet, Vespa crabro (Hymenoptera: Vespidae): Implications on the attachment mechanism. Arthr. Struct. Dev. 2004, 33, 77–89. [Google Scholar] [CrossRef]
- Gladun, D.; Gorb, S.N. Insect walking techniques on thin stems. Arthr. Plant Interact. 2007, 1, 77–91. [Google Scholar] [CrossRef]
- Clemente, C.J.; Federle, W. Pushing versus pulling: Division of labour between tarsal attachment pads in cockroaches. Proc. R. Soc. B 2008, 275, 1329–1336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bußhardt, P.; Gorb, S.N.; Wolf, H. Activity of the claw retractor muscle in stick insects in wall and ceiling situations. J. Exp. Biol. 2011, 214, 1678–1684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labonte, D.; Federle, W. Functionally different pads on the same foot allow control of attachment: Stick insects have load-sensitive “heel” pads for friction and shear-sensitive “toe” pads for adhesion. PLoS ONE 2013, 8, e81943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niederegger, S.; Gorb, S.N. Friction and adhesion in the tarsal and metatarsal scopulae of spiders. J. Comp. Physiol. A 2006, 192, 1223–1232. [Google Scholar]
- Endlein, T.; Federle, W. On heels and toes: How ants climb with adhesive pads and tarsal friction hair arrays. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russell, A.P.; Bauer, A.M.; Laroiya, R. Morphological correlates of the secondarily symmetrical pes of gekkotan lizards. J. Zool. 1997, 241, 767–790. [Google Scholar] [CrossRef]
- Russell, A.P.; Oetelaar, G.S. Limb and digit orientation during vertical clinging in Bibron’s gecko, Chondrodactylus bibronii (A. Smith, 1846) and its bearing on the adhesive capabilities of geckos. Acta Zool. 2015, 97, 345–360. [Google Scholar] [CrossRef]
- Hanna, G.; Barnes, W.J.P. Adhesion and detachment of the toe pads of tree frogs. J. Exp. Biol. 1991, 155, 103–125. [Google Scholar]
Figure 1.
Schematic of the set-up used to study single leg and toe reaction forces of the gecko. The gecko was put on the force platform consisting of 3 × 8 measurement arrays. Helium-filled balloons were used to balance the animal’s body weight. Note that the balloon and connecting thread are not drawn to scale.
Figure 1.
Schematic of the set-up used to study single leg and toe reaction forces of the gecko. The gecko was put on the force platform consisting of 3 × 8 measurement arrays. Helium-filled balloons were used to balance the animal’s body weight. Note that the balloon and connecting thread are not drawn to scale.

Figure 2.
(a) Top view of the gecko running on the force measurement array (FMA). The front left limb is contact with three force sensors. (b) Three-dimensional force-time curves of different toes in front left limbs.
Figure 2.
(a) Top view of the gecko running on the force measurement array (FMA). The front left limb is contact with three force sensors. (b) Three-dimensional force-time curves of different toes in front left limbs.

Figure 3.
(a) The spread angle of the forelimb and hindlimb in the gravity condition and in the reduced gravity condition. (b) The orientation angle of the forelimb and hindlimb in the gravity condition and in the reduced gravity condition.
Figure 3.
(a) The spread angle of the forelimb and hindlimb in the gravity condition and in the reduced gravity condition. (b) The orientation angle of the forelimb and hindlimb in the gravity condition and in the reduced gravity condition.

Figure 4.
Relationship between the normal force and shear force of the single toe and single pad in the gravity condition and in the reduced gravity condition.
Figure 4.
Relationship between the normal force and shear force of the single toe and single pad in the gravity condition and in the reduced gravity condition.

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, Z.; Gorb, S.N.; Dai, Z. Control strategies of gecko’s toe in response to reduced gravity. Appl. Sci. 2020, 10, 2257. https://0-doi-org.brum.beds.ac.uk/10.3390/app10072257
AMA Style
Wang Z, Gorb SN, Dai Z. Control strategies of gecko’s toe in response to reduced gravity. Applied Sciences. 2020; 10(7):2257. https://0-doi-org.brum.beds.ac.uk/10.3390/app10072257
Chicago/Turabian StyleWang, Zhongyuan, Stanislav N. Gorb, and Zhendong Dai. 2020. "Control strategies of gecko’s toe in response to reduced gravity" Applied Sciences 10, no. 7: 2257. https://0-doi-org.brum.beds.ac.uk/10.3390/app10072257
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.