
Role of Irisin in Endocrine and Metabolic Disorders—Possible New Therapeutic Agent?

 and
and
Abstract
:1. Introduction
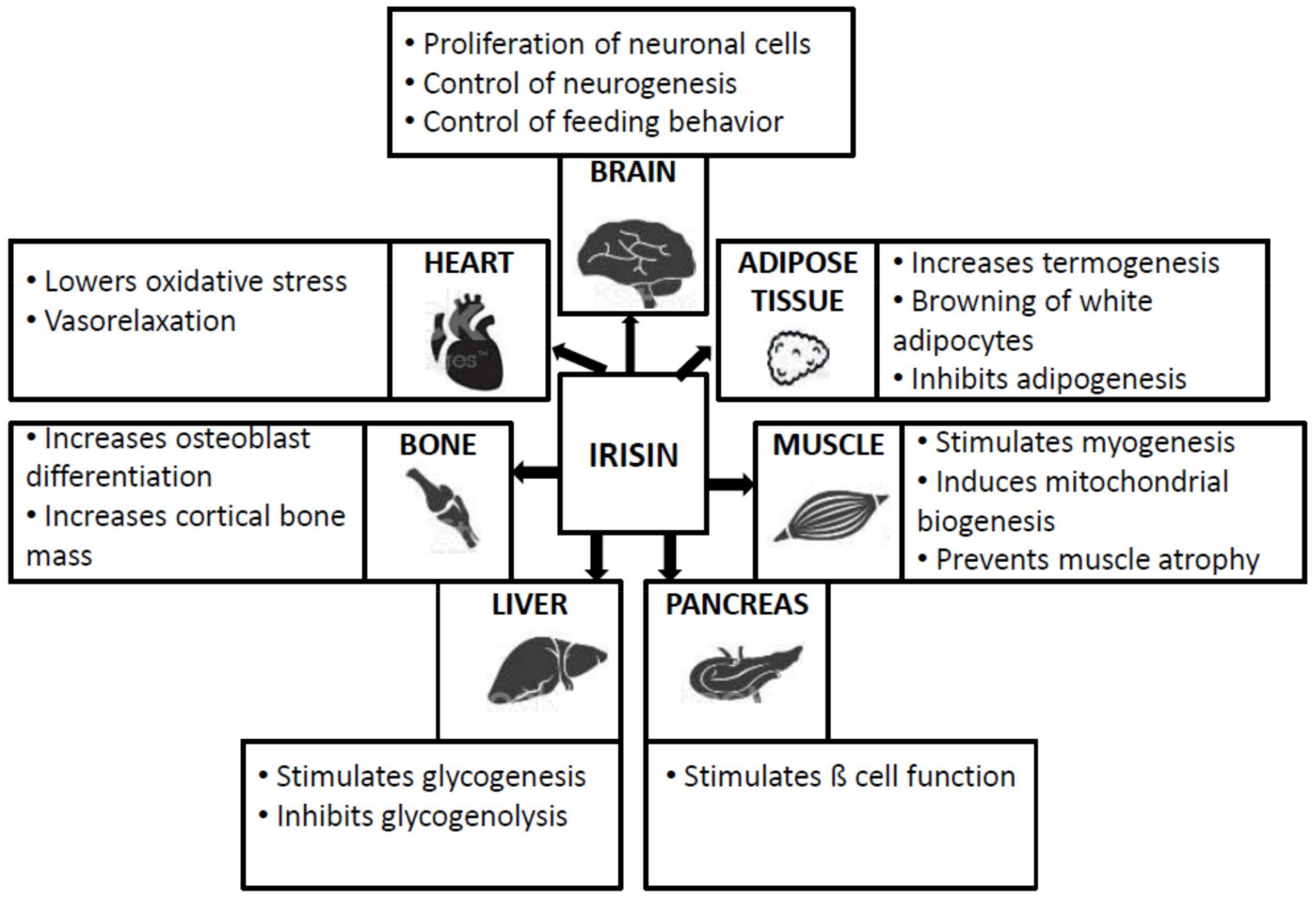
2. General Data
3. Thyroid Dysfunction
4. Small for Gestational Age (SGA)
5. Diabetes Mellitus Type 2
6. Osteoporosis
7. Prader–Willi Syndrome (PW)
8. Polycystic Ovary Syndrome
9. Obesity
10. Irisin: New Therapeutical Agent?
11. Irisin and Pharmacological Agents
12. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mahgoub, M.O.; D’Souza, C.; Al Darmaki, R.S.M.H.; Baniyas, M.M.Y.H.; Adeghate, E. An update on the role of irisin in the regulation of endocrine and metabolic functions. Peptides 2018, 104, 15–23. [Google Scholar] [CrossRef]
- Grygiel-Górniak, B.; Puszczewicz, M. A review on irisin, a new protagonist that mediates muscle-adipose-bone-neuron connectivity. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 4687–4693. [Google Scholar]
- Irving, B.A.; Still, C.D.; Argyropoulos, G. Does irisin have a brite future as a therapeutic agent in humans? Curr. Obes. Rep. 2014, 3, 235–241. [Google Scholar] [CrossRef] [Green Version]
- Perakakis, N.; Triantafyllou, G.A.; Fernández-Real, J.M.; Huh, J.Y.; Park, K.H.; Seufert, J.; Mantzoros, C.S. Physiology and role of irisin in glucose homeostasis. Nat. Rev. Endocrinol. 2017, 13, 324–337. [Google Scholar]
- Raschke, S.; Elsen, M.; Gassenhuber, H.; Sommerfeld, M.; Schwahn, U.; Brockmann, B. Evidence against a beneficial effect of irisin in humans. PLoS ONE 2013, 8, e73680. [Google Scholar]
- Colaianni, G.; Grano, M. Role of irisin on the bone-muscle functional unit. Bonekey Rep. 2015, 4, 765. [Google Scholar]
- Kaji, H. Effects of myokines on bone. Bonekey Rep. 2016, 5, 826. [Google Scholar]
- Shoukry, A.; Shalaby, S.M.; El-ArabiBdeer, S.; Mahmoud, A.A.; Mousa, M.M.; Khalifa, A. Circulating serum irisin levels in obesity and type 2 diabetes mellitus. IUBMB Life 2016, 68, 544–556. [Google Scholar] [CrossRef] [PubMed]
- Al-Daghri, N.M.; Mohammed, A.K.; Al-Attas, O.S.; Amer, O.E.; Clerici, M.; Alenad, A.; Alokail, M.S. SNPs in FNDC5 (irisin) are associated with obesity and modulation of glucose and lipid metabolism in Saudi subjects. Lipids Health Dis. 2016, 15, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rana, K.S.; Pararasa, C.; Afzal, I.; Nagel, D.A.; Hill, E.J.; Bailey, C.J.; Griffiths, H.R.; Kyrou, I.; Randeva, H.S.; Bellary, S.; et al. Plasma irisin is elevated in type 2 diabetes and is associated with increased E-selectin levels. Cardiovasc. Diabetol. 2017, 16, 147. [Google Scholar]
- Leung, P.S. The potential of irisin as a therapeutic for diabetes. Future Med. Chem. 2017, 9, 529–532. [Google Scholar] [CrossRef]
- Zhao, L.; Li, J.; Li, Z.L.; Yang, J.; Li, M.L.; Wang, G.L. Circulating irisin is lower in gestational diabetes mellitus. Endocr. J. 2015, 62, 921–926. [Google Scholar]
- Askari, H.; Rajani, S.F.; Poorebrahim, M.; Haghi-Aminjan, H.; Raeis-Abdollahi, E.; Abdollahi, M. A glance at the therapeutic potential of irisin against diseases involving inflammation, oxidative stress, and apoptosis: An introductory review. Pharmacol. Res. 2018, 129, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Marrano, N.; Biondi, G.; Borrelli, A.; Cignarelli, A.; Perrini, S.; Laviola, L.; Giorgino, F.; Natalicchio, A. Irisin and incretin hormones: Similarities, differences, and implications in type 2 diabetes and obesity. Biomolecules 2021, 11, 286. [Google Scholar] [CrossRef]
- Rabiee, F.; Lachinani, L.; Ghaedi, S.; Nasr-Esfahani, M.H.; Megraw, T.L.; Ghaedi, K. New insights into the cellular activities of Fndc5/irisin and its signaling pathways. Cell Biosci. 2020, 10, 51. [Google Scholar] [CrossRef] [PubMed]
- Moon, H.S.; Dincer, F.; Mantzoros, C.S. Pharmacological concentrations of irisin increase cell proliferation without influencing markers of neurite outgrowth and synaptogenesis in mouse H19-7 hippocampal cell lines. Metabolism 2013, 62, 1131–1136. [Google Scholar] [CrossRef] [Green Version]
- Tekin, S.; Erden, Y.; Ozyalin, F.; Cigremis, Y.; Colak, C.; Sandal, S. The effects of intracerebroventricular infusion of irisin on feeding behaviour in rats. Neurosci. Lett. 2017, 645, 25–32. [Google Scholar] [CrossRef]
- Wrann, C.D. FNDC5/irisin–their role in the nervous system and as a mediator for beneficial effects of exercise on the brain. Brain Plast. 2015, 1, 55–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaughan, R.A.; Gannon, N.P.; Barberena, M.A.; Garcia-Smith, R.; Bisoffi, M.; Mermier, C.M.; Conn, C.A.; Trujillo, K.A. Characterization of the metabolic effects of irisin on skeletal muscle in vitro. Diabetes Obes. Metab. 2014, 16, 711–718. [Google Scholar]
- Rodriguez, A.; Becerril, S.; Mendez-Gimenez, L.; Ramirez, B.; Sainz, N.; Catalan, V.; Gómez-Ambrosi, J.; Frühbeck, G. Leptin administration activates irisin-induced myogenesis via nitric oxidedependent mechanisms, but reduces its effect on subcutaneous fat browning in mice. Int. J. Obes. 2015, 39, 397–407. [Google Scholar] [CrossRef]
- Jiang, M.; Wan, F.; Wang, F.; Wu, Q. Irisin relaxes mouse mesenteric arteries through endothelium-dependent and endothelium-independent mechanisms. Biochem. Biophys. Res. Commun. 2015, 468, 832–836. [Google Scholar] [CrossRef] [PubMed]
- Han, F.; Zhang, S.; Hou, N.; Wang, D.; Sun, X. Irisin improves endothelial function in obese mice through the AMPK-eNOS pathway. Am. J. Physiol. Heart Circ. Physiol. 2015, 309, 1501–1508. [Google Scholar] [CrossRef] [Green Version]
- Xiong, X.Q.; Chen, D.; Sun, H.J.; Ding, L.; Wang, J.J.; Chen, Q.; Li, Y.H.; Zhou, Y.B.; Han, Y.; Zhang, F.; et al. FNDC5 overexpression and irisin ameliorate glucose/lipid metabolic derangements and enhance lipolysis in obesity. Biochim. Biophys. Acta 2015, 1852, 1867–1875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- So, W.Y.; Leung, P.S. Irisin ameliorates hepatic glucose/lipid metabolism and enhances cell survival in insulin-resistant human HepG2 cells through adenosine monophosphate-activated protein kinase signaling. Int. J. Biochem. Cell. Biol. 2016, 78, 237–247. [Google Scholar] [CrossRef]
- Albrecht, E.; Schering, L.; Buck, F.; Vlach, K.; Schober, H.C.; Drevon, C.A.; Maak, S. Irisin: Still chasing shadows. Mol. Metab. 2020, 34, 124–135. [Google Scholar]
- Zybek-Kocik, A.; Sawicka-Gutaj, N.; Wrotkowska, E.; Sowiński, J.; Ruchała, M. Time-dependent irisin concentration changes in patients affected by overt hypothyroidism. Endokrynol. Pol. 2016, 67, 476–480. [Google Scholar]
- Huh, J.Y.; Panagiotou, G.; Mougios, V.; Brinkoetter, M.; Vamvini, M.T.; Schneider, B.E.; Mantzoros, C.S. FNDC5 and irisin in humans: I. predictors of circulating concentrations in serum and plasma and II. mRNA expression and circulating concentrations in response to weight loss and exercise. Metabolism 2012, 61, 1725–1738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dua, A.; Hennes, M.I.; Hoffmann, R.G.; Maas, D.L.; Krakower, G.R.; Sonnenberg, G.E.; Kissebah, A.H. Leptin: A significant indicator of total body fat but not of visceral fat and insulin insensitivity in African-American women. Diabetes 1996, 45, 1635–1637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruchala, M.; Zybek, A.; Szczepanek-Parulska, E. Serum irisin levels and thyroid function–newly discovered association. Peptides 2014, 60, 51–55. [Google Scholar] [CrossRef]
- Ateş, İ.; Altay, M.; Topçuoğlu, C.; Yılmaz, F.M. Circulating levels of irisin is elevated in hypothyroidism, a case-control study. Arch. Endocrinol. Metab. 2016, 60, 95–100. [Google Scholar] [CrossRef] [Green Version]
- Samy, D.M.; Ismail, C.A.; Nassra, R.A. Circulating irisin concentrations in rat models of thyroid dysfunction-effect of exercise. Metabolism 2015, 64, 804–813. [Google Scholar] [CrossRef] [PubMed]
- Keleş, E.; Turan, F.F. Evaluation of cord blood irisin levels in term newborns with small gestational age and appropriate gestational age. Springerplus 2016, 5, 1757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colaianni, G.; Cuscito, C.; Mongelli, T.; Oranger, A.; Mori, G.; Brunetti, G.; Colucci, S.; Cinti, S.; Grano, M. Irisin enhances osteoblast differentiation in vitro. Int. J. Endocrinol. 2014, 2014, 902186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Zhang, Y.; Zhao, F.; Yin, C.; Yang, C.; Wang, X.; Wu, Z.; Liang, S.; Li, D.; Lin, X.; et al. Recombinant irisin prevents the reduction of osteoblast differentiation induced by stimulated microgravity through increasing β-catenin expression. Int. J. Mol. Sci. 2020, 21, 1259. [Google Scholar] [CrossRef] [Green Version]
- Qiao, X.; Nie, Y.; Ma, Y.; Chen, Y.; Cheng, R.; Yin, W.; Hu, Y.; Xu, W.; Xu, L. Irisin promotes osteoblast proliferation and differentiation via activating the MAP kinase signaling pathways. Sci. Rep. 2016, 6, 18732. [Google Scholar] [CrossRef] [Green Version]
- Karras, S.N.; Koufakis, T.; Adamidou, L.; Dimakopoulos, G.; Karalazou, P.; Thisiadou, K.; Makedou, K.; Kotsa, K. Effects of christian orthodox fasting versus time-restricted eating on plasma irisin concentrations among overweight metabolically healthy individuals. Nutrients 2021, 13, 1071. [Google Scholar] [CrossRef] [PubMed]
- Engin-Üstün, Y.; Çağlayan, E.K.; Göçmen, A.Y.; Polat, M.F. Postmenopausal osteoporosis is associated with serum chemerin and irisin but not with apolipoprotein M levels. J. Menopausal Med. 2016, 22, 76–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gouveia, M.C.; Vella, J.P.; Cafeo, F.R.; Affonso Fonseca, F.L.; Bacci, M.R. Association between irisin and major chronic diseases: A review. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 4072–4077. [Google Scholar]
- Varela-Rodríguez, B.M.; Pena-Bello, L.; Juiz-Valiña, P.; Vidal-Bretal, B.; Cordido, F.; Sangiao-Alvarellos, S. FNDC5 expression and circulating irisin levels are modified by diet and hormonal conditions in hypothalamus, adipose tissue and muscle. Sci. Rep. 2016, 6, 29898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirsch, H.J.; Gross, I.; Pollak, Y.; Eldar-Geva, T.; Gross-Tsur, V. Irisin and the metabolic phenotype of adults with prader-willi syndrome. PLoS ONE 2015, 10, e0136864. [Google Scholar]
- Bakhshalizadeh, S.; Rabiee, F.; Shirazi, R.; Ghaedi, K.; Amidi, F.; Nasr-Esfahani, M.H. Assessment of PGC1α-FNDC5 axis in granulosa cells of PCOS mouse model. J. Reprod. Infertil. 2018, 19, 89–94. [Google Scholar]
- Adamska, A.; Karczewska-Kupczewska, M.; Lebkowska, A.; Milewski, R.; Gorska, M.; Otziomek, E. Serum irisin and its regulation by hyperinsulinemia in women with polycystic ovary syndrome. Endocrine. J. 2017, 63, 1107–1112. [Google Scholar]
- Polak, K.; Czyzyk, A.; Simoncini, T.; Meczekalski, B. New markers of insulin resistance in polycystic ovary syndrome. J. Endocrinol. Investig. 2017, 40, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acet, M.; Celik, N.; Acet, T.; Ilhan, S.; Yardim, M.; Aktun, H.L.; Basaranoglu, S.; Deregozu, A.; Aydin, S. Serum and follicular fluid irisin levels in poor and high responder women undergoing IVF/ICSI. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 1940–1946. [Google Scholar] [PubMed]
- Bostancı, M.S.; Akdemir, N.; Cinemre, B.; Cevrioglu, A.S.; Özden, S.; Ünal, O. Serum irisin levels in patients with polycystic ovary syndrome. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 4462–4468. [Google Scholar]
- Bonfante, I.L.; Chacon-Mikahil, M.P.T.; Brunelli, D.T.; Gáspari, A.F.; Duft, R.G.; Oliveira, A.G.; Araujo, T.G.; Saad, M.J.A.; Cavaglieri, C.R. Obese with higher FNDC5/irisin levels have a better metabolic profile, lower lipopolysaccharide levels and type 2 diabetes risk. Arch. Endocrinol. Metab. 2017, 61, 524–533. [Google Scholar]
- Elizondo-Montemayor, L.; Silva-Platas, C.; Torres-Quintanilla, A.; Rodríguez-López, C.; Ruiz-Esparza, G.U.; Reyes-Mendoza, E.; Garcia-Rivas, G. Association of irisin plasma levels with anthropometric parameters in children with underweight, normal weight, overweight, and obesity. Biomed. Res. Int. 2017, 2017, 2628968. [Google Scholar]
- Fukushima, Y.; Kurose, S.; Shinno, H.; Thi Thu, H.C.; Takao, N.; Tsutsumi, H.; Hasegawa, T.; Nakajima, T.; Kimura, Y. Effects of body weight reduction on serum irisin and metabolic parameters in obese subjects. Diabetes Metab. J. 2016, 40, 386–395. [Google Scholar] [CrossRef]
- Novelle, M.G.; Contreras, C.; Romero-Picó, A.; López, M.; Diéguez, C. Irisin, two years later. Int. J. Endocrinol. 2013, 2013, 746281. [Google Scholar]
- De Oliveira, M.; De Sibio, M.T.; Mathias, L.S.; Rodrigues, B.M.; Sakalem, M.E.; Nogueira, C.R. Irisin modulates genes associated with severe coronavirus disease (COVID-19) outcome in human subcutaneous adipocytes cell culture. Mol. Cell Endocrinol. 2020, 515, 110917. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Rampersad, S.; Wang, X.; Cheng, X.; Qu, S. Serum irisin concentrations were increased after transient continuous subcutaneous insulin infusion in type 2 diabetes mellitus patients. Diabetes Res. Clin. Pract. 2016, 113, 44–47. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, K.C.D.C.; Pereira, R.M.; de Campos, T.D.P.; De Moura, R.F.; Da Silva, A.S.R.; Cintra, D.E.; Ropelle, E.R.; Pauli, J.R.; de Araújo, M.B.; Moura, L.P. The role of physical exercise to improve the browning of white adipose tissue via POMC neurons. Front. Cell Neurosci. 2018, 12, 88. [Google Scholar] [CrossRef]
- Li, D.J.; Huang, F.; Lu, W.J.; Jiang, G.J.; Deng, Y.P.; Shen, F.M. Metformin promotes irisin release from murine skeletal muscle independently of AMP-activated protein kinase activation. Acta Physiol. 2015, 213, 711–721. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Chen, X.; Chen, Y.; Zhao, Q. PGC-1 mediates the regulation of metformin in muscle irisin expression and function. Am. J. Transl. Res. 2015, 7, 1850–1859. [Google Scholar]
- Rachid, T.L.; Penna-de-Carvalho, A.; Bringhenti, I.; Aguila, M.B.; Mandarim-de Lacerda, C.A.; Souza-Mello, V. Fenofibrate, (PPARalpha agonist) induces beige cell formation in subcutaneous white adipose tissue from diet-induced male obese mice. Mol. Cell. Endocrinol. 2015, 402, 86–94. [Google Scholar] [CrossRef]
- Feng, X.; Gao, X.; Jia, Y.; Zhang, H.; Pan, Q.; Yao, Z.; Yang, N.; Liu, J.; Xu, Y.; Wang, G.; et al. PPAR-α agonist fenofibrate decreased serum irisin levels in type 2 diabetes patients with hypertriglyceridemia. PPAR Res. 2015, 2015, 924131. [Google Scholar]
- Gouni-Berthold, I.; Berthold, H.K.; Huh, J.Y.; Berman, R.; Spenrath, N.; Krone, W.; Mantzoros, C.S. Effects of lipid-lowering drugs on irisin in human subjects in vivo and in human skeletal muscle cells ex vivo. PLoS ONE 2013, 8, e72858. [Google Scholar]
- Liu, J.; Hu, Y.; Zhang, H.; Xu, Y.; Wang, G. Exenatide treatment increases serum irisin levels in patients with obesity and newly diagnosed type 2 diabetes. J. Diabetes Complicat. 2016, 30, 1555–1559. [Google Scholar]
- Zhou, Q.; Chen, K.; Liu, P.; Gao, Y.; Zou, D.; Deng, H.; Huang, Y.; Zhang, Q.; Zhu, J.; Mi, M. Dihydromyricetin stimulates irisin secretion partially via the PGC-1α pathway. Mol. Cell. Endocrinol. 2015, 412, 349–357. [Google Scholar]
- Jiménez-Aranda, A.; Fernández-Vázquez, G.; Campos, D.; Tassi, M.; Velasco-Perez, L.; Tan, D.X.; Reiter, R.J.; Agil, A. Melatonin induces browning of inguinal white adipose tissue in Zucker diabetic fatty rats. J. Pineal Res. 2013, 55, 416–423. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Endocrine and Metabolic Disorders | Irisin Profile | References |
|---|---|---|
| THYROID DYSFUNCTIONS | Variation according to the thyroid function (hypothyroidism or hyperthyroidism) Correlation with TSH and fT4 | A. Zybek-Kocik et al., 2016 [26] J. Y. Huh et al., 2012 [27] M. Ruchala et al., 2014 [29] |
| SGA | Decreased in SGA Positive correlation with gestational age and birth weight | E. Keles et al., 2016 [32] |
| PRADER–WILLI SD. | Increased in saliva Compensatory role for the amelioration of lipid profile | H. J. Hirsh et al., 2015 [40] |
| OSTEOPOROSIS | Positive correlation with BMD and bone strength | B. Grygiel-Goriak et al., 2017 [2] G. Colaianni et al., 2014 [33] X. Qiao et al., 2016 [35] |
| OBESITY | Negative/positive (?) correlation with adiposity Positive correlation with BMI in children | B. Grygiel-Goriak et al., 2017 [2] L. Elizondo-Montemayor et al., 2017 [47] |
| PCOS | Increased in serum Irisin resistance (?) | A. Adamska et al., 2017 [42] K. Polak et al., 2017 [43] M. Acet et al., 2016 [44] |
| DIABETES MELLITUS | Decreased in T2DM Increased in obese T2DM | A. Shoukry et al., 2016 [8] K. S. Rana et al., 2017 [10] L. Zhao et al., 2015 [10] |
| Therapeutical Agent | Study Type | Action | References |
|---|---|---|---|
| INSULIN | humans with T2DM | ↑ serum irisin | L. Li et al., 2016 [51] |
| LEPTIN | obese mice | Up-regulation of FNDC5 in skeletal muscle → ↑ irisin ↓ irisin expression in subcutaneous fat | A. Rodriguez et al., 2015 [52] |
| METFORMIN | diabetic obese mice | Up-regulation of FNDC5 in skeletal muscle → ↑ irisin | D. J. Li et al., 2015 [53] |
| METFORMIN | obese mice | Up-regulation of FNDC5 in skeletal muscle → ↑ irisin | Z. Yang et al., 2015 [54] |
| FENOFIBRATE | male C57BL/6 mice | irisin induction | T. L. Rachid et al., 2015 [55] |
| FENOFIBRATE | humans with T2DM | ↓ irisin in serum | X. Feng et al., 2015 [56] |
| SIMVASTATIN | humans with hypercholesterolemia | ↑ irisin | I. Gouni-Berthold et al., 2013 [57] |
| EXENATIDE | humans with T2DM | ↑ irisin | J. Liu et al., 2016 [58] |
| DIHYDROMYRICETIN | humans rats | ↑ irisin | Q. Zhou et al., 2015 [59] |
| MELATONIN | Diabetic fatty rats | ↑ irisin | A. Jimenez-Aranda et al., 2013 [60] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leustean, L.; Preda, C.; Teodoriu, L.; Mihalache, L.; Arhire, L.; Ungureanu, M.-C. Role of Irisin in Endocrine and Metabolic Disorders—Possible New Therapeutic Agent? Appl. Sci. 2021, 11, 5579. https://0-doi-org.brum.beds.ac.uk/10.3390/app11125579
Leustean L, Preda C, Teodoriu L, Mihalache L, Arhire L, Ungureanu M-C. Role of Irisin in Endocrine and Metabolic Disorders—Possible New Therapeutic Agent? Applied Sciences. 2021; 11(12):5579. https://0-doi-org.brum.beds.ac.uk/10.3390/app11125579
Chicago/Turabian StyleLeustean, Letitia, Cristina Preda, Laura Teodoriu, Laura Mihalache, Lidia Arhire, and Maria-Christina Ungureanu. 2021. "Role of Irisin in Endocrine and Metabolic Disorders—Possible New Therapeutic Agent?" Applied Sciences 11, no. 12: 5579. https://0-doi-org.brum.beds.ac.uk/10.3390/app11125579