The Effect of Elevated CO2 on Berry Development and Bunch Structure of Vitis vinifera L. cvs. Riesling and Cabernet Sauvignon

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Site, Experimental Design and Plant Material
2.2. Weather Conditions
2.3. Berry Development Sampling
2.4. Berry Analysis
2.5. Bunch Structure Measures
2.6. Statistical Analysis
3. Results
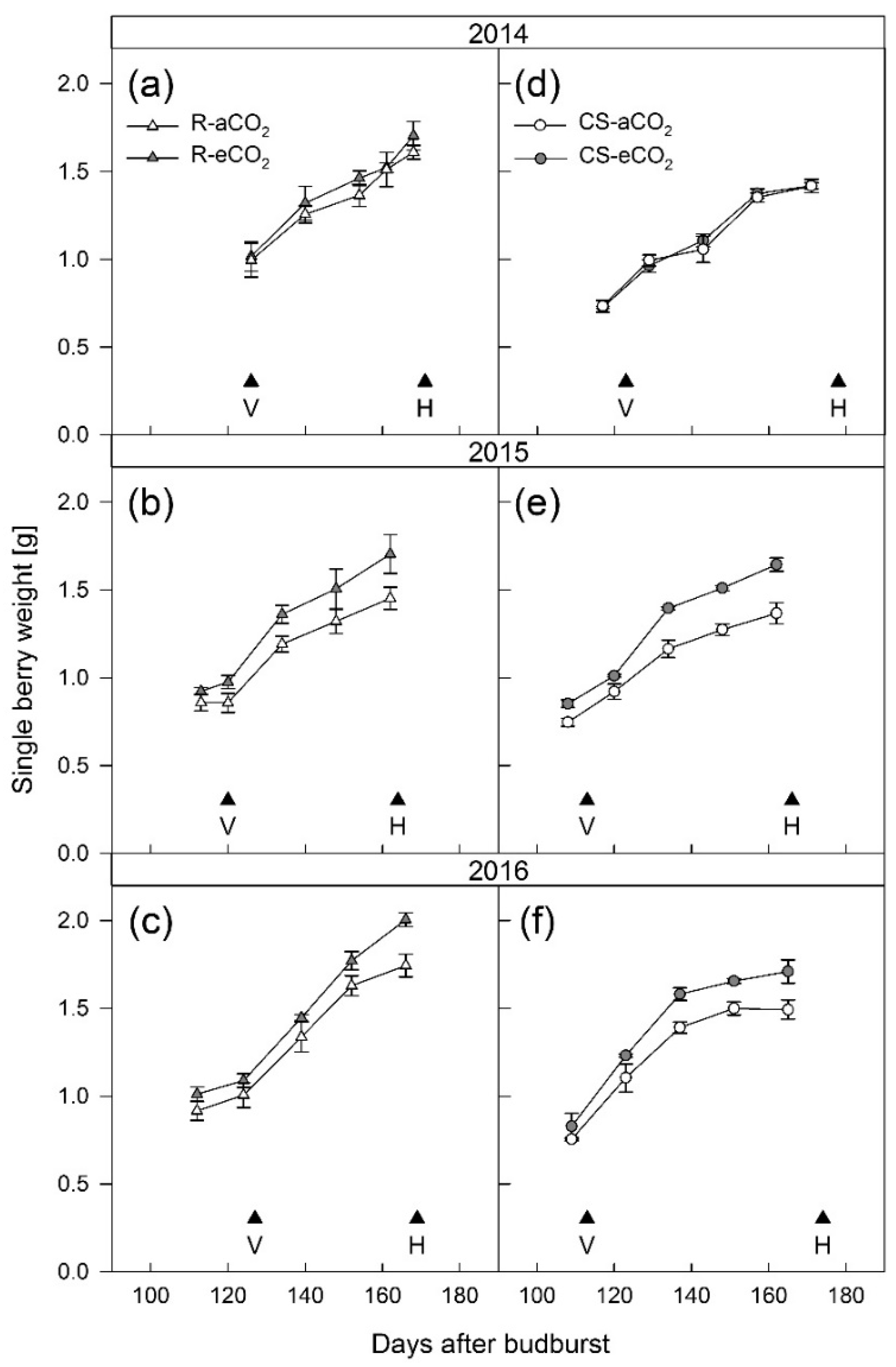
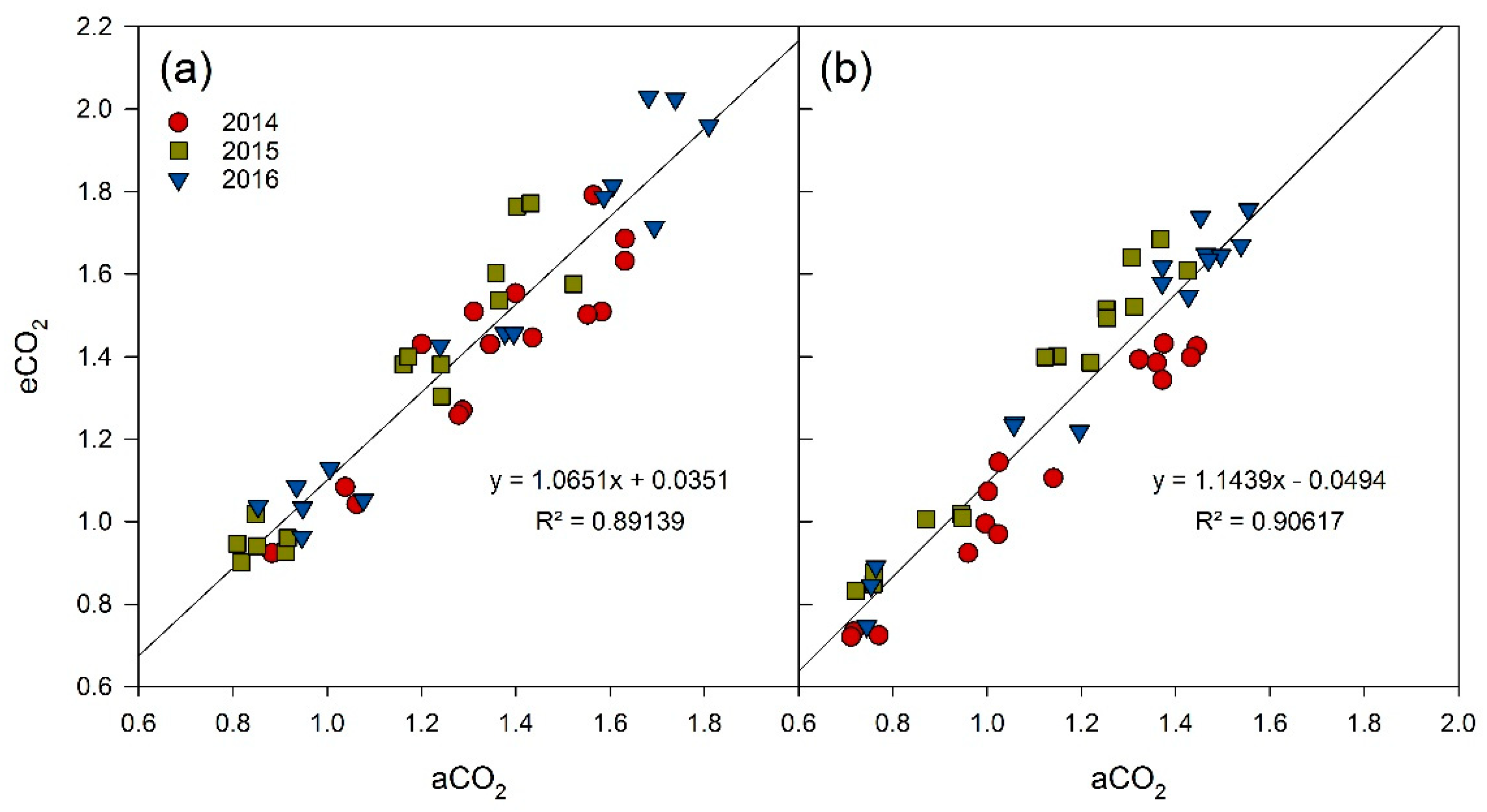
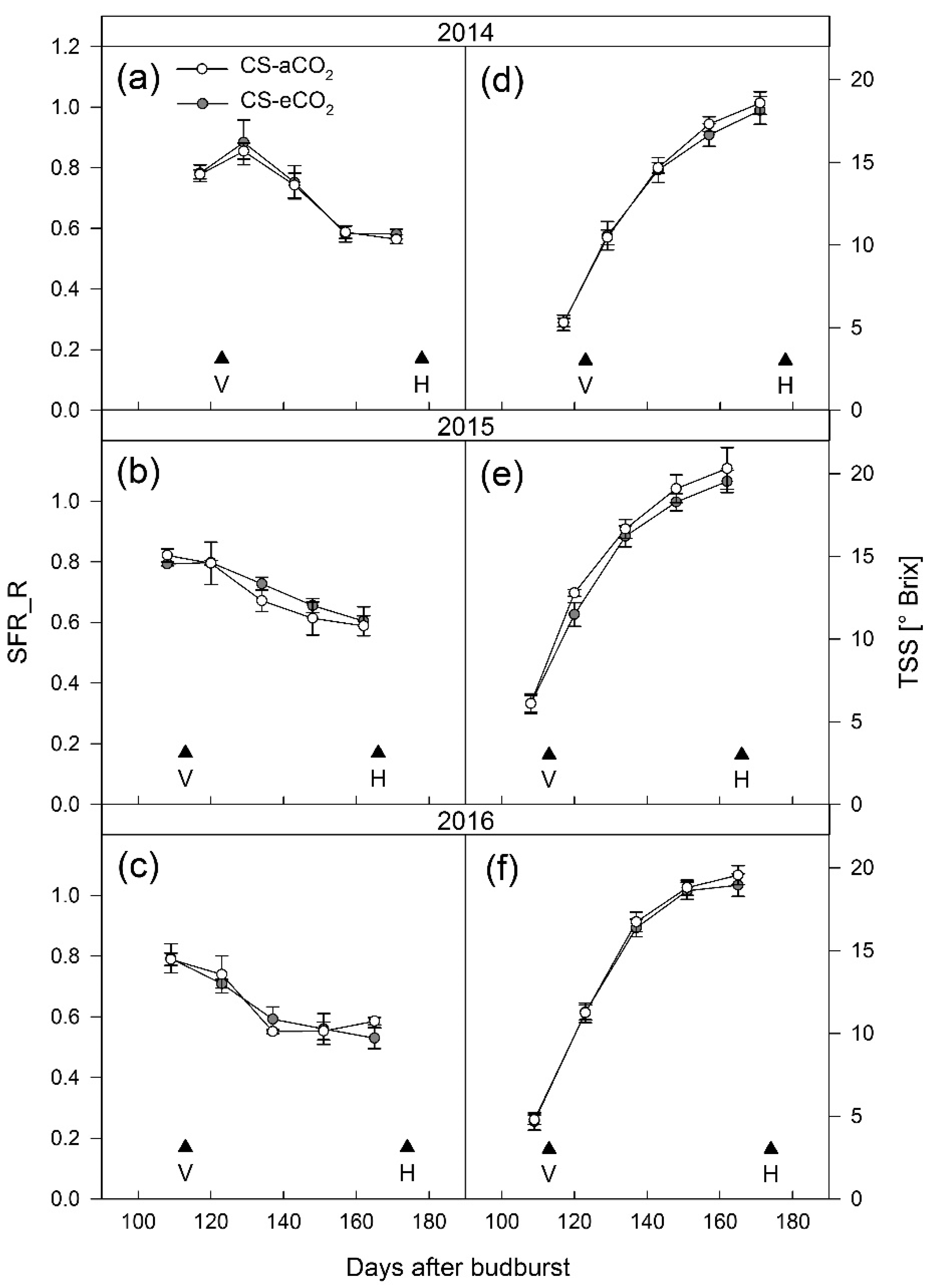
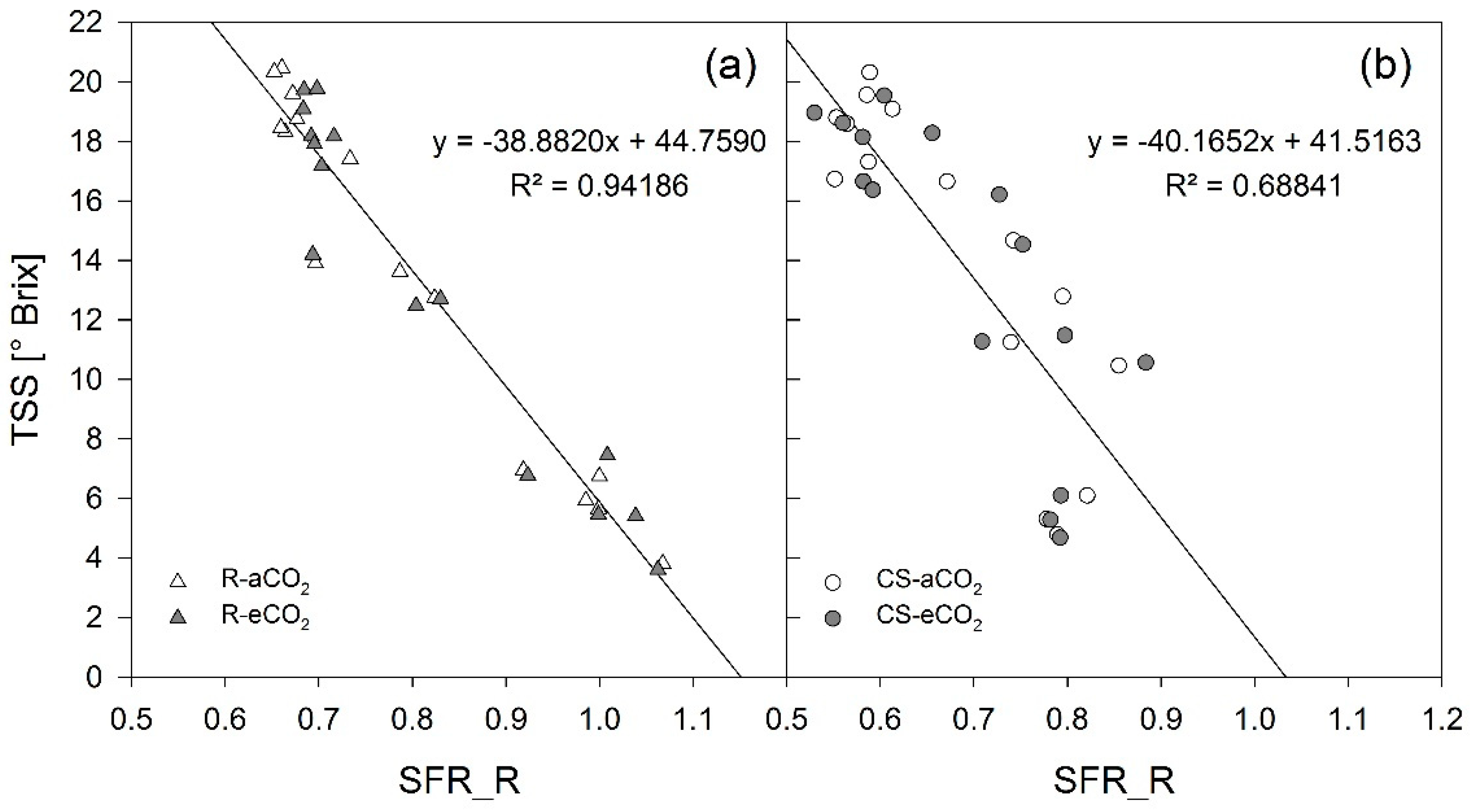
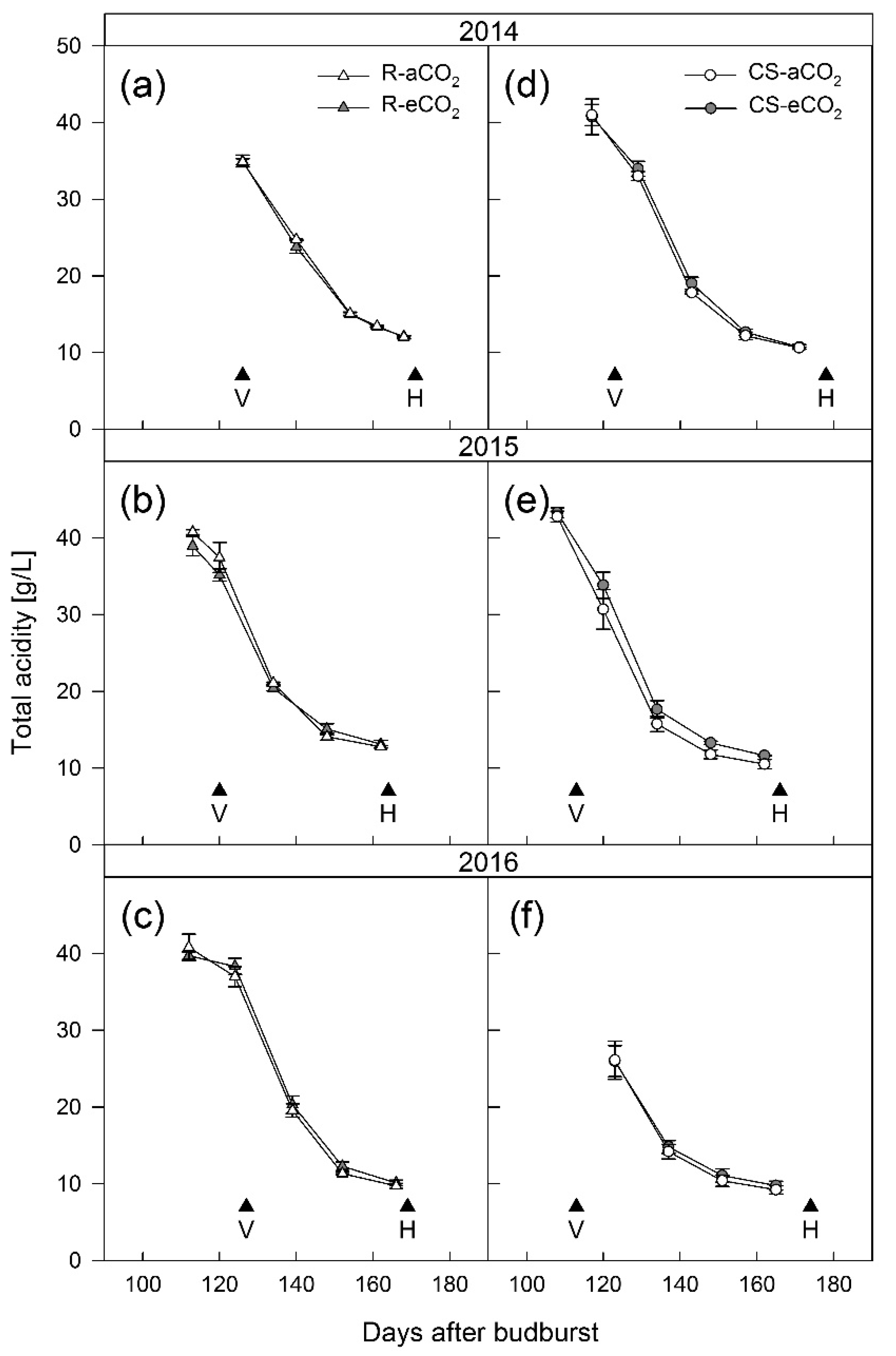
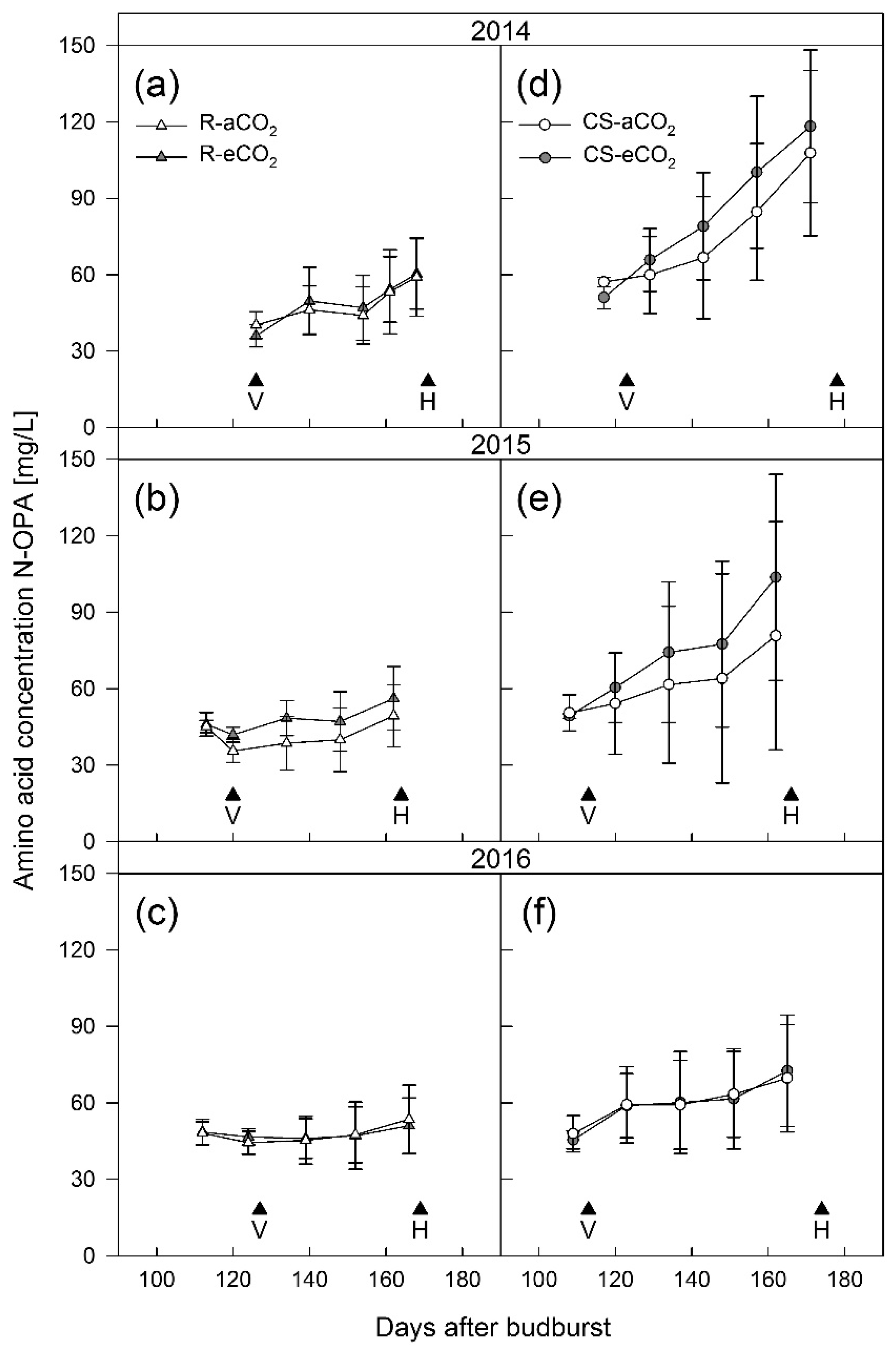
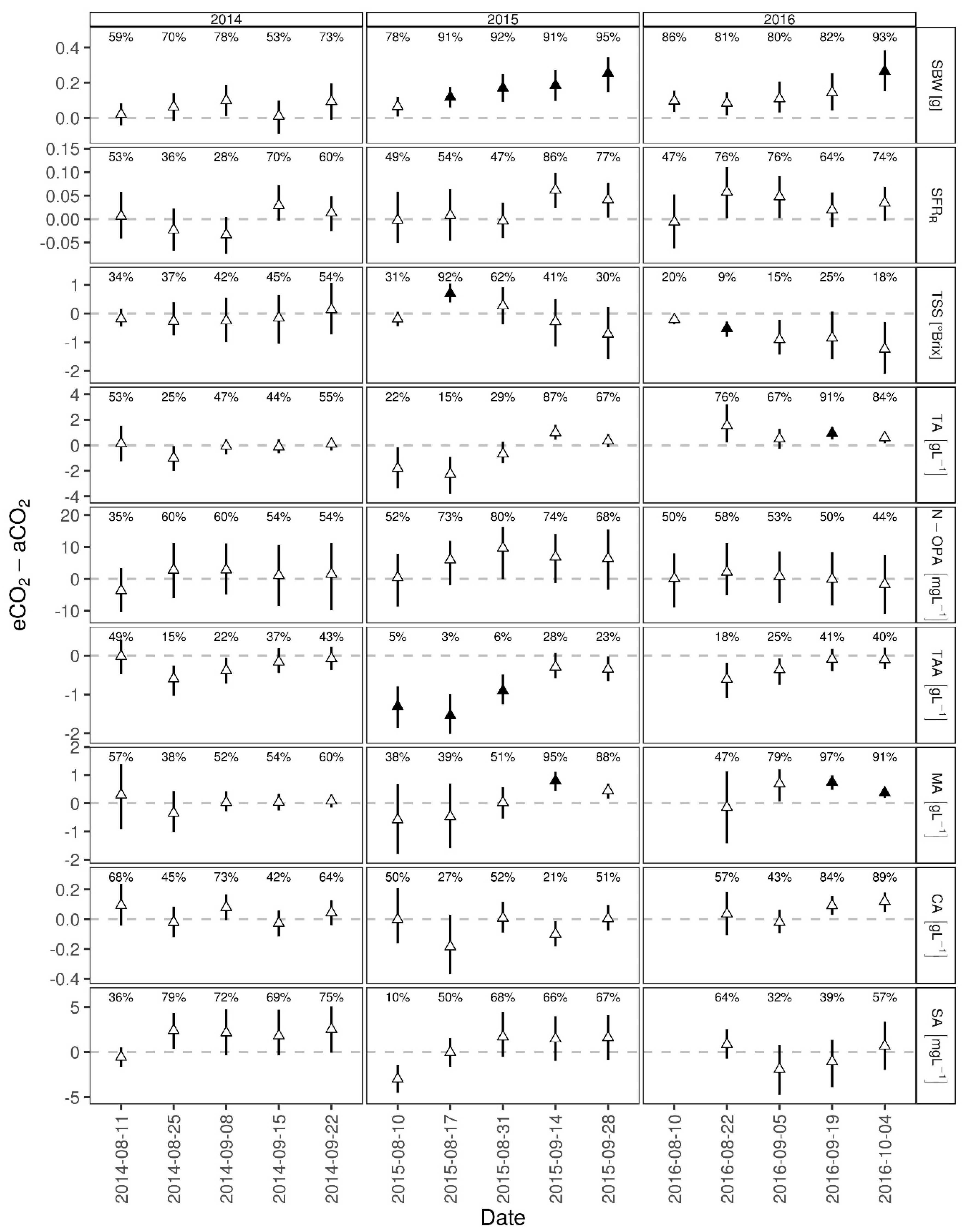
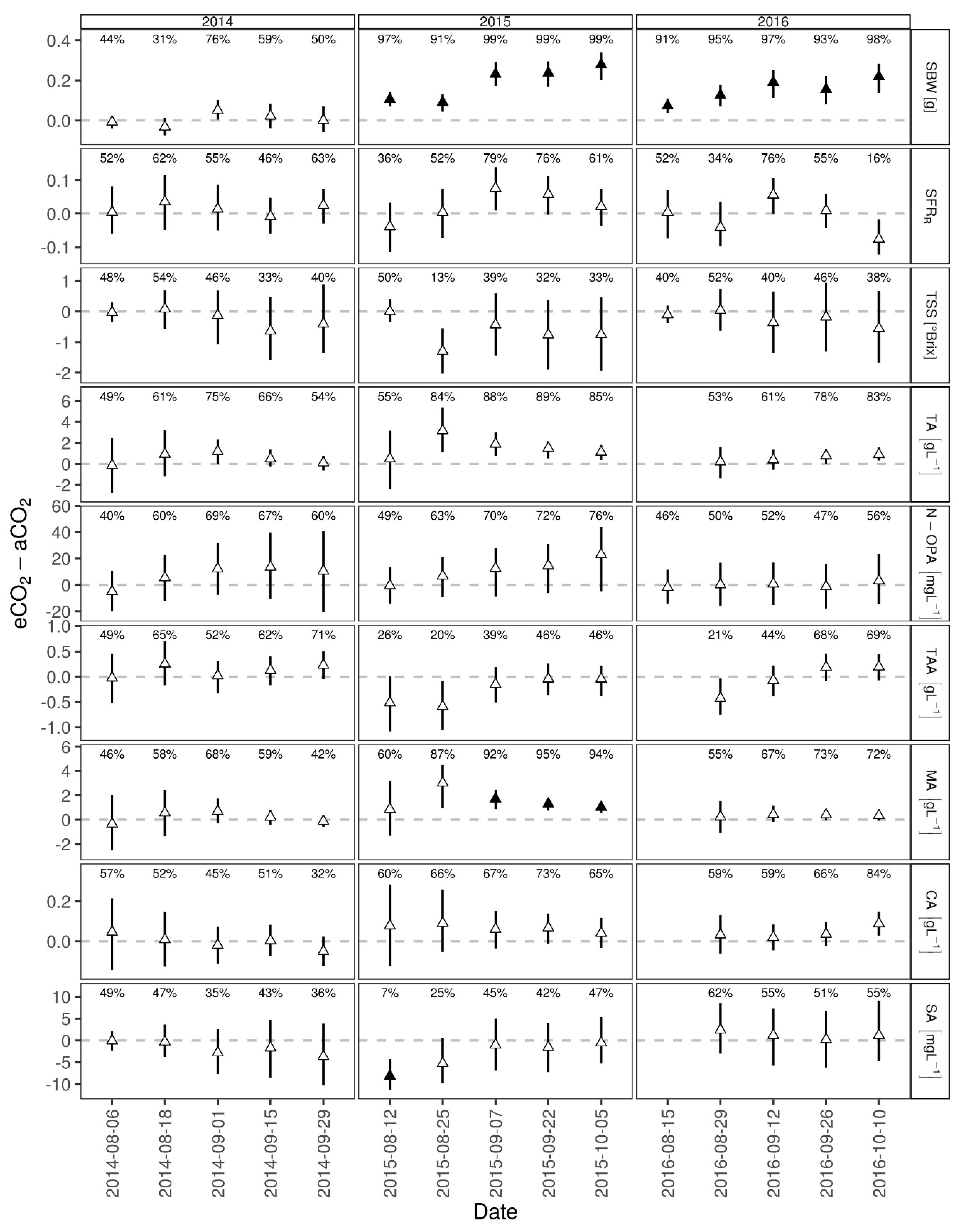
3.1. Berry Development
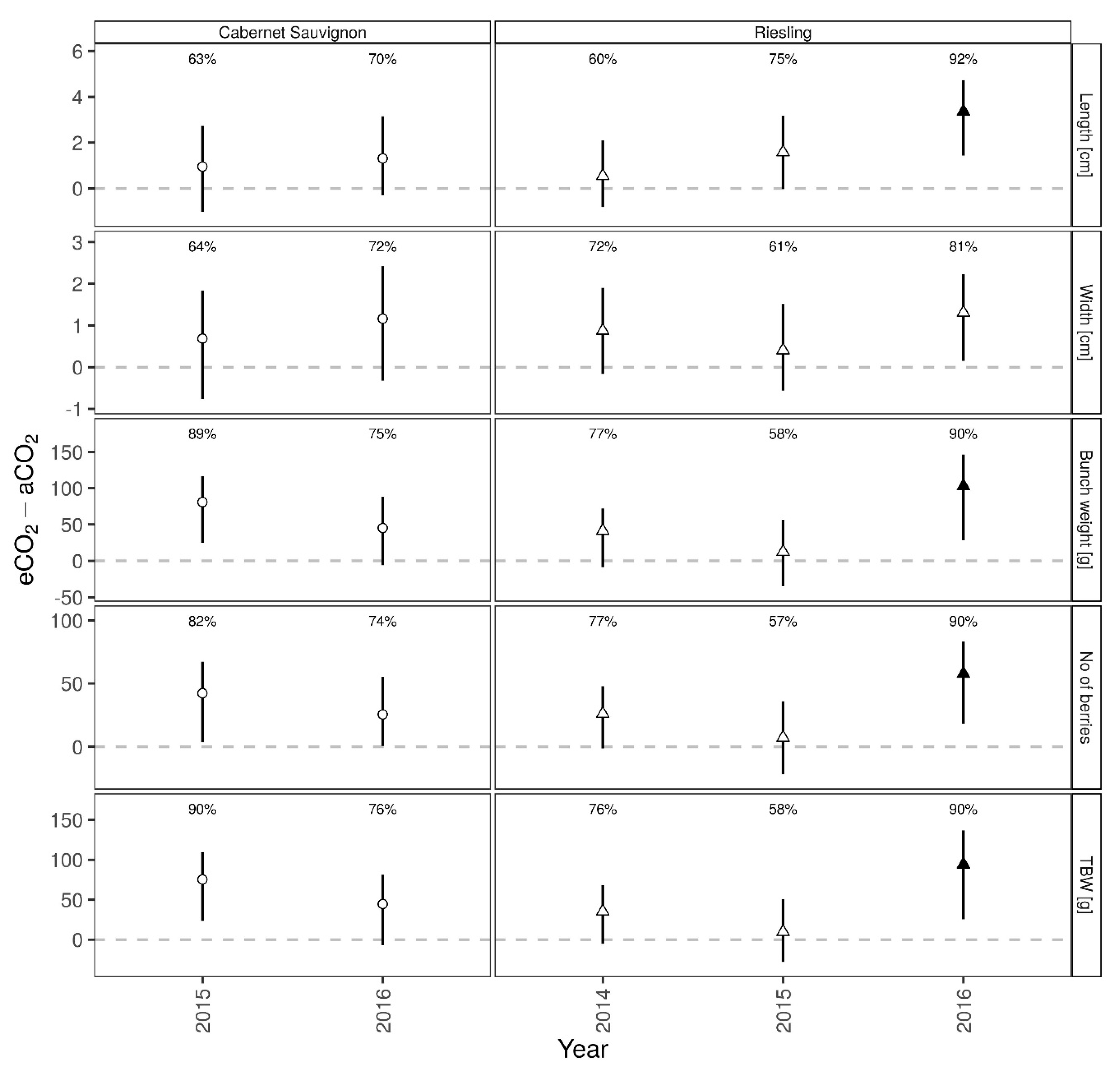
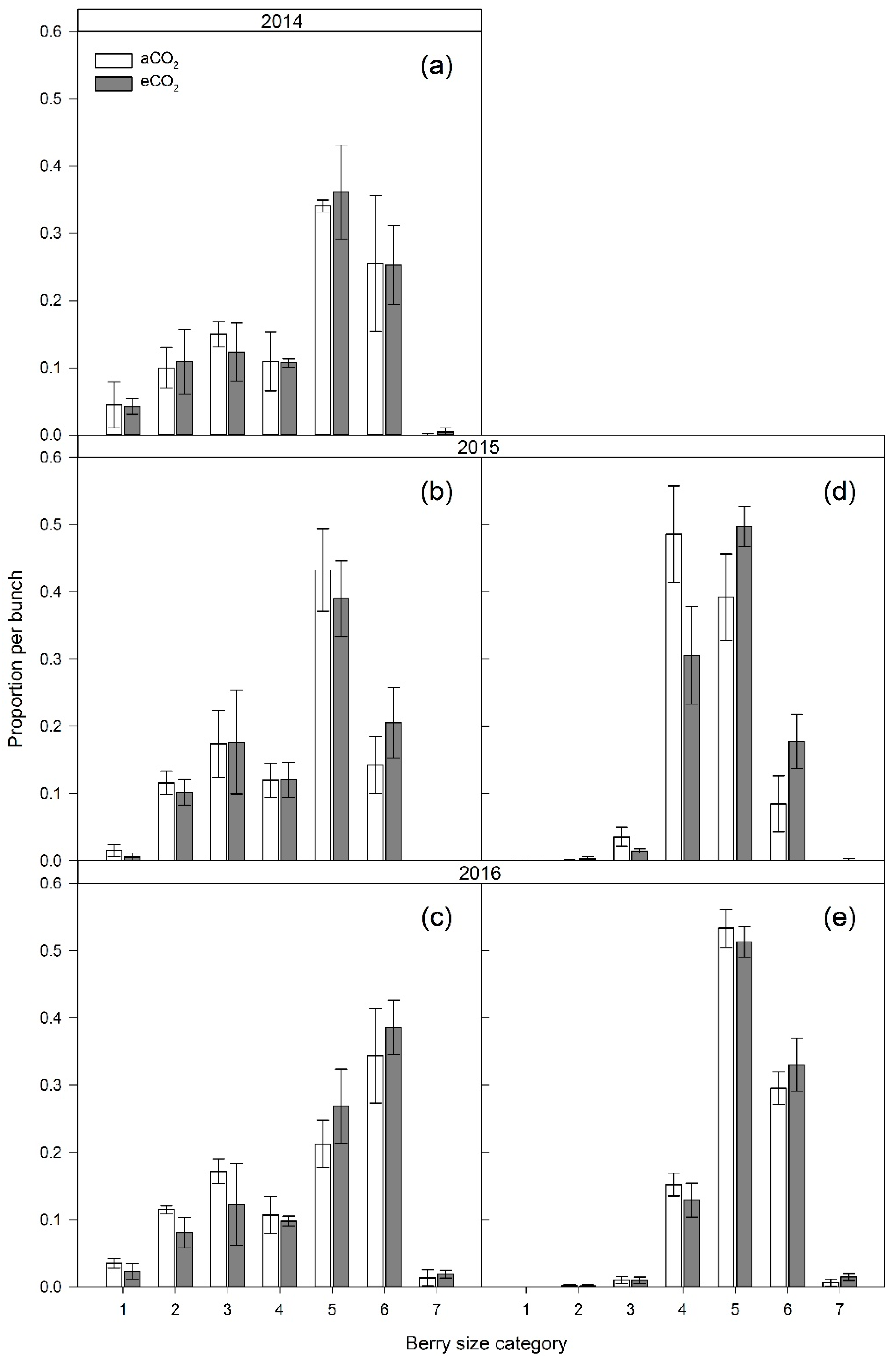
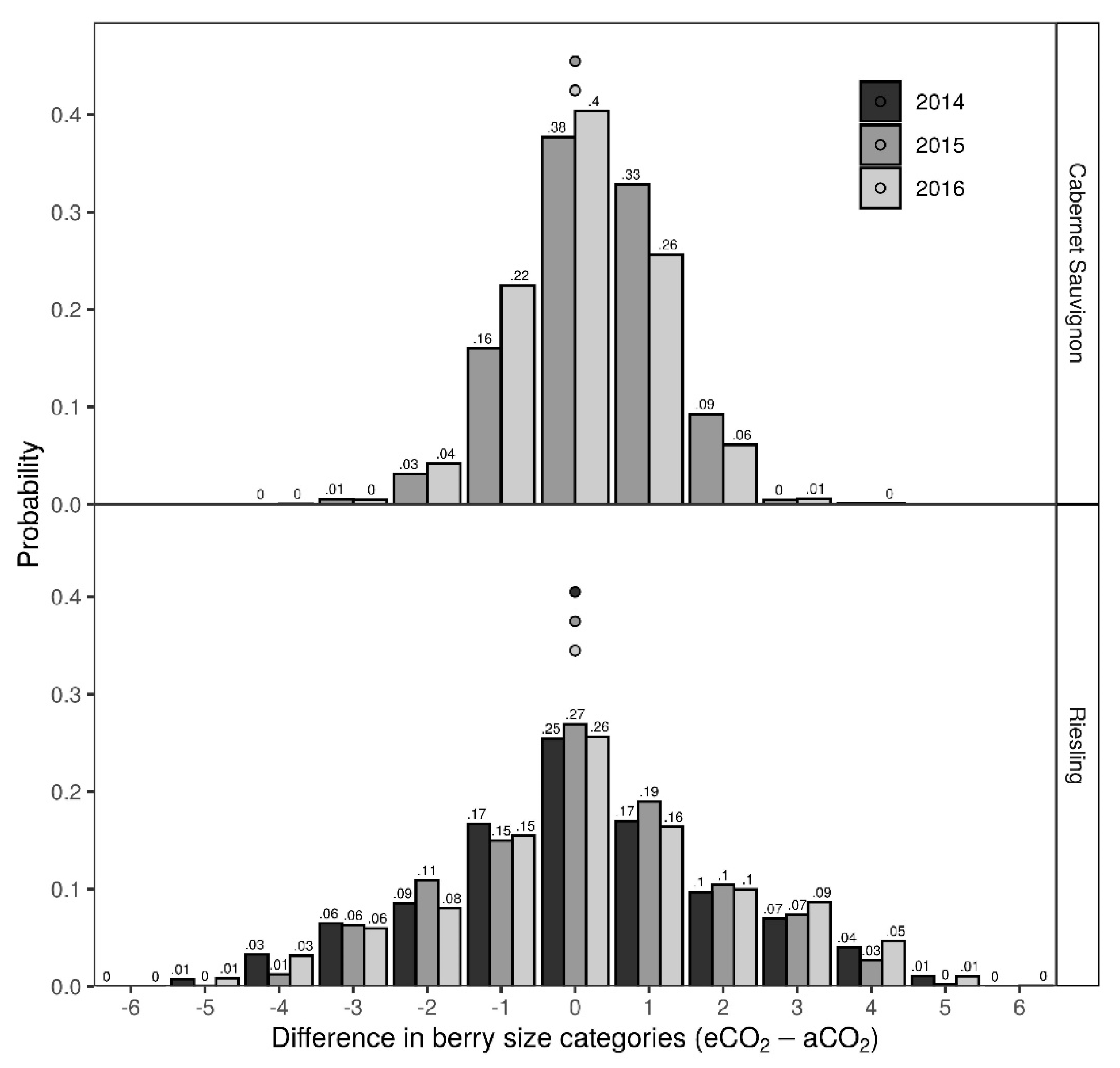
3.2. Bunch Structure Measures
4. Discussion
4.1. Berry Development
4.2. Bunch Structure Measures
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ciais, P.; Sabine, C.; Bala, G.; Bopp, L.; Brovkin, V.; Canadell, J.; Chhabra, A.A.; Defries, R.; Galloway, J.; Heimann, M.; et al. Carbon and other biogeochemical cycles. In Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.-K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK, 2013; pp. 465–570. [Google Scholar]
- Gonçalves, B.; Falco, V.; Moutinho-Pereira, J.M.; Bacelar, E.; Peixoto, F.; Correia, C. Effects of elevated CO2 on grapevine (Vitis vinifera L.): Volatile composition, phenolic content and in vitro antioxidant activity of red wine. J. Agric. Food Chem. 2009, 57, 265–273. [Google Scholar]
- Moutinho-Pereira, J.M.; Goncalves, B.; Bacelar, E.; Boaventura, C.; Coutinho, J.; Correia, C.M. Effects of elevated CO2 on grapevine (Vitis vinifera L.): Physiological and yield attributes. Vitis 2009, 48, 159–165. [Google Scholar]
- Edwards, E.J.; Unwin, D.J.; Sommer, K.J.; Downey, M.O.; Mollah, M. The response of commercially managed, field grown, grapevines (Vitis vinifera L.) to a simulated future climate consisting of elevated CO2 in combination with elevated air temperature. Acta Hortic. 2016, 1115, 103–110. [Google Scholar] [CrossRef]
- Edwards, E.J.; Unwin, D.; Kilmister, R.; Treeby, M. Multi-seasonal effects of warming and elevated CO2 on the physiology, growth and production of mature, field grown, Shiraz grapevines. Oeno One 2017, 51, 127–132. [Google Scholar] [CrossRef] [Green Version]
- Bindi, M.; Fibbi, L.; Gozzini, B.; Orlandini, S.; Seghi, L. The effect of elevated CO2 concentration on grapevine growth under field conditions. Acta Hort. 1996, 427, 325–330. [Google Scholar] [CrossRef]
- Bindi, M.; Fibbi, L.; Miglieta, F. Free air CO2 enrichment (FACE) of grapevine (Vitis vinifera L.): II. Growth and quality of grape and wine in response to elevated CO2 concentrations. Eur. J. Agron. 2001, 14, 145–155. [Google Scholar] [CrossRef]
- Bindi, M.; Raschi, A.; Lanini, M.; Miglietta, F.; Tognetti, R. Physiological and yield responses of grapevine (Vitis vinifera L.) exposed to elevated CO2 concentrations in a free air CO2 enrichment (FACE). J. Crop Improv. 2005, 13, 345–359. [Google Scholar] [CrossRef]
- Wohlfahrt, Y.; Smith, J.; Tittmann, S.; Honermeier, B.; Stoll, M. Primary productivity and physiological responses of Vitis vinifera L. cvs. under free air carbon dioxide enrichment (FACE). Eur. J. Agron. 2018, 101, 149–162. [Google Scholar] [CrossRef]
- Wohlfahrt, Y.; Collins, C.; Stoll, M. Grapevine bud fertility under conditions of elevated carbon dioxide. Oeno One 2019, 2, 303–314. [Google Scholar] [CrossRef]
- Salazar Parra, C.; Aguirreolea, J.; Sánchez-Díaz, M.; Irigoyen, J.J.; Morales, F. Effects of climate change scenarios on Tempranillo grapevine (Vitis vinifera L.) ripening: Response to a combination of elevated CO2 and temperature, and moderate drought. Plant Soil 2010, 337, 179–191. [Google Scholar] [CrossRef]
- Kizildeniz, T.; Mekni, I.; Santesteban, H.; Pascual, I.; Morales, F.; Irigoyen, J.J. Effects of climate change including elevated CO2 concentration, temperature and water deficit on growth, water status, and yield quality of grapevine (Vitis vinifera L.) cultivars. Agric. Water Manag. 2015, 159, 155–164. [Google Scholar] [CrossRef] [Green Version]
- Kizildeniz, T.; Pascual, I.; Irigoyen, J.J.; Morales, F. Using fruit-bearing cuttings of grapevine and temperature gradient greenhouses to evaluate effects of climate change (elevated CO2 and temperature, and water deficit) on the cv. red and white Tempranillo. Yield and must quality in three consecutive growing seasons (2013–2015). Agric. Water Manag. 2018, 202, 299–310. [Google Scholar]
- Singleton, V.L. Effects on red wine quality of removing juice before fermentation to simulate variation in berry size. Am. J. Enol. Vitic. 1972, 23, 106–113. [Google Scholar]
- Roby, G.; Matthews, M. Relative proportions of seed, skin and flesh, in ripe berries from Cabernet Sauvignon grapevines grown in a vineyard either well irrigated or under water deficit. Aust. J. Grape Wine Res. 2004, 10, 74–82. [Google Scholar] [CrossRef]
- Roby, G.; Harbertson, J.F.; Adams, D.A.; Matthews, M.A. Berry size and vine water deficits as factors in winegrape composition: Anthocyanins and tannins. Aust. J. Grape Wine Res. 2004, 10, 100–107. [Google Scholar] [CrossRef]
- Šuklje, K.; Lisjak, K.; Ĉesnik, H.B.; Janeš, L.; Du Toit, W.; Coetzee, Z.; Vanzo, A.; Deloire, A. Classification of grape berries according to diameter and total soluble solids to study the effect of light and temperature on methoxypyrazine, glutathione, and hydroxycinnamate evolution during ripening of Sauvignon blanc (Vitis vinifera L.). J. Agric. Food Chem. 2012, 60, 9454–9461. [Google Scholar]
- Melo, M.S.; Schultz, H.R.; Volschenk, C.G.; Hunter, J.J. Berry size variation of Vitis vinifera L. cv. Syrah: Morphological dimensions, berry composition and wine quality. S. Afr. J. Enol. Vitic. 2015, 36, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Friedel, M.; Sorrentino, V.; Blank, M.; Schuettler, A. Influence of berry diameter and colour on some determinants of wine composition of Vitis vinifera L. cv. Riesling. Aust. J. Grape Wine Res. 2016, 22, 215–225. [Google Scholar] [CrossRef]
- Chen, W.-K.; He, F.; Wang, Y.-X.; Liu, X.; Duan, C.-Q.; Wang, J. Influences of berry size on fruit composition and wine quality of Vitis vinifera L. cv. Cabernet Sauvignon grapes. S. Afr. J. Enol. Vitic. 2018, 39, 67–76. [Google Scholar] [CrossRef] [Green Version]
- Ziegler, M.; Wegmann-Herr, P.; Schmarr, H.-G.; Gök, R.; Winterhalter, P.; Fischer, U. Impact of rootstock, clonal selection and berry size of Vitis vinifera sp. Riesling on the formation of TDN, vitispiranes and other volatile compounds. J. Agric. Food Chem. 2020, 68, 3834–3849. [Google Scholar] [CrossRef]
- Ito, J.; Hasegawa, S.; Fujita, K.; Ogasawara, S.; Fujiwara, T. Effect of CO2 enrichment on fruit growth and quality in Japanese pear (Pyrus serotina Reheder cv. Kosui). Soil Sci. Plant Nutr. 1999, 45, 385–393. [Google Scholar] [CrossRef] [Green Version]
- Han, J.-H.; Cho, J.G.; Son, I.-C.; Kim, S.H.; Lee, I.-B.; Choi, I.M.; Kim, D. Effects of elevated carbon dioxide and temperature on photosynthesis and fruit characteristics of ‘Niitaka’ pear (Pyrus pyrifolia Nakai). Hort. Env. Biotechnol. 2012, 53, 357–361. [Google Scholar] [CrossRef]
- Idso, S.B.; Kimball, B.A. Effects of long-term atmospheric CO2 enrichment on the growth and fruit production of sour orange trees. Glob. Chang. Biol. 1997, 3, 89–96. [Google Scholar] [CrossRef]
- Sung, F.J.M.; Chen, J.J. Gas exchange rate and yield response of strawberry to carbon dioxide enrichment. Sci. Hortic. 1991, 48, 241–251. [Google Scholar] [CrossRef]
- Wang, S.Y.; Bunce, J.A. Elevated carbon dioxide affects fruit flavor in field-grown strawberries (Fragaria × Ananassa Duch). J. Sci. Food Agric. 2004, 84, 1464–1468. [Google Scholar] [CrossRef]
- Mochizuki, M.J.; Daugovish, O.; Ahumada, M.H.; Ashkan, S.; Lovatt, C.J. Carbon dioxide enrichment may increase yield of field-grown red raspberry under high tunnels. HortTechnology 2010, 20, 213–219. [Google Scholar] [CrossRef] [Green Version]
- Cozzolino, D.; Cynkar, W.U.; Dambergs, R.G.; Gishen, M.; Smith, P. Grape (Vitis vinifera) compositional data spanning ten successive vintages in the context of abiotic growing parameters. Agric. Ecosyst. Env. 2010, 139, 565–570. [Google Scholar] [CrossRef]
- Schultz, H.R. Climate change and viticulture: A European perspective on climatology, carbon dioxide and UV-B effects. Aust. J. Grape Wine Res. 2000, 6, 2–12. [Google Scholar] [CrossRef]
- Ben Ghozlen, N.; Cerovic, Z.G.; Germain, C.; Toutain, S.; Latouche, G. Non-destructive optical monitoring of grape maturation by proximal sensing. Sensors 2010, 10, 10040–10068. [Google Scholar] [CrossRef]
- Agati, G.; D’Onofrio, C.; Ducci, E.; Cuzzola, A.; Remorini, D.; Tuccio, L.; Lazzini, F.; Mattii, G. Potential of a multiparametric optical sensor for determining in situ the maturity components or red and white Vitis vinifera wine grapes. J. Agric. Food Chem. 2013, 61, 12211–12218. [Google Scholar] [CrossRef]
- Dukes, B.C.; Butzke, C.E. Rapid determination of primary amino acids in grape juice using an o-Phthaldialdehyde/N-Acetyl-L-Cysteine spectrophotometric assay. Am. J. Enol. Vitic. 1998, 49, 125–134. [Google Scholar]
- Schneider, A.; Gerbi, V.; Redoglia, M. A rapid HPLC method for separation and determination of major organic acids in grape musts and wines. Am. J. Enol. Vitic. 1987, 38, 151–155. [Google Scholar]
- Knoll, C.; Fritsch, S.; Schnell, S.; Grossmann, M.; Rauhut, D.; du Toit, M. Influence of pH and ethanol on malolactic fermentation and volatile aroma compound composition in white wines. Lwt. Food Sci. Technol. 2011, 44, 2077–2086. [Google Scholar] [CrossRef]
- The R Project for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 4 December 2019).
- Bürkner, P.-C. Brms: An R package for bayesian multilevel models using stan. J. Stat. Softw. 2017, 80, 1–28. [Google Scholar] [CrossRef] [Green Version]
- Bürkner, P.-C. Advanced bayesian multilevel modeling with the R package brms. R. J. 2018, 10, 395–411. [Google Scholar] [CrossRef]
- Carpenter, B.; Gelman, A.; Hoffman, M.D.; Lee, D.; Goodrich, B.; Betancourt, M.; Brubaker, M.; Guo, J.; Li, P.; Riddell, A. Stan: A probabilistic programming language. J. Stat. Softw. 2017, 76, 1–32. [Google Scholar] [CrossRef] [Green Version]
- Kaur, A.; Gregori, D.; Patil, G.P.; Taillie, C. Ecological applications of generalized linear models and quasi-likelihood methods–An overview. Stat. Appl. 1996, 8, 59–82. [Google Scholar]
- Bürkner, P.-C.; Vuorre, M. Ordinal regression models in psychology: A tutorial. Adv. Methods Pract. Psychol. Sci. 2019, 2, 77–101. [Google Scholar] [CrossRef] [Green Version]
- Vehtari, A.; Gelman, A.; Simpson, D.; Carpenter, B.; Bürkner, P.-C. Rank-normalization, folding, and localization: An improved $\widehat{R}$ for assessing convergence of MCMC. arxiv 2019, arXiv:1903.08008. [Google Scholar]
- Flegal, J.M.; Hughes, J.; Vats, D.; Dai, N. mcmcse: Monte Carlo Standard Errors for MCMC. Available online: https://rdrr.io/cran/mcmcse/ (accessed on 4 December 2019).
- Vats, D.; Flegal, J.M.; Jones, G.L. Multivariate output analysis for Markov chain Monte Carlo. Biometrika 2019, 106, 321–337. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Lüscher, J.; Kizildeniz, T.; Vucetic, V.; Dai, Z.; Luedeling, E.; Van Leeuwen, C.; Gomes, E.; Pascual, I.; Irigoyen, J.J.; Morales, F.; et al. Sensitivity of grapevine phenology to water availability, temperature and CO2 concentration. Front. Env. Sci. 2016, 4, 48. [Google Scholar] [CrossRef]
- Mira de Orduna, R. Climate change associated effects on grape and wine quality and production. Food Res. Int. 2010, 43, 1844–1855. [Google Scholar] [CrossRef]
- Coombe, B.G. Influence of temperature on composition and quality of grapes. Acta Hort. 1987, 206, 23–35. [Google Scholar] [CrossRef]
- Ollat, N.; Diakou-Verdin, P.; Carde, J.-P.; Barrieu, F.; Gaudillère, J.-P.; Moing, A. Grape berry development: A review. J. Int. Sci. Vigne Vin 2002, 36, 109–131. [Google Scholar]
- Keller, M. Managing grapevines to optimise fruit development in a challenging environment: A climate change primer for viticulturists. Aust. J. Grape Wine Res. 2010, 16, 56–69. [Google Scholar] [CrossRef]
- Jackson, D.I.; Lombard, P.B. Environmental and management practices affecting grape composition and wine quality—A review. Am. J. Enol. Vitic. 1993, 44, 409–430. [Google Scholar]
- Kliewer, W.M.; Bledsoe, A.M. Influence of hedging and leaf removal on canopy microclimate, grape composition, and wine quality under California conditions. Acta Hort. 1987, 206, 157–168. [Google Scholar] [CrossRef]
- Zoecklein, B.W.; Wolf, T.K.; Duncan, N.W.; Judge, J.M.; Cook, M.K. Effects of fruit zone leaf removal on yield, fruit composition, and fruit rot incidence of chardonnay and white riesling (Vitis vinifera L.) grapes. Am. J. Enol. Vitic. 1992, 43, 139–148. [Google Scholar]
- Smart, R.E. Principles of grapevine canopy microclimate manipulation with implications for yield and quality. A review. Am. J. Enol. Vitic. 1985, 36, 230–239. [Google Scholar]
- Lakso, A.N.; Kliewer, W.M. The influence of temperature on malic acid metabolism in grape berries: I. Enzyme responses. Plant. Physiol. 1975, 56, 370–372. [Google Scholar] [CrossRef] [Green Version]
- Ruffner, H.P. Metabolism of tartaric and malic acids in Vitis: A review—part B. Vitis 1982, 21, 346–358. [Google Scholar]
- Etienne, A.; Genard, M.; Lobit, P.; Mbeguie-A.-Mbeguie, D.; Bugaud, C. What controls fleshy fruit acidity? A review of malate and citrate accumulation in fruit cells. J. Exp. Bot. 2013, 64, 1451–1469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greer, D.H.; Weston, C. Heat stress affects flowering, berry growth, sugar accumulation and photosynthesis of Vitis vinifera cv. Semillon grapevines grown in a controlled environment. Funct. Plant. Biol. 2010, 37, 206–214. [Google Scholar] [CrossRef]
- Tello, J.; Ibanez, J. What do we know about grapevine bunch compactness? A state-of-the-art review. Aust. J. Grape Wine Res. 2018, 24, 6–23. [Google Scholar] [CrossRef]
- Vail, M.E.; Marois, J.J. Grape cluster architecture and the susceptibility of berries to Botrytis cinerea. Phytopathology 1991, 81, 188–191. [Google Scholar] [CrossRef]
- Vail, M.E.; Wolpert, J.A.; Gubler, W.D.; Rademacher, M.R. Effect of cluster tightness on botrytis bunch rot in six Chardonnay clones. Plant. Dis. 1998, 82, 107–109. [Google Scholar] [CrossRef] [Green Version]
- Alonso-Villaverde, V.; Boso, S.; Santiago, J.L.; Gago, P.; Martínez, M.C. Relationship between susceptibility to botrytis bunch rot and grape cluster morphology in the Vitis vinifera L. cultivar Albariño. Int. J. Fruit Sci. 2008, 8, 251–265. [Google Scholar] [CrossRef]
- Hed, B.; Ngugi, H.K.; Travis, J.W. Relationship between cluster compactness and bunch rot in vignoles grapes. Plant. Dis. 2009, 93, 1195–1201. [Google Scholar] [CrossRef] [Green Version]
- Simpson, R.F. 1,1,6-Trimethyl-1,2-dihydronaphthalene: An important contributor to the bottle aged bouquet of wine. Chem. Ind. 1978, 1, 37. [Google Scholar]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vegetation Period | Mean Daily Max. Temperature (°C) | Mean Daily Min. Temperature (°C) | No. of Heat Days (>30 °C) | Precipitation (mm) |
|---|---|---|---|---|
| 2014 | 21.6 | 11.3 | 10 | 441 |
| 2015 | 21.5 | 10.5 | 26 | 227 |
| 2016 | 21.5 | 10.5 | 17 | 371 |
| Cultivar | Date | Tartaric Acid (g∙L−1) | Malic Acid (g∙L−1) | Citric Acid (g∙L−1) | Shikimic Acid (mg∙L−1) | ||||
| R | 2014 | aCO2 | eCO2 | aCO2 | eCO2 | aCO2 | eCO2 | aCO2 | eCO2 |
| 11/08 | 13.86 ± 0.32 | 13.84 ± 0.29 | 19.12 ± 0.43 | 19.38 ± 0.42 | 0.28 ± 0.01 | 0.30 ± 0.02 | 15.2 ± 1.7 | 14.6 ± 0.8 | |
| 25/08 | 12.17 ± 0.11 | 11.58 ± 0.48 | 12.53 ± 0.24 | 12.19 ± 0.25 | 0.21 ± 0.01 | 0.21 ± 0.02 | 26.4 ± 0.7 | 28.8 ± 2.0 | |
| 08/09 | 10.67 ± 0.11 | 10.29 ± 0.10 | 6.14 ± 0.15 | 6.17 ± 0.16 | 0.17 ± 0.02 | 0.18 ± 0.02 | 33.9 ± 2.0 | 36.1 ± 1.3 | |
| 15/09 | 9.91 ± 0.14 | 9.75 ± 0.12 | 4.87 ± 0.02 | 4.92 ± 0.12 | 0.18 ± 0.02 | 0.17 ± 0.00 | 34.0 ± 3.1 | 35.9 ± 1.2 | |
| 22/09 | 9.31 ± 0.08 | 9.23 ± 0.08 | 3.87 ± 0.31 | 3.96 ± 0.16 | 0.17 ± 0.01 | 0.18 ± 0.00 | 34.3 ± 2.8 | 36.8 ± 1.8 | |
| 2015 | |||||||||
| 10/08 | 17.21 ± 0.15 | 15.91 ± 0.51 | 21.10 ± 0.34 | 20.53 ± 0.64 | 0.38 ± 0.02 | 0.38 ± 0.02 | 22.4 ± 3.2 | 19.3 ± 0.6 | |
| 17/08 | 16.57 ± 0.51 | 15.02 ± 0.30 | 19.96 ± 1.22 | 19.48 ± 0.67 | 0.42 ± 0.02 | 0.39 ± 0.01 | 21.9 ± 2.3 | 21.8 ± 1.6 | |
| 31/08 | 12.42 ± 0.05 | 11.50 ± 0.20 | 9.40 ± 0.09 | 9.43 ± 0.17 | 0.21 ± 0.00 | 0.21 ± 0.00 | 33.3 ± 1.2 | 35.0 ± 1.0 | |
| 14/09 | 10.31 ± 0.45 | 10.01 ± 0.39 | 5.17 ± 0.37 | 5.97 ± 0.44 | 0.18 ± 0.01 | 0.16 ± 0.00 | 33.8 ± 2.7 | 35.3 ± 1.1 | |
| 28/09 | 10.02 ± 0.24 | 9.67 ± 0.08 | 4.15 ± 0.26 | 4.61 ± 0.34 | 0.17 ± 0.02 | 0.17 ± 0.02 | 34.0 ± 2.5 | 35.6 ± 1.2 | |
| 2016 | |||||||||
| 22/08 | 14.56 ± 0.50 | 13.94 ± 0.10 | 22.11 ± 0.80 | 21.96 ± 0.39 | 0.29 ± 0.01 | 0.30 ± 0.01 | 21.7 ± 1.3 | 22.5 ± 0.8 | |
| 05/09 | 10.71 ± 0.22 | 10.35 ± 0.11 | 9.07 ± 0.95 | 9.76 ± 0.89 | 0.16 ± 0.03 | 0.16 ± 0.01 | 38.3 ± 3.8 | 36.3 ± 1.7 | |
| 19/09 | 8.96 ± 0.10 | 8.86 ± 0.04 | 3.96 ± 0.24 | 4.72 ± 0.50 | 0.12 ± 0.00 | 0.14 ± 0.01 | 36.7 ± 3.0 | 35.6 ± 0.7 | |
| 04/10 | 8.38 ± 0.24 | 8.29 ± 0.10 | 3.07 ± 0.02 | 3.45 ± 0.35 | 0.12 ± 0.02 | 0.14 ± 0.01 | 36.6 ± 1.2 | 37.4 ± 1.1 | |
| Cultivar | Date | Tartaric Acid (g∙L−1) | Malic Acid (g∙L−1) | Citric Acid (g∙L−1) | Shikimic Acid (mg∙L−1) | ||||
| CS | 2014 | aCO2 | eCO2 | aCO2 | eCO2 | aCO2 | eCO2 | aCO2 | eCO2 |
| 06/08 | 13.45 ± 0.38 | 13.41 ± 0.34 | 24.85 ± 0.71 | 24.42 ± 1.26 | 0.49 ± 0.02 | 0.50 ± 0.02 | 27.1 ± 3.1 | 27.0 ± 2.0 | |
| 18/08 | 11.37 ± 0.23 | 11.63 ± 0.43 | 20.30 ± 0.67 | 20.85 ± 0.72 | 0.36 ± 0.02 | 0.37 ± 0.02 | 44.5 ± 3.3 | 44.2 ± 3.8 | |
| 01/09 | 8.68 ± 0.16 | 8.69 ± 0.17 | 10.56 ± 0.11 | 11.27 ± 0.92 | 0.25 ± 0.01 | 0.25 ± 0.01 | 62.7 ± 3.2 | 59.7 ± 3.4 | |
| 15/09 | 7.42 ± 0.30 | 7.55 ± 0.32 | 6.52 ± 0.45 | 6.72 ± 0.29 | 0.20 ± 0.01 | 0.21 ± 0.01 | 79.1 ± 3.7 | 77.2 ± 3.8 | |
| 29/09 | 7.20 ± 0.18 | 7.42 ± 0.11 | 4.97 ± 0.40 | 4.83 ± 0.24 | 0.20 ± 0.01 | 0.19 ± 0.01 | 86.5 ± 5.3 | 82.7 ± 6.4 | |
| 2015 | |||||||||
| 12/08 | 14.99 ± 0.52 | 14.47 ± 0.30 | 24.11 ± 0.70 | 24.96 ± 0.20 | 0.54 ± 0.04 | 0.56 ± 0.02 | 45.1 ± 9.1 | 36.6 ± 3.9 | |
| 25/08 | 13.20 ± 0.15 | 12.61 ± 0.24 | 17.15 ± 2.25 | 20.13 ± 1.42 | 0.41 ± 0.01 | 0.43 ± 0.02 | 65.3 ± 6.8 | 59.6 ± 1.7 | |
| 07/09 | 9.50 ± 0.30 | 9.35 ± 0.23 | 7.45 ± 1.05 | 9.17 ± 1.05 | 0.20 ± 0.01 | 0.22 ± 0.01 | 71.4 ± 3.5 | 70.2 ± 1.4 | |
| 22/09 | 8.44 ± 0.44 | 8.39 ± 0.16 | 4.77 ± 0.58 | 6.08 ± 0.39 | 0.20 ± 0.01 | 0.22 ± 0.01 | 68.2 ± 6.5 | 66.4 ± 1.5 | |
| 05/10 | 8.08 ± 0.38 | 8.03 ± 0.20 | 3.95 ± 0.34 | 4.97 ± 0.27 | 0.20 ± 0.01 | 0.21 ± 0.02 | 64.8 ± 5.6 | 64.0 ± 2.9 | |
| 2016 | |||||||||
| 29/08 | 9.94 ± 0.11 | 9.51 ± 0.16 | 14.17 ± 1.83 | 14.43 ± 1.44 | 0.26 ± 0.03 | 0.27 ± 0.03 | 68.1 ± 4.5 | 70.5 ± 2.4 | |
| 12/09 | 8.44 ± 0.13 | 8.36 ± 0.07 | 6.97 ± 0.80 | 7.38 ± 0.73 | 0.17 ± 0.02 | 0.18 ± 0.01 | 77.1 ± 5.4 | 78.1 ± 3.4 | |
| 26/09 | 7.21 ± 0.17 | 7.40 ± 0.16 | 4.72 ± 0.54 | 5.14 ± 0.71 | 0.15 ± 0.02 | 0.16 ± 0.01 | 78.0 ± 6.7 | 77.9 ± 4.0 | |
| 10/10 | 7.00 ± 0.20 | 7.20 ± 0.04 | 3.96 ± 0.35 | 4.32 ± 0.60 | 0.15 ± 0.02 | 0.17 ± 0.02 | 82.2 ± 8.2 | 83.3 ± 7.0 | |
| Cultivar | Year | Length (cm) | Width (cm) | Bunch Weight (g) | No of Berries | Total Berry Weight (g) | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| R | aCO2 | eCO2 | aCO2 | eCO2 | aCO2 | eCO2 | aCO2 | eCO2 | aCO2 | eCO2 | |
| 2014 | 9.92 ± 0.19 | 10.46 ± 0.31 | 6.29 ± 0.95 | 7.17 ± 0.89 | 108.14 ± 5.90 | 151.88 ± 9.11 | 85.9 ± 8.20 | 112.7 ± 1.8 | 100.26 ± 15.36 | 138.77 ± 7.24 | |
| % | +5.4 | +14.0 | +40.4 | +31.2 | +38.4 | ||||||
| 2015 | 10.56 ± 0.24 | 12.19 ± 0.82 | 6.69 ± 0.65 | 7.08 ± 0.22 | 143.67 ± 7.35 | 158.40 ± 28.82 | 113.6 ± 9.5 | 122.1 ± 18.4 | 125.18 ± 6.47 | 137.86 ± 25.29 | |
| % | +15.4 | +5.8 | +10.3 | +7.5 | +10.1 | ||||||
| 2016 | 9.67 ± 0.22 | 13.06 ± 0.97 | 6.00 ± 0.22 | 7.33 ± 0.51 | 130.07 ± 13.09 | 237.98 ± 16.46 | 97.2 ± 10.8 | 156.3 ± 3.1 | 121.06 ± 12.95 | 222.02 ± 15.90 | |
| % | +35.1 | +22.2 | +83.0 | +60.8 | +83.4 | ||||||
| CS | 2015 | 15.58 ± 0.46 | 16.50 ± 0.36 | 6.81 ± 1.00 | 7.47 ± 0.39 | 136.81 ± 20.38 | 219.00 ± 3.97 | 110.9 ± 9.2 | 154.4 ± 3.9 | 127.79 ± 19.75 | 205.48 ± 4.22 |
| % | +5.9 | +9.7 | +60.1 | +39.2 | +60.8 | ||||||
| 2016 | 14.31 ± 0.17 | 15.64 ± 0.21 | 6.86 ± 0.24 | 8.08 ± 0.66 | 164.04 ± 10.95 | 212.15 ± 11.11 | 102.1 ± 4.9 | 128.9 ± 4.6 | 153.95 ± 9.12 | 200.40 ± 10.68 | |
| % | +9.3 | +17.8 | +29.3 | +26.2 | +30.2 | ||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wohlfahrt, Y.; Tittmann, S.; Schmidt, D.; Rauhut, D.; Honermeier, B.; Stoll, M. The Effect of Elevated CO2 on Berry Development and Bunch Structure of Vitis vinifera L. cvs. Riesling and Cabernet Sauvignon. Appl. Sci. 2020, 10, 2486. https://0-doi-org.brum.beds.ac.uk/10.3390/app10072486
Wohlfahrt Y, Tittmann S, Schmidt D, Rauhut D, Honermeier B, Stoll M. The Effect of Elevated CO2 on Berry Development and Bunch Structure of Vitis vinifera L. cvs. Riesling and Cabernet Sauvignon. Applied Sciences. 2020; 10(7):2486. https://0-doi-org.brum.beds.ac.uk/10.3390/app10072486
Chicago/Turabian StyleWohlfahrt, Yvette, Susanne Tittmann, Dominik Schmidt, Doris Rauhut, Bernd Honermeier, and Manfred Stoll. 2020. "The Effect of Elevated CO2 on Berry Development and Bunch Structure of Vitis vinifera L. cvs. Riesling and Cabernet Sauvignon" Applied Sciences 10, no. 7: 2486. https://0-doi-org.brum.beds.ac.uk/10.3390/app10072486