
Quantitative Optical Coherence Tomography for Longitudinal Monitoring of Postnatal Retinal Development in Developing Mouse Eyes


Abstract
:Featured Application
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Preparation
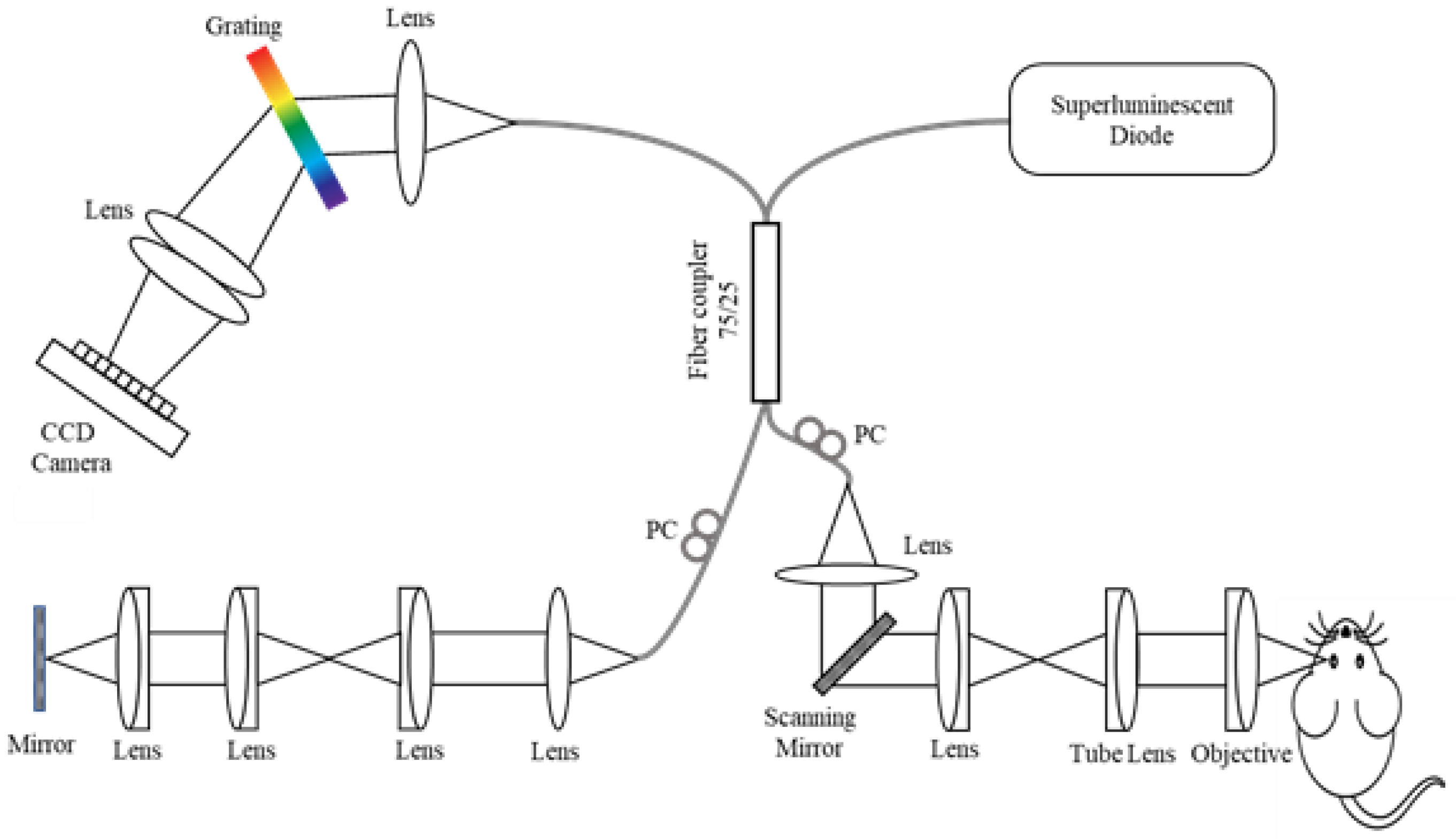
2.2. Imaging System Setup
2.3. Data Acquisition and Image Processing
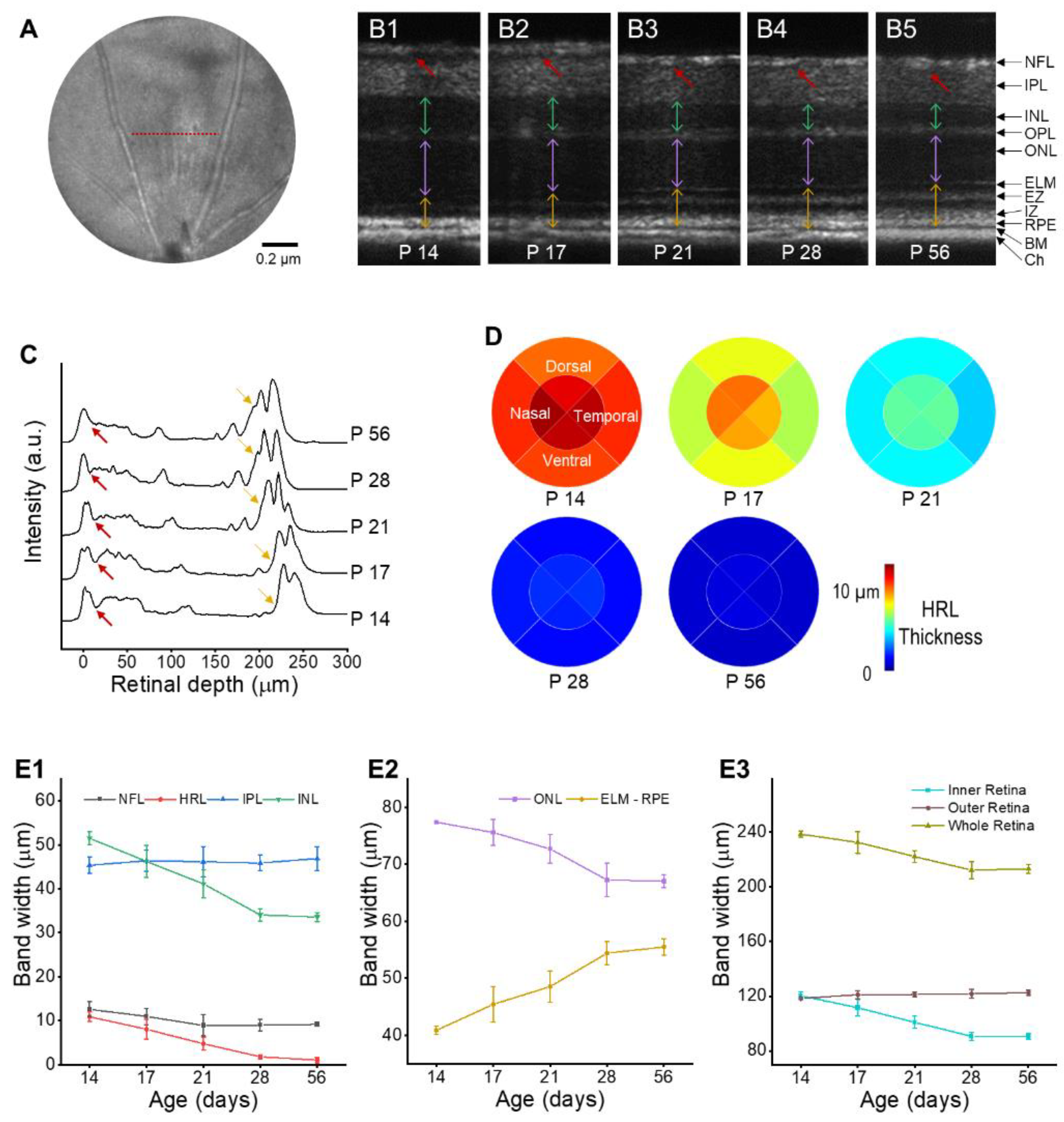
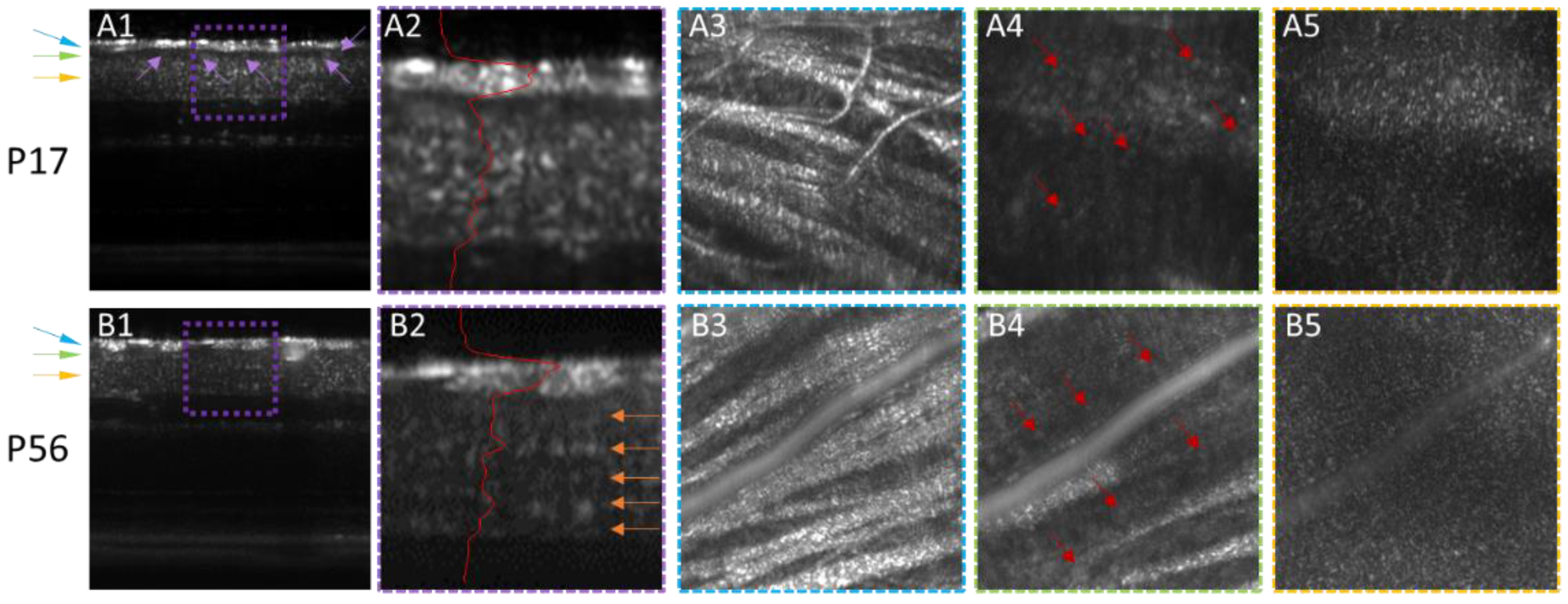
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Morgan, J.; Wong, R. Development of cell types and synaptic connections in the retina. In Webvision: The Organization of the Retina Visual System; University of Utah Health Sciences Center: Salt Lake City, UT, USA, 2007. [Google Scholar]
- Chan, Y.-C.; Chiao, C.-C. Effect of visual experience on the maturation of ON–OFF direction selective ganglion cells in the rabbit retina. Vis. Res. 2008, 48, 2466–2475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, N.; Copenhagen, D.R. Visual stimulation is required for refinement of ON and OFF pathways in postnatal retina. Neuron 2003, 39, 85–96. [Google Scholar] [CrossRef] [Green Version]
- Tian, N. Development of retinal ganglion cell dendritic structure and synaptic connections. In Webvision: The Organization of the Retina Visual System; University of Utah Health Sciences Center: Salt Lake City, UT, USA, 2012. [Google Scholar]
- Coombs, J.L.; Van Der List, D.; Chalupa, L.M. Morphological properties of mouse retinal ganglion cells during postnatal development. J. Comp. Neurol. 2007, 503, 803–814. [Google Scholar] [CrossRef] [PubMed]
- Tian, N. Visual experience and maturation of retinal synaptic pathways. Vis. Res. 2004, 44, 3307–3316. [Google Scholar] [CrossRef] [Green Version]
- Fisher, L.J. Development of synaptic arrays in the inner plexiform layer of neonatal mouse retina. J. Comp. Neurol. 1979, 187, 359–372. [Google Scholar] [CrossRef]
- Koehler, C.L.; Akimov, N.P.; Rentería, R.C. Receptive field center size decreases and firing properties mature in ON and OFF retinal ganglion cells after eye opening in the mouse. J. Neurophysiol. 2011, 106, 895–904. [Google Scholar] [CrossRef] [Green Version]
- Drexler, W.; Fujimoto, J.G. Optical Coherence Tomography: Technology and Applications; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2008. [Google Scholar]
- Hariri, S.; Moayed, A.A.; Choh, V.; Bizheva, K. In vivo assessment of thickness and reflectivity in a rat outer retinal degeneration model with ultrahigh resolution optical coherence tomography. Investig. Ophthalmol. Vis. Sci. 2012, 53, 1982–1989. [Google Scholar] [CrossRef]
- Nivison-Smith, L.; Wang, H.; Assaad, N.; Kalloniatis, M. Retinal Thickness Changes throughout the Natural History of Drusen in Age-related Macular Degeneration. Optom. Vis. Sci. 2018, 95, 648–655. [Google Scholar] [CrossRef]
- Kim, T.-H.; Son, T.; Klatt, D.; Yao, X. Concurrent OCT and OCT angiography of retinal neurovascular degeneration in the 5XFAD Alzheimer’s disease mice. Neurophotonics 2021, 8, 035002. [Google Scholar] [CrossRef]
- Chauhan, B.C.; Stevens, K.T.; Levesque, J.M.; Nuschke, A.C.; Sharpe, G.P.; O’Leary, N.; Archibald, M.L.; Wang, X. Longitudinal in vivo imaging of retinal ganglion cells and retinal thickness changes following optic nerve injury in mice. PLoS ONE 2012, 7, e40352. [Google Scholar]
- Zeimer, R.; Asrani, S.; Zou, S.; Quigley, H.; Jampel, H. Quantitative detection of glaucomatous damage at the posterior pole by retinal thickness mapping: A pilot study. Ophthalmology 1998, 105, 224–231. [Google Scholar] [CrossRef]
- Maldonado, R.S.; O’Connell, R.V.; Sarin, N.; Freedman, S.F.; Wallace, D.K.; Cotten, C.M.; Winter, K.P.; Stinnett, S.; Chiu, S.J.; Izatt, J.A. Dynamics of human foveal development after premature birth. Ophthalmology 2011, 118, 2315–2325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, T.H.; Son, T.; Yao, X. Feature article: Functional OCT angiography reveals early physiological dysfunction of hyaloid vasculature in developing mouse eye. Exp. Biol. Med. 2019, 244, 819–823. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.H.; Son, T.; Le, D.; Yao, X. Longitudinal OCT and OCTA monitoring reveals accelerated regression of hyaloid vessels in retinal degeneration 10 (rd10) mice. Sci. Rep. 2019, 9, 16685. [Google Scholar] [CrossRef]
- Kim, T.H.; Son, T.; Lu, Y.; Alam, M.; Yao, X. Comparative Optical Coherence Tomography Angiography of Wild-Type and rd10 Mouse Retinas. Transl. Vis. Sci. Technol. 2018, 7, 42. [Google Scholar] [CrossRef] [Green Version]
- Pi, S.; Hormel, T.T.; Wei, X.; Cepurna, W.; Morrison, J.C.; Jia, Y. Imaging retinal structures at cellular-level resolution by visible-light optical coherence tomography. Optics Lett. 2020, 45, 2107–2110. [Google Scholar] [CrossRef]
- Liu, Z.; Kurokawa, K.; Zhang, F.; Lee, J.J.; Miller, D.T. Imaging and quantifying ganglion cells and other transparent neurons in the living human retina. Proc. Natl. Acad. Sci. USA 2017, 114, 12803–12808. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Miller, E.B.; Manna, S.K.; Meleppat, R.K.; Pugh, E.N.; Zawadzki, R. Temporal speckle-averaging of optical coherence tomography volumes for in-vivo cellular resolution neuronal and vascular retinal imaging. Neurophotonics 2019, 6, 041105. [Google Scholar] [CrossRef]
- Ma, G.; Son, T.; Kim, T.H.; Yao, X. In vivo optoretinography of phototransduction activation and energy metabolism in retinal photoreceptors. J. Biophotonics 2021, 14, e202000462. [Google Scholar] [CrossRef]
- Son, T.; Alam, M.; Toslak, D.; Wang, B.; Lu, Y.; Yao, X. Functional optical coherence tomography of neurovascular coupling interactions in the retina. J. Biophotonics 2018, 11, e201800089. [Google Scholar] [CrossRef]
- Tkatchenko, T.V.; Shen, Y.; Tkatchenko, A.V. Analysis of postnatal eye development in the mouse with high-resolution small animal magnetic resonance imaging. Investig. Ophthalmol. Vis. Sci. 2010, 51, 21–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, R.W. Cell death during differentiation of the retina in the mouse. J. Comp. Neurol. 1984, 229, 362–373. [Google Scholar] [CrossRef] [PubMed]
- Caley, D.W.; Johnson, C.; Liebelt, R.A. The postnatal development of the retina in the normal and rodless CBA mouse: A light and electron microscopic study. Am. J. Anat. 1972, 133, 179–211. [Google Scholar] [CrossRef] [PubMed]
- Farah, M.H.; Easter, S.S., Jr. Cell birth and death in the mouse retinal ganglion cell layer. J. Comp. Neurol. 2005, 489, 120–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dangata, Y.; Findlater, G.; Kaufman, M. Postnatal development of the optic nerve in (C57BL × CBA) F1 hybrid mice: General changes in morphometric parameters. J. Anat. 1996, 189, 117. [Google Scholar] [PubMed]
- Gyllensten, L.; Malmfors, T.R. Myelinization of the optic nerve and its dependence on visual function—A quantitative investigation in mice. Embryol. Exp. Morphol. 1963, 255–266. [Google Scholar] [CrossRef]
- Gyllensten, L.; Malmfors, T.; Norrlin-Grettve, M.L. Developmental and functional alterations in the fiber composition of the optic nerve in visually deprived mice. J. Comp. Neurol. 1966, 128, 413–418. [Google Scholar] [CrossRef]
- Antony, B.J.; Jeong, W.; Abramoff, M.D.; Vance, J.; Sohn, E.H.; Garvin, M.K. Automated 3D segmentation of intraretinal surfaces in SD-OCT volumes in normal and diabetic mice. Transl. Vis. Sci. Technol. 2014, 3, 8. [Google Scholar] [CrossRef] [Green Version]
- Ruggeri, M.; Wehbe, H.; Jiao, S.; Gregori, G.; Jockovich, M.E.; Hackam, A.; Duan, Y.; Puliafito, C.A. In vivo three-dimensional high-resolution imaging of rodent retina with spectral-domain optical coherence tomography. Investig. Ophthalmol. Vis. Sci. 2007, 48, 1808–1814. [Google Scholar] [CrossRef]
- Zhang, P.; Zam, A.; Jian, Y.; Wang, X.; Li, Y.; Lam, K.S.; Burns, M.E.; Sarunic, M.V.; Pugh, E.N., Jr.; Zawadzki, R.J. In vivo wide-field multispectral scanning laser ophthalmoscopy–optical coherence tomography mouse retinal imager: Longitudinal imaging of ganglion cells, microglia, and Müller glia, and mapping of the mouse retinal and choroidal vasculature. J. Biomed. Opt. 2015, 20, 126005. [Google Scholar] [CrossRef] [Green Version]
- Harper, D.J.; Augustin, M.; Lichtenegger, A.; Gesperger, J.; Himmel, T.; Muck, M.; Merkle, C.W.; Eugui, P.; Kummer, S.; Woehrer, A. Retinal analysis of a mouse model of Alzheimer’s disease with multicontrast optical coherence tomography. Neurophotonics 2020, 7, 015006. [Google Scholar] [CrossRef] [PubMed]
- Dysli, C.; Enzmann, V.; Sznitman, R.; Zinkernagel, M.S. Quantitative analysis of mouse retinal layers using automated segmentation of spectral domain optical coherence tomography images. Transl. Vis. Sci. Technol. 2015, 4, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srinivasan, P.P.; Heflin, S.J.; Izatt, J.A.; Arshavsky, V.Y.; Farsiu, S. Automatic segmentation of up to ten layer boundaries in SD-OCT images of the mouse retina with and without missing layers due to pathology. Biomed. Opt. Express 2014, 5, 348–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pennesi, M.E.; Michaels, K.V.; Magee, S.S.; Maricle, A.; Davin, S.P.; Garg, A.K.; Gale, M.J.; Tu, D.C.; Wen, Y.; Erker, L.R. Long-term characterization of retinal degeneration in rd1 and rd10 mice using spectral domain optical coherence tomography. Investig. Ophthalmol. Vis. Sci. 2012, 53, 4644–4656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanna, J.; Yücel, Y.H.; Zhou, X.; Mathieu, E.; Paczka-Giorgi, L.A.; Gupta, N. Progressive loss of retinal blood vessels in a live model of retinitis pigmentosa. Can. J. Ophthalmol. 2018, 53, 391–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinekar, A.; Mangalesh, S.; Jayadev, C.; Maldonado, R.S.; Bauer, N.; Toth, C.A. Retinal imaging of infants on spectral domain optical coherence tomography. BioMed Res. Int. 2015, 2015, 782420. [Google Scholar] [CrossRef] [Green Version]
- Vajzovic, L.; Hendrickson, A.E.; O’Connell, R.V.; Clark, L.A.; Tran-Viet, D.; Possin, D.; Chiu, S.J.; Farsiu, S.; Toth, C.A. Maturation of the human fovea: Correlation of spectral-domain optical coherence tomography findings with histology. Am. J. Ophthalmol. 2012, 154, 779–789. e772. [Google Scholar] [CrossRef] [Green Version]
- Vajzovic, L.; Rothman, A.L.; Tran-Viet, D.; Cabrera, M.T.; Freedman, S.F.; Toth, C.A. Delay in retinal photoreceptor development in very preterm compared to term infants. Investig. Ophthalmol. Vis. Sci. 2015, 56, 908–913. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Age | NFL (μm) | HRL (μm) | IPL (μm) | INL (μm) | ONL (μm) | ELM-RPE (μm) | Inner Retina (μm) | Outer Retina (μm) | Whole Retina (μm) |
|---|---|---|---|---|---|---|---|---|---|
| P14 | 12.6 ± 1.7 | 10.9 ± 1.1 | 45.3 ± 1.9 | 51.5 ± 1.5 | 77.4 ± 0.3 | 40.9 ± 0.7 | 120.3 ± 2.5 | 118.3 ± 0.9 | 238.5 ± 2.3 |
| P17 | 10.9 ± 1.8 | 8 ± 2.3 | 46.4 ± 2.4 | 46.2 ± 3.6 | 75.6 ± 2.2 | 45.4 ± 3.1 | 111.5 ± 6.2 | 121 ± 3 | 232.5 ± 8 |
| P21 | 8.9 ± 2.4 | 4.7 ± 1.5 | 46.1 ± 3.5 | 41.1 ± 3.2 | 72.7 ± 2.5 | 48.6 ± 2.7 | 100.9 ± 4.4 | 121.3 ± 1.9 | 222.1 ± 4.4 |
| P28 | 8.9 ± 1.3 | 1.7 ± 0.5 | 45.9 ± 1.8 | 34 ± 1.4 | 67.3 ± 2.9 | 54.4 ± 2 | 90.6 ± 3.1 | 121.7 ± 3.2 | 212.3 ± 6.2 |
| P56 | 9.1 ± 0.5 | 1 ± 0.6 | 46.9 ± 2.7 | 33.6 ± 1 | 67 ± 1.1 | 55.5 ± 1.4 | 90.5 ± 2.2 | 122.6 ± 1.9 | 213.1 ± 3.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, G.; Ding, J.; Kim, T.-H.; Yao, X. Quantitative Optical Coherence Tomography for Longitudinal Monitoring of Postnatal Retinal Development in Developing Mouse Eyes. Appl. Sci. 2022, 12, 1860. https://0-doi-org.brum.beds.ac.uk/10.3390/app12041860
Ma G, Ding J, Kim T-H, Yao X. Quantitative Optical Coherence Tomography for Longitudinal Monitoring of Postnatal Retinal Development in Developing Mouse Eyes. Applied Sciences. 2022; 12(4):1860. https://0-doi-org.brum.beds.ac.uk/10.3390/app12041860
Chicago/Turabian StyleMa, Guangying, Jie Ding, Tae-Hoon Kim, and Xincheng Yao. 2022. "Quantitative Optical Coherence Tomography for Longitudinal Monitoring of Postnatal Retinal Development in Developing Mouse Eyes" Applied Sciences 12, no. 4: 1860. https://0-doi-org.brum.beds.ac.uk/10.3390/app12041860