
Informative Biomarkers for Autism Spectrum Disorder Diagnosis in Functional Magnetic Resonance Imaging Data on the Default Mode Network

 , , and
, , and
Abstract
:1. Introduction
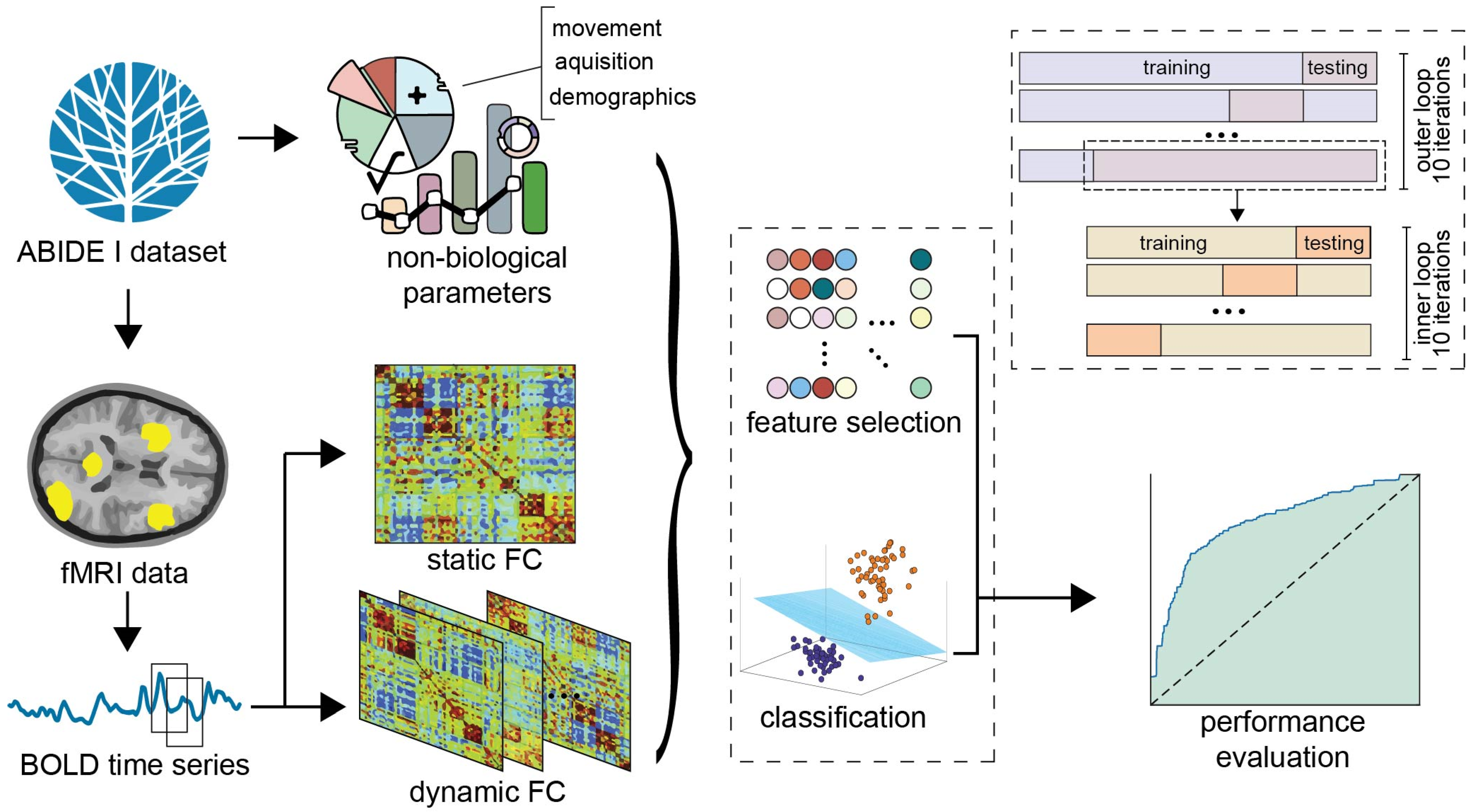
2. Materials and Methods
2.1. fMRI Data Acquisition
2.2. Inclusion/Exclusion Criteria and Pre-Processing
2.3. Feature Extraction
2.3.1. Static Functional Connectivity
2.3.2. Dynamic Functional Connectivity
2.3.3. Demographics
2.3.4. Acquisition Parameters
2.3.5. Motion Parameters
2.4. Feature Selection and Biomarker Assessment
2.5. Classification
3. Results
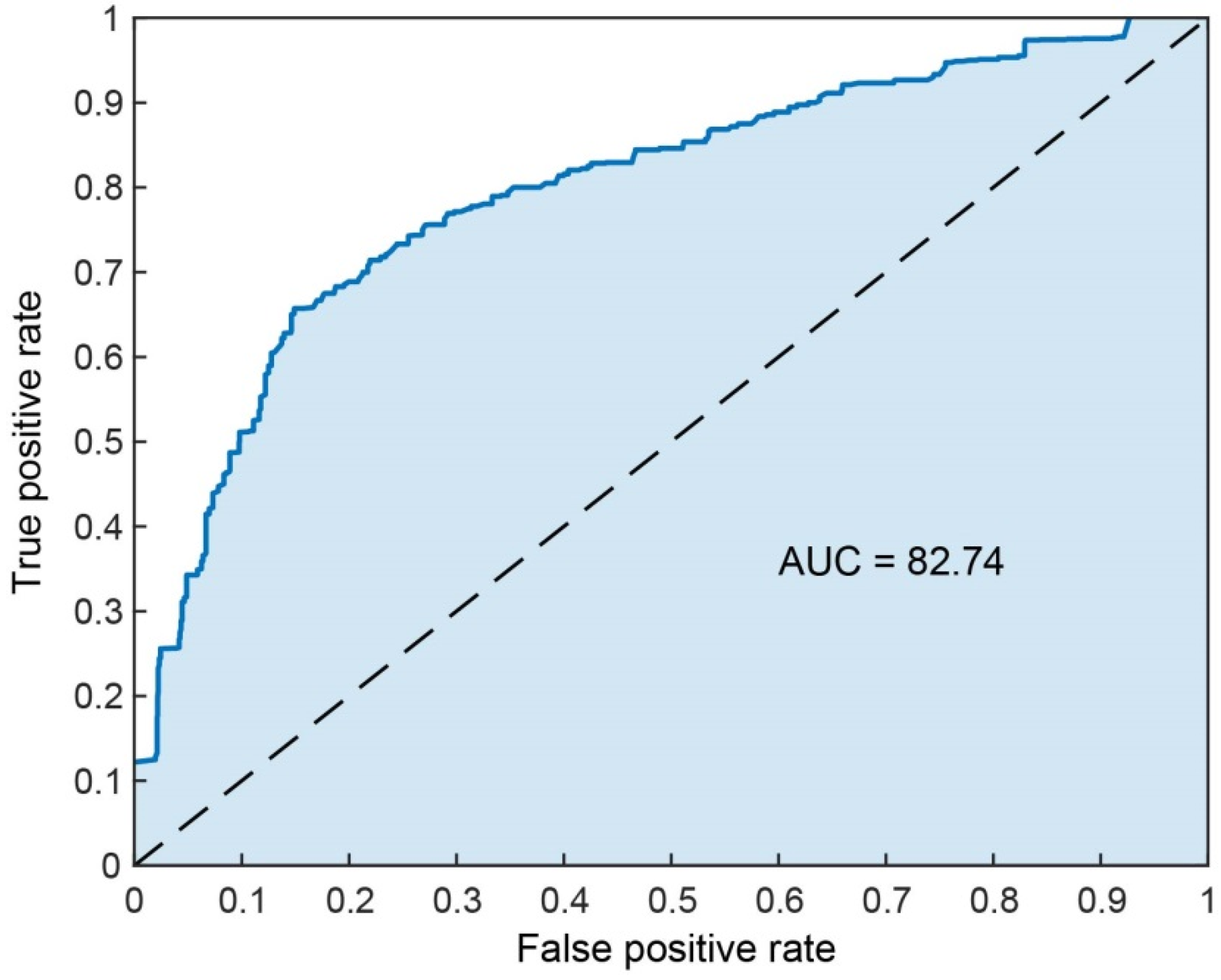
3.1. Classification Performance Results
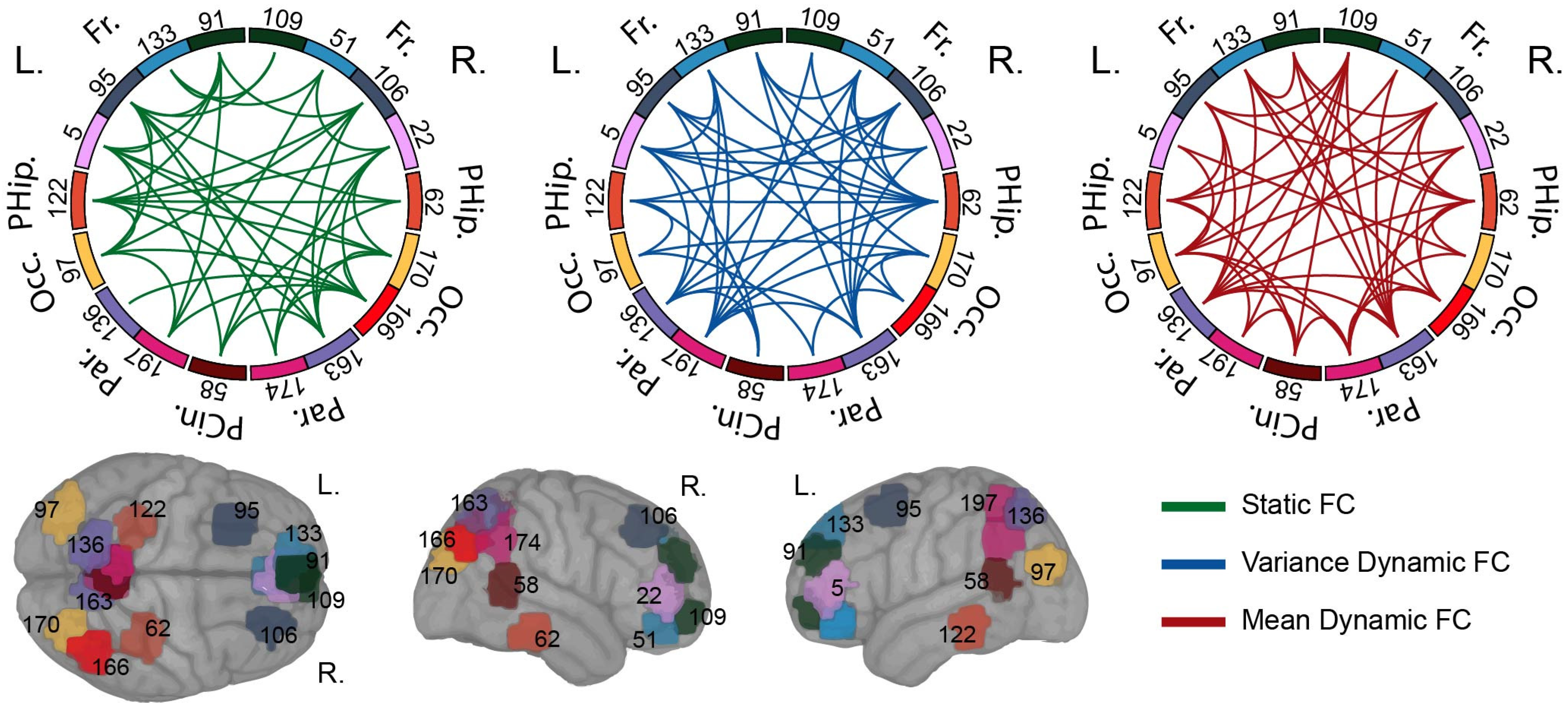
3.2. Analysis of Selected Biomarkers
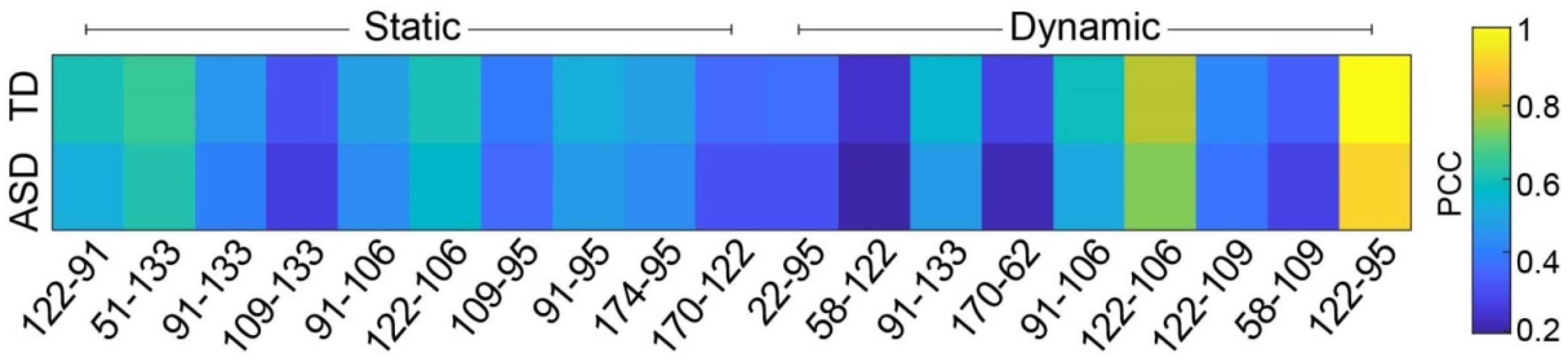
3.3. Functional Connectivity Features Validation
3.4. Evaluation of the Adopted Framework and Relationship to Prior Work
4. Discussion
4.1. Classification Performance
4.2. Informative Biomarkers
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Association: Arlington, VA, USA, 2013. [Google Scholar]
- Kasari, C. Are we there yet? The state of early prediction and intervention in autism spectrum disorder. J. Am. Acad. Child Adolesc. Psychiatry 2014, 53, 133–134. [Google Scholar] [CrossRef] [Green Version]
- Shen, M.D.; Piven, J. Brain and behavior development in autism from birth through infancy. Dialog-Clin. Neurosci. 2017, 19, 325–333. [Google Scholar]
- Ha, S.; Sohn, I.-J.; Kim, N.; Sim, H.J.; Cheon, K.-A. Characteristics of Brains in Autism Spectrum Disorder: Structure, Function and Connectivity across the Lifespan. Exp. Neurobiol. 2015, 24, 273–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Im, W.Y.; Ha, J.H.; Kim, E.J.; Cheon, K.-A.; Cho, J.; Song, D.-H. Impaired White Matter Integrity and Social Cognition in High-Function Autism: Diffusion Tensor Imaging Study. Psychiatry Investig. 2018, 15, 292–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lange, N.; Travers, B.G.; Bigler, E.D.; Prigge, M.B.; Froehlich, A.L.; Nielsen, J.A.; Cariello, A.N.; Zielinski, B.A.; Anderson, J.S.; Fletcher, P.T.; et al. Longitudinal Volumetric Brain Changes in Autism Spectrum Disorder Ages 6-35 Years. Autism Res. 2015, 8, 82–93. [Google Scholar] [CrossRef]
- Friston, K.J. Functional and Effective Connectivity: A Review. Brain Connect. 2011, 1, 13–36. [Google Scholar] [CrossRef]
- Abbott, A.E.; Nair, A.; Keown, C.L.; Datko, M.; Jahedi, A.; Fishman, I.; Müller, R.-A. Patterns of Atypical Functional Connectivity and Behavioral Links in Autism Differ Between Default, Salience, and Executive Networks. Cereb. Cortex 2016, 26, 4034–4045. [Google Scholar] [CrossRef]
- Rolls, E.T.; Zhou, Y.; Cheng, W.; Gilson, M.; Deco, G.; Feng, J. Effective connectivity in autism. Autism Res. 2019, 13, 32–44. [Google Scholar] [CrossRef] [Green Version]
- Jung, M.; Kosaka, H.; Saito, D.N.; Ishitobi, M.; Morita, T.; Inohara, K.; Asano, M.; Arai, S.; Munesue, T.; Tomoda, A.; et al. Default mode network in young male adults with autism spectrum disorder: Relationship with autism spectrum traits. Mol. Autism 2014, 5, 35. [Google Scholar] [CrossRef] [Green Version]
- Hyatt, C.J.; Calhoun, V.D.; Pittman, B.; Corbera, S.; Bell, M.D.; Rabany, L.; Pelphrey, K.; Pearlson, G.D.; Assaf, M. Default mode network modulation by mentalizing in young adults with autism spectrum disorder or schizophrenia. NeuroImage Clin. 2020, 27, 102343. [Google Scholar] [CrossRef]
- Nair, A.; Jolliffe, M.; Lograsso, Y.S.S.; Bearden, C.E. A Review of Default Mode Network Connectivity and Its Association with Social Cognition in Adolescents with Autism Spectrum Disorder and Early-Onset Psychosis. Front. Psychiatry 2020, 11, 614. [Google Scholar] [CrossRef]
- Neufeld, J.; Kuja-Halkola, R.; Mevel, K.; Cauvet, É.; Fransson, P.; Bölte, S. Alterations in resting state connectivity along the autism trait continuum: A twin study. Mol. Psychiatry 2017, 23, 1659–1665. [Google Scholar] [CrossRef]
- Janardhanan, P.; Heena, L.; Sabika, F. Effectiveness of Support Vector Machines in Medical Data mining. J. Commun. Softw. Syst. 2015, 11, 25–30. [Google Scholar] [CrossRef]
- Shahid, N.; Rappon, T.; Berta, W. Applications of artificial neural networks in health care organizational decision-making: A scoping review. PLoS ONE 2019, 14, e0212356. [Google Scholar] [CrossRef] [PubMed]
- Di Martino, A.; Yan, C.-G.; Li, Q.; Denio, E.; Castellanos, F.; Alaerts, K.; Anderson, J.S.; Assaf, M.; Bookheimer, S.Y.; Dapretto, M.; et al. The autism brain imaging data exchange: Towards a large-scale evaluation of the intrinsic brain architecture in autism. Mol. Psychiatry 2014, 19, 659–667. [Google Scholar] [CrossRef] [PubMed]
- Plitt, M.; Barnes, K.A.; Martin, A. Functional connectivity classification of autism identifies highly predictive brain features but falls short of biomarker standards. NeuroImage Clin. 2015, 7, 359–366. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.P.; Keown, C.L.; Jahedi, A.; Nair, A.; Pflieger, M.E.; Bailey, B.A.; Müller, R.-A. Diagnostic classification of intrinsic functional connectivity highlights somatosensory, default mode, and visual regions in autism. NeuroImage Clin. 2015, 8, 238–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Xiao, Z.; Wu, J. Functional connectivity-based classification of autism and control using SVM-RFECV on rs-fMRI data. Phys. Med. 2019, 65, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Aghdam, M.A.; Sharifi, A.; Pedram, M.M. Diagnosis of Autism Spectrum Disorders in Young Children Based on Resting-State Functional Magnetic Resonance Imaging Data Using Convolutional Neural Networks. J. Digit. Imaging 2019, 32, 899–918. [Google Scholar] [CrossRef]
- Sherkatghanad, Z.; Akhondzadeh, M.; Salari, S.; Zomorodi-Moghadam, M.; Abdar, M.; Acharya, U.R.; Khosrowabadi, R.; Salari, V. Automated Detection of Autism Spectrum Disorder Using a Convolutional Neural Network. Front. Neurosci. 2020, 13, 1325. [Google Scholar] [CrossRef] [Green Version]
- Abraham, A.; Milham, M.P.; Di Martino, A.; Craddock, R.C.; Samaras, D.; Thirion, B.; Varoquaux, G. Deriving reproducible biomarkers from multi-site resting-state data: An Autism-based example. NeuroImage 2017, 147, 736–745. [Google Scholar] [CrossRef] [Green Version]
- Parisot, S.; Ktena, S.I.; Ferrante, E.; Lee, M.; Guerrero, R.; Glocker, B.; Rueckert, D. Disease prediction using graph convolutional networks: Application to Autism Spectrum Disorder and Alzheimer’s disease. Med. Image Anal. 2018, 48, 117–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinsfeld, A.S.; Franco, A.R.; Craddock, R.C.; Buchweitz, A.; Meneguzzi, F. Identification of autism spectrum disorder using deep learning and the ABIDE dataset. NeuroImage Clin. 2018, 17, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Epalle, T.M.; Song, Y.; Liu, Z.; Lu, H. Multi-atlas classification of autism spectrum disorder with hinge loss trained deep architectures: ABIDE I results. Appl. Soft Comput. 2021, 107, 107375. [Google Scholar] [CrossRef]
- Karampasi, A.; Kakkos, I.; Miloulis, S.-T.; Zorzos, I.; Dimitrakopoulos, G.N.; Gkiatis, K.; Asvestas, P.; Matsopoulos, G. A Machine Learning fMRI Approach in the Diagnosis of Autism. In Proceedings of the 2020 IEEE International Conference on Big Data (Big Data), Atlanta, GA, USA, 10–13 December 2020; Institute of Electrical and Electronics Engineers (IEEE): Piscataway, NJ, USA, 2020; pp. 3628–3631. [Google Scholar]
- Cameron, C.; Sharad, S.; Brian, C.; Ranjeet, K.; Satrajit, G.; Chaogan, Y.; Qingyang, L.; Daniel, L.; Joshua, V.; Randal, B.; et al. Towards Automated Analysis of Connectomes: The Configurable Pipeline for the Analysis of Connectomes (C-PAC). Front. Aging Neurosci. 2013, 42, 3310–3389. [Google Scholar] [CrossRef]
- Power, J.D.; Barnes, K.A.; Snyder, A.Z.; Schlaggar, B.L.; Petersen, S.E. Spurious but systematic correlations in functional connectivity MRI networks arise from subject motion. NeuroImage 2012, 59, 2142–2154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ABIDE Preprocessed. Available online: http://preprocessed-connectomes-project.org/abide/cpac.html (accessed on 3 July 2021).
- Smith, S.M.; Fox, P.; Miller, K.; Glahn, D.C.; Mackay, C.; Filippini, N.; Watkins, K.E.; Toro, R.; Laird, A.R.; Beckmann, C.F. Correspondence of the brain’s functional architecture during activation and rest. Proc. Natl. Acad. Sci. USA 2009, 106, 13040–13045. [Google Scholar] [CrossRef] [Green Version]
- Shirer, W.R.; Ryali, S.; Rykhlevskaia, E.; Menon, V.; Greicius, M.D. Decoding Subject-Driven Cognitive States with Whole-Brain Connectivity Patterns. Cereb. Cortex 2011, 22, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Harlalka, V.; Bapi, R.S.; Vinod, P.K.; Roy, D. Atypical Flexibility in Dynamic Functional Connectivity Quantifies the Severity in Autism Spectrum Disorder. Front. Hum. Neurosci. 2019, 13, 6. [Google Scholar] [CrossRef] [Green Version]
- Preti, M.G.; Bolton, T.W.A.; Van De Ville, D. The dynamic functional connectome: State-of-the-art and perspectives. NeuroImage 2017, 160, 41–54. [Google Scholar] [CrossRef]
- Savva, A.D.; Karampasi, A.S.; Matsopoulos, G.K. Chapter 6-Deriving Resting-State fMRI Biomarkers for Classification of Autism Spectrum Disorder. In Autism 360°; Das, U., Papaneophytou, N., El-Kour, T., Eds.; Academic Press: Amsterdam, The Netherlands, 2020; pp. 101–123. ISBN 978-0-12-818466-0. [Google Scholar]
- Ambroise, C.; McLachlan, G.J. Selection bias in gene extraction on the basis of microarray gene-expression data. Proc. Natl. Acad. Sci. USA 2002, 99, 6562–6566. [Google Scholar] [CrossRef] [Green Version]
- Yan, K.; Zhang, D. Feature selection and analysis on correlated gas sensor data with recursive feature elimination. Sens. Actuators B Chem. 2015, 212, 353–363. [Google Scholar] [CrossRef]
- Kakkos, I.; Dimitrakopoulos, G.N.; Gao, L.; Zhang, Y.; Qi, P.; Matsopoulos, G.K.; Thakor, N.; Bezerianos, A.; Sun, Y. Mental Workload Drives Different Reorganizations of Functional Cortical Connectivity Between 2D and 3D Simulated Flight Experiments. IEEE Trans. Neural Syst. Rehabil. Eng. 2019, 27, 1704–1713. [Google Scholar] [CrossRef] [PubMed]
- Burges, C.J. A Tutorial on Support Vector Machines for Pattern Recognition. Data Min. Knowl. Discov. 1998, 2, 121–167. [Google Scholar] [CrossRef]
- Ariafar, S.; Coll-Font, J.; Brooks, D.; Dy, J. ADMMBO: Bayesian Optimization with Unknown Constraints using ADMM. J. Mach. Learn. Res. 2019, 20, 20. [Google Scholar]
- Eslami, T.; Mirjalili, V.; Fong, A.; Laird, A.R.; Saeed, F. ASD-DiagNet: A Hybrid Learning Approach for Detection of Autism Spectrum Disorder Using fMRI Data. Front. Aging Neurosci. 2019, 13, 70. [Google Scholar] [CrossRef] [PubMed]
- Kunda, M.; Zhou, S.; Gong, G.; Lu, H. Improving Multi-Site Autism Classification Based on Site-Dependence Minimisation and Second-Order Functional Connectivity. bioRxiv 2020, 2, 930073. [Google Scholar] [CrossRef] [Green Version]
- Niu, K.; Guo, J.; Pan, Y.; Gao, X.; Peng, X.; Li, N.; Li, H. Multichannel Deep Attention Neural Networks for the Classification of Autism Spectrum Disorder Using Neuroimaging and Personal Characteristic Data. Complexity 2020, 2020, 1–9. [Google Scholar] [CrossRef]
- Liu, Y.; Xu, L.; Li, J.; Yu, J.Y.A.X.; Yu, X. Attentional Connectivity-based Prediction of Autism Using Heterogeneous rs-fMRI Data from CC200 Atlas. Exp. Neurobiol. 2020, 29, 27–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Islam, M.S.; Khaled, A.M.A. Functional connectivity magnetic resonance imaging classification of autism spectrum disorder using the multisite ABIDE dataset. In Proceedings of the 2019 IEEE EMBS International Conference on Biomedical & Health Informatics (BHI), Hong Kong, China, 5–7 January 2012; Institute of Electrical and Electronics Engineers (IEEE): Piscataway, NJ, USA, 2019; pp. 1–4. [Google Scholar]
- Wang, C.; Xiao, Z.; Wang, B.; Wu, J. Identification of Autism Based on SVM-RFE and Stacked Sparse Auto-Encoder. IEEE Access 2019, 7, 118030–118036. [Google Scholar] [CrossRef]
- Min, S.; Lee, B.; Yoon, S. Deep learning in bioinformatics. Brief. Bioinform. 2016, 18, bbw068–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazeminejad, A.; Sotero, R.C. Topological Properties of Resting-State fMRI Functional Networks Improve Machine Learning-Based Autism Classification. Front. Neurosci. 2019, 12, 1018. [Google Scholar] [CrossRef] [PubMed]
- Kiviniemi, V.; Vire, T.; Remes, J.; Elseoud, A.A.; Starck, T.; Tervonen, O.; Nikkinen, J. A Sliding Time-Window ICA Reveals Spatial Variability of the Default Mode Network in Time. Brain Connect. 2011, 1, 339–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Washington, S.D.; Gordon, E.; Brar, J.; Warburton, S.; Sawyer, A.T.; Wolfe, A.; Mease-Ference, E.R.; Girton, L.; Hailu, A.; Mbwana, J.; et al. Dysmaturation of the default mode network in autism. Hum. Brain Mapp. 2014, 35, 1284–1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yerys, B.E.; Gordon, E.; Abrams, D.N.; Satterthwaite, T.; Weinblatt, R.; Jankowski, K.F.; Strang, J.; Kenworthy, L.; Gaillard, W.D.; Vaidya, C.J. Default mode network segregation and social deficits in autism spectrum disorder: Evidence from non-medicated children. NeuroImage Clin. 2015, 9, 223–232. [Google Scholar] [CrossRef] [Green Version]
- Garrison, J.R.; Fernyhough, C.; McCarthy-Jones, S.; Haggard, M.; Simons, J.S. Paracingulate Sulcus Morphology Is Associated with Hallucinations in the Human Brain. Nat. Commun. 2015, 6, 8956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeuner, K.E.; Sidiropoulos, C. Cognitive behavioral therapy in functional tremor. Neurology 2019, 93, 825–826. [Google Scholar] [CrossRef]
- Warrier, V.; Toro, R.; Won, H.; Leblond, C.S.; Cliquet, F.; Delorme, R.; De Witte, W.; Bralten, J.; Chakrabarti, B.; Børglum, A.D.; et al. Social and non-social autism symptoms and trait domains are genetically dissociable. Commun. Biol. 2019, 2, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Gaigg, S.B.; Gardiner, J.M.; Bowler, D. Free recall in autism spectrum disorder: The role of relational and item-specific encoding. Neuropsychology 2008, 46, 983–992. [Google Scholar] [CrossRef] [Green Version]
- Cooper, R.; Richter, F.R.; Bays, P.M.; Plaisted-Grant, K.C.; Baron-Cohen, S.; Simons, J.S. Reduced Hippocampal Functional Connectivity During Episodic Memory Retrieval in Autism. Cereb. Cortex 2017, 27, 888–902. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, H.; Taki, Y.; Nouchi, R.; Sekiguchi, A.; Kotozaki, Y.; Miyauchi, C.M.; Yokoyama, R.; Iizuka, K.; Hashizume, H.; Nakagawa, S.; et al. Regional gray matter density is associated with achievement motivation: Evidence from voxel-based morphometry. Brain Struct. Funct. 2014, 219, 71–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borsook, D.; Maleki, N.; Burstein, R. Chapter 42-Migraine. In Neurobiology of Brain Disorders; Zigmond, M.J., Rowland, L.P., Coyle, J.T., Eds.; Academic Press: San Diego, CA, USA, 2015; pp. 693–708. ISBN 978-0-12-398270-4. [Google Scholar]
- Zhang, B.; He, S.; Weng, X. Localization and Functional Characterization of an Occipital Visual Word form Sensitive Area. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef]
- Zhen, Z.; Fang, H.; Liu, J. The Hierarchical Brain Network for Face Recognition. PLoS ONE 2013, 8, e59886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weigelt, S.; Koldewyn, K.; Kanwisher, N. Face Recognition Deficits in Autism Spectrum Disorders Are Both Domain Specific and Process Specific. PLoS ONE 2013, 8, e74541. [Google Scholar] [CrossRef] [Green Version]
- Cerliani, L.; Mennes, M.; Thomas, R.M.; Di Martino, A.; Thioux, M.; Keysers, C. Increased Functional Connectivity between Subcortical and Cortical Resting-State Networks in Autism Spectrum Disorder. JAMA Psychiatry 2015, 72, 767–777. [Google Scholar] [CrossRef]
- Chen, H.; Nomi, J.S.; Uddin, L.Q.; Duan, X.; Chen, H. Intrinsic functional connectivity variance and state-specific under-connectivity in autism. Hum. Brain Mapp. 2017, 38, 5740–5755. [Google Scholar] [CrossRef] [Green Version]
- Graña, M.; Silva, M. Impact of Machine Learning Pipeline Choices in Autism Prediction from Functional Connectivity Data. Int. J. Neural Syst. 2021, 31, 2150009. [Google Scholar] [CrossRef]
- Cheng, W.; Rolls, E.; Gu, H.; Zhang, J.; Feng, J. Autism: Reduced connectivity between cortical areas involved in face expression, theory of mind, and the sense of self. Brain 2015, 138, 1382–1393. [Google Scholar] [CrossRef] [Green Version]
- Ghuman, A.S.; Honert, R.N.V.D.; Huppert, T.J.; Wallace, G.; Martin, A. Aberrant Oscillatory Synchrony Is Biased Toward Specific Frequencies and Processing Domains in the Autistic Brain. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2017, 2, 245–252. [Google Scholar] [CrossRef] [Green Version]
- Aykan, S.; Puglia, M.H.; Kalaycıoğlu, C.; Pelphrey, K.A.; Tuncalı, T.; Nalçacı, E. Right Anterior Theta Hypersynchrony as a Quantitative Measure Associated with Autistic Traits and K-Cl Cotransporter KCC2 Polymorphism. J. Autism Dev. Disord. 2021, 1–12. [Google Scholar] [CrossRef]
- Arbab, T.; Battaglia, F.P.; Pennartz, C.M.; Bosman, C.A. Abnormal hippocampal theta and gamma hypersynchrony produces network and spike timing disturbances in the Fmr1-KO mouse model of Fragile X syndrome. Neurobiol. Dis. 2018, 114, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Kissinger, S.T.; Wu, Q.; Quinn, C.J.; Anderson, A.K.; Pak, A.; Chubykin, A.A. Visual Experience-Dependent Oscillations and Underlying Circuit Connectivity Changes Are Impaired in Fmr1 KO Mice. Cell Rep. 2020, 31, 107486. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ASD | TD | |
|---|---|---|
| Handedness | 334 R/54 L/11 A | 432 R/32 L/8 A |
| Age | 16.4197 F/17.7835 M | 17.3202 F/14.5929 M |
| Sex | 49 F/350 M | 93 F/379 M |
| Reference | Classifier | Data | #Total Features | #Selected Features | Accuracy (%) |
|---|---|---|---|---|---|
| Proposed Work | RFE-CBR/SVM | 472 TD/399 ASD | 473 | 136 | 76.63 |
| Karampasi et al. (2020) [26] | RFE/SVM | 472 TD/399 ASD | 561 | 117 | 72.5 |
| Heinsfeld et al. (2017) [24] | SVM | 530 TD 505 ASD | 19,900 | 600 | 65 |
| Eslami et al. (2019) [40] | SVM | 530 TD/505 ASD | 19,900 | 9950 | 68.3 |
| Kunda et al. (2020) [41] | SVM | 530 TD/505 ASD | - | - | 71 |
| Niu et al. (2020) [42] | SVM | 401 TD/408 ASD | 19,905 | 19,902 | 69.3 |
| Liu et al. (2020) [43] | Extra-Trees/SVM | 548 TD/506 ASD | 19,900 | 1935 | 72.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karampasi, A.S.; Savva, A.D.; Korfiatis, V.C.; Kakkos, I.; Matsopoulos, G.K. Informative Biomarkers for Autism Spectrum Disorder Diagnosis in Functional Magnetic Resonance Imaging Data on the Default Mode Network. Appl. Sci. 2021, 11, 6216. https://0-doi-org.brum.beds.ac.uk/10.3390/app11136216
Karampasi AS, Savva AD, Korfiatis VC, Kakkos I, Matsopoulos GK. Informative Biomarkers for Autism Spectrum Disorder Diagnosis in Functional Magnetic Resonance Imaging Data on the Default Mode Network. Applied Sciences. 2021; 11(13):6216. https://0-doi-org.brum.beds.ac.uk/10.3390/app11136216
Chicago/Turabian StyleKarampasi, Aikaterini S., Antonis D. Savva, Vasileios Ch. Korfiatis, Ioannis Kakkos, and George K. Matsopoulos. 2021. "Informative Biomarkers for Autism Spectrum Disorder Diagnosis in Functional Magnetic Resonance Imaging Data on the Default Mode Network" Applied Sciences 11, no. 13: 6216. https://0-doi-org.brum.beds.ac.uk/10.3390/app11136216