Metabolic Syndrome-Preventive Peptides Derived from Milk Proteins and Their Presence in Cheeses: A Review



Faculty of Food Science, University of Warmia and Mazury in Olsztyn, Pl. Cieszyński 1, 10-726 Olsztyn-Kortowo, Poland
*
Authors to whom correspondence should be addressed.
Appl. Sci. 2020, 10(8), 2772; https://0-doi-org.brum.beds.ac.uk/10.3390/app10082772
Submission received: 11 March 2020
/
Revised: 10 April 2020
/
Accepted: 14 April 2020
/
Published: 17 April 2020
(This article belongs to the Special Issue New Trends in Environmental Engineering, Agriculture, Food Production, and Analysis)
Abstract
:The metabolic syndrome (MetS) is defined as the occurrence of diet-related diseases such as abdominal obesity, atherogenic dyslipidemia, hyperglycemia (insulin resistance) and hypertension. Milk-derived peptides are well-known agents acting against high blood pressure, blood glucose level, and lipoprotein disproportion. The aim of this review are metabolic syndrome-preventive peptides derived from milk proteins which were identified in cheeses. Special attention was paid to the sequences acting as angiotensin converting enzyme (ACE), dipeptidyl peptidase IV (DDP4), and α-glucosidase inhibitors, as well as antioxidative, hypocholesterolemic, antiobesity, and anti-inflammatory agents. Some results of meta-analyses concerning the consumption of cheese and the risk of MetS diseases were also presented.

1. Introduction
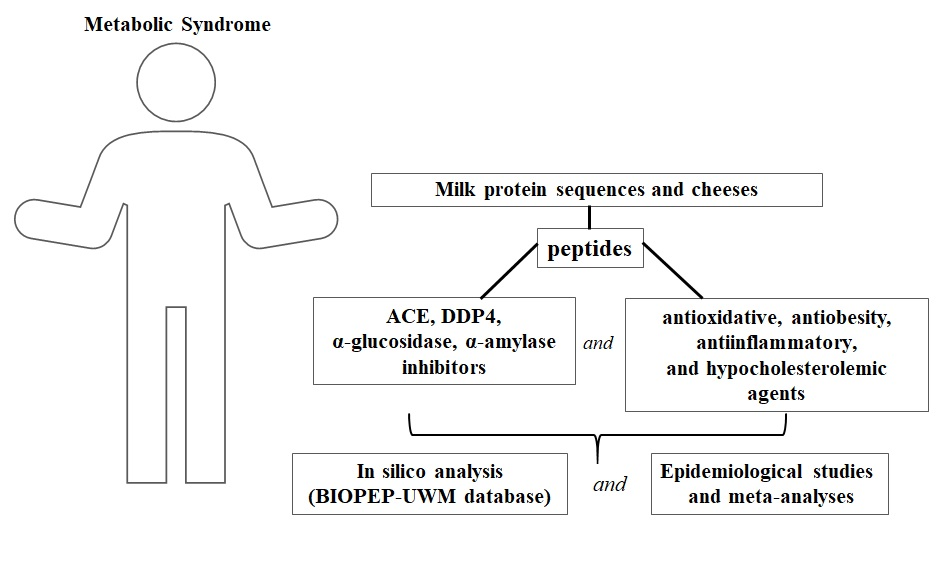
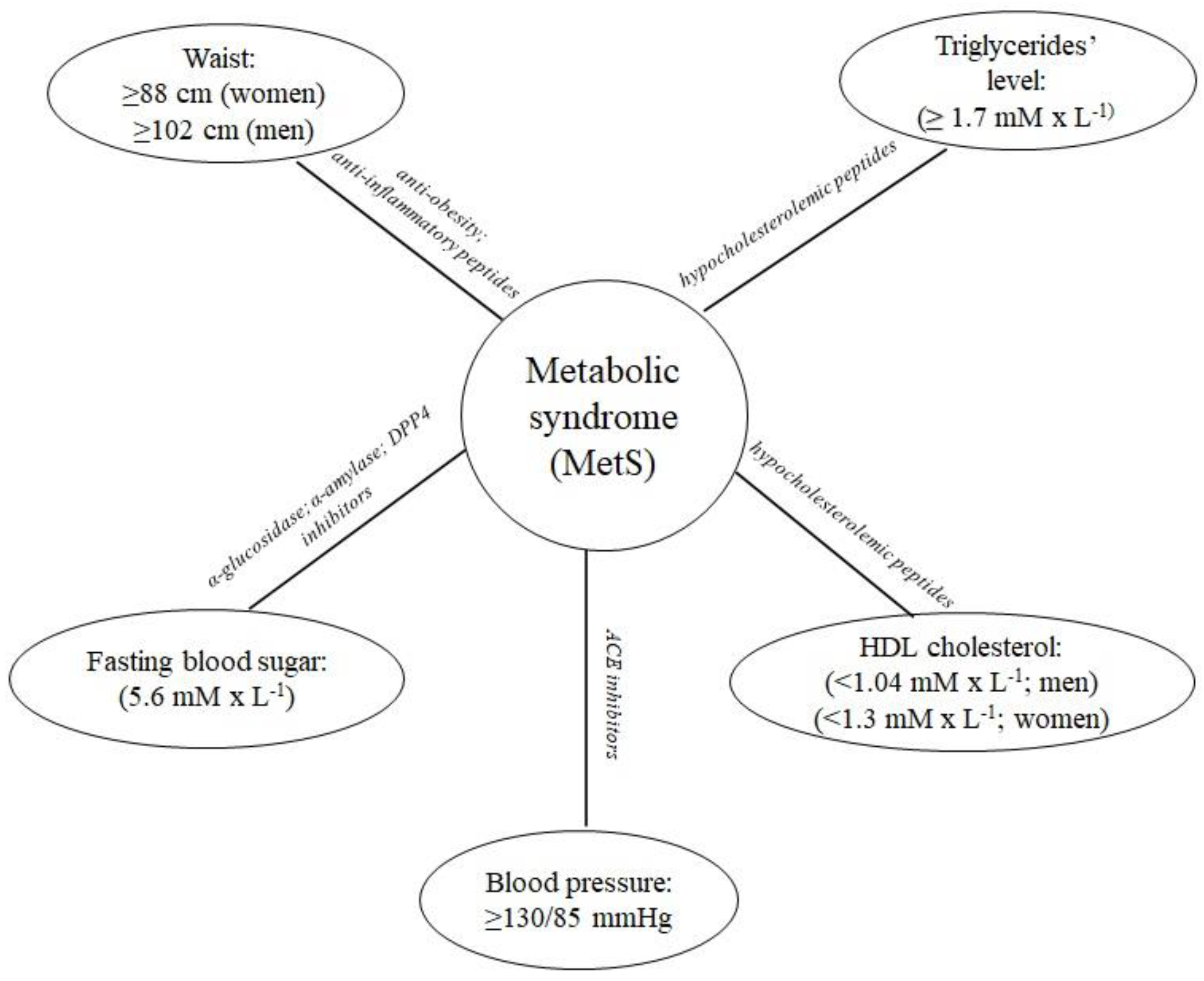
The metabolic syndrome (MetS) is defined as a clustering of four major cardiovascular risk factors, namely: abdominal obesity, atherogenic dyslipidemia, hyperglycemia (insulin resistance), and hypertension (Figure 1) [1]. Moreover, the pathogenesis of MetS involves both genetic and acquired factors contributing to the inflammation [2]. MetS also represents a significant risk factor for the development of cardiovascular disease (CVD), type 2 diabetes mellitus (T2D), and all-cause mortality [1]. Moore et al. [3] analyzed data derived from the National Health and Nutrition Examination Survey (NHANES). Based on the analysis including the period from 1988 to 2012, MetS was defined as at least three conditions from these listed further that have to occur in a patient: elevated waist circumference, elevated level of triglycerides, reduced high-density lipoprotein cholesterol level, high blood pressure, and elevated fasting blood glucose level. According to the above-mentioned definition of MetS, in 2012 there were more than 66 million adults in the US with recognized MetS symptoms. They were observed in every sociodemographic group [3].
The following enzymes are involved in MetS: dipeptidyl peptidase IV (EC 3.4.14.5; DPP4), α-glucosidase (EC 3.2.1.20), α-amylase (EC 3.2.1.1), and angiotensin-converting enzyme (EC 3.4.15.1; ACE). The first three affect the postprandial blood glucose level, whereas ACE is the key enzyme involved in the regulation of the blood pressure. Thus, peptides being the inhibitors of the above-mentioned enzymes may be considered anti-MetS agents [5]. Additionally, peptides also reduce cholesterol level, antioxidative stress [6], act as anti-obesity [7], and anti-inflammatory agents [8].
Milk is the first functional food for every mammalian species. The contents of its nutrients, such as proteins (and their proportions), fat, sugar, vitamins, and minerals, vary between species as well as breeds. Moreover, milk composition is affected by feeding methods, age of an animal, and many other [9]. Milk and dairy products were also studied as anti-metabolic syndrome foods [10].
A cohort study made in 2018 on 136,384 individuals from 21 countries on five continents demonstrated that dairy product consumption (especially of milk and yogurt) can be associated with a lower risk, of not only CVD, but also mortality. According to this study, even one serving of milk or yogurt significantly lowered the risks of the above-mentioned events. Additionally, the higher consumption of dairy products can be associated with significantly lower risk of stroke [10]. According to another study, every 200 mL increase of daily milk consumption reduced the risk of CVD by 6%. Based on the data from 2015 it was shown that three main milk constituents were responsible for the MetS-preventive activity, namely calcium, fat, and proteins [11]. One of the MetS-preventive potentials of milk and dairy products resulting from the presence of proteins might be related to the action of biologically active peptides [5]. It also reflects vast information found in. e.g., Scopus database [www.scopus.com]. Providing the following words as the input query “ALL (metabolic AND syndrome AND milk AND peptides)” revealed 7459 hits (accessed in September 2019).
The first mentions about cheese manufacturing appeared around 5500 BC. Early adult humans were unable to digest lactose. Thus, making cheese was a good way to consume health beneficial nutrients derived from milk, without getting ill [11]. The casein, main milk protein, during cheese maturation is hydrolyzed into a variety of peptides. Their composition varies and depends on the activity of endogenous proteases and peptidases of milk, rennet, starter culture, and microbial flora [12]. Bovine casein has been shown to possess a variety of bioactivities, including MetS-preventive ones. In silico analyses and reviews conducted on bovine casein confirmed the casein-derived peptides to exhibit these activities [13,14]. Hence, considering the increasing role of biopeptides in the prevention of MetS, the aim of this paper was to describe MetS-preventive peptides derived mainly from bovine milk proteins which were also identified in the ripened cheeses. The main attention was paid to the peptides exerting antidiabetic, antihypertensive, antioxidative, cholesterol-reducing, antiobesity, and anti-inflammatory effects.
2. Antidiabetic Agents
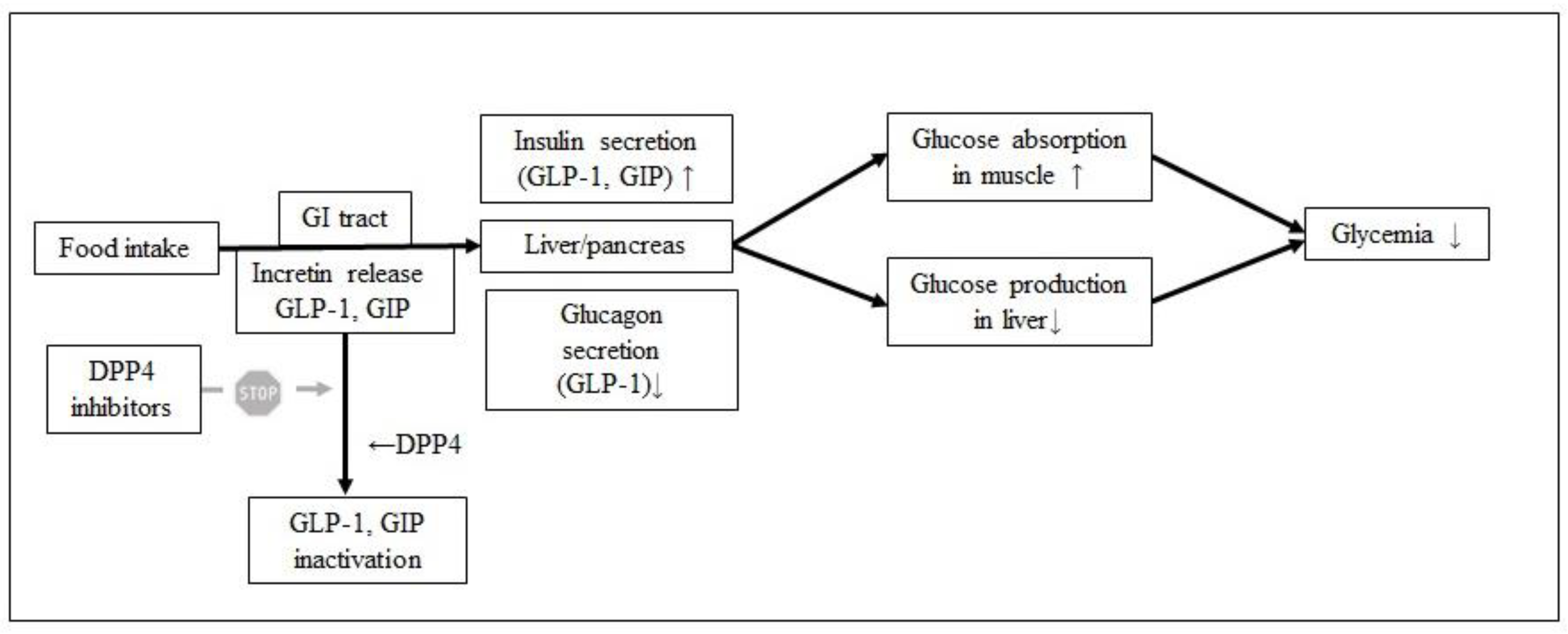
One of the body enzymes involved in the regulation of the glucose level is DPP4 [15]. This enzyme is involved in the incretin effect after meal consumption. Incretins, like glucagon-like peptide 1 (GLP-1) and glucose-dependent insulinotropic polypeptide (GIP), are hormones that decrease blood glucose levels by stimulating insulin secretion and inhibiting glucagon release. GIP serves several additional functions including promotion of growth and survival of the pancreatic beta-cell and stimulation of adipogenesis [15]. This effect is hindered by DPP4, therefore inhibition of DPP4 prolongs the incretin effect and lowers the postprandial blood glucose level (Figure 2) [16].
Uenishi et al. [17] identified DPP4 inhibitory peptides in water-soluble Gouda cheese extracts. Peptides derived from β-casein (LPQNIPPL) and αS1-, αS2-, β-caseins (LPQ) evoked the highest DPP4-inhibiting effect in vitro. Their IC50 values were 46.0 and 56.7 µM, respectively. The bioactivity of the LPQNIPPL sequence was also tested on rats (Female Sprague Dawley, 8-week-old). The administration of this peptide at 300 mg × kg−1 body weight reduced significantly the postprandial glucose level in those rats [16]. Other peptides with DPP4 inhibitory bioactivity that were identified in Gouda cheese water-soluble-extracts (WSE) were characterized with the following sequences: VPITPTL (110 µM), VPITPT (130 µM), FPGPIPN (260 µM), YPFPGPIPN (670 µM), PGPIHNS (1000 µM), IPPLTQTPV (1300 µM), PQNIPPL (1500 µM), and VPPFIQPE (2500 µM). Their activity expressed in IC50 values was provided in brackets [17].
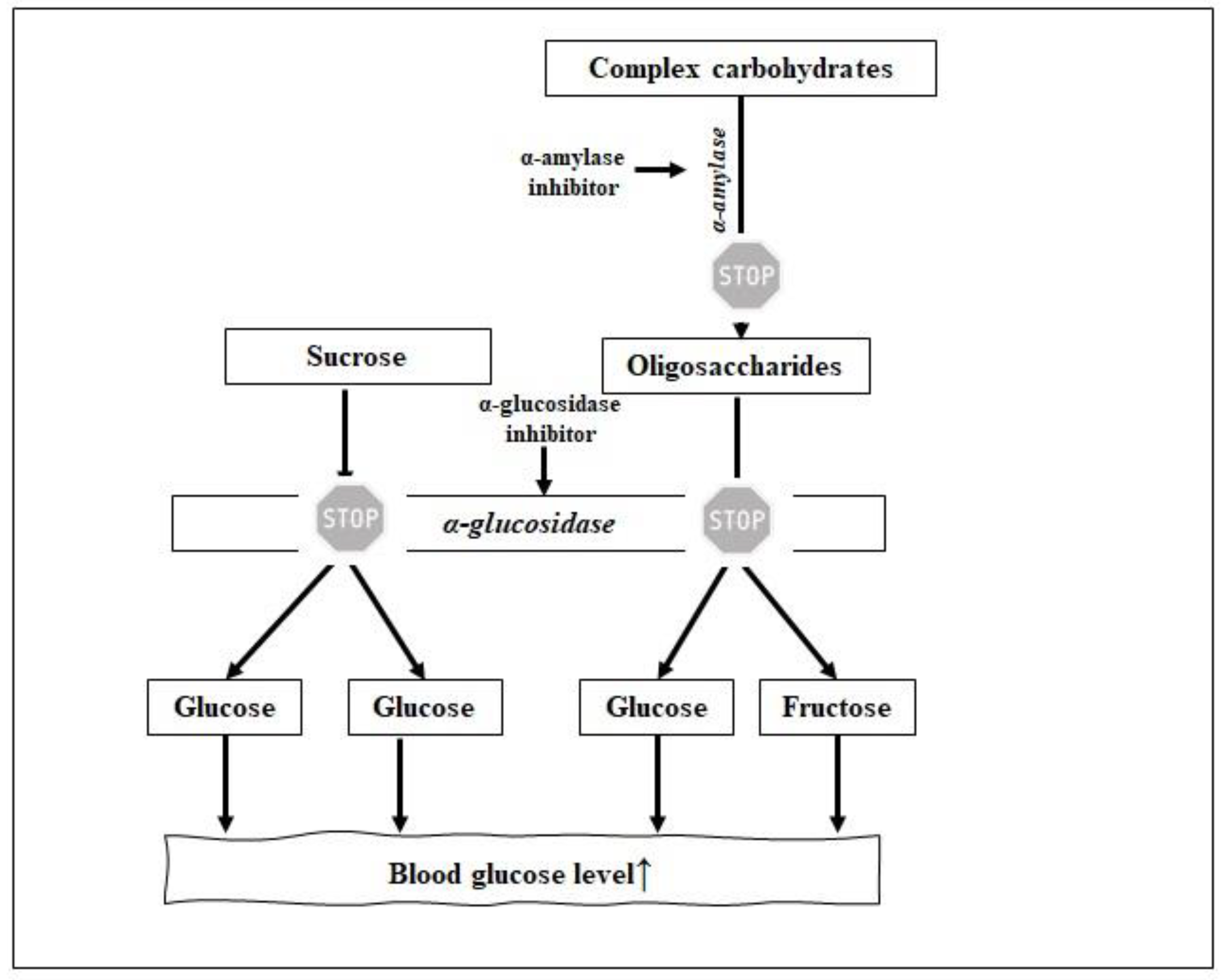
Antidiabetic properties of peptides may also result from their ability to inhibit α-glucosidase (EC 3.2.1.20). α-Glucosidase is an enzyme located in the brush border of enterocytes of the jejunum in the small intestine and is a key enzyme in carbohydrate breakdown. It cleaves carbohydrate to easy absorbable glucose. Thus, one of the T2D preventive strategies is to decrease the postprandial hyperglycemia by delaying glucose absorption through the inhibition of α-glucosidase (Figure 3). α-Glucosidase inhibitors also play a crucial role in the secretion of GLP-1 in healthy and diabetic patients, due to the prolonged incretin effect [18].
α-Amylase is another enzyme involved in the regulation of the blood sugar level [5]. This enzyme is responsible for the initial hydrolysis of the α-D-(1-4) glycosidic bonds of polysaccharides like glycogen and starch to oligosaccharides. Then, oligosaccharides are further hydrolyzed into monosaccharides by α-glucosidase (see above), which are next transferred into bloodstream [5].
Some milk proteins were reported as the sources of α-glucosidase inhibitors. Whey proteins: α-lactalbumin, β-lactoglobulin, serum albumin, and lactoferrin hydrolysates obtained by peptic digestion were confirmed to exert such an inhibiting effect. β-Lactoglobulin and whey protein isolate hydrolysates were found to inhibit α-glucosidase (IC50 value of 3.5 and 4.5 mg × mL−1, respectively) [18]. Dipeptide, like arginine-proline (RP), derived from whey proteins and being the component of commercial Pep2Dia® product, affected the inhibition of α-glucosidase with an IC50 value of 0.0025 mg × mL−1 [19]. To the best of our knowledge, there is no research papers about α-amylase inhibitors identified milk proteins and dairy products. Only one article concerned cheese as a source of α-glucosidase inhibitors. According to Grom et al. [20], Prato cheese inhibited α-glucosidase activity in vitro by 52.6%. Although, it was concluded that the consumption of probiotic Prato cheese can contribute to the reduction of postprandial glycemia in healthy individuals, but no peptide was identified [20].
It was evidenced on mice that peptides from cheese effectively decreased glucose level. Some epidemiological surveys showed a slightly higher risk of T2D development with increasing cheese consumption, but most of the meta-analyses concluded that increasing cheese consumption was correlated with a significant decreasing in T2D development risk [21]. Another epidemiological and cohort studies showed that dairy products can be beneficial in type 2 diabetes prevention. The latest research carried out on 34,224 men in the Health Professionals Follow-Up Study (1986–2012), 76,531 women in the Nurses’ Health Study (1986–2012), and 81,597 women in the Nurses’ Health Study II (1991–2013) suggested that the decrease of total dairy intake by > 1.0 serving per day over a period of 4 years was associated with an 11% increase of T2D development risk [22]. However, there were some opposite remarks to these above. For example, it was stated that increasing cheese consumption by > 0.5 serving per day, caused a 9% increase of T2D development risk, when compared to the maintained, stable cheese intake [22]. In turn, these findings were not consistent with other studies. For example, the meta-analysis made by Gao et al. [23], concerning seven studies with a total of 178,429 subjects and 14,810 cases, led to opposite conclusions. They found an inverse linear correlation between the consumption of cheese and the risk of T2D development. The consumption of 30 g per day cheese decreased the relative risk (RR) of T2D development (RR 0.80 ± 0.12). The obtained results suggested that a modest increase in cheese consumption might prevent from T2D but further research is needed [23]. Another eight cohort studies concerning the meta-analysis of dairy food consumption and its effect on the T2D risk revealed that the consumption of 50 g cheese per day decreased the risk of T2D development (RR 0.92 ± 0.07) and a non-linear negative correlation was observed between cheese consumption and T2D risk [24]. The cohort study conducted on 2809 participants for 12 years showed a negative dose-response relation between cheese consumption and risk of T2D development. Out of the 1867 participants with no T2D symptoms, 902 were diagnosed as prediabetic. Consumption of ≥ 4 servings of cheese per week lowered the risk of T2D incident by 70%. Less than one cheese serving per week was associated with a 63% reduction of T2D incidents. None of the abovementioned meta-analyses, however, considered what type of cheese caused the risk-reducing effect [25].
3. Peptides with Antihypertensive Activity
The regulation of blood pressure that takes place in the organism involves several modes of ACE action. They include renin-angiotensin, kinin-nitric-oxide, and neutral endopeptidase systems. The first one (RAS; renin-angiotensin system), also called renin-angiotensin-aldosterone system (RAAS), is considered as the major in blood pressure regulation. Briefly, renin (EC 3.4.22.15) activates angiotensinogen and releases angiotensin I. Then, ACE (Figure 4) transforms angiotensin I into angiotensin II (known as potent vasoconstrictor) and induces the release of aldosterone. Moreover, the increase of the blood pressure caused by angiotensin II causes some dysfunctions like damages of body organs leading to e.g., heart and kidney failure. Thus, the action of ACE receptor blockers or ACE-inhibiting peptides contributes to the blood pressure reduction [26].
Milk proteins have been reported as the reservoir of ACE inhibitors for many years [27]. The most known sequences are VPP and IPP identified in β- and κ-casein, respectively. Due to their antihypertensive potential tested in animals and humans, they became the components of commercial milk drink nutraceuticals such as Calpis, Evolus, and Valio [28]. VPP and IPP sequences were also identified in 36 cheeses of Swiss origin (Table 1). Depending on the type of Swiss cheese, the content of these two sequences varied. For VPP, it ranged from 0 to 224 mg per kg of cheese, whereas for IPP from 0 to 95.4 mg. The highest content of both peptides, VPP and IPP, was identified in the hard cheese “Hobelkäse from the Bernese Oberland” (224.1 and 95.4 mg × kg−1, respectively). That cheese sample had the highest ACE inhibitory activity of all analyzed chesses, with an IC50 value of 2.6 mg × mL−1 [29]. Different concentrations of these two peptides in cheeses were probably related to milk pretreatment, cultures, scalding conditions, and ripening time. Thus, it is recommended to develop a reproducible cheese-making process to produce higher concentrations of these peptides that could be used for clinical trials [30].
The potential of some cheeses containing the high concentrations of VPP and IPP to reduce blood pressure was tested in spontaneously hypertensive rats (SHR). Blood pressure was monitored after 6 h since oral administration. It was observed that the decrease of systolic blood pressure (SBP) differed depending on the cheese variety. The level of the SBP reduction (mmHg) in SHR was as follows: −24.7 and −17.2 (Gouda, aged 8 and 24 months, respectively), −17.2 (Gouda, aged 24 months), −20.7 (Edam), −20.3 (Blue), −20.0 (Havarti), and −13.0 (Emmental) [31].
According to the scientific reports, sequential motif located between 60 and 70 residue of β-casein is considered as strategic zone due to its ACE-inhibiting, immunostimulating, and opioid potential [32]. The example of peptide derived from β-casein showing dual bioactivity is β-casomorphin-7 (YPFPGPI; fragment 60–66) acting as opioid and ACE inhibitory agent [26]. De Noni and Cattaneo [33] identified this peptide in different dairy products including commercial cheeses like e.g., Brie, Cheddar, Gouda, Fontina, and Gorgonzola (see Table 2).
Some ACE inhibitors which matched the sequences of caseins were identified in Spanish cheeses. Those peptides which sequences fulfilled the structural requirements for being a potent ACE inhibitors were synthesized [34]. Their structural characteristics included the presence amino acids like lysine, arginine or residues possessing hydrophobic side chains as well as C-terminal proline [35]. Finally, it was reported that eight peptides (out of 11) inhibited ACE. Among them DKIHP peptide was found as the most potent (IC50 = 113.1 µM) but the addition C-terminal phenylalanine (DKIHPF) caused the dramatic reduction of its ACE-inhibiting potential (IC50 = 2419.4 µM) [36]. The ACE-inhibiting activity of synthesized peptide DKIHP (source Manchego cheese; β-casein) was also measured using different protecting groups, namely tert-butyloxycarbonyl (Boc) and fluorenyl-methyloxycarbonyl (Fmoc). The IC50 values for DKIHP were 113.18 and 577.92 μM, respectively. It was concluded that the differences above were attributed to the presence of less active conformers in the structure of the peptide [37]. The other sequences of ACE inhibitors identified in some Spanish cheeses are given in Table 3.
Sequences of caseins were also the sources of other ACE-inhibiting peptides that were identified in cheeses: RPKHPIKHQ (source: αs1-casein; Cheddar and Gouda; IC50 = n.d. and 13.4 µM, respectively), RPKHPIKHQGLPQ (source: αs1-casein; Gouda; IC50 = n.d.), YPFPGPIPN (source: β-casein; Gouda; IC50 = 14.8 µM), MPFPKYPVQPF (source: β-casein; Gouda; IC50 = n.d.), RPKHPIK, RPKHPI, FVAPFPEVFGK (source: αs1-casein, Cheddar; IC50 = n.d.), YQEPVLGPVRGPFPIIV (source: β-casein; Cheddar; IC50 = n.d.), RPKHPIKHQGLPQEV, RPKHPIKHQGLPQEVLNEN, LLR, EVLNENLLRF (source: αs1-casein, Fresco; IC50 = n.d.), FVAPFPEVFGK, YQEPVLGPVRGPF, YQEPVLGPVRGPFPI, YQEPVLGPVRGPFPIIV (source: β-casein; Fresco; IC50 = n.d.) [30]. Tonouchi et al. [38] isolated two peptides (LQP and MAP) from Danish skimmed milk cheese. LQP was originally known as ACE inhibitor identified in α-zein hydrolyzed with thermolysin. MAP sequence was identified in β-casein and exhibited ACE inhibition (IC50 = 0.8 µM) and antihypertensive effect in SHR. Significant hypotensive effect of MAP tripeptide was observed at 8 h since oral administration (−17.0 mm Hg; dose 3 mg × kg−1 body weight) [38].
The potential of cheeses as diet components that may affect the reduction of blood pressure is also in the focus of interest of the scientists aiming at meta-analyses [39]. Nilsen et al. [39] carried out a cross-sectional study to analyze the impact of the consumption of Gamalost cheese (a traditional Norwegian cheese) on the reduction of the risk of hypertension development. Based on data obtained from the questionnaires of 168 participants, the average intake of Gamalost servings per week was 2 and the average value of systolic and diastolic blood pressures was 128 and 78 mmHg, respectively. The increase of Gamalost servings reduced the systolic blood pressure by 0.72 mm Hg. Moreover, it was concluded that blood pressure reduction related to Gamalost consumption was associated with the presence of ACE inhibitory peptides [39]. Crippa et al. [40] investigated the antihypertensive effect resulting from the consumption of Grana Padano cheese and observed that supplementation of Grana Padano cheese at a dose 30 g per day significantly lowered the systolic and diastolic blood pressure in mild-hypertensive subjects (–3.5 and–2.4 mm Hg, respectively) [40].
One of the meta-analyses concerned studies evaluating the association of CVD, coronary heart disease (CHD), and stroke with high vs. low cheese consumption. This analysis revealed that high cheese consumption lowered the relative risk for: total CVD (RR 0.90; 7; 8076), CHD (RR 0.86; 8; 7631), and stroke (RR 0.90; 7; 10,449). Respective numbers in the brackets represent: relative risk, number of studies, and number of documented events provided. Finally, the results indicated a nonlinear relationship between cheese consumption and decreasing risks of CVD, CHD, and stroke. Consumption of approximately 40 g of cheese per day ensured the best results in reducing the risks of dysfunctions related to hypertension [41].
To conclude, epidemiological studies suggested that the antihypertensive potential of cheeses can be linked to the presence of biopeptides. According to above-mentioned analyses, the optimal daily cheese dose to reduce the risk of hypertensive dysfunctions was 30–40 g. These results suggest that cheese-as part of a diet-can prevent from the hypertension-related diseases.
4. Antioxidative Peptides
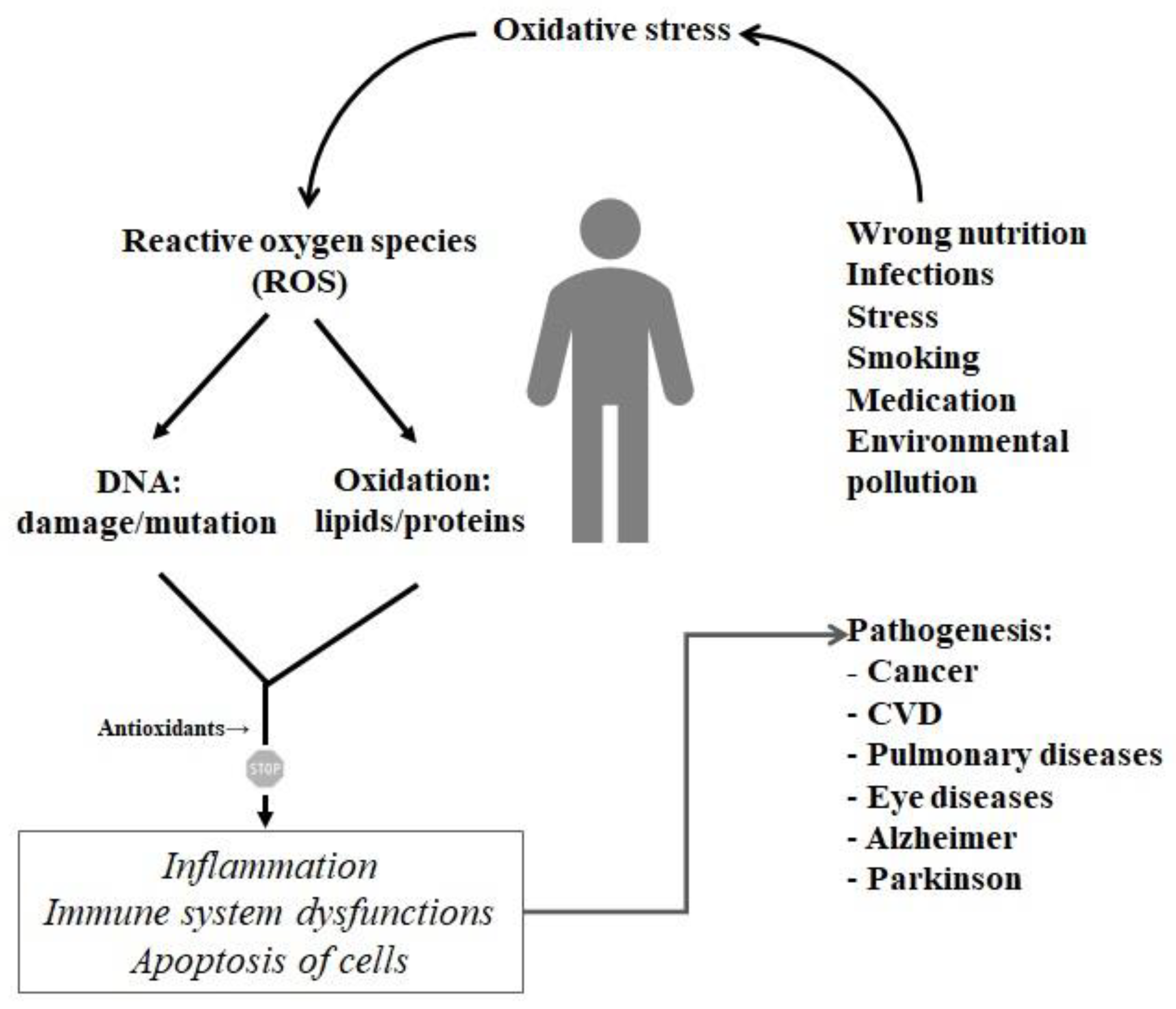
Free radicals are involved in many signaling processes, i.e., as defending the body from infections. Thus, the accumulation of free radicals in the organism can lead to unwanted cell damage. Oxidized cells can then initiate several body dysfunctions like e.g., rapid ageing, atherosclerosis, arthritis, diabetes, and cancer (Figure 5). It is the so-called oxidative stress defined as the process of formation of highly reactive molecules (that is, reactive oxygen species-ROS). In order to act against the destructive activity of ROS, it is important to look for diet-derived compounds acting as antioxidants [42].
Gupta et al. [43] produced Cheddar cheese using different Lactobacillus casei cultures and screened for antioxidant activity. Two antioxidative peptides matching the fragments of bovine β-casein (VKEAMAPK) and αS1-casein (HIQKEDVPSER) were identified in Cheddar. Both peptides exhibited 2,2-diphenyl-1-picryl-hydrazyl (DPPH) radical scavenging activity comparable to the commercial antioxidants such as butylated hydroxyanisole (BHA), tert-butyl hydroquinone (t-BHQ) or ferulic acid [44]. The results indicated that the antioxidant activity of the water-soluble cheese extracts (WSE) was dependent on the ripening period of cheese. The highest bioactivity was observed after 4 months of ripening. After that, the activity started to decrease, up until the 7th month of ripening and remained similar after 9 months [44]. Another research group evaluated the inhibition of DPPH radicals by the WSE of Australian Cheddar cheeses. They found a moderate antioxidant activity of the fractions containing peptides with a molecular weight > 10 kDa, although peptide identification was not carried out [45]. A study conducted to analyze the antioxidative potential of Mexican Cotija cheese revealed that the maximal antioxidant activity of this cheese was achieved after 24 weeks of ripening. Although no specific peptides were identified, it was concluded that the antioxidative potential of cheese was related to a significantly high content of bioactive sequences [46].
Timón et al. [47] produced hard cow cheese and analyzed the antioxidative potential of its WSE. After the fractionation of WSE, it was discovered that some of the extracts exhibited DPPH-radical scavenging and metal chelating bioactivity. Moreover, two peptides, namely EIVPN and DKIHPF which were known from their ACE-inhibiting function, were identified in WSE of the hard cheese. Another sequence i.e., VAPFPQ identified in cheese-derived extract was discovered as a new metal chelating peptide. To conclude, it was postulated that antioxidative potential of cheese water-soluble extracts may be the result of the presence of the above-mentioned peptides. However, the bioactivity of these sequences should be confirmed on synthetic peptides [47].
Some peptides with the antioxidative potential were identified in WSE of the Burgos-type cheese. It was shown that 22 fractions derived from Burgos-type cheese extracts exhibited radical scavenging activity (oxygen radical absorbance capacity with fluorescein (ORAC-FL) assay). Liquid chromatography and tandem mass spectrometry (LC-MS/MS) identification revealed the presence of the aromatic amino acids like tyrosine, phenylalanine or tryptophan in some fractions of cheeses extracts. These amino acids itself possess the antioxidative potential [48]. It was also reported that some of the Burgos-type cheese-originating peptide fractions contained the sequences, namely: SDIPNPIGSENSEKTTMPLW (source: αs1-casein), YQQPVLGPVRGPFPIIV (source: β-casein), and LLYQQPVLGPVRGPFPIIV (source: β-casein) [49]. According to Timón et al. [49], peptides matching the sequences of β-casein have already been known as antioxidative agents. The sequence SDIPNPIGSENSEKTTMPLW is probably found as the new peptidic antioxidant, however further tests to measure this activity need to be performed [49].
5. Hypocholesterolemic Peptides
Plasma lipoproteins are composed of the following elements: cholesterol, triglycerides, phospholipids, and apoproteins. Their varying composition affects the density, size, and electrophoretic mobility of each lipoprotein particle which enables for the classification of lipoprotein disorders [50]. According to Cox et al. [50], cholesterol is an essential precursor of some substances as adrenal and gonadal steroid hormones and bile acids. Moreover, it is an element of the animal cell membranes. Triglycerides are the major component of dietary fat as well as animal fat depots. Abnormal concentrations of cholesterol and triglycerides in blood indicate some dysfunctions in synthesis, degradation, and transport of lipoproteins and their increase levels lead to the different variants of hypercholesterolemia and hypertriglyceridemia [50]. In turn, phospholipids are the constituents of the natural membranes. They are polar lipids which are e.g., the components of the milk fat globule membrane involved in the circulation of the lipid droplets secreted by the mammary gland cells [51].
It was reported that some peptides/protein-derived hydrolysates exhibited activity against metabolic lipids’ dysfunctions and their effect was tested in cellular systems and animal models [52]. To the best of our knowledge, no peptides acting as hypolipidemic were identified in cheeses. When thinking about milk, most of the studies concerning their “anticholesterolemic effect” conducted so far, have concerned the whey proteins. The ingestion of whey protein was observed to be correlated with a significant reduction in total cholesterol levels in rats fed cholesterol-free and cholesterol-enriched diets [53]. The cholesterol reducing activity of β-lactoglobulin was due to the presence of two major biopeptides, namely lactostatin (IIAEK) and β-lactotensin (HIRL) [54]. According to the literature, consumption of 300 mg IIAEK × kg−1 body weight per day reduced LDL level as well as increased HDL cholesterol level, and fecal steroid excretion. The lactostatin activity was linked to the degradation of cholesterol into bile acids in hepatic cells [55,56]. The HIRL sequence was shown to be involved in the reduction of total (22.7%), and LDL plus VLDL (32.0%) cholesterol levels in mice after 2 days of administration at the dose of 30 mg × kg−1 body weight [57]. The activity of both peptides was tested on mice fed a cholesterol-enriched diet. Despite the optimistic results, it should be noted that the exact mechanisms of the hypocholesterolemic activity of peptides are still unclear. It is speculated that it might be linked to the amino acid composition, but further research is needed in this respect [54].
6. Antiobesity and Anti-Inflammatory Peptides
The studies concerning the effectiveness of different types of diets and proportions of macronutrients on the weight loss is in the focus of the interest of the many scientists [58]. Some studies have shown that the high-protein content diet is the most effective when thinking about the reduction of body weight. It results from the fact that consumption of the protein contributes to the increased satiety and the reduction of the energy intake in addition to a greater dietary thermogenesis [59].
Although proteins accelerate caloric metabolism, some of them are more efficient on weight loss than others. It was observed that soybean-originating proteins more efficiently enhance weight loss than bovine milk proteins [60]. One of the explanations of this phenomenon results from the fact that peptides produced from proteins during gastrointestinal digestion initiate satiety signals from the gut and hence suppress food intake. Thus, scientists try to explain reliably the mechanisms of a protein-rich diet on the body’s health [5]. Some of them include e.g., the impact of dietary protein on cholecystokinin (CKK) release, the activation of GLP-1 receptor to suppress appetite, and/or modulating the adipose genes affecting metabolic and vascular functions [60].
It was reported that opioid peptides may also contribute to the weight loss due to the reduction of food intake [61]. For example, casomorphins interact with gastric opioid receptors and slow down the motility of the gastrointestinal tract, which inhibits food intake [60]. The example of the opioid peptide identified both in bovine casein and several types of cheeses is β-casomorphin-7 (YPFPGPI) which was described above (see Section 2). It was indicated that dose of 179 mg × kg−1 body weight of modified β-casomorphins effectively impaired gastric emptying in pup rats. It was probably related to direct interaction of β-casomorphins with opioid receptors in the gut [62].
Some studies confirmed the causal direction of the relationship between adiposity and inflammation (the greater adiposity, the higher C-reactive protein level) [63]. Citing the words by Zielińska et al. [64], inflammation is defined as a “complex biological process, occurring through a variety of mechanisms and leading to changes in local blood flow and the release of several molecular mediators”. Uncontrolled inflammation may lead to chronic dysfunctions such as rheumatoid arthritis, atherosclerosis, and it is linked to cancer [65]. It was reported that VPP and IPP milk-derived ACE-inhibiting tripeptides exhibited anti-inflammatory activity. First reduced leukocyte-endothelial cell interaction in vitro and this effect was related to that of inhibition of MAPK (mitogen-activated protein kinase) signaling pathway. The anti-inflammatory effect of both peptides was also tested in a model of intestinal enterocolitis. It was found that VPP and IPP were protective against atherosclerosis in mice with defect of endogenous apoprotein E (ApoE). These peptides suppressed the mRNA for inflammatory cytokines, oxidized low density lipoprotein receptor, and transcription regulators [65]. These two peptides were also identified in the different types of cheeses (see Section 2).
To the best of our knowledge, there is no more specific anti-inflammatory peptides that could be identified in cheeses. However, it was found that water-soluble extracts of Cheddar cheese possessed such an activity which was determined based on nitric oxide (NO) production in lipopolysaccharide-stimulated macrophage (RAW-264.7) cells [66].
7. Multi-Activity of Peptides in MetS Prevention: A Screening Using BIOPEP-UWM Database
Loads of publication data concern the analysis of the proteins as the sources of biopeptides exhibiting only one biological effect while the information about dual or multifunctional properties of peptides is rather poor [5]. Such statement may also refer to the MetS-preventive peptides. To the best of our knowledge, there is no a “universal” peptide sequence that would show all bioactivities described in this paper. Thus, databases of biological information like BIOPEP-UWM, available at: http://www.uwm.edu.pl/biochemia, may be a suitable option to acquire the knowledge about peptides exhibiting more than one biological function [67]. BIOPEP-UWM is a curated database providing comprehensive information on 3899 bioactive peptide sequences (accessed: March 2020).
We applied BIOPEP-UWM database to retrieve the profiles of potential biological activities of the four major casein fractions (see Table A1). Profile of the potential biological activity of the protein is defined as the type and the location of bioactive fragment in a protein chain [67]. Briefly, such analysis enables to answer the question: what bioactive fragments match the protein sequence? Then, each peptide sequence was pasted again to BIOPEP-UWM database of bioactive peptide sequences using the search option “sequence” and ticking “exact” [67]. Such searching was helpful to show what additional activities are exhibited by this peptide (the pasted one). This approach led us find that 66 peptides had dual i.e., ACE and DPP4 inhibitory function, and 17 were acting as ACE inhibitory and antioxidant agents. Some peptides exerted a triple bioactivity, i.e., eight dipeptides inhibited ACE and DPP4 as well as exerted an antioxidant activity. The example of the sequence with triple inhibitory bioactivity is YP peptide present in αS1-, β-, and κ-casein sequence. It has proven ACE, α-glucosidase and DPP4 inhibitory function. The brief summary of peptides encrypted in the sequences of caseins and exerting e.g., more than one biological effect is shown in Table 4. For checking what particular sequences were found as dual or multiactive, see Table A1.
Although, our bioinformatic approach is useful for searching multifunctions of peptides, it has some limitations. The important one is based on the idea of the positive selection assuming that sequence of interest matches the peptide sequences present in databases. Thus, it may not include peptides that are “new” and they are, at the time of screening, not displayed in the database. Thus, some authors postulate for the regular update of the databases with newly-reported bioactive peptides [68]. Nevertheless, such databases, if curated and reliable, may contribute to acquiring of some information about the peptides which multi-activity can be described as “MetS-preventive”.
8. Conclusions
Some milk-derived peptides were identified in different types of cheeses. These biopeptides exhibited bioactivities playing an important role in the prevention from the dysfunctions related to metabolic syndrome. It needs to be noted that majority of studies concerned the identification of peptides in cheeses and did not included studies on animal and humans. Several epidemiological and cohort studies confirmed the positive effect of cheese consumption on the reduction of the risk of MetS diseases. A majority of studies concerning milk-derived peptides and their identification in cheeses focus on the determination of only one biological effect. Despite its limitations, bioinformatic-assisted approach can be helpful in the screening of multifunctional effect of peptides being the promising agents in the prevention of MetS.
Author Contributions
D.M. wrote Section 3, Section 4, Section 5, Section 6 and Section 7, and was involved in draft preparation and editing; A.I. was responsible for conceptualization, wrote Section 1, Section 2, Section 8, and reviewed the article. All authors have read and agreed to the published version of the manuscript.
Funding
Project financially supported by Minister of Science and Higher Education in the range of the program entitled “Regional Initiative of Excellence” for the years 2019–2022, Project No. 010/RID / 2018 / 19, amount of funding 12.000.000 PLN as well as the funds of the University of Warmia and Mazury in Olsztyn (Project No. 17.610.014-110).
Acknowledgments
The authors wish to thank Małgorzata Darewicz for her valuable scientific suggestions when preparing this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| ACE | Angiotensin-converting enzyme (EC 3.4.15.1) |
| ApoE | ndogenous apoprotein E |
| BHA | butylated hydroxyanisole |
| BIOPEP-UWM | database of bioactive peptide sequences [68] |
| Boc | tert-butyloxycarbonyl |
| CHD | coronary heart disease |
| CVD | cardiovascular disease |
| DPP4 | dipeptidyl peptidase IV (EC 3.4.14.5) |
| DPPH | 2,2-diphenyl-1-picryl-hydrazyl) |
| Fmoc | fluorenylmethyloxycarbonyl |
| GIP | glucose-dependent insulinotropic polypeptide |
| GLP-1 | glucagon-like peptide 1 |
| IP3 | inositol 1,4,5-triphosphate |
| LC-MS/MS | Liquid Chromatography and Tandem Mass Spectrometry |
| LDL cholesterol | low density lipoprotein cholesterol |
| MAPK | mitogen-activated protein kinase |
| MetS | metabolic syndrome |
| NHANES | Health and Nutrition Examination Survey |
| NO | nitric oxide |
| NTR2 | neurotensin receptor 2 |
| ORAC-FL | Oxygen Radical Absorbance Capacity with Fluorescein |
| RAAS | renin-angiotensin-aldosterone system |
| RAS | renin-angiotensin system |
| ROS | reactive oxygen species |
| RR | relative risk |
| SBP | systolic blood pressure |
| SHR | spontaneously hypertensive rats |
| T2D | type 2 diabetes mellitus |
| t-BHQ | tert-butyl hydroquinone |
| VLDL cholesterol | very low density lipoprotein cholesterol |
| WSE | water-soluble cheese extracts |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table A1.
Biopeptides (MetS-preventive sequences) Present in the Sequences of Bovine Caseins - Data Acquired from the BIOPEP-UWM Database (accessed on March 2020).
Table A1.
Biopeptides (MetS-preventive sequences) Present in the Sequences of Bovine Caseins - Data Acquired from the BIOPEP-UWM Database (accessed on March 2020).
| BIOPEP-UWM ID | Activity | Sequence | Protein | Location |
|---|---|---|---|---|
| 7590/8637 | 1* | AA | κ-casein | (86–87) |
| 8758 | 12 | AE | αS1-casein | (77–78), (131–132) |
| αS2-casein | (77–78) | |||
| 7583/8759 | 1 | AF | β-casein | (189–190) |
| 8193 | 9 | AI | αS2-casein | (42–43) |
| κ-casein | (128–129) | |||
| 8559 | 12 | AL | αS1-casein | (13–14) |
| αS2-casein | (13–14), (96–97), (190–191) | |||
| κ-casein | (13–14), (70–71) | |||
| 3177/7584 | 1 | AP | αS1-casein | (41–42), (191–192) |
| β-casein | (103–104) | |||
| 7742 | 9 | AR | αS1-casein | (15–16) |
| κ-casein | (117–118) | |||
| 8762 | 12 | AS | κ-casein | (147–148) |
| 8763 | 12 | AT | αS2-casein | (80–81) |
| κ-casein | (165–166) | |||
| 8764/8951 | 1 | AV | αS1-casein | (11–12) |
| αS2-casein | (11–12), (131–132) | |||
| β-casein | (177–178) | |||
| κ-casein | (87–88), (159–160), (189–190) | |||
| 7543/8460/8695 | 7 | AW | αS1-casein | (178–179) |
| 3563/7866/8765 | 7 | AY | αS1-casein | (158–159), (173–174) |
| 7606 | 9 | DA | αS1-casein | (172–173), (190–191) |
| 9075 | 9 | DM | αS2-casein | (155–156) |
| β-casein | (184–185) | |||
| 8768 | 12 | DQ | αS1-casein | (66–67) |
| αS2-casein | (125–126) | |||
| 7623 | 9 | EA | αS1-casein | (76-77) |
| β-casein | (100–101) | |||
| κ-casein | (158–159) | |||
| 7622/8770 | 1 | EG | αS1-casein | (140–141) |
| 8771 | 12 | EH | αS2-casein | (20–21) |
| 7826/8772 | 1 | EI | αS1-casein | (85–86), (125–126) |
| αS2-casein | (99–100) | |||
| β-casein | (11–12) | |||
| κ-casein | (139–140), (179–180) | |||
| 7840/8558 | 1 | EK | αS1-casein | (50–51), (207–208) |
| αS2-casein | (38–39), (172–173) | |||
| β-casein | (31–32) | |||
| κ-casein | (33–34) | |||
| 7888 | 11 | EL | αS1-casein | (54–55), (156–157), (163–164) |
| β-casein | (2–3), (5–6), (44–45) | |||
| 8529 | 12 | EP | αS1-casein | (148–149) |
| β-casein | (117–118), (195–196) | |||
| κ-casein | (150–151) | |||
| 8773 | 12 | ES | αS1-casein | (62–63), (78–79) |
| αS2-casein | (27–28), (75–76), (157–158) | |||
| β-casein | (14–15), (21–22) | |||
| κ-casein | (161–162), (175–176) | |||
| 8774 | 12 | ET | αS2-casein | (33–34) |
| 7828/8775 | 1 | EV | αS1-casein | (29–30), (45–46) |
| αS2-casein | (57–58), (78–79), (83–84), (160–161) | |||
| β-casein | (91–92) | |||
| κ-casein | (172–173) | |||
| 7752/8777 | 1 | EY | αS2-casein | (66–67) |
| 3176 | 12 | FA | αS2-casein | (189–190) |
| β-casein | (52–53) | |||
| 9342 | 11 | FC | αS2-casein | (54–55) |
| 7605 | 9 | FG | αS1-casein | (47–48) |
| 8555 | 12 | FL | αS2-casein | (178–179) |
| β-casein | (87–88), (190–191) | |||
| κ-casein | (6–7), (18–19), (76–77) | |||
| 3502/8506 | 1 | FP | αS1-casein | (43–44) |
| αS2-casein | (107–108) | |||
| β-casein | (62–63), (111–112), (157–158), (205–206) | |||
| 8779/9076 | 1 | FQ | β-casein | (33–34) |
| 7592/8780 | 1 | FR | αS1-casein | (165–166) |
| 3556 | 9 | FY | αS1-casein | (160–161), (168–169) |
| αS2-casein | (103–104) | |||
| 7598/8524 | 1 | GA | αS1-casein | (177–178) |
| κ-casein | (20–21) | |||
| 7615/8781 | 1 | GE | β-casein | (10–11) |
| κ-casein | (149–150) | |||
| 7596/8785 | 1 | GI | αS1-casein | (141–142) |
| 7611 | 9 | GK | αS1-casein | (48–49) |
| 7599/8561 | 1 | GL | αS1-casein | (25–26) |
| κ-casein | (60–61) | |||
| 3169/7512 | 1 | GP | αS2-casein | (117–118) |
| β-casein | (64–65), (199–200), (203–204) | |||
| 7607 | 9 | GS | αS1-casein | (60–61), (202–203) |
| αS2-casein | (70–71) | |||
| 7612 | 9 | GT | αS1-casein | (185–186) |
| 7608/8786 | 1 | GV | αS1-casein | (152–153) |
| β-casein | (94–95) | |||
| 3532/8788 | 1 | GY | αS1-casein | (108–109) |
| 3184 | 12 | HA | αS1-casein | (143–144) |
| 8793 | 12 | HI | αS1-casein | (95–96) |
| 7844 | 9 | HK | β-casein | (106–107) |
| 3317/7602/8557 | 7 | HL | β-casein | (134–135) |
| κ-casein | (123–124) | |||
| 7842/8520 | 1 | HP | αS1-casein | (19–20) |
| κ-casein | (119–120), (121–122) | |||
| β-casein | (50–51) | |||
| 8795 | 12 | HS | αS1-casein | (136–137) |
| 8797 | 12 | HV | αS2-casein | (21–22) |
| 3494/8799 | 1 | HY | αS2-casein | (92–93) |
| 7562/8525 | 1 | IA | κ-casein | (43–44), (146–147) |
| 7827 | 9 | IE | β-casein | (30–31) |
| κ-casein | (174–175) | |||
| 7593 | 9 | IF | αS2-casein | (5–6) |
| 7595 | 9 | IG | αS1-casein | (59-60), (151-152), (201-202) |
| αS2-casein | (69–70) | |||
| 8800 | 12 | IH | αS1-casein | (142–143) |
| β-casein | (49–50) | |||
| 8801 | 12 | II | αS2-casein | (29–30) |
| β-casein | (207–208) | |||
| 8802/9079 | 1 | IL | αS1-casein | (5–6) |
| κ-casein | (11–12), (94–95) | |||
| 8804 | 12 | IN | αS2-casein | (43–44), (100–101) |
| β-casein | (26–27) | |||
| κ-casein | (72–73), (143–144), (180–181) | |||
| 7581/8501 | 1 | IP | αS1-casein | (197–198) |
| αS2-casein | (216–217) | |||
| β-casein | (66-67), (74-75) | |||
| κ-casein | (47–48), (129–130), (140–141) | |||
| 8805 | 12 | IQ | αS1-casein | (96–97) |
| αS2-casein | (209–210) | |||
| β-casein | (187–188) | |||
| κ-casein | (49–50) | |||
| 3258/8215/8806 | 7 | IR | κ-casein | (30–31) |
| 3174/7743 | 1 | KA | αS2-casein | (95–96), (203–204) |
| β-casein | (176–177) | |||
| 8134 | 11 | KD | αS1-casein | (57–58) |
| κ-casein | (34–35) | |||
| 7841/8808 | 1 | KE | αS1-casein | (49–50), (98–99), (139–140), (147–148) |
| αS2-casein | (47–48), (56–57) | |||
| β-casein | (99–100), (107–108) | |||
| 7692/8809 | 1 | KF | αS2-casein | (2–3), (106–107), (188–189) |
| β-casein | (32–33) | |||
| 8811 | 12 | KH | αS1-casein | (18–19), (22–23), (94–95) |
| αS2-casein | (91–92) | |||
| β-casein | (105–106) | |||
| 8812 | 12 | KI | αS2-casein | (85–86), (181–182) |
| β-casein | (29–30), (48–49) | |||
| κ-casein | (42–43) | |||
| 8813 | 12 | KK | αS1-casein | (117–118) |
| αS2-casein | (151–152), (164–165), (180–81) | |||
| β-casein | (28–29) | |||
| κ-casein | (132–133) | |||
| 7693 | 9 | KL | αS1-casein | (2–3) |
| αS2-casein | (167–168) | |||
| 7810/8218/8519 | 7 | KP | αS2-casein | (206–207) |
| κ-casein | (67–68), (84–85) | |||
| 7586/8814 | 1 | KR | αS2-casein | (128–129) |
| 8815 | 12 | KS | κ-casein | (3–4), (107–108) |
| 8816 | 12 | KT | αS1-casein | (208–209) |
| αS2-casein | (152–153), (165–166), (196–197), (212–213) | |||
| κ-casein | (137–138) | |||
| 8817 | 12 | KV | αS1-casein | (51–52), (120–121) |
| αS2-casein | (214–215) | |||
| β-casein | (97–98), (169–170) | |||
| 7691/8819 | 1 | KY | αS1-casein | (118–119) |
| β-casein | (113–114) | |||
| κ-casein | (45–46) | |||
| 3175/7585 | 1 | LA | αS1-casein | (14–15), (157–158) |
| αS2-casein | (10–11), (14–15) | |||
| κ-casein | (12–13) | |||
| 3551 | 9 | LF | αS1-casein | (164–165) |
| 7619 | 9 | LG | αS1-casein | (107–108), (184–185) |
| β-casein | (198–199) | |||
| κ-casein | (19–20) | |||
| 3305/8820 | 6 | LH | αS1-casein | (135–136) |
| β-casein | (133–134) | |||
| 8821 | 12 | LI | αS1-casein | (4–5) |
| κ-casein | (71–72) | |||
| 8217 | 11 | LK | αS1-casein | (116–117) |
| αS2-casein | (179–180), (195–196) | |||
| 3182 | 12 | LL | αS1-casein | (3–4), (35–36), (113–114) |
| αS2-casein | (9–10) | |||
| β-casein | (139–140), (191–192) | |||
| 7832/8823 | 1 | LN | αS1-casein | (31–32) |
| αS2-casein | (97–98), (121–122), (138–139), (176–177) | |||
| β-casein | (6–7) | |||
| κ-casein | (61–62) | |||
| 3180 | 12 | LP | αS1-casein | (26–27) |
| αS2-casein | (191–192) | |||
| β-casein | (70–71), (135–136), (137–138), (151–152), (171–172) | |||
| κ-casein | (16–17), (77–78) | |||
| 7831 | 9 | LQ | αS2-casein | (111–112) |
| β-casein | (45–46), (88–89), (140–141) | |||
| κ-casein | (95–96) | |||
| 9213 | 9 | LR | αS1-casein | (36–37), (114–115) |
| 8824 | 12 | LT | αS1-casein | (6–7) |
| αS2-casein | (168–169) | |||
| β-casein | (77–78), (125–126), (127–128) | |||
| κ-casein | (14–15) | |||
| 8825 | 12 | LV | αS1-casein | (9–10) |
| β-casein | (58–59) | |||
| κ-casein | (7–8) | |||
| 3389/8462/8688 | 7 | LW | αS1-casein | (213–214) |
| 3381/7872 | 2 | LY | αS2-casein | (114–115) |
| β-casein | (192–193) | |||
| 3173 | 12 | MA | αS2-casein | (41–42) |
| β-casein | (102–103) | |||
| κ-casein | (116–117), (127–128) | |||
| 7839/8826 | 1 | ME | αS1-casein | (69–70), (75–76) |
| αS2-casein | (19–20), (156–157) | |||
| 3385/8827 | 1 | MF | β-casein | (156–157) |
| 7609/8828 | 1 | MG | β-casein | (93–94) |
| 8829 | 12 | MH | β-casein | (144–145) |
| 8830 | 12 | MI | αS1-casein | (150–151) |
| 8831 | 12 | MK | αS1-casein | (1–2), (138–139) |
| αS2-casein | (1–2), (205–206) | |||
| κ-casein | (2–3) | |||
| 8833/9085/9086 | 7 | MM | κ-casein | (1–2) |
| 3171 | 12 | MP | αS1-casein | (211–212) |
| β-casein | (109–110), (185–186) | |||
| 8839 | 12 | NA | αS2-casein | (61–62), (130–131) |
| 8841 | 12 | NE | αS1-casein | (32–33), (53–54) |
| αS2-casein | (63–64), (98–99) | |||
| 7683/8842 | 1 | NF | αS2-casein | (177–178) |
| 7698 | 9 | NK | β-casein | (27–28) |
| 8845 | 12 | NL | αS1-casein | (34–35) |
| αS2-casein | (49–50) | |||
| β-casein | (132–133) | |||
| 8846 | 12 | NM | αS2-casein | (40–41) |
| 8847 | 12 | NN | κ-casein | (73–74) |
| 8530 | 12 | NP | αS1-casein | (199–200) |
| αS2-casein | (44–45), (122–123) | |||
| 8848 | 12 | NQ | αS1-casein | (154–155) |
| αS2-casein | (101–102) | |||
| κ-casein | (25–26), (74–75), (134–135) | |||
| 8849 | 12 | NR | αS2-casein | (139–140), (174–175) |
| 8850 | 12 | NT | αS2-casein | (17–18) |
| κ-casein | (102–103), (144–145), (181–182) | |||
| 8851 | 12 | NV | β-casein | (7–8) |
| 7682/8853 | 1 | NY | κ-casein | (62–63) |
| 3179 | 12 | PA | κ-casein | (85–86), (91–92), (105–106) |
| 8854 | 12 | PF | αS1-casein | (42–43) |
| β-casein | (51–52), (61–62), (86–87), (110–111), (118–119), (204–205) | |||
| κ-casein | (17–18) | |||
| 7625/8855 | 1 | PG | β-casein | (9–10), (63–64) |
| 7843/8856 | 1 | PH | β-casein | (147–148) |
| κ-casein | (120–121), (122–123) | |||
| 8857 | 12 | PI | αS1-casein | (20–21), (200–201) |
| αS2-casein | (118–119), (133–134) | |||
| β-casein | (65–66), (186–187), (206–207) | |||
| κ-casein | (29–30), (48–49) | |||
| 8858 | 12 | PK | αS1-casein | (17–18) |
| αS2-casein | (211–212) | |||
| β-casein | (104–105), (112–113) | |||
| κ-casein | (131–132) | |||
| 7513/8638 | 1 | PL | αS1-casein | (183–184), (212–213) |
| β-casein | (76–77), (136–137), (138–139), (150–151) | |||
| 8859 | 12 | PM | αS1-casein | (149–150) |
| 8860 | 12 | PN | αS1-casein | (88–89), (128–129), (198–199) |
| β-casein | (67–68) | |||
| 3170/7836 | 1 | PP | β-casein | (75–76), (85–86), (152–153), (158–159) |
| κ-casein | (130–131), (177–178) | |||
| 7837/8861 | 1 | PQ | αS1-casein | (27–28), (122–123) |
| αS2-casein | (108–109), (192–193) | |||
| β-casein | (71–72), (159–160), (174–175), (181–182) | |||
| 8862 | 12 | PS | αS1-casein | (102–103), (175–176), (192–193) |
| αS2-casein | (45–46) | |||
| κ-casein | (57–58) | |||
| 7833/8863 | 1 | PT | αS2-casein | (136–137) |
| β-casein | (153–154) | |||
| κ-casein | (113–114), (141–142), (151–152), (155–156) | |||
| 8864 | 12 | PV | β-casein | (81–82), (115–116), (172–173), (196–197), (200–201) |
| κ-casein | (68–69) | |||
| 8190/8865 | 6 | PW | αS2-casein | (123–124), (207–208) |
| 8866 | 12 | PY | αS2-casein | (217–218) |
| β-casein | (179–180) | |||
| κ-casein | (78–79), (80–81) | |||
| 8867 | 12 | QA | αS1-casein | (67–68) |
| β-casein | (188–189) | |||
| κ-casein | (110–111) | |||
| 8868 | 12 | QD | β-casein | (46–47) |
| κ-casein | (135–136) | |||
| 8869 | 12 | QE | αS1-casein | (28–29), (155–156) |
| αS2-casein | (32–33), (37–38) | |||
| β-casein | (194–195) | |||
| κ-casein | (22–23), (26–27) | |||
| 8870 | 12 | QF | αS1-casein | (167–168) |
| αS2-casein | (102–103) | |||
| κ-casein | (75–76) | |||
| 7617/8871 | 1 | QG | αS1-casein | (24–25) |
| αS2-casein | (116–117) | |||
| 8872 | 12 | QH | αS2-casein | (200–201) |
| 8873 | 12 | QI | κ-casein | (93–94) |
| 7680 | 9 | QK | αS1-casein | (93–94), (97–98), (146–147) |
| αS2-casein | (94–95), (105–106), (187–188), (202–203) | |||
| β-casein | (175–176) | |||
| κ-casein | (66–67) | |||
| 8874 | 12 | QL | αS1-casein | (112–113), (123–124), (170–171) |
| αS2-casein | (142–143) | |||
| 8875 | 12 | QN | β-casein | (72–73) |
| κ-casein | (24–25) | |||
| 8532 | 12 | QP | αS2-casein | (210–211) |
| β-casein | (89–90), (146–147), (149–150) | |||
| κ-casein | (28–29), (112–113) | |||
| 8876 | 12 | αS1-casein | (145–146) | |
| β-casein | (38–39), (39–40) | |||
| κ-casein | (65–66) | |||
| 8877 | 12 | QS | β-casein | (34–35), (56–57), (123–124), (141–142), (160–161), (167–168) |
| 8878 | 12 | QT | β-casein | (40–41), (54–55), (79–80) |
| 8879 | 12 | QV | αS2-casein | (126–127) |
| κ-casein | (98–99), (184–185) | |||
| 8880 | 12 | QW | κ-casein | (96–97) |
| 8881 | 12 | QY | αS1-casein | (187–188) |
| αS2-casein | (109–110), (112–113), (193–194) | |||
| κ-casein | (50–51) | |||
| 3489 | 9 | RF | αS1-casein | (37–38) |
| κ-casein | (37–38) | |||
| 8882/9173 | 1 | RG | β-casein | (202–203) |
| 8883 | 12 | RH | κ-casein | (118–119) |
| 8884 | 12 | RI | β-casein | (25–26) |
| 3257/8886 | 1 | RL | αS1-casein | (115–116), (134–135) |
| αS2-casein | (175–176) | |||
| 8888 | 12 | RN | αS2-casein | (60–61), (129–130) |
| 7582/8518 | 1 | RP | αS1-casein | (16–17) |
| 3380 | 9 | RY | αS1-casein | (105–106) |
| αS2-casein | (185–186), (220–221) | |||
| κ-casein | (55–56) | |||
| 7685/8891 | 1 | SF | αS1-casein | (193–194) |
| κ-casein | (4–5), (125–126) | |||
| 7618 | 9 | SG | αS1-casein | (176–177) |
| κ-casein | (148–149) | |||
| 8893 | 12 | SI | αS1-casein | (79–80) |
| αS2-casein | (28–29), (68–69) | |||
| β-casein | (22–23) | |||
| 8894 | 12 | SK | αS1-casein | (56–57) |
| αS2-casein | (46–47), (150–151) | |||
| β-casein | (96–97), (168–169) | |||
| 8560 | 12 | SL | β-casein | (15–16), (57–58), (69–70), (124–125), (164–165) |
| 8505 | 12 | SP | κ-casein | (90–91), (170–171), (176–177) |
| 9184 | 9 | ST | αS1-casein | (63–64) |
| αS2-casein | (52–53), (144–145), (158–159) | |||
| κ-casein | (153–154), (162–163), (187–188) | |||
| 8895 | 12 | SV | αS1-casein | (90–91) |
| β-casein | (161–162) | |||
| 8896 | 12 | SW | β-casein | (142–143) |
| 7684/8897 | 1 | SY | κ-casein | (58–59) |
| 8531 | 12 | TA | κ-casein | (188–189) |
| 8898 | 12 | TD | αS1-casein | (189–190) |
| β-casein | (128–129) | |||
| 7830/8899 | 1 | TE | αS1-casein | (64–65) |
| αS2-casein | (81–82), (159–160), (169–170) | |||
| β-casein | (41–42), (120–121) | |||
| κ-casein | (138–-139), (157–158) | |||
| 8185/8900 | 1 | TF | αS2-casein | (53–54) |
| 8903 | 12 | TI | κ-casein | (10–11), (142–143), (145–146) |
| 8904 | 12 | TK | αS2-casein | (163–164), (166–167), (213–214) |
| 8905 | 12 | TL | αS2-casein | (137–138) |
| β-casein | (126–127) | |||
| κ-casein | (15–16), (166–167) | |||
| 8906 | 12 | TM | αS1-casein | (210–211) |
| αS2-casein | (18–19) | |||
| κ-casein | (115–116) | |||
| 8503/9073 | 1 | TP | αS2-casein | (135–136) |
| β-casein | (80–81) | |||
| κ-casein | (154–155) | |||
| 7834/8908 | 1 | TQ | αS1-casein | (186–187) |
| β-casein | (55–56), (78–79) | |||
| 8909 | 12 | TR | β-casein | (24–25) |
| 8910 | 12 | TS | αS2-casein | (145–146) |
| κ-casein | (152–153), (186–187) | |||
| 8911 | 12 | TT | αS1-casein | (209–210) |
| κ-casein | (114–115), (156–157) | |||
| 8912 | 12 | TV | αS2-casein | (87–88), (153–154), (197–198) |
| β-casein | (154–155) | |||
| κ-casein | (103–104), (163–164), (182–183) | |||
| 8219/8914 | 6 | TY | αS2-casein | (34–35) |
| 3172 | 12 | VA | αS1-casein | (10–11), (12–13), (40–41) |
| αS2-casein | (12–13), (79–80) | |||
| κ-casein | (69–70), (164–165) | |||
| 8915 | 12 | VD | αS2-casein | (88–89), (154–155) |
| 7829/8916 | 1 | VE | αS1-casein | (91–92) |
| β-casein | (13–14), (116–117), (130–131) | |||
| κ-casein | (160–161) | |||
| 3384/8917 | 1 | VF | αS1-casein | (46–47) |
| αS2-casein | (161–162) | |||
| 8920 | 12 | VI | αS2-casein | (215–216) |
| κ-casein | (173–174) | |||
| 7558/8921 | 1 | VK | αS2-casein | (84–85), (127–128) |
| β-casein | (98–99) | |||
| 8922 | 12 | VL | αS1-casein | (30–31) |
| αS2-casein | (120–121) | |||
| β-casein | (162–163), (170–171), (197–198) | |||
| κ-casein | (52–53), (99–100) | |||
| 8923 | 12 | VM | β-casein | (92–93), (155–156) |
| 8924 | 12 | VN | αS1-casein | (52–53), (153–154) |
| 3181/7587 | 1 | VP | αS1-casein | (87–88), (101–102), (121–122), (127–128), (182–183) |
| αS2-casein | (132–133) | |||
| β-casein | (8–9), (84–85), (173–174), (178–179) | |||
| κ-casein | (104–105) | |||
| 8925 | 12 | VQ | κ-casein | (183–184) |
| 7628/8594 | 1 | VR | αS2-casein | (59–60), (219–220) |
| β-casein | (201–202) | |||
| κ-casein | (88–89) | |||
| 8926 | 12 | VS | αS2-casein | (22–23) |
| β-casein | (95–96) | |||
| 8927 | 12 | VT | κ-casein | (9–10), (185–186) |
| 3183 | 12 | VV | αS2-casein | (58–59) |
| β-casein | (82–83), (83–84) | |||
| κ-casein | (8–9) | |||
| 3492/8224/8929 | 7 | VY | αS2-casein | (198–199) |
| β-casein | (59–60) | |||
| 8930 | 12 | WD | αS2-casein | (124–125) |
| 8679 | 12 | WI | αS2-casein | (208–209) |
| 8682/9090 | 1 | WM | β-casein | (143–144) |
| 8678 | 12 | WQ | κ-casein | (97–98) |
| 7898/8683 | 6 | WY | αS1-casein | (179–180) |
| 7589/8932 | 1 | YA | κ-casein | (82–83) |
| 8935 | 12 | YF | αS1-casein | (159–160) |
| 3553/8936 | 1 | YG | κ-casein | (59–60) |
| 8938 | 12 | YI | κ-casein | (46–47) |
| 7697/8939 | 1 | YK | αS1-casein | (119–120) |
| αS2-casein | (35–36) | |||
| 8940 | 12 | YL | αS1-casein | (106–107), (109–110) |
| αS2-casein | (110–111), (113–114), (194–195), (221–222) | |||
| 3666/8521/9548 | 8 | YP | αS1-casein | (161–162), (174–175) |
| β-casein | (60–61), (114–115), (180–181) | |||
| κ-casein | (56–57), (79–80) | |||
| 8943 | 12 | YQ | αS1-casein | (169–170) |
| αS2-casein | (93–94), (104–105), (115–116), (186–187), (199–200) | |||
| β-casein | (193–194) | |||
| κ-casein | (64–65) | |||
| 8945 | 12 | YS | αS2-casein | (67–68) |
| 8696 | 12 | YT | αS1-casein | (188–189) |
| 8946/9077 | 1 | YV | αS1-casein | (181–182) |
| αS2-casein | (218–219) | |||
| κ-casein | (51–52) | |||
| 8948 | 12 | YY | αS1-casein | (180–181) |
| κ-casein | (63–64), (81–82) | |||
| 9042 | 9 | AFL | β-casein | (189–191) |
| 3597 | 9 | AIP | κ-casein | (128–130) |
| 9029 | 9 | ALP | αS2-casein | (190–192) |
| 3370 | 9 | AVP | αS2-casein | (131–133) |
| β-casein | (177–179) | |||
| 9359 | 11 | CLV | αS1-casein | (8–10) |
| 7823 | 9 | FAL | αS2-casein | (189–191) |
| 7560 | 9 | FFL | κ-casein | (5–7) |
| 3377 | 9 | FGK | αS1-casein | (47–49) |
| 7554 | 9 | GEP | κ-casein | (149–151) |
| 7509 | 9 | GLP | αS1-casein | (25–27) |
| 7545/9116 | 1 | GPV | β-casein | (199–201) |
| 3311 | 11 | HPH | κ-casein | (119–121), (121–123) |
| 9027 | 9 | HQG | αS1-casein | (23–25) |
| 7626 | 9 | IAK | κ-casein | (43–45) |
| 3167 | 12 | IPI | κ-casein | (47–49) |
| 3522 | 9 | IPP | β-casein | (74–76) |
| κ-casein | (129–131) | |||
| 7803 | 9 | IPY | αS2-casein | (216–218) |
| 8184/8693 | 1 | IQP | αS2-casein | (209–211) |
| 8149/8180 | 2 | IQY | κ-casein | (49–51) |
| 9365 | 11 | KKY | αS1-casein | (117–119) |
| 8133 | 11 | KVI | αS2-casein | (214–216) |
| 3558 | 9 | LAY | αS1-casein | (157–159) |
| 9031 | 9 | LEE | β-casein | (3–5) |
| 9206 | 9 | LFR | αS1-casein | (164–166) |
| 7508 | 9 | LGP | β-casein | (198–200) |
| 7995 | 11 | LHL | β-casein | (133–135) |
| 8000 | 11 | LHS | αS1-casein | (135–137) |
| 8484 | 11 | LLR | αS1-casein | (35–37), (113–115) |
| 8509 | 9 | LNF | αS2-casein | (176–178) |
| 3544 | 9 | LNP | αS2-casein | (121–123) |
| 9026 | 9 | LNY | κ-casein | (61–63) |
| 9040 | 9 | LPF | κ-casein | (16–18) |
| 8616 | 12 | LPL | β-casein | (135–137), (137–139) |
| 3391 | 9 | LPP | β-casein | (151–153) |
| 9339 | 12 | LPQ | αS1-casein | (26–28) |
| αS2-casein | (191–193) | |||
| β-casein | (70–72) | |||
| 3542/8689 | 1 | LQP | β-casein | (88–90) |
| 9163 | 11 | LTC | αS1-casein | (6–8) |
| 8402 | 9 | LVY | β-casein | (58–60) |
| 9241 | 9 | MAP | β-casein | (102–104) |
| 8958 | 9 | MKP | αS2-casein | (205–207) |
| 9363 | 11 | NEN | αS1-casein | (32–34) |
| 7965 | 11 | NYY | κ-casein | (62–64) |
| 8139 | 11 | PEL | αS1-casein | (162–164) |
| 8029 | 11 | PHL | κ-casein | (122–124) |
| 8032 | 11 | PHQ | β-casein | (147–149) |
| 3094/7510 | 5 | PLG | αS1-casein | (183–185) |
| 2664 | 9 | PLP | β-casein | (136–138), (150–152) |
| 3531 | 9 | PLW | αS1-casein | (212–214) |
| 7645 | 9 | PPK | κ-casein | (130–132) |
| 8652 | 12 | PPL | β-casein | (75–77) |
| 3373 | 9 | PQR | β-casein | (181–183) |
| 7559 | 9 | PSY | κ-casein | (57–59) |
| 8040 | 11 | PWD | αS2-casein | (123–125) |
| 8044 | 11 | PWI | αS2-casein | (207–209) |
| 3372 | 9 | PYP | β-casein | (179–181) |
| κ-casein | (78–80) | |||
| 3307 | 11 | PYY | κ-casein | (80–82) |
| 9146 | 9 | QGP | αS2-casein | (116–118) |
| 9543 | 9 | RYL | αS1-casein | (105–107) |
| αS2-casein | (220–222) | |||
| 8382/8474 | 2 | RYQ | αS2-casein | (185–187) |
| 7498 | 9 | TVY | αS2-casein | (197–199) |
| 3521 | 9 | VAP | αS1-casein | (40–42) |
| 7635 | 9 | VAV | αS1-casein | (10–12) |
| 2653 | 9 | VLP | β-casein | (170–172) |
| 8347 | 12 | VPL | αS1-casein | (182–184) |
| 3524 | 9 | VPP | β-casein | (84–86) |
| 9046 | 9 | VQV | κ-casein | (183–185) |
| 8384 | 9 | VRY | αS2-casein | (219–221) |
| 3505 | 9 | VYP | β-casein | (59–61) |
| 7962 | 11 | WYY | αS1-casein | (179–181) |
| 7963 | 11 | YFY | αS1-casein | (159–161) |
| 3973 | 9 | YGL | κ-casein | (59–61) |
| 7959 | 11 | YLY | αS2-casein | (113–115) |
| 2870 | 15 | YPF | β-casein | (60–62) |
| 8617/9550 | 3 | YPY | κ-casein | (79–81) |
| 8487 | 11 | YQL | αS1-casein | (169–171) |
| 8150 | 11 | YVL | κ-casein | (51–53) |
| 7634 | 9 | YVP | αS1-casein | (181–183) |
| 7939 | 11 | YYA | κ-casein | (81–83) |
| 7948 | 11 | YYQ | κ-casein | (63–65) |
| 7942 | 11 | YYV | αS1-casein | (180–182) |
| 8369 | 9 | AFLL | β-casein | (189–192) |
| 7668 | 9 | AIPP | κ-casein | (128–131) |
| 8380/8472 | 2 | AYPS | αS1-casein | (173–176) |
| 8591 | 12 | FLQP | β-casein | (87–90) |
| 7499 | 9 | FVAP | αS1-casein | (39–42) |
| 8489 | 11 | FYQL | αS1-casein | (168–171) |
| 8371/8467 | 2 | GTQY | αS1-casein | (185–188) |
| 3306 | 11 | HPHL | κ-casein | (121–124) |
| 7661 | 9 | LFRQ | αS1-casein | (164–167) |
| 2668 | 9 | LHLP | β-casein | (133–136) |
| 2665 | 9 | LPLP | β-casein | (135–138) |
| 3397 | 9 | LQSW | β-casein | (140–143) |
| 9236 | 9 | LRFF | αS1-casein | (36–39) |
| 3331 | 9 | LVYP | β-casein | (58–61) |
| 3374 | 9 | PFPE | αS1-casein | (42–45) |
| 7896 | 11 | PYPQ | β-casein | (179–182) |
| 8357 | 9 | RYLG | αS1-casein | (105–108) |
| 8378/8470 | 2 | RYPS | κ-casein | (55–58) |
| 8221 | 11 | TSTA | κ-casein | (186–189) |
| 8483 | 11 | TVYQ | αS2-casein | (197–200) |
| 8593 | 12 | VLGP | β-casein | (197–200) |
| 8309 | 9 | VRSP | κ-casein | (88–91) |
| 7801 | 9 | VRYL | αS2-casein | (219–222) |
| 8308 | 9 | VVPP | β-casein | (83–86) |
| 8592 | 12 | WIQP | αS2-casein | (208–211) |
| 8381/8473 | 2 | YAKP | κ-casein | (82–85) |
| 8485 | 11 | YGLN | κ-casein | (59–62) |
| 9420/9421 | 2 | YLGY | αS1-casein | (106–109) |
| 8482 | 11 | YLKT | αS2-casein | (194–197) |
| 8140 | 11 | YPEL | αS1-casein | (161–164) |
| 2868 | 15 | YPFP | β-casein | (60–63) |
| 3218/8615 | 4 | YPYY | κ-casein | (79–82) |
| 7878 | 11 | YQEP | β-casein | (193–196) |
| 8477 | 11 | YQLD | αS1-casein | (169–172) |
| 3399/7660 | 9 | AMKPW | αS2-casein | (204–208) |
| 9229 | 9 | ARHPH | κ-casein | (117–121) |
| 3371 | 9 | AVPYP | β-casein | (177–181) |
| 3519 | 9 | AYFYP | αS1-casein | (158–162) |
| 3497 | 9 | FFVAP | αS1-casein | (38–42) |
| 8368 | 9 | FPIIV | β-casein | (205–209) |
| 9378 | 9 | FPPQS | β-casein | (157–161) |
| 8141 | 11 | FYPEL | αS1-casein | (160–164) |
| 9375 | 9 | GLPQE | αS1-casein | (25–29) |
| 8478 | 11 | GYLEQ | αS1-casein | (108–112) |
| 2666 | 9 | HLPLP | β-casein | (134–138) |
| 8613 | 12 | IPIQY | κ-casein | (47–51) |
| 7627 | 9 | KDERF | κ-casein | (34–38) |
| 9552 | 9 | KFPQY | αS2-casein | (106–110) |
| 8618 | 12 | LPLPL | β-casein | (135–139) |
| 7631/8614 | 1 | LPYPY | κ-casein | (77–81) |
| 2671 | 9 | NLHLP | β-casein | (132–136) |
| 8157 | 9 | PGPIP | β-casein | (63–67) |
| 9242 | 9 | PLPLL | β-casein | (136–140) |
| 9243 | 9 | PPEIN | κ-casein | (177–181) |
| 8373/8469 | 2 | RHPHP | κ-casein | (118–122) |
| 9244 | 9 | RINKK | β-casein | (25–29) |
| 8481 | 11 | RLKKY | αS1-casein | (115–119) |
| 8358 | 9 | RYLGY | αS1-casein | (105–109) |
| 8375 | 9 | RYQKF | αS2-casein | (185–189) |
| 9246 | 9 | SLPQN | β-casein | (69–73) |
| 3332 | 9 | SLVYP | β-casein | (57–61) |
| 8379/8471 | 2 | SRYPS | κ-casein | (54–58) |
| 7485 | 9 | TKVIP | αS2-casein | (213–217) |
| 8278/9556 | 6 | VPYPQ | β-casein | (178–182) |
| 8370/8466 | 2 | YAKPA | κ-casein | (82–86) |
| 8480 | 11 | YPELF | αS1-casein | (161–165) |
| 3263 | 13 | YPFPG | β-casein | (60–64) |
| 8486 | 11 | YQKFP | αS2-casein | (104–108) |
| 8372/8468 | 2 | ARHPHP | κ-casein | (117–122) |
| 3501 | 9 | AYFYPE | αS1-casein | (158–163) |
| 7493 | 9 | DKIHPF | β-casein | (47–52) |
| 7629 | 9 | EKDERF | κ-casein | (33–38) |
| 3511 | 9 | EMPFPK | β-casein | (108–113) |
| 7480 | 9 | FALPQY | αS2-casein | (189–194) |
| 7487 | 9 | KVLPVP | β-casein | (169–174) |
| 3513 | 9 | LAYFYP | αS1-casein | (157–162) |
| 2667 | 9 | LHLPLP | β-casein | (133–138) |
| 8179 | 9 | LKKISQ | αS2-casein | (179–184) |
| 9238 | 9 | LVYPFP | β-casein | (58–63) |
| 8152/8178 | 2 | PYVRYL | αS2-casein | (217–222) |
| 3426 | 9 | QSLVYP | β-casein | (56–61) |
| 8479 | 11 | RDMPIQ | β-casein | (183–188) |
| 3128 | 14 | RYLGYL | αS1-casein | (105–110) |
| 3217 | 14 | SRYPSY | κ-casein | (54–59) |
| 3127/3530 | 5 | TTMPLW | αS1-casein | (209–214) |
| 9248 | 9 | VLSRYP | κ-casein | (52–57) |
| 8658 | 12 | VPITPT | αS2-casein | (132–137) |
| 7481 | 9 | VTSTAV | κ-casein | (185–190) |
| 9251 | 9 | VVVPPF | β-casein | (82–87) |
| 3504 | 9 | VYPFPG | β-casein | (59–64) |
| 7887/8376 | 2 | YFYPEL | αS1-casein | (159–164) |
| 7630 | 9 | YIPIQY | κ-casein | (46–51) |
| 3500 | 9 | YKVPQL | αS1-casein | (119–124) |
| 3265 | 15 | YLGYLE | αS1-casein | (106–111) |
| 9252 | 12 | YPVEPF | β-casein | (114–119) |
| 7662/9253 | 9 | YQEPVL | β-casein | (193–198) |
| 8383 | 9 | YYQQKP | κ-casein | (63–68) |
| 3480/7875 | 2 | AVPYPQR | β-casein | (177–183) |
| 8359 | 9 | AYFYPEL | αS1-casein | (158–164) |
| 7496 | 9 | FPEVFGK | αS1-casein | (43–49) |
| 8659 | 12 | FPGPIPN | β-casein | (62–68) |
| 7659 | 9 | FPQYLQY | αS2-casein | (107–113) |
| 8181 | 9 | FSDKIAK | κ-casein | (39–45) |
| 8159 | 9 | GPFPIIV | β-casein | (203–209) |
| 8285 | 11 | IPIQYVL | κ-casein | (47–53) |
| 9373 | 9 | KEDVPSE | αS1-casein | (98–104) |
| 9234 | 9 | KKYKVPQ | αS1-casein | (117–123) |
| 3498 | 9 | KVLPVPQ | β-casein | (169–175) |
| 7565 | 9 | LHLPLPL | β-casein | (133–139) |
| 8657 | 12 | LPQNIPP | β-casein | (70–76) |
| 8155 | 9 | LTLTDVE | β-casein | (125–131) |
| 2669 | 9 | NLHLPLP | β-casein | (132–138) |
| 9058 | 12 | PQNIPPL | β-casein | (71–77) |
| 3473 | 13 | RYLGYLE | αS1-casein | (105–111) |
| 8476 | 11 | TIASGEP | κ-casein | (145–151) |
| 3333 | 9 | TQSLVYP | β-casein | (55–61) |
| 7877 | 11 | VLPVPQK | β-casein | (170–176) |
| 8475 | 11 | VLSRYPS | κ-casein | (52–58) |
| 9249 | 12 | VPITPTL | αS2-casein | (132–138) |
| 9250 | 9 | VPSERYL | αS1-casein | (101–107) |
| 3262/7665 | 5 | YPFPGPI | β-casein | (60–66) |
| 3214 | 15 | YPSYGLN | κ-casein | (56–62) |
| 9254 | 9 | YQKFPQY | αS2-casein | (104–110) |
| 8377 | 9 | AYFYPELF | αS1-casein | (158–165) |
| 3512 | 9 | DAYPSGAW | αS1-casein | (172–179) |
| 9539 | 9 | EKVNELSK | αS1-casein | (50–57) |
| 2672 | 9 | ENLHLPLP | β-casein | (131–138) |
| 7658/8151 | 2 | FALPQYLK | αS2-casein | (189–196) |
| 7876 | 11 | KVLPVPQK | β-casein | (169–176) |
| 9235 | 12 | LPQNIPPL | β-casein | (70–77) |
| 3398 | 9 | MKPWIQPK | αS2-casein | (205–212) |
| 7802 | 9 | NMAINPSK | αS2-casein | (40–47) |
| 9168/9273 | 9 | PFPEVFGK | αS1-casein | (42–49) |
| 3334 | 9 | QTQSLVYP | β-casein | (54–61) |
| 8160 | 9 | RGPFPIIV | β-casein | (202–209) |
| 7796 | 11 | VKEAMAPK | β-casein | (98–105) |
| 7492 | 9 | VYPFPGPI | β-casein | (59–66) |
| 2869/7501 | 5 | YPFPGPIP | β-casein | (60–67) |
| 7879 | 11 | YQEPVLGP | β-casein | (193–200) |
| 7804 | 9 | ALNEINQFY | αS2-casein | (96–104) |
| 3395/7497 | 9 | AMKPWIQPK | αS2-casein | (204–212) |
| 3335 | 9 | AQTQSLVYP | β-casein | (53–61) |
| 8660 | 12 | IPPLTQTPV | β-casein | (74–82) |
| 9380 | 9 | LGPVRGPFP | β-casein | (198–206) |
| 7494 | 9 | LNVPGEIVE | β-casein | (6–14) |
| 8396 | 9 | QSWMHQPHQ | β-casein | (141–149) |
| 7483 | 9 | RPKHPIKHQ | αS1-casein | (16–24) |
| 9376 | 9 | VEQKHIQKE | αS1-casein | (91–99) |
| 7566 | 9 | VRGPFPIIV | β-casein | (201–209) |
| 7486 | 9 | YPFPGPIPN | β-casein | (60–68) |
| 8156 | 9 | YPQRDMPIQ | β-casein | (180–188) |
| 8397 | 9 | YYAKPAAVR | κ-casein | (81–89) |
| 3336 | 9 | FAQTQSLVYP | β-casein | (52–61) |
| 8169/8286 | 2 | GPVRGPFPII | β-casein | (199–208) |
| 9231 | 12 | INNQFLPYPY | κ-casein | (72–81) |
| 7495 | 9 | NIPPLTQTPV | β-casein | (73–82) |
| 3216/3495 | 5 | YIPIQYVLSR | κ-casein | (46–55) |
| 7805 | 9 | ALNEINQFYQK | αS2-casein | (96–106) |
| 7797 | 11 | ARHPHPHLSFM | κ-casein | (117–127) |
| 9237 | 9 | LTQTPVVVPPF | β-casein | (77–87) |
| 3503 | 9 | TPVVVPPFLQP | β-casein | (80–90) |
| 3251 | 13 | YPFPGPIPNSL | β-casein | (60–70) |
| 9377 | 9 | EIVPNSAEERLH | αS1-casein | (125–136) |
| 7800 | 9 | FFVAPFPEVFGK | αS1-casein | (38–49) |
| 3337 | 9 | HPFAQTQSLVYP | β-casein | (50–61) |
| 3338 | 9 | IHPFAQTQSLVYP | β-casein | (49–61) |
| 9379 | 9 | QEPVLGPVRGPFP | β-casein | (194–206) |
| 3339 | 9 | KIHPFAQTQSLVYP | β-casein | (48–61) |
| 9240 | 9 | LVYPFPGPIPNSLPQN | β-casein | (58–73) |
| 7564 | 9 | LVYPFPGPIPNSLPQNIPP | β-casein | (58–76) |
*1-ACE and DPP4 inhibitor; 2-ACE inhibitory and antioxidative activity; 3-DPP4 and α-glucosidase inhibitor; 4-DPP4 inhibitory and opioid activity; 5- ACE inhibitory and opioid activity; 6- DPP4 inhibitory and antioxidative activity; 7- ACE, DPP4 inhibitory, and antioxidative activity; 8- ACE, DPP4, and α-glucosidase inhibitor; 9-ACE inhibitor; 10- α-glucosidase inhibitor; 11- antioxidative activity; 12- DPP4 inhibitor; 13-opioid agonist activity; 14 - opioid antagonist activity; 15-opioid activity.
References
- Karadag, A.S.; Ozlu, E.; Lavery, M.J. Cutaneous manifestations of diabetes mellitus and the metabolic syndrome. Clin. Dermatol. 2018, 36, 89–93. [Google Scholar] [CrossRef]
- Rochlani, Y.; Pothineni, N.V.; Kovelamudi, S.; Mehta, J.L. Metabolic syndrome: Pathophysiology, management, and modulation by natural compounds. Ther. Adv. Cardiovasc. Dis. 2017, 11, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.X.; Chaudhary, N.; Akinyemiju, T. Metabolic syndrome prevalence by race/ethnicity and sex in the united states, national health and nutrition examination survey, 1988–2012. Prev. Chronic Dis. 2017, 14, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Huang, P.L. A comprehensive definition for metabolic syndrome. Dis. Model. Mech. 2009, 2, 231–237. [Google Scholar] [CrossRef] [Green Version]
- Iwaniak, A.; Darewicz, M.; Minkiewicz, P. Peptides Derived from Foods as Supportive Diet Components in the Prevention of Metabolic Syndrome. Compr. Rev. Food Sci. Food Saf. 2018, 17, 63–81. [Google Scholar] [CrossRef] [Green Version]
- González-Ortega, O.; López-Limón, A.R.; Morales-Domínguez, J.F.; Soria-Guerra, R.E. Production and purification of recombinant hypocholesterolemic peptides. Biotechnol. Lett. 2015, 37, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.P.; Vij, S.; Hati, S. Functional significance of bioactive peptides derived from soybean. Peptides 2014, 54, 171–179. [Google Scholar] [CrossRef]
- Yuan, L.; Shu, W.; Shen, Y.; Jin, W.; Gao, R. Inflammation and food-derived anti-inflammatory peptides. Biomed. Res. Sci. 2019, 1, 1001. [Google Scholar]
- Xu, Q.; Yan, X.; Zhang, Y.; Wu, J. Current understanding of transport and bioavailability of bioactive peptides derived from dairy proteins: A review. Int. J. Food Sci. Technol. 2019, 54, 1930–1941. [Google Scholar] [CrossRef] [Green Version]
- Dehghan, M.; Mente, A.; Rangarajan, S.; Sheridan, P.; Mohan, V.; Iqbal, R.; Gupta, R.; Lear, S.; Wentzel-Viljoen, E.; Avezum, A.; et al. Association of dairy intake with cardiovascular disease and mortality in 21 countries from five continents (PURE): A prospective cohort study. Lancet 2018, 392, 2288–2297. [Google Scholar] [CrossRef]
- Abedini, M.; Falahi, E.; Roosta, S. Dairy product consumption and the metabolic syndrome. Diabetes Metab. Syndr. Clin. Res. Rev. 2015, 9, 34–37. [Google Scholar] [CrossRef] [PubMed]
- López-Expósito, I.; Amigo, L.; Recio, I. A mini-review on health and nutritional aspects of cheese with a focus on bioactive peptides. Dairy Sci. Technol. 2012, 92, 419–438. [Google Scholar] [CrossRef] [Green Version]
- Darewicz, M.; Dziuba, B.; Minkiewicz, P.; Dziuba, J. The Preventive Potential of Milk and Colostrum Proteins and Protein Fragments. Food Rev. Int. 2011, 27, 357–388. [Google Scholar] [CrossRef]
- Skwarek, A.; Darewicz, M.; Borawska-Dziadkiewicz, J. Ripened cheese as a source of bioactive peptides. Biotechnol. Food Sci. 2018, 82, 49–60. [Google Scholar]
- McIntosh, C.H.S.; Widenmaier, S.; Kim, S. Chapter 15 Glucose-Dependent Insulinotropic Polypeptide (Gastric Inhibitory Polypeptide; GIP). Vitam. Horm. 2009, 80, 409–471. [Google Scholar] [CrossRef]
- Mkele, G. Dipeptidyl peptidase-4 inhibitors: Their role in the management of type 2 diabetes. S. Afr. Fam. Pr. 2013, 55, 508–510. [Google Scholar] [CrossRef]
- Uenishi, H.; Kabuki, T.; Seto, Y.; Serizawa, A.; Nakajima, H. Isolation and identification of casein-derived dipeptidyl-peptidase 4 (DPP-4)-inhibitory peptide LPQNIPPL from gouda-type cheese and its effect on plasma glucose in rats. Int. Dairy J. 2012, 22, 24–30. [Google Scholar] [CrossRef]
- Patil, P.; Mandal, S.; Tomar, S.K.; Anand, S. Food protein-derived bioactive peptides in management of type 2 diabetes. Eur. J. Nutr. 2015, 54, 863–880. [Google Scholar] [CrossRef]
- Sartorius, T.; Weidner, A.; Dharsono, T.; Boulier, A.; Wilhelm, M.; Schön, C. Postprandial Effects of a Proprietary Milk Protein Hydrolysate Containing Bioactive Peptides in Prediabetic Subjects. Nutrients 2019, 11, 1700. [Google Scholar] [CrossRef] [Green Version]
- Grom, L.C.; Rocha, R.S.; Balthazar, C.F.; Guimarães, J.T.; Coutinho, N.M.; Barros, C.P.; Pimentel, T.C.; Venâncio, E.L.; Collopy Junior, I.; Maciel, P.M.C.; et al. Postprandial glycemia in healthy subjects: Which probiotic dairy food is more adequate? J. Dairy Sci. 2020, 103, 1110–1119. [Google Scholar] [CrossRef]
- Hutchison, A.T.; Piscitelli, D.; Horowitz, M.; Jones, K.L.; Clifton, P.M.; Standfield, S.; Hausken, T.; Feinle-Bisset, C.; Luscombe-Marsh, N.D. Acute load-dependent effects of oral whey protein on gastric emptying, gut hormone release, glycemia, appetite, and energy intake in healthy men. Am. J. Clin. Nutr. 2015, 102, 1574–1584. [Google Scholar] [CrossRef] [PubMed]
- Drouin-Chartier, J.-P.; Li, Y.; Ardisson Korat, A.V.; Ding, M.; Lamarche, B.; Manson, J.E.; Rimm, E.B.; Willett, W.C.; Hu, F.B. Changes in dairy product consumption and risk of type 2 diabetes: Results from 3 large prospective cohorts of US men and women. Am. J. Clin. Nutr. 2019, 110, 1201–1212. [Google Scholar] [CrossRef] [PubMed]
- Gao, D.; Ning, N.; Wang, C.; Wang, Y.; Li, Q.; Meng, Z.; Liu, Y.; Li, Q. Dairy Products Consumption and Risk of Type 2 Diabetes: Systematic Review and Dose-Response Meta-Analysis. PLoS ONE 2013, 8, e73965. [Google Scholar] [CrossRef]
- Aune, D.; Norat, T.; Romundstad, P.; Vatten, L.J. Dairy products and the risk of type 2 diabetes: A systematic review and dose-response meta-analysis of cohort studies. Am. J. Clin. Nutr. 2013, 98, 1066–1083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hruby, A.; Ma, J.; Rogers, G.; Meigs, J.B.; Jacques, P.F. Associations of Dairy Intake with Incident Prediabetes or Diabetes in Middle-Aged Adults Vary by Both Dairy Type and Glycemic Status. J. Nutr. 2017, 147, 1764–1775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwaniak, A.; Minkiewicz, P.; Darewicz, M. Food-Originating ACE Inhibitors, Including Antihypertensive Peptides, as Preventive Food Components in Blood Pressure Reduction. Compr. Rev. Food Sci. Food Saf. 2014, 13, 114–134. [Google Scholar] [CrossRef]
- Hrynkiewicz, M.; Iwaniak, A.; Bucholska, J.; Minkiewicz, P.; Darewicz, M. Structure-activity prediction of ACE inhibitory/bitter dipeptides - a chemometric approach based on stepwise regression. Molecules 2019, 24, 950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beltrán-Barrientos, L.M.; Hernández-Mendoza, A.; Torres-Llanez, M.J.; González-Córdova, A.F.; Vallejo-Córdoba, B. Inivited review: Fermented milk as antihypertensive functional food. J. Dairy Sci. 2016, 99, 499–4110. [Google Scholar] [CrossRef] [Green Version]
- Bütikofer, U.; Meyer, J.; Sieber, R.; Wechsler, D. Quantification of the angiotensin-converting enzyme-inhibiting tripeptides Val-Pro-Pro and Ile-Pro-Pro in hard, semi-hard and soft cheeses. Int. Dairy J. 2007, 17, 968–975. [Google Scholar] [CrossRef]
- Pihlanto, A. Lactic fermentation and bioactive peptides. In Lactic Acid Bacteria. R & D for Food, Health and Livestock Purposes; Kongo, J.M., Ed.; IntechOpen: London, UK, 2013; pp. 309–332. [Google Scholar] [CrossRef]
- Sieber, R.; Bütikofer, U.; Egger, C.; Portmann, R.; Walther, B.; Wechsler, D. ACE inhibitory activity and ACE-inhibiting peptides. Dairy Sci. Technol. 2010, 90, 47–73. [Google Scholar] [CrossRef]
- Haque, E.; Chand, R. Antihypertensive and Antimicrobial Bioactive Peptides from Milk Proteins. Eur. Food Res. Technol. 2008, 227, 7–15. [Google Scholar] [CrossRef]
- De Noni, I.; Cattaneo, S. Occurrence of β-casomorphins 5 and 7 in commercial dairy products and in their digests following in vitro simulated gastro-intestinal digestion. Food Chem. 2010, 119, 560–566. [Google Scholar] [CrossRef]
- Gómez-Ruiz, J.Á.; Ramos, M.; Recio, I. Angiotensin-converting enzyme-inhibitory peptides in Manchego cheeses manufactured with different starter cultures. Int. Dairy J. 2002, 12, 697–706. [Google Scholar] [CrossRef]
- Pripp, A.H.; Isaksson, T.; Stepaniak, L.; Sørhaug, T. Quantitative structure-activity relationship modelling of ACE-inhibitory peptides derived from milk proteins. Eur. Food Res. Technol. 2004, 219, 579–583. [Google Scholar] [CrossRef]
- Gómez-Ruiz, J.Á.; Taborda, G.; Amigo, L.; Recio, I.; Ramos, M. Identification of ACE-inhibitory peptides in different Spanish cheeses by tandem mass spectrometry. Eur. Food Res. Technol. 2006, 223, 595–601. [Google Scholar] [CrossRef]
- Gómez-Ruiz, J.A.; Recio, I.; Belloque, J. ACE-Inhibitory Activity and Structural Properties of Peptide Asp-Lys-Ile-His-Pro [β-CN f(47−51)]. Study of the Peptide Forms Synthesized by Different Methods. J. Agric. Food Chem. 2004, 52, 6315–6319. [Google Scholar] [CrossRef]
- Tonouchi, H.; Suzuki, M.; Uchida, M.; Oda, M. Antihypertensive effect of an angiotensin converting enzyme inhibitory peptide from enzyme modified cheese. J. Dairy Sci. 2008, 75, 284–290. [Google Scholar] [CrossRef]
- Nilsen, R.; Pripp, A.H.; Høstmark, A.T.; Haug, A.; Skeie, S. Short communication: Is consumption of a cheese rich in angiotensin-converting enzyme-inhibiting peptides, such as the Norwegian cheese Gamalost, associated with reduced blood pressure? J. Dairy Sci. 2014, 97, 2662–2668. [Google Scholar] [CrossRef] [Green Version]
- Crippa, G.; Zabzuni, D.; Bravi, E.; Piva, G.; De Noni, I.; Bighi, E.; Rossi, F. Randomized, double blind placebo-controlled pilot study of the antihypertensive effects of Grana Padano D.O.P. cheese consumption in mild—Moderate hypertensive subjects. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 7573–7581. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.-C.; Wang, Y.; Tong, X.; Szeto, I.M.Y.; Smit, G.; Li, Z.-N.; Qin, L.-Q. Cheese consumption and risk of cardiovascular disease: A meta-analysis of prospective studies. Eur. J. Nutr. 2017, 56, 2565–2575. [Google Scholar] [CrossRef]
- Sarmadi, B.H.; Ismail, A. Antioxidative peptides from food proteins: A review. Peptides 2010, 31, 1949–1956. [Google Scholar] [CrossRef]
- Gupta, A.; Mann, B.; Kumar, R.; Sangwan, R.B. Antioxidant activity of Cheddar cheeses at different stages of ripening. Int. J. Dairy Technol. 2009, 62, 339–347. [Google Scholar] [CrossRef]
- Gupta, A.K.; Mann, B.; Kumar, R.; Sangwan, R. Identification of antioxidant peptides in cheddar cheese made with adjunct culture Lactobacillus casei ssp. casei 300. Milchwissenschaft 2010, 65, 396–399. [Google Scholar]
- Pritchard, S.R.; Phillips, M.; Kailasapathy, K. Identification of bioactive peptides in commercial Australian organic cheddar cheeses. Aust. J. Dairy Technol. 2010, 65, 170–173. [Google Scholar] [CrossRef]
- Hernández-Galán, L.; Cardador-Martínez, A.; López-del-Castillo, M.; Picque, D.; Spinnler, H.E.; Martín del Campo, S.T. Antioxidant and angiotensin-converting enzyme inhibitory activity in fresh goat cheese prepared without starter culture: A preliminary study. CyTA J. Food 2016, 15, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Timón, M.L.; Andrés, A.; Otte, J.; Petrón, M.J. Antioxidant peptides (<3 kDa) identified on hard cow milk cheese with rennet from different origin. Food Res. Int. 2019, 120, 643–649. [Google Scholar] [CrossRef]
- Udenigwe, C.C.; Aluko, R.E. Chemometric analysis of the amino acid requirements of antioxidant food protein hydrolysates. Int. J. Mol. Sci. 2011, 12, 3148–3161. [Google Scholar] [CrossRef] [Green Version]
- Timón, M.L.; Parra, V.; Otte, J.; Broncano, J.M.; Petrón, M.J. Identification of radical scavenging peptides (<3 kDa) from Burgos-type cheese. LWT-Food Sci. Technol. 2014, 57, 359–365. [Google Scholar] [CrossRef]
- Cox, R.A.; García-Palmieri, M.R. Cholesterol, Triglycerides, and Associated Lipoproteins. In Clinical Methods: The History, Physical, and Laboratory Examinations, 3rd ed.; Walker, H.K., Hall, W.D., Hurst, J.W., Eds.; Butterworths: Boston, MA, USA, 1990; Chapter 31; pp. 153–160. [Google Scholar]
- Contarini, G.; Povolo, M. Phospholipids in milk fat: Composition, biological and technological significance, and analytical strategies. Int. J. Mol. Sci. 2013, 14, 2808–2831. [Google Scholar] [CrossRef] [Green Version]
- Udenigwe, C.C.; Rouvinen-Watt, K. The Role of Food Peptides in Lipid Metabolism during Dyslipidemia and Associated Health Conditions. Int. J. Mol. Sci. 2015, 16, 9303–9313. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, C.-C.; Hernández-Ledesma, B.; Fernández-Tomé, S.; Weinborn, V.; Barile, D.; de Moura Bell, J.M.L.N. Milk Proteins, Peptides, and Oligosaccharides: Effects against the 21st Century Disorders. Biomed Res. Int. 2015, 2015, 146840. [Google Scholar] [CrossRef]
- Nagaoka, S. Structure-function properties of hypolipidemic peptides. J. Food Biochem. 2019, 43, e12539. [Google Scholar] [CrossRef] [Green Version]
- Wakasa, Y.; Tamakoshi, C.; Ohno, T.; Hirose, S.; Goto, T.; Nagaoka, S.; Takaiwa, F. The Hypocholesterolemic Activity of Transgenic Rice Seed Accumulating Lactostatin, a Bioactive Peptide Derived from Bovine Milk β-Lactoglobulin. J. Agric. Food Chem. 2011, 59, 3845–3850. [Google Scholar] [CrossRef] [PubMed]
- Cabanos, C.; Ekyo, A.; Amari, Y.; Kato, N.; Kuroda, M.; Nagaoka, S.; Takaiwa, F.; Utsumi, S.; Maruyama, N. High-level production of lactostatin, a hypocholesterolemic peptide, in transgenic rice using soybean A1aB1b as carrier. Transgenic Res. 2013, 22, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, R.; Ohinata, K.; Yoshikawa, M. β-Lactotensin and neurotensin rapidly reduce serum cholesterol via NT2 receptor. Peptides 2003, 24, 1955–1961. [Google Scholar] [CrossRef]
- Keller, U. Dietary Proteins in Obesity and in Diabetes. Int. J. Vitam. Nutr. Res. 2011, 81, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Campos-Nonato, I.; Hernandez, L.; Barquera, S. Effect of a High-Protein Diet versus Standard-Protein Diet on Weight Loss and Biomarkers of Metabolic Syndrome: A Randomized Clinical Trial. Obes Facts. 2017, 10, 238–251. [Google Scholar] [CrossRef] [PubMed]
- Erdmann, K.; Cheung, B.W.; Schröder, H. The possible role of food-derived peptides in reducing the risk of cardiovascular disease. J. Nutr. Biochem. 2008, 19, 643–654. [Google Scholar] [CrossRef] [PubMed]
- Bhat, Z.F.; Kumar, S.; Bhat, H.F. Bioactive peptides of animal origin: A review. J. Food Sci. Technol. 2015, 52, 5377–5392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondrashina, A.; Brodkorb, A.; Giblin, L. Dairy-derived peptides in satiety. J. Funct. Foods 2020, 66, 103801. [Google Scholar] [CrossRef]
- Monteiro, R.; Azevedo, I. Chronic inflammation in obesity and the metabolic syndrome. Mediat. Inflamm. 2010, 2010, 289645. [Google Scholar] [CrossRef]
- Zielińska, E.; Baraniak, B.; Karaś, M. Antioxidant and Anti-Inflammatory Activities of Hydrolysates and Peptide Fractions Obtained by Enzymatic Hydrolysis of Selected Heat-Treated Edible Insects. Nutrients 2017, 9, 970. [Google Scholar] [CrossRef] [Green Version]
- Marcone, S.; Belton, O.; FitzGerald, D.J. Milk-derived bioactive peptides and their health-promoting effects: A potential role in atherosclerosis. Br. J. Clin. Pharmacol. 2017, 83, 152–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rafiq, S.; Huma, N.; Rakariyatham, K.; Hussain, I.; Gulzar, N.; Hayat, I. Anti-inflammatory and anticancer activities of water-soluble peptide extracts of buffalo and cow milk Cheddar cheeses. Int. J. Dairy Technol. 2017, 71, 432–438. [Google Scholar] [CrossRef]
- Minkiewicz, P.; Iwaniak, A.; Darewicz, M. BIOPEP-UWM Database of Bioactive Peptides: Current Opportunities. Int. J. Mol. Sci. 2019, 20, 5978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwaniak, A.; Darewicz, M.; Mogut, D.; Minkiewicz, P. Elucidation of the role of in silico methodologies in approaches to studying bioactive peptides derived from foods. J. Funct. Foods 2019, 61, 103486. [Google Scholar] [CrossRef]
Figure 1.
Metabolic syndrome criteria and related preventive agents according to Huang [4].
Figure 1.
Metabolic syndrome criteria and related preventive agents according to Huang [4].

Figure 2.
The involvement of the dipeptidyl peptidase IV inhibitors in prolonging the incretin effect according to Mkele [16].
Figure 2.
The involvement of the dipeptidyl peptidase IV inhibitors in prolonging the incretin effect according to Mkele [16].

Figure 3.
The delay of carbohydrate breakdown by α-glucosidase and α-amylase inhibitors according to Iwaniak et al. [5] and Patil et al. [18].

Figure 4.
Brief description of the mechanism of action of ACE and its inhibitors according to Iwaniak et al. [26].
Figure 4.
Brief description of the mechanism of action of ACE and its inhibitors according to Iwaniak et al. [26].

Figure 5.
The preventive activity of antioxidants on the formation of diseases according to Sarmadi and Ismail [42].
Figure 5.
The preventive activity of antioxidants on the formation of diseases according to Sarmadi and Ismail [42].

Table 1.
Quantification of VPP and IPP in cheeses of Swiss origin with their corresponding ACE inhibitory activity expressed with IC50 values. Adapted with permission from [29], Copyright Elsevier, 2007.
Table 1.
Quantification of VPP and IPP in cheeses of Swiss origin with their corresponding ACE inhibitory activity expressed with IC50 values. Adapted with permission from [29], Copyright Elsevier, 2007.
| Cheese | VPP | IPP | IC50 |
|---|---|---|---|
| (mg × kg−1) | (mg × kg−1) | (mg cheese × mL−1) | |
| Hard cheeses | |||
| Emmental: | |||
| cave-aged, aged 12 months | 100.1 | 23.5 | 7.1 ± 0.2 |
| reserve, aged 8 months | 68.6 | 13.2 | 9.9 ± 0.2 |
| organic, aged 4 months | 153.7 | 35.8 | 7.5 ± 1.8 |
| classic, aged 4 months | 42.6 | 4.6 | 10.5 ± 0.6 |
| Gouda old | 97.7 | 90.1 | 2.0 ± 0.3 |
| Gruyere AOC: | |||
| vieux, aged 12 months | 13.7 | 7.9 | 29.4 ± 16.8 |
| salé, aged 10 months | 68.7 | 20.4 | 14.2 ± 0.9 |
| mi-salé, aged 8 months | 22.7 | 14.9 | 18.7 ± 6.1 |
| doux, aged 5 months | 53.0 | 9.3 | 14.9 ± 0.7 |
| Hobelkäse from the Bernese Oberland | *224.1 | 95.4 | 2.6 ± 0.2 |
| Parmino | 2.0 | 0.8 | 28.6 ± 4.2 |
| Sbrinz | 28.0 | 9.4 | 16.4 ± 5.5 |
| Semi-hard cheeses | |||
| Appenzeller: | |||
| aged 7–10 months (1/4 fat) | 94.4 | 64.9 | 4.2 ± 1.6 |
| extra, aged 6–7 months | 29.3 | 14.5 | 16.8 ± 1.1 |
| surchoix, aged 4 months | 26.7 | 17.8 | 8.0 ± 1.0 |
| aged 3 months (1/4 fat) | 56.0 | 10.5 | 7.1 ± 1.9 |
| classic, aged 3 months | 22.0 | 8.5 | 17.6 ± 7.6 |
| Edam | 1.0 | 0.1 | 13.3 ± 0.4 |
| Küsnachter | 25.7 | 18.0 | 19.6 ± 2.4 |
| Mountain cheese Graubünden: | |||
| ¼ fat | 32.0 | 22.7 | 4.8 ± 0.0 |
| ½ fat | 10.4 | 7.1 | 8.9 ± 4.1 |
| full fat | 25.1 | 4.3 | 9.2 ± 1.4 |
| Raclette: | |||
| from raw milk | 12.6 | 2.6 | 8.8 ± 0.0 |
| from pasteurized milk | 1.0 | 0.1 | 9.0 ± 0.3 |
| St. Paulin | 5.2 | 1.0 | 23.5 ± 6.3 |
| Tête de Moine | 98.0 | 15.5 | 7.1 ± 1.9 |
| Tilsit: | |||
| from raw milk | 120.2 | 30.0 | 4.4 ± 0.2 |
| from pasteurized milk | 1.2 | 0.1 | 18.0 ± 6.3 |
| Vacherin fribourgeois | 62.1 | 46.7 | 5.0 ± 0.3 |
| Wangener Geissmutschli (goat cheese) | 33.2 | 0.0 | 7.9 ± 0.5 |
| Winzerkäse | 97.7 | 27.1 | 9.9 ± 1.8 |
| Soft cheeses | |||
| Brie | 0.4 | 0.3 | 25.9 ± 3.0 |
| Camembert | 0.2 | 0.2 | Very weak |
| Mozarella: | |||
| from buffalo milk | 0.1 | 0.0 | - |
| from cow milk | 0.0 | 0.0 | - |
| Tomme vaudoise | 0.0 | 0.0 | 14.9 ± 3.7 |
| Vacherin Mont d’Or | 0.2 | 0.1 | 21.4 ± 9.6 |
*Bold - the highest concentration of peptide in the analyzed category (hard, semi-hard, and soft cheese).
Table 2.
The Concentration of YPFPGPI in commercial cheeses before and after in vitro simulated gastro-intestinal digestion. Adapted with permission from [33], Copyright Elsevier, 2010.
Table 2.
The Concentration of YPFPGPI in commercial cheeses before and after in vitro simulated gastro-intestinal digestion. Adapted with permission from [33], Copyright Elsevier, 2010.
| Cheese | Concentration of YPFPGPI (mg × kg−1) | |
|---|---|---|
| Before Simulated Digestion | After Simulated Digestion | |
| Brie | 0.15 | 2.07 ± 0.09 |
| Caprino | 0 | 1.51 ± 0.09 |
| Cheddar | 0.11 | 15.22 ± 0.13 |
| Fontina | 0.04 | 10.43 ± 0.12 |
| Gorgonzola | 0.01 | 5.41 ± 0.08 |
| Gouda | 0.10 | 21.77 ± 0.12 |
| Grana Padano (aged 25 months) | 0 | 11.06 ± 0.08 |
| Grana Padano (aged 17 months) | 0 | 12.55 ± 0.09 |
| Grana Padano (aged 10 months) | 0 | 8.79 ± 0.04 |
| Taleggio | 0 | 7.15 ± 0.07 |
Table 3.
ACE inhibitors identified in Spanish cheeses. Adapted by permission from Springer, European Food Research and Technology [36], 2006.
Table 3.
ACE inhibitors identified in Spanish cheeses. Adapted by permission from Springer, European Food Research and Technology [36], 2006.
| Cheese | Peptide | IC50 (µM) | Precursor | Reference |
| Manchego, Roncal, Idiazábal, Cabrales, goat | DKIHP | 113.1 | β-casein | [36] |
| DKIHPF | 2419.4 | |||
| Goat | VRGP | 120.9 | ||
| Manchego | PFP | 144.4 | various fragments | [36] |
| FP | 1215.7 | |||
| PP | 2284.7 | |||
| KKYNVPQL | 77.2 | αs1-casein | [30] | |
| VRYL | 24.1 | |||
| Manchego, Roncal, Idiazabal, Goat | QP | 598.1 | various fragments | [36] |
| Roncal | PKHP | 709.1 | αS1- casein |
Table 4.
The summary of casein-derived peptides showing more than one biological activity (based on BIOPEP-UWM database screening; http://www.uwm.edu.pl/biochemia; accessed on March 2020).
Table 4.
The summary of casein-derived peptides showing more than one biological activity (based on BIOPEP-UWM database screening; http://www.uwm.edu.pl/biochemia; accessed on March 2020).
| Dual and Triple Activity of Casein-Derived Peptides | Number of Peptides Encrypted in the All Casein Sequences |
|---|---|
| ACE and DPP4 inhibitor | 66 |
| ACE inhibitor and antioxidative | 17 |
| DPP4 and α-glucosidase inhibitor | 1 |
| DPP4 inhibitor and opioid | 1 |
| ACE inhibitor and opioid | 5 |
| DPP4 inhibitor and antioxidative | 5 |
| ACE, DPP4 inhibitor, and antioxidative | 8 |
| ACE, DPP4, and α-glucosidase inhibitor | 1 |
| Total | 104 |
| Peptides Showing one Bioactivity | |
| ACE inhibitor | 171 |
| α-glucosidase inhibitor | 0 |
| antioxidative | 52 |
| DPP4 inhibitor | 127 |
| opioid agonist | 3 |
| opioid antagonist | 2 |
| opioid | 4 |
| Total | 359 |
| Total Number of Bioactive Sequences | 463 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Iwaniak, A.; Mogut, D. Metabolic Syndrome-Preventive Peptides Derived from Milk Proteins and Their Presence in Cheeses: A Review. Appl. Sci. 2020, 10, 2772. https://0-doi-org.brum.beds.ac.uk/10.3390/app10082772
AMA Style
Iwaniak A, Mogut D. Metabolic Syndrome-Preventive Peptides Derived from Milk Proteins and Their Presence in Cheeses: A Review. Applied Sciences. 2020; 10(8):2772. https://0-doi-org.brum.beds.ac.uk/10.3390/app10082772
Chicago/Turabian StyleIwaniak, Anna, and Damir Mogut. 2020. "Metabolic Syndrome-Preventive Peptides Derived from Milk Proteins and Their Presence in Cheeses: A Review" Applied Sciences 10, no. 8: 2772. https://0-doi-org.brum.beds.ac.uk/10.3390/app10082772
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.