
Allelopathic Interactions between Seeds of Portulaca oleracea L. and Crop Species

 , , , and
, , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Statistical Analysis
3. Results
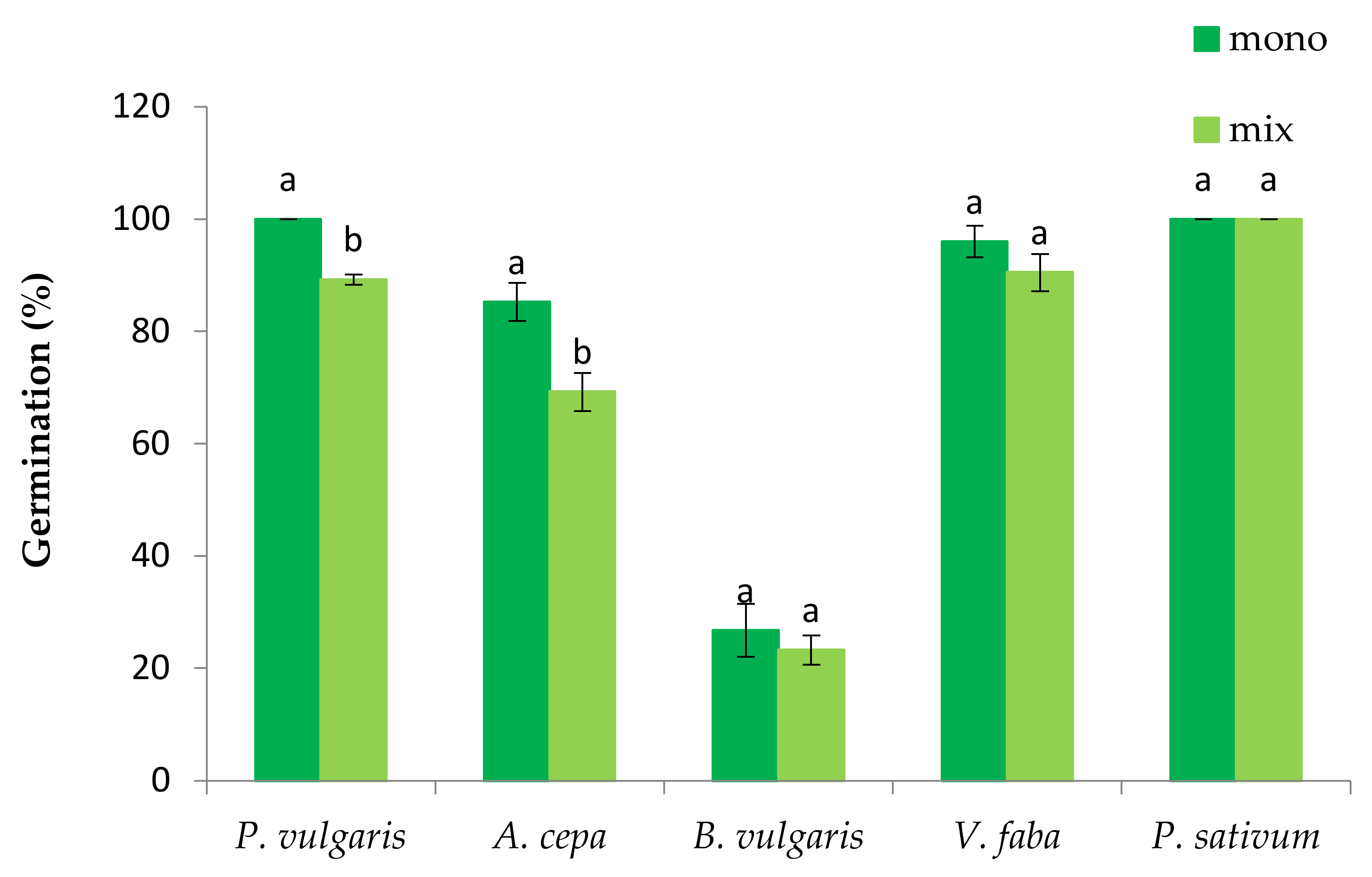
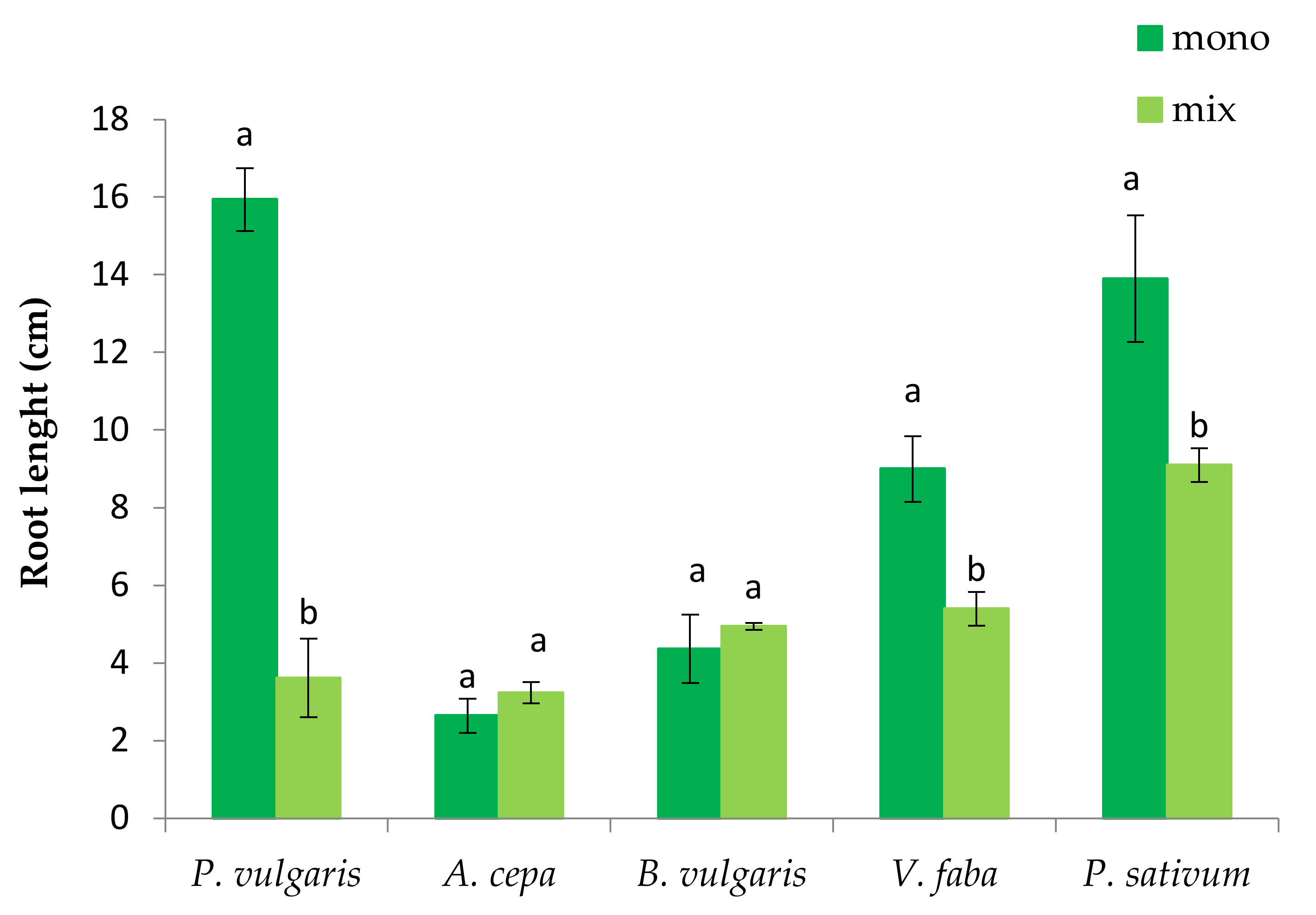
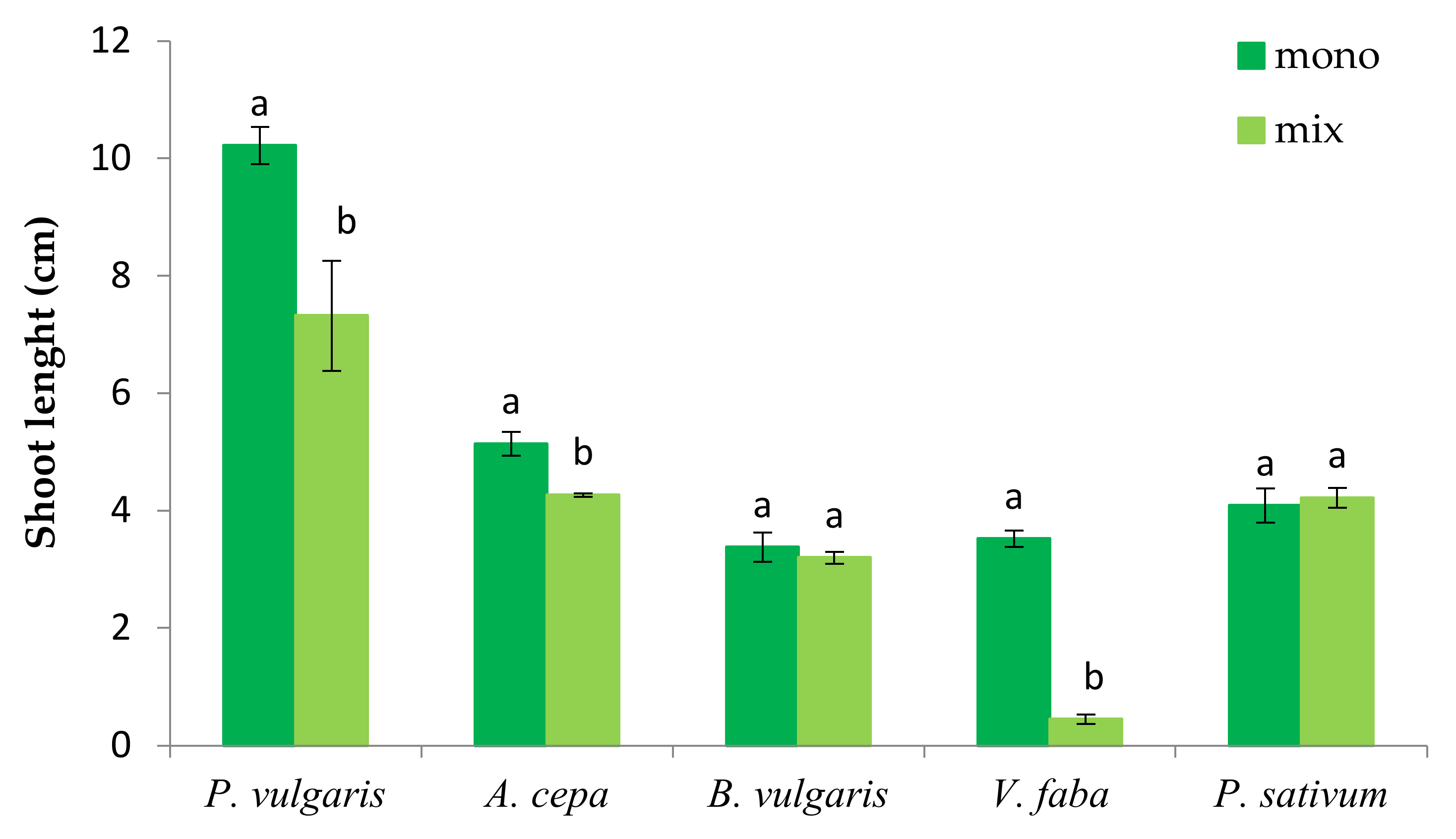
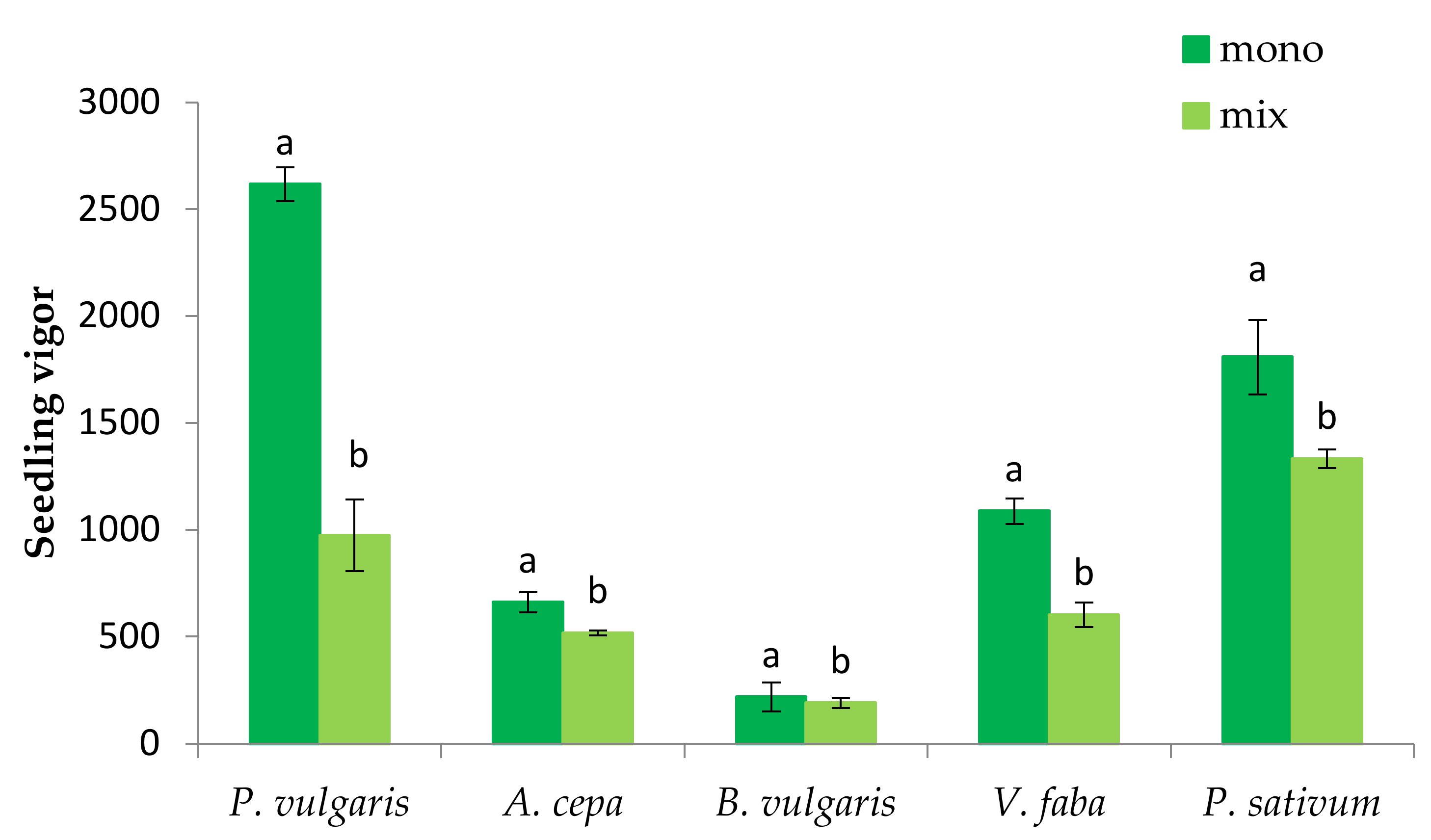
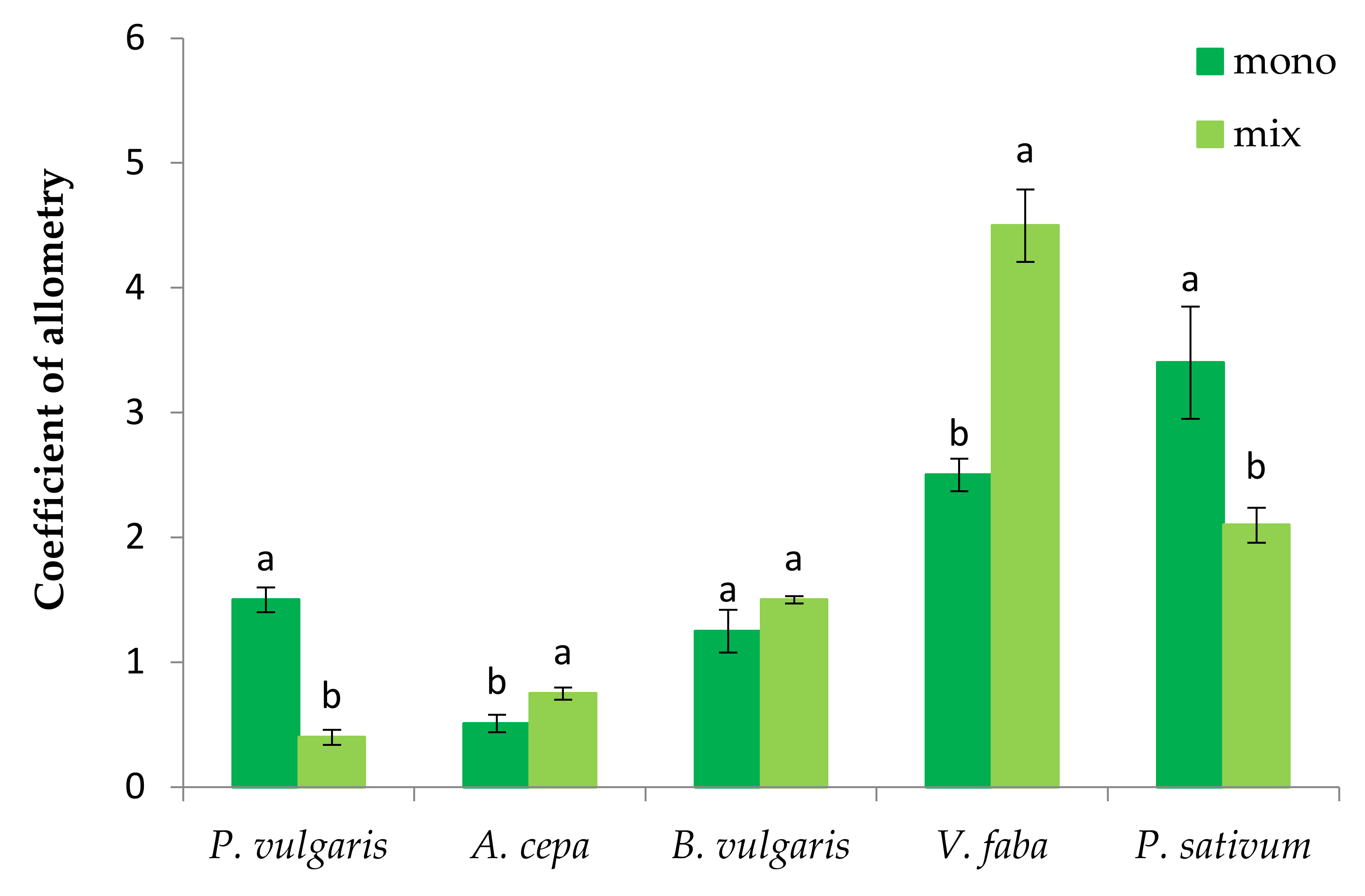
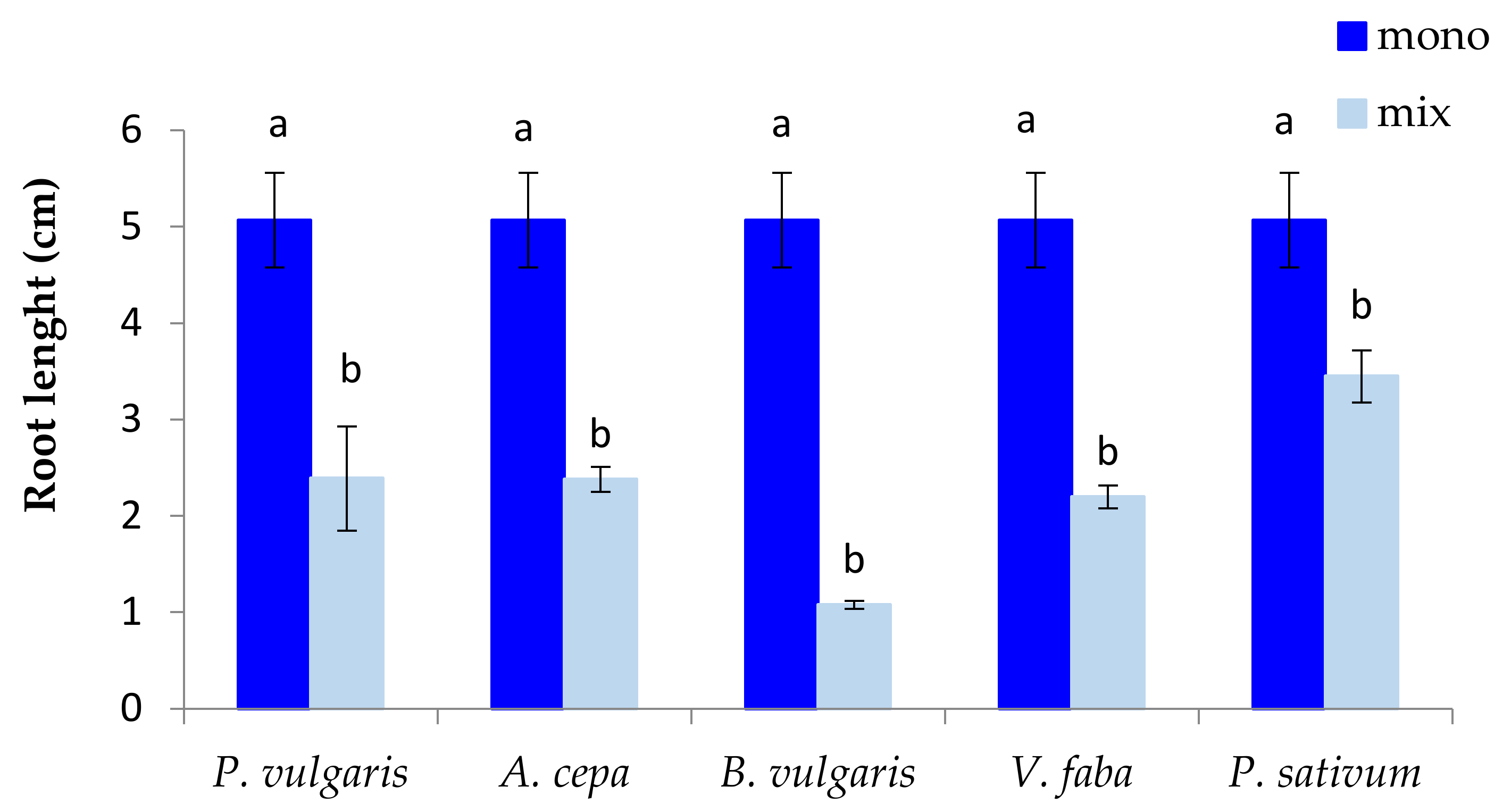
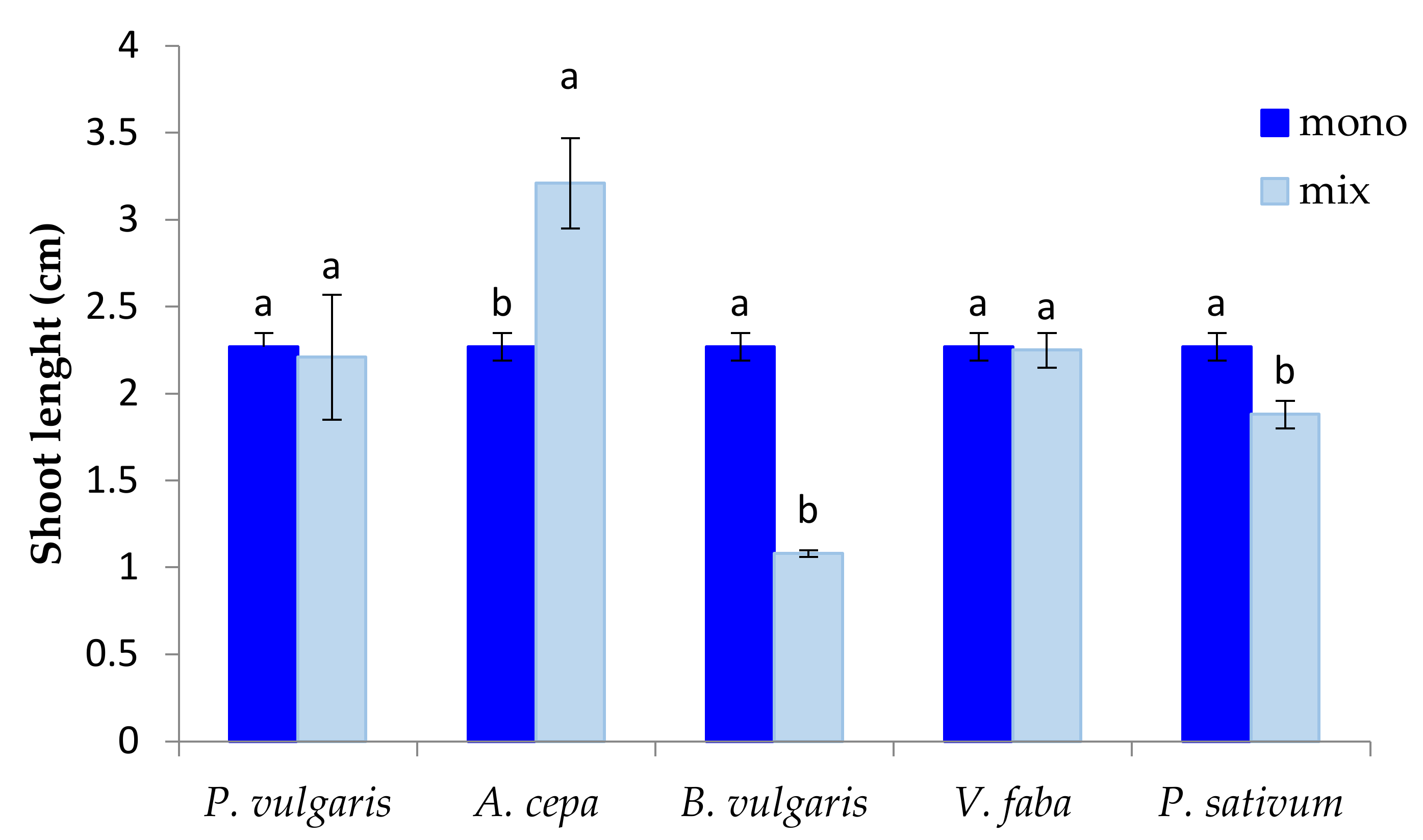
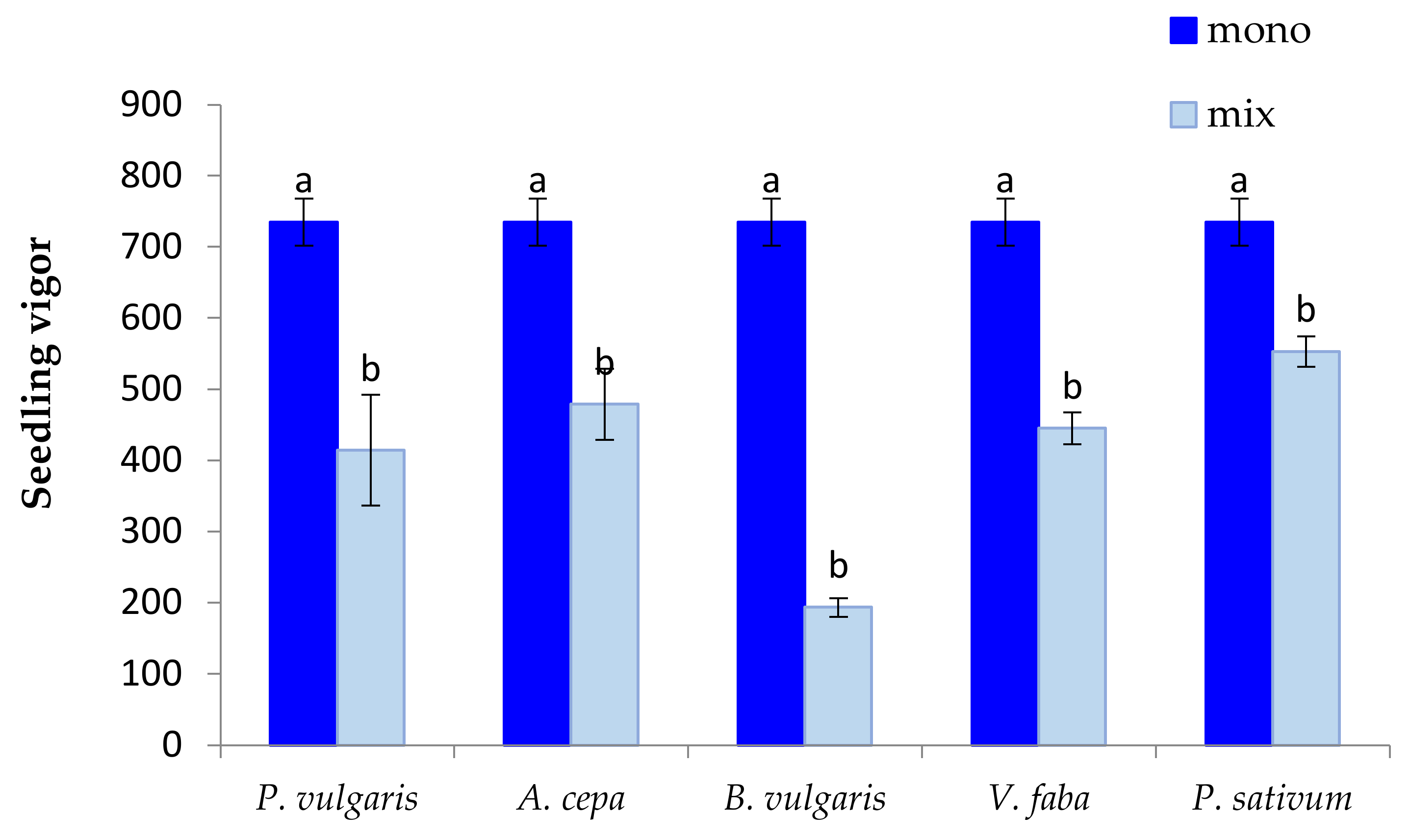
3.1. The Effect of P. oleracea on the Germination and Seedling Growth of Crop Species
3.2. The Effect of Crop Species on the Germination and Seedling Growth of P. oleracea
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zimdahl, R.L. Fundamentals of Weed Science; Academic Press: London, UK, 2018. [Google Scholar]
- Le Tourneau, D.; Failes, G.D.; Heggeness, H.G. The effect of aqueous extracts of plant tissue on germination of seeds and growth of seedlings. Weeds 1956, 4, 363–368. [Google Scholar] [CrossRef]
- Cheema, Z.; Khaliq, A. Use of sorghum allelopathic properties to control weeds in irrigated wheat in a semi arid region of Punjab. Agric. Ecosyst. Environ. 2000, 79, 105–112. [Google Scholar] [CrossRef]
- Heidarzade, A.; Pirdashti, H.; Esmaeili, M. Esmaeili, Quantification of allelopathic substances and inhibitory potential in root exudates of rice (Oryza sativa) varieties on barnyardgrass (Echinochloa crus-galli L.). Plant Omics 2010, 3, 204–209. [Google Scholar]
- Gholami, P.; Ghorbani, J.; Ghaderi, S. Allelopathic effects of Artemisia aucheri on seed germination and Dactylis glomerata properties of Festuca arundinacea Schreb. Plant Ecophysiol. 2011, 9, 42–52, (In Persian with English Summary). [Google Scholar]
- Afendi, F.M.; Okada, T.; Yamazaki, M.; Hirai-Morita, A.; Nakamura, Y.; Nakamura, K.; Ikeda, S.; Takahashi, H.; Altaf-Ul-Amin, M.; Darusman, L.K.; et al. KNApSAcK family databases: Integrated metabolite—Plant species databases for multifaceted plant research. Plant Cell Physiol. 2012, 53, e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahuja, I.; Kissen, R.; Bones, A.M. Phytoalexins in defense against pathogens. Trends Plant Sci. 2012, 17, 73–90. [Google Scholar] [CrossRef]
- Weston, L.A.; Duke, S.O. Weed and crop allelopathy. Crit. Rev. Plant Sci. 2003, 22, 367–389. [Google Scholar] [CrossRef]
- Jabran, K.; Mahajan, G.; Sardana, V.; Chauhan, B.S. Allelopathy for weed control in agricultural systems. Crop Prot. 2015, 72, 57–65. [Google Scholar] [CrossRef]
- Muzell Trezzi, M.; Vidal, R.A.; Balbinot Junior, A.A.; von Hertwig Bittencourt, H.; da Silva Souza Filho, A.P. Allelopathy: Driving mechanisms governing its activity in agriculture. J. Plant Interact. 2016, 11, 53–60. [Google Scholar] [CrossRef]
- Ogata, T.; Hamachi, M.; Nishi, K. Organic Herbicide for Paddy Field. Japan Patent No. JP2008050329A, 6 March 2008. [Google Scholar]
- Gorbanli, M.L.; Bakhshi, K.G.R.; Shojaei, A.A. Examination of the effects of Allelopathy of Artemisia sieberi Besser Subsp. sieberi on seed germination and Avena lodoviciana and Amaranthus retroflexus seedlings growth. Pajouhesh-Va-Sazandegi 2008, 21, 129–134, (In Persian with English Summary). [Google Scholar]
- Scrivanti, L.R.; Anton, A.M.; Zygadlo, J.A. Allelopathic potential of South American Bothriochloa species (Poaceae: Andropogoneae). Allelopath. J. 2011, 28, 189–200. [Google Scholar]
- Holm, L.G.; Plucknett, D.L.; Pancho, J.V.; Herberger, J.P. The World’s Worst Weeds: Distribution and Biology; University Press of Hawaii: Honolulu, HI, USA, 1977. [Google Scholar]
- Xin, H.L.; Xu, Y.F.; Hou, Y.H.; Zhang, Y.N.; Yue, X.Q.; Lu, J.C.; Ling, C.Q. Two novel triterpenoids from Portulaca oleracea L. Helv. Chim. Acta 2008, 91, 2075–2080. [Google Scholar] [CrossRef]
- Keys, J.D. Chinese Herbs: Their Botany, Chemistry, and Pharmacodynamics; Charles, E., Ed.; Tuttle Company, Inc.: Rutland, VT, USA; Tokyo, Japan, 1976. [Google Scholar]
- Iwu, M. Handbook of African Medicinal Plants; CRC Press: Boca Raton, FL, USA, 1993; pp. 183–184. [Google Scholar]
- Xiang, L.; Xing, D.; Wang, W.; Wang, R.; Ding, Y.; Du, L. Alkaloids from Portulaca oleracea L. Phytochemistry 2005, 66, 2595–2601. [Google Scholar] [CrossRef] [PubMed]
- Erkan, N. Antioxidant activity and phenolic compounds of fractions from Portulaca oleracea L. Food Chem. 2012, 133, 775–781. [Google Scholar] [CrossRef]
- Zhu, H.; Wang, Y.; Liu, Y.; Xia, Y.; Tang, T. Analysis of flavonoids in Portulaca oleracea L. by UV–vis spectrophotometry with comparative study on different extraction technologies. Food Anal. Methods 2010, 3, 90–97. [Google Scholar] [CrossRef]
- Silva, M.; Magrico, S.; Dias, A.S.; Dias, L.S. Allelopathic plants. 20. Portulaca oleracea L. Allelopathy J. 2007, 19, 275–286. [Google Scholar]
- Wang, C.; Liu, J.; Xiao, H.; Du, D. Response of Leaf Functional Traits of Cerasus yedoensis (Mats.) Yü Li to Serious Insect Attack. Pol. J. Environ. Stud. 2016, 25, 333–339. [Google Scholar] [CrossRef]
- Agrawal, R. Seed Technology; Oxford IBH Publishing: Oxford, UK, 1991; 658p. [Google Scholar]
- Kulkarni, M.G.; Sparg, S.G.; Van Staden, J. Germination and post-germination response of Acacia seeds to smoke-water and butenolide, a smoke-derived compound. J. Arid Environ. 2007, 69, 177–187. [Google Scholar] [CrossRef]
- Saxena, A.; Singh, D.V.; Joshi, N.L. Autotoxic effects of pearl millet aqueous extracts on seed germination and seedling growth. J. Arid Environ. 1996, 33, 255–260. [Google Scholar] [CrossRef]
- Yue, M.E.; Jiang, T.F.; Shi, Y.P. Simultaneous determination of noradrenaline and dopamine in Portulaca oleracea L. by capillary zone electrophoresis. J. Sep. Sci. 2005, 28, 360–364. [Google Scholar] [CrossRef]
- Haig, T. Allelochemicals in plants. In Allelopathy in Sustainable Agriculture and Forestry; Springer: New York, NY, USA, 2008; pp. 63–104. [Google Scholar]
- Kaletha, M.S.; Bhatt, B.P.; Todaria, N.P. Tree-crop interactions in traditional agroforestry systems of Garhwal Himalaya. 1. Phytotoxic effects of farm trees on food crops. Allelopath. J. 1996, 3, 247–250. [Google Scholar]
- Mishra, A. Allelopathic properties of Lantana camara: A review article. Int. J. Innov. Res. Rev. 2014, 2, 32–52. [Google Scholar]
- Sharma, S.B.; Nasir, A.; Prabhu, K.M.; Murthy, P.S.; Dev, G. Hypoglycaemic and hypolipidemic effect of ethanolic extract of seeds of Eugenia jambolana in alloxan-induced diabetic rabbits. J. Ethnopharmacol. 2003, 85, 201–206. [Google Scholar] [CrossRef]
- Dadkhah, A. Allelopathic effect of sugar beet (Beta vulgaris) and eucalyptus (Eucalyptus camaldulensis) on seed germination and growth of Portulaca oleracea. Russ. Agric. Sci. 2013, 39, 117–123. [Google Scholar] [CrossRef]
- El-Shora, H.M.; Abd El-Gawad, A.M. Physiological and biochemical responses of Cucurbita pepo L. mediated by Portulaca oleracea L. allelopathy. Fresenius Environ. Bull. J. 2015, 24, 386–393. [Google Scholar]
- Galindo, J.C.; Molinillo, J.M.; Macias, F.A. Allelopathy: Chemistry and Mode of Action of Allelochemicals; CRC Press: Boca Raton, FL, USA, 2003. [Google Scholar]
- Furuya, M.; Galston, A.W.; Stowe, B.B. Isolation from peas of co-factors and inhibitors of indolyl-3-acetic acid oxidase. Nature 1962, 193, 456–457. [Google Scholar] [CrossRef]
- Stenlid, G. The effects of flavonoid compounds on oxidative phosphorylation and on the enzymatic destruction of indoleacetic acid. Physiol. Plant. 1963, 16, 110–120. [Google Scholar] [CrossRef]
- Mathesius, U. Flavonoids induced in cells undergoing nodule organogenesis in white clover are regulators of auxin breakdown by peroxidase. J. Exp. Bot. 2001, 52, 419–426. [Google Scholar] [CrossRef]
- Jacobs, M.; Rubery, P.H. Naturally occurring auxin transport regulators. Science 1988, 241, 346–349. [Google Scholar] [CrossRef] [PubMed]
- Peer, W.A.; Brown, D.E.; Tague, B.W.; Muday, G.K.; Taiz, L.; Murphy, A.S. Flavonoid accumulation patterns of transparent testa mutants of Arabidopsis. Plant Physiol. 2001, 126, 536–548. [Google Scholar] [CrossRef] [Green Version]
- Levizou, E.; Karageorgou, P.; Petropoulou, Y.; Grammatikopoulos, G.; Manetas, Y. Induction of ageotropic response in lettuce radicle growth by epicuticular flavonoid aglycons of Dittrichia viscosa. Biol. Plant. 2004, 48, 305–307. [Google Scholar] [CrossRef]
- Kiran, B.; Lalitha, V.; Raveesha, K.A. In vitro evaluation of aqueous seed extract of Psoralea corylifolia L. on seed germination and seedling vigour of maize. J. Appl. Pharm. Sci. 2011, 1, 128–130. [Google Scholar]
- Dhungana, S.K.; Kim, I.D.; Adhikari, B.; Kim, J.H.; Shin, D.H. Reduced Germination and Seedling Vigor of Weeds with Root Extracts of Maize and Soybean, and the Mechanism Defined as Allelopathic. J. Crop Sci. Biotechnol. 2019, 22, 11–16. [Google Scholar] [CrossRef]
- Pasricha, V.; Satpathy, G.; Gupta, R.K. Phytochemical & Antioxidant activity of underutilized legume Vicia faba seeds and formulation of its fortified biscuits. J. Pharmacogn. Phytochem. 2014, 3, 75–80. [Google Scholar]
- Geil, P.B.; Anderson, J.W. Nutrition and health implications of dry beans: A review. J. Am. Coll. Nutr. 1994, 13, 549–558. [Google Scholar] [CrossRef] [PubMed]
- Mao, J.; Yang, L.; Shi, Y.; Hu, J.; Piao, Z.; Mei, L.; Yin, S. Crude extract of Astragalus mongholicus root inhibits crop seed germination and soil nitrifying activity. Soil Biol. Biochem. 2006, 38, 201–208. [Google Scholar] [CrossRef]
- Farooq, M.; Jabran, K.; Cheema, Z.A.; Wahid, A.; Siddique, K.H. The role of allelopathy in agricultural pest management. Pest Manag. Sci. 2011, 67, 493–506. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rashidi, S.; Reza Yousefi, A.; Goicoechea, N.; Pouryousef, M.; Moradi, P.; Vitalini, S.; Iriti, M. Allelopathic Interactions between Seeds of Portulaca oleracea L. and Crop Species. Appl. Sci. 2021, 11, 3539. https://0-doi-org.brum.beds.ac.uk/10.3390/app11083539
Rashidi S, Reza Yousefi A, Goicoechea N, Pouryousef M, Moradi P, Vitalini S, Iriti M. Allelopathic Interactions between Seeds of Portulaca oleracea L. and Crop Species. Applied Sciences. 2021; 11(8):3539. https://0-doi-org.brum.beds.ac.uk/10.3390/app11083539
Chicago/Turabian StyleRashidi, Sakineh, Ali Reza Yousefi, Nieves Goicoechea, Majid Pouryousef, Parviz Moradi, Sara Vitalini, and Marcello Iriti. 2021. "Allelopathic Interactions between Seeds of Portulaca oleracea L. and Crop Species" Applied Sciences 11, no. 8: 3539. https://0-doi-org.brum.beds.ac.uk/10.3390/app11083539