
Selection and Validation of Reliable Reference Genes for qRT-PCR Normalization of Bursaphelenchus xylophilus from Different Temperature Conditions and Developmental Stages

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Reproductive Juveniles Nematode
2.2. Collection of Dispersal Juveniles
2.3. Collection of Nematodes Treated with Different Temperatures
2.4. Total RNA Isolation and cDNA Synthesis
2.5. Selection of Candidate Reference Genes and qPCR Primer Design
2.6. Quantitative Reverse Transcription-Polymerase Chain Reaction
2.7. Assessing Expression Stability of Reference Genes
2.8. Validation of Reference Genes
3. Results
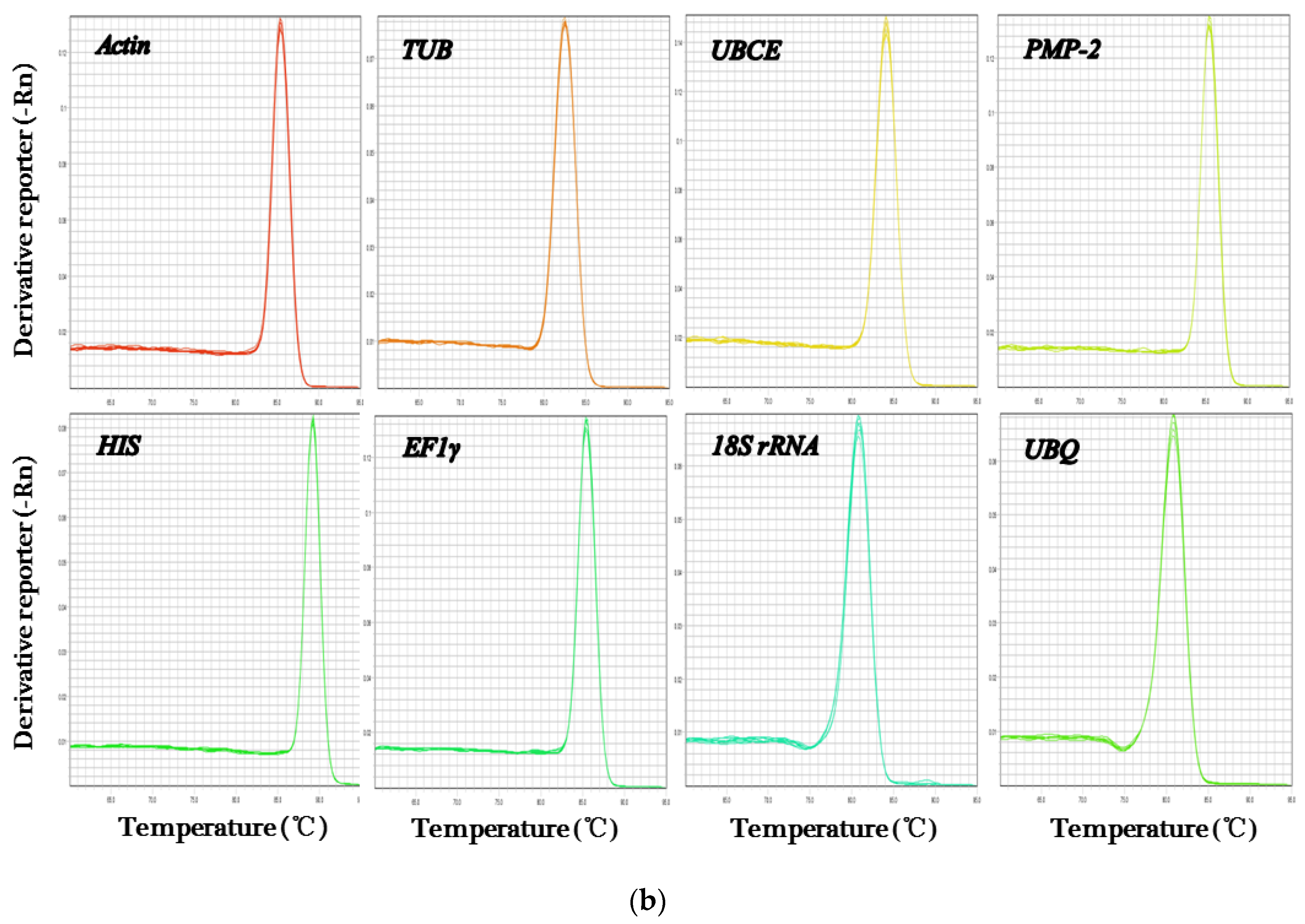
3.1. Performance of qRT-PCR Primers
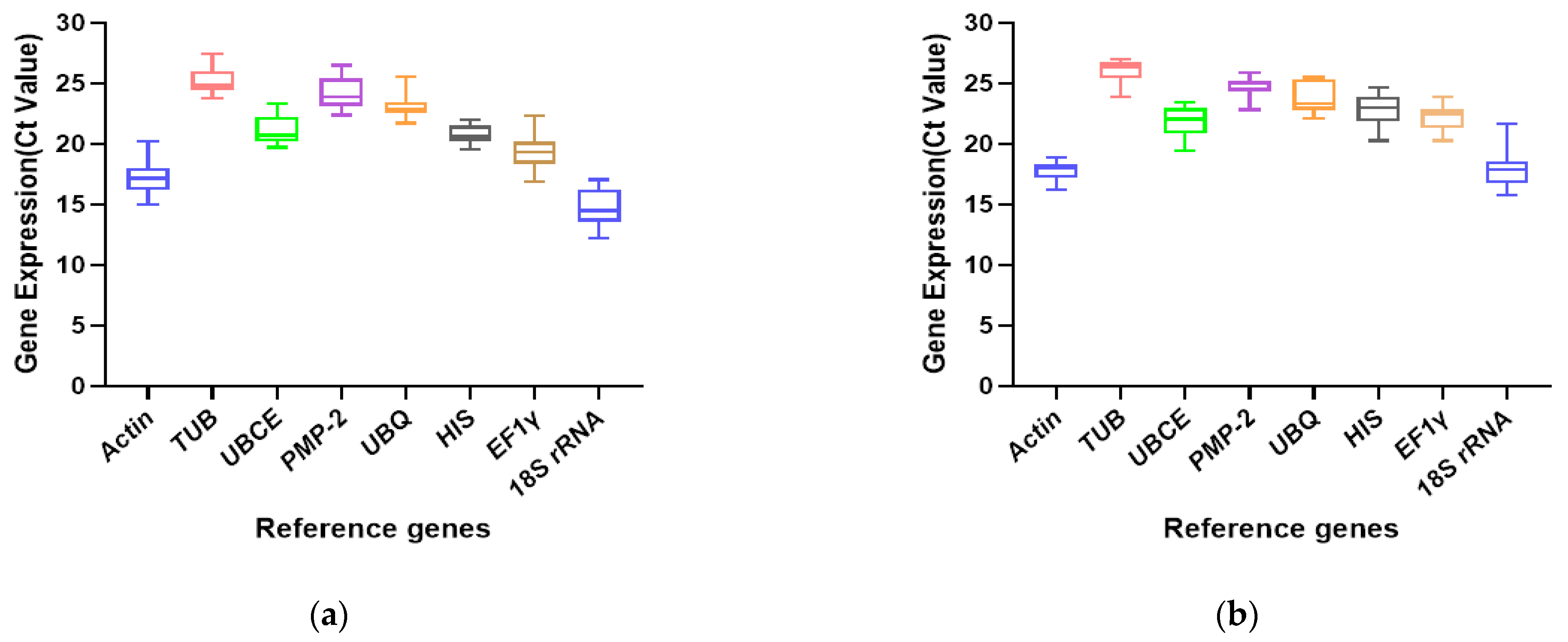
3.2. Expression Profiles of Candidate Reference Genes
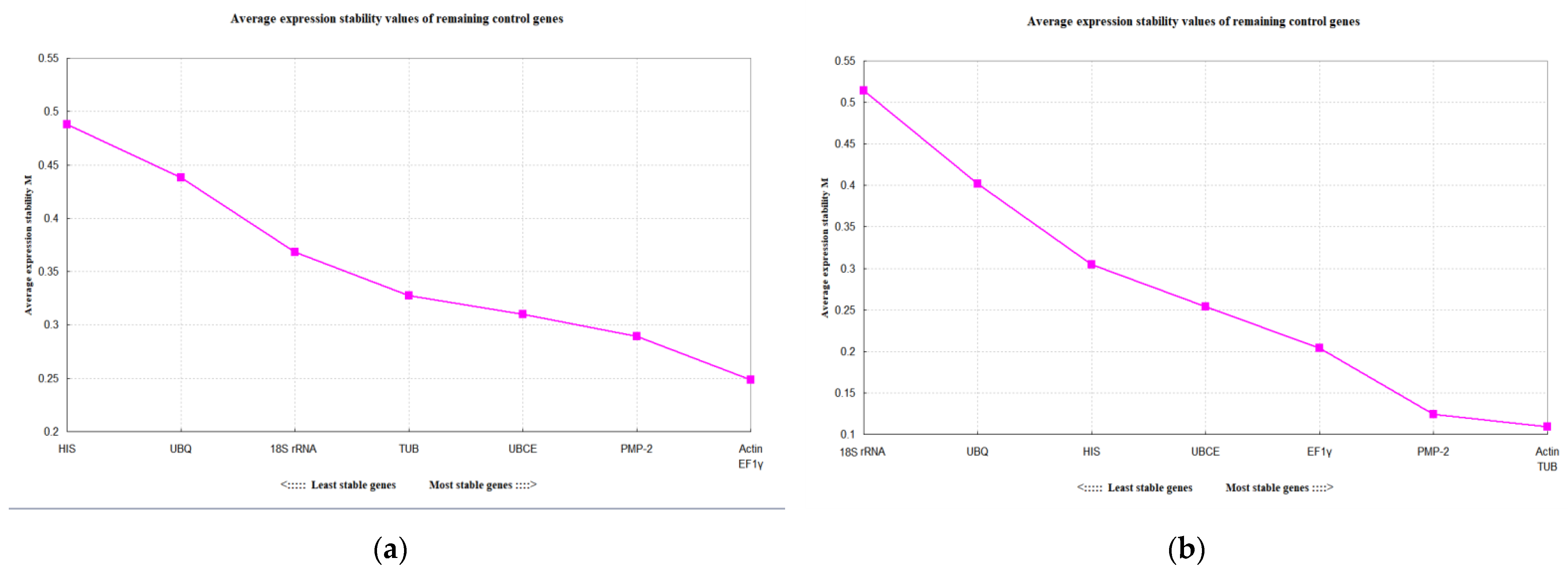
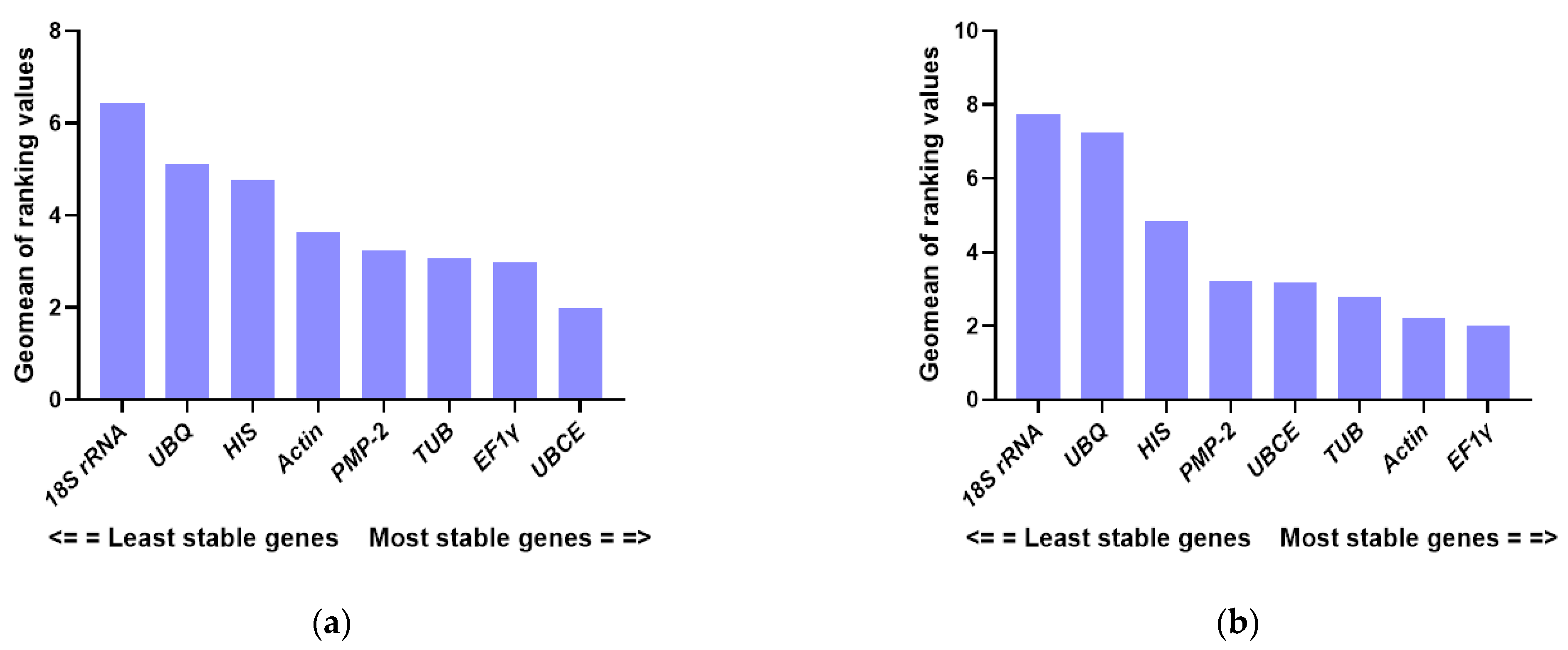
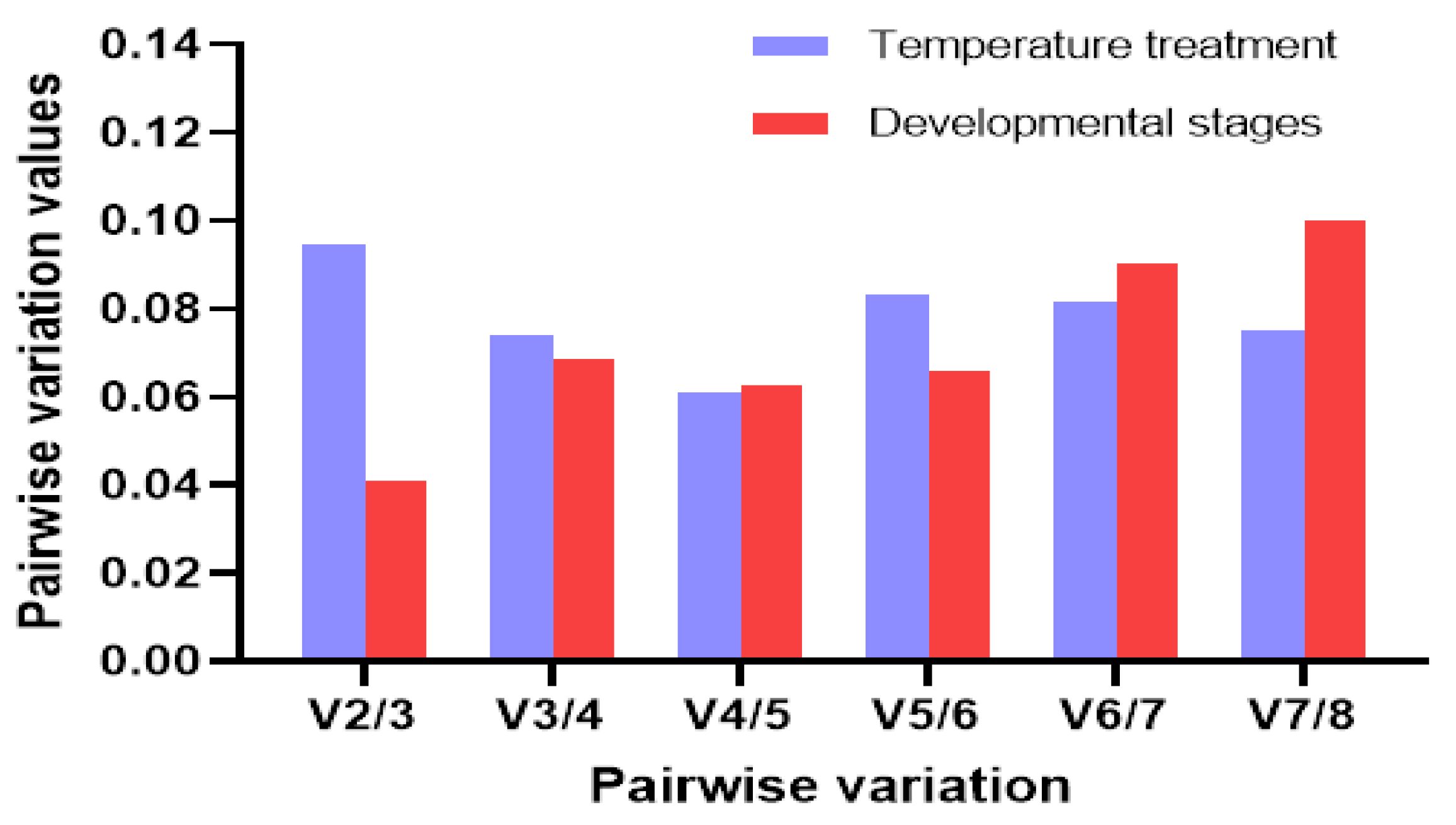
3.3. Stability of the Candidate Reference Genes
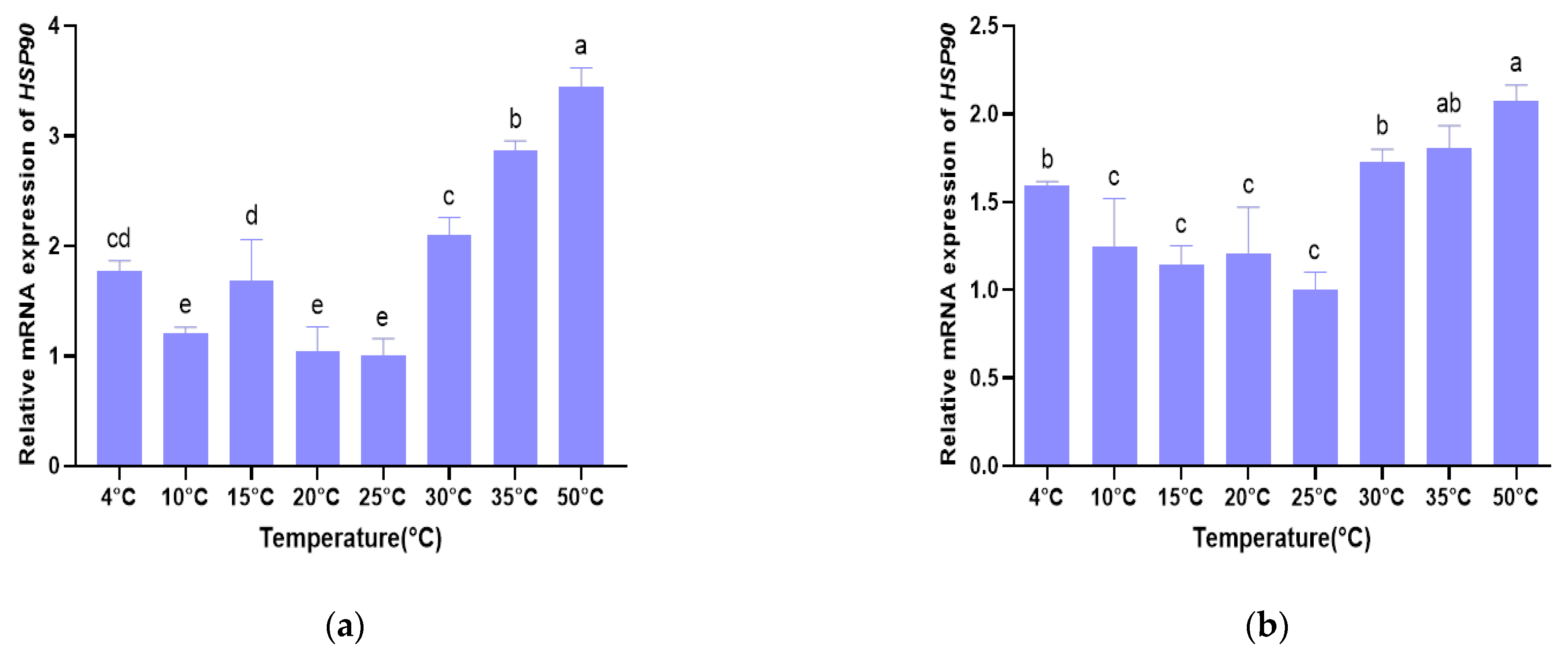
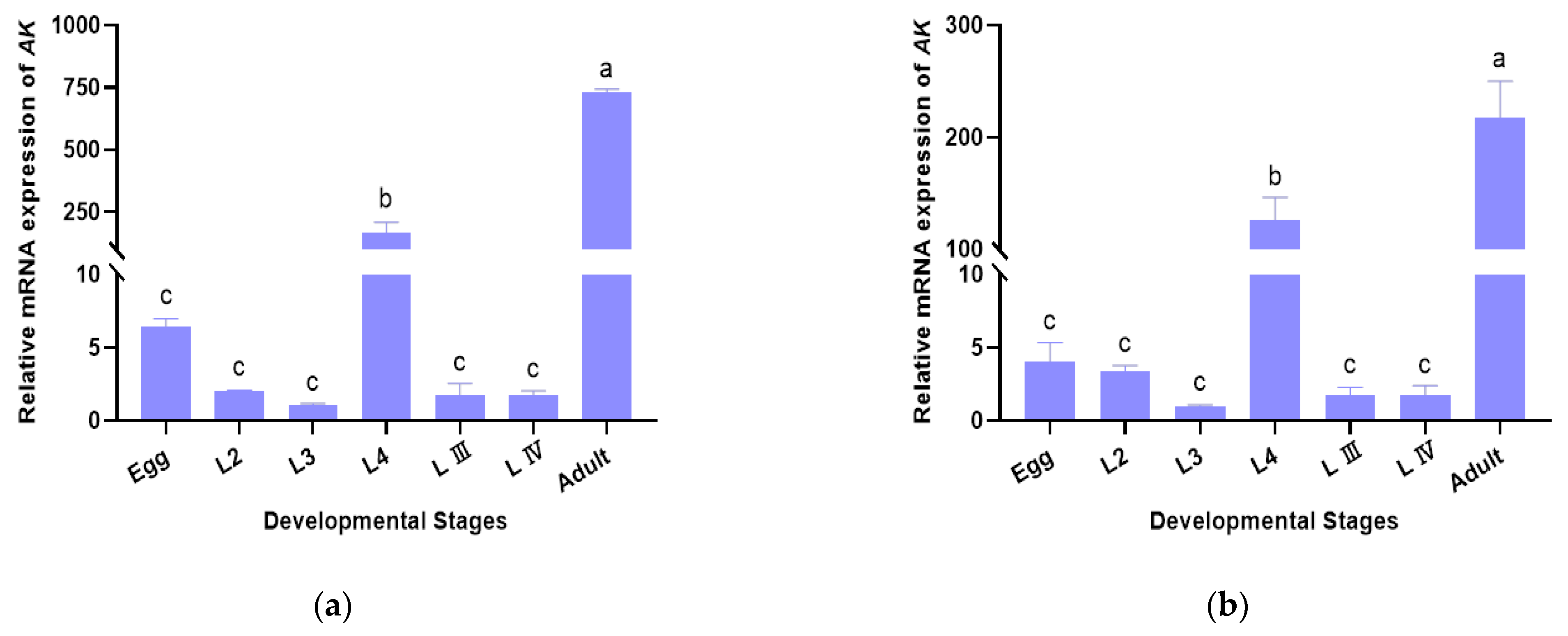
3.4. Validation of Selected Reference Genes in B. xylophilus
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Rodrigues, J.M. A Worldwide Threat to Forest Ecosystems. In Pine Wilt Disease; Mota, M.M., Vieira, P., Eds.; Springer: Dordrechet, The Netherlands, 2008; pp. 5–14. [Google Scholar]
- Zhao, L.; Mota, M.; Vieira, P.; Butcher, R.A.; Sun, J. Interspecific Communication between Inewood Nematode, Its Insect Vector, and Associated Microbes. Trends Parasitol. 2014, 30, 299–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Mota, M.; Vieira, P.; Butcher, R.A.; Sun, J. A Native Fungal Symbiont Facilitates the Prevalence and Development of an Invasive Pathogen-native Vector Symbiosis. Ecol. Ecol. 2013, 94, 2817–2826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maehara, N.; Futai, K. Effect of Fungal Interactions on the Numbers of the Pinewood Nematode, Bursaphelenchus xylophilus (Nematoda: Aphelenchoididae), Carried by the Japanese Pine Sawyer, Monochamus alternatus (Coleoptera: Cerambycidae). Fundam. Appl. Nematol. 1997, 20, 611–617. [Google Scholar]
- Zhao, B.G.; Tao, J.; Ju, Y.W.; Wang, P.K.; Ye, J.L. The Role of Wood-inhabiting Bacteria in Pine Wilt Disease. J. Nematol. 2011, 43, 129–134. [Google Scholar] [PubMed]
- Zhao, L.; Zhang, X.; Wei, Y.; Zhou, J.; Zhang, W.; Qin, P.; Chinta, S.; Kong, X.; Liu, Y.; Yu, H.; et al. Ascarosides Coordinate the Dispersal of a Plant-parasitic Nematode with the Metamorphosis of Its Vector Beetle. Nat. Commun. 2016, 7, 8. [Google Scholar] [CrossRef] [Green Version]
- Tomminen, J.; Halik, S.; Bergdahl, D.R. Incubation-Temperature and Time Effects on Life Stages of Bursaphelenchus Xylophilus in Wood Chips. J. Nematol. 1991, 23, 477–484. [Google Scholar]
- Cha, D.J.; Kim, J.; Kim, D.S. Nematicidal Activities of Three Naphthoquinones against the Pine Wood Nematode, Bursaphelenchus xylophilus. Molecules 2019, 24, 3634. [Google Scholar] [CrossRef] [Green Version]
- de la Fuente, B.; Saura, S.; Beck, P.S.A. Predicting the Spread of an Invasive Tree Pest: The Pine Wood Nematode in Southern Europe. J. Appl. Ecol. 2018, 55, 2374–2385. [Google Scholar] [CrossRef]
- Firmino, P.N.; Calvao, T.; Ayres, M.P.; Pimentel, C.S. Monochamus galloprovincialis and Bursaphelenchus xylophilus Life History in an Area Severely Affected by Pine Wilt Disease: Implications for Forest Management. For. Ecol. Manag. 2017, 389, 105–115. [Google Scholar] [CrossRef]
- Huang, X.; Hu, L.J.; Wu, X.Q. Identification of a Novel Effector BxSapB3 that Enhances the Virulence of Pine Wood Nematode Bursaphelenchus xylophilus. Acta Biochim. Biophys. Sin. 2019, 51, 1071–1078. [Google Scholar] [CrossRef]
- Qiu, X.; Yang, L.; Ye, J.; Wang, W.; Zhao, T.; Hu, H.; Zhou, G. Silencing of Cyp-33C9 Gene Affects the Reproduction and Pathogenicity of the Pine Wood Nematode, Bursaphelenchus xylophilus. Int. J. Mol. Sci. 2019, 20, 4520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Meng, F.; Deng, X.; Wang, X.; Feng, Y.; Zhang, W.; Pan, L.; Zhang, X. Comparative Transcriptome Analysis of the Pinewood Nematode Bursaphelenchus xylophilus Reveals the Molecular Mechanism Underlying Its Defense Response to Host-Derived -pinene. Int. J. Mol. Sci. 2019, 20, 911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espada, M.; Silva, A.C.; Akker, S.E.V.D.; Cock, P.; Mota, M.; Jones, J.T. Identification and Characterization of Parasitism Genes from the Pinewood Nematode Bursaphelenchus xylophilus Reveals a Multilayered Detoxification Strategy. Mol. Plant Pathol. 2016, 17, 286–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, L.X.; Wu, X.Q.; Xue, Q.; Qiu, X.W. Effects of Endobacterium (Stenotrophomonas maltophilia) on Pathogenesis-Related Gene Expression of Pine Wood Nematode (Bursaphelenchus xylophilus) and Pine Wilt Disease. Int. J. Mol. Sci. 2016, 17, 778. [Google Scholar] [CrossRef] [Green Version]
- Ma, H.B.; Lu, Q.; Liang, J.; Zhang, X.Y. Functional Analysis of the Cellulose Gene of the Pine Wood Nematode, Bursaphelenchus xylophilus, Using RNA Interference. Genet. Mol. Res. 2011, 10, 1931–1941. [Google Scholar] [CrossRef]
- Qiu, X.W.; Wu, X.Q.; Huang, L.; Ye, J.R. Influence of Bxpel1 Gene Silencing by dsRNA Interference on the Development and Pathogenicity of the Pine Wood Nematode, Bursaphelenchus xylophilus. Int. J. Mol. Sci. 2016, 17, 125. [Google Scholar] [CrossRef] [Green Version]
- Tang, J.; Ma, R.; Zhu, N.; Guo, K.; Guo, Y.; Bai, L.; Yu, H.; Hu, J.; Zhang, X. Bxy-fuca Encoding Alpha-L-fucosidase Plays Crucial Roles in Development and Reproduction of the Pathogenic Pinewood Nematode, Bursaphelenchus xylophilus. Pest Manag. Sci. 2020, 76, 205–214. [Google Scholar] [CrossRef]
- Bustin, S.A. Developments in Real-Time PCR Research and Molecular Diagnostics. Expert Rev. Mol. Diagn. 2010, 10, 713–715. [Google Scholar] [CrossRef]
- Jozefczuk, J.; Adjaye, J. Methods in Systems Biology. In Methods in Enzymology; Jameson, D., Verma, M., Westerhoff, H.V., Eds.; Academic Press: San Diego, CA, USA, 2011; Volume 500, pp. 99–109. [Google Scholar]
- Singh, C.; Roy-Chowdhuri, S. Methods in Molecular Biology. In Clinical Applications of PCR, 3rd ed.; Luthra, R., Singh, R.R., Patel, K.P., Eds.; Humana Press: Totowa, NJ, USA, 2016; Volume 1392, pp. 161–176. [Google Scholar]
- Wen-Kai, W.U.; Cheng-Qian, L.I.U.; Zhi-Gang, Z.; Shan, L.U. The Selection of Reference Genes in Chlamydomonas reinhardtii P.A. Dangeard by Real-Time Quantitative PCR. Plant Physiol. Commun. 2009, 45, 667–672. [Google Scholar]
- Condeelis, J. Elongation-Factor 1-Alpha, Translation and the Cytoskeleton. Trends Biochem. Sci. 1995, 20, 169–170. [Google Scholar] [CrossRef]
- Thellin, O.; Zorzi, W.; Lakaye, B.; De Borman, B.; Coumans, B.; Hennen, G.; Grisar, T.; Igout, A.; Heinen, E. Housekeeping Genes as Internal Standards: Use andLimits. J. Biotechnol. 1999, 75, 291–295. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Yuan, M.; Gao, X.; Kang, T.; Zhan, S.; Wan, H.; Li, J. Identification and Validation of Reference Genes for Gene Expression Analysis Using Quantitative PCR in Spodoptera litura (Lepidoptera: Noctuidae). PLoS ONE 2013, 8, e68059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Jonge, H.J.M.; Fehrmann, R.; De Bont, E.S.J.M.; Hofstra, R.; Gerbens, F.; Kamps, W.A.; de Vries, E.; Van Der Zee, A.G.J.; Meerman, G.J.T.; Ter Elst, A. Evidence Based Selection of Housekeeping Genes. PLoS ONE 2007, 2, e898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, H.-F.; Meng, Y.-P.; Cui, G.-M.; Cao, Q.-F.; Li, J.; Liang, A.-H. Selection of Housekeeping Genes for Gene Expression Studies on the Development of Fruit Bearing Shoots in Chinese Jujube (Ziziphus jujube Mill.). Mol. Biol. Rep. 2009, 36, 2183–2190. [Google Scholar] [CrossRef]
- Huis, R.; Hawkins, S.; Neutelings, G. Selection of Reference Genes for Quantitative Gene Expression Normalization in Flax (Linum usitatissimum L.). BMC Plant Biol. 2010, 10, 14. [Google Scholar] [CrossRef] [Green Version]
- Silver, N.; Best, S.; Jiang, J.; Thein, S.L. Selection of Housekeeping Genes for Gene Expression Studies in Human Reticulocytes Using Real-Time PCR. BMC Mol. Biol. 2006, 7, 9. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of Stable Housekeeping Genes, Differentially Regulated Target Genes and Sample Integrity: BestKeeper-Excel-based Tool Using Pair-wise Correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate Nnormalization of Real-Time Quantitative RT-PCR Data by Geometric Averaging of Multiple Internal Control Genes. Genome Biol. 2002, 3, research0034.1. [Google Scholar] [CrossRef] [Green Version]
- Andersen, C.L.; Jensen, J.L.; Orntoft, T.F. Normalization of Real-Time Quantitative Reverse Transcription-PCR Data: A Model-based Variance Estimation Approach to Identify Genes Suited for Normalization, Applied to Bladder and Colon Cancer Data Sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [Green Version]
- Xie, F.; Xiao, P.; Chen, D.; Xu, L.; Zhang, B. miRDeepFinder: A miRNA Analysis Tool for Deep Sequencing of Plant Small RNAs. Plant Mol. Biol. 2012, 80, 75–84. [Google Scholar] [CrossRef]
- Viglierchio, D.R.; Schmitt, R.V. On the Methodology of Nematode Extraction from Field Samples-baermann Funnel Modifications. J. Nematol. 1983, 15, 438–444. [Google Scholar] [PubMed]
- Kikuchi, T.; Cotton, J.A.; Dalzell, J.J.; Hasegawa, K.; Kanzaki, N.; McVeigh, P.; Takanashi, T.; Tsai, I.J.; Assefa, S.A.; Cock, P.J.; et al. Genomic Insights into the Origin of Parasitism in the Emerging Plant Pathogen Bursaphelenchus xylophilus. PLoS Pathog. 2011, 7, e1002219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baermann, G. A Simple Method for the Detection of Ankylostomum (Nematode) Larvae in Soil Tests. In Mededelingen uit het Geneeskundig Laboratorium te Weltevreden; Javasche Boekhandel & Drukkerij: Batavia, IL, USA, 1917; pp. 41–47. [Google Scholar]
- Freckman, D.W.; Mankau, R.; Ferris, H. Nematode Community Structure in Desert Soils-Nematode Recovery. J. Nematol. 1975, 7, 343–346. [Google Scholar] [PubMed]
- Zhou, L.; Chen, F.; Pan, H.; Ye, J.; Dong, X.; Li, C.; Lin, F. Identifying Virulence-Associated Genes Using Transcriptomic and Proteomic Association Analyses of the Plant Parasitic Nematode Bursaphelenchus mucronatus. Int. J. Mol. Sci. 2016, 17, 1492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.F.; Chen, F.M.; Ye, J.R.; Pan, H.Y. Selection of Reliable Reference Genes for RT-qPCR Analysis of Bursaphelenchus mucronatus Gene Expression from Different Habitats and Developmental Stages. Front. Genet. 2018, 9, 11. [Google Scholar] [CrossRef] [Green Version]
- Hellemans, J.; Mortier, G.; De Paepe, A.; Speleman, F.; Vandesompele, J. qBase Relative Quantification Framework and Software for Management and Automated Analysis of Real-Time Quantitative PCR Data. Genome Biol. 2007, 8, 14. [Google Scholar] [CrossRef] [Green Version]
- Kulathunga, D.G.R.S.; Wickramasinghe, S.; Rajapakse, R.P.V.J.; Yatawara, L.; Jayaweera, W.R.; Agatsuma, T. Immunolocalization of Arginine Kinase (AK) in Toxocara canis, Toxocara vitulorum, and Ascaris lumbricoides. Parasitol. Res. 2012, 111, 663–671. [Google Scholar] [CrossRef]
- Qi, X.-L.; Su, X.-F.; Lu, G.-Q.; Liu, C.-X.; Liang, G.-M.; Cheng, H.-M. The effect of Silencing Arginine Kinase by RNAi on the Larval Development of Helicoverpa armigera. Bull. Entomol. Res. 2015, 105, 555–565. [Google Scholar] [CrossRef]
- Wang, F.; Wang, Z.Y.; Li, D.L.; Chen, Q.L. Identification and Characterization of a Bursaphelenchus xylophilus (Aphelenchida: Aphelenchoididae) Thermotolerance-Related Gene: Bx-HSP90. Int. J. Mol. Sci. 2012, 13, 8819–8833. [Google Scholar] [CrossRef]
- Hunter, T.; Garrels, J.I. Characterization of Messenger-Rnas for Alpha-Actin, Beta-Actin and Gamma-Actin. Cell 1977, 12, 767–781. [Google Scholar] [CrossRef]
- Liang, P.; Guo, Y.; Zhou, X.; Gao, X. Expression Profiling in Bemisia tabaci under Insecticide Treatment: Indicating the Necessity for Custom Reference Gene Selection. PLoS ONE 2014, 9, e87514. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Pan, H.; Liu, Y.; Zhou, X. Selection of Reference Genes for Expression Analysis Using Quantitative Real-Time PCR in the Pea Aphid, Acyrthosiphon pisum (Harris) (Hemiptera, Aphidiae). PLoS ONE 2014, 9, e110454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ladror, D.T.; Frey, B.L.; Scalf, M.; Levenstein, M.E.; Artymiuk, J.M.; Smith, L.M. Methylation of Yeast Ribosomal Protein S2 is Elevated During Stationary Phase Growth Conditions. Biochem. Biophys. Res. Commun. 2014, 445, 535–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lü, J.; Chen, S.; Guo, M.; Ye, C.; Qiu, B.; Wu, J.; Yang, C.; Pan, H. Selection and Validation of Reference Genes for RT-qPCR Analysis of the Ladybird Beetle Henosepilachna vigintioctopunctata. Front. Physiol. 2019, 10, 981. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Meng, Q.; Zhu, X.; Sun, S.; Gao, S.; Gou, Y.; Liu, A. Evaluation and Validation of Reference Genes for Quantitative Real-Time PCR in Helopeltis theivora Waterhouse (Hemiptera: Miridae). Sci. Rep. 2019, 9, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dzaki, N.; Ramli, K.N.; Azlan, A.; Ishak, I.H.; Azzam, G. Evaluation of Reference Genes at Different Developmental Stages for QuantitativeReal-Time PCR in Aedes aegypti. Sci. Rep. 2017, 7, srep43618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaros, L.G.; Coutinho, L.; Sider, L.H.; de Medeiros, H.R.; Das Neves, M.R.M.; Benvenuti, C.L.; Navarro, A.M.D.C.; Vieira, L.D.S. Evaluation of Reference Genes for Real-Time PCR Studies of Brazilian Somalis Sheep Infected by Gastrointestinal Nematodes. Genet. Mol. Biol. 2010, 33, 486–490. [Google Scholar] [CrossRef]
- Yuan, M.; Lu, Y.; Zhu, X.; Wan, H.; Shakeel, M.; Zhan, S.; Jin, B.-R.; Li, J. Selection and Evaluation of Potential Reference Genes for Gene Expression Analysis in the Brown Planthopper, Nilaparvata lugens (Hemiptera: Delphacidae) Using Reverse-Transcription Quantitative PCR. PLoS ONE 2014, 9, e86503. [Google Scholar] [CrossRef] [Green Version]
- Shakeel, M.; Zhu, X.; Kang, T.H.; Wan, H.; Li, J.H. Selection and Evaluation of Reference Genes for Quantitative Gene Expression Studies in Cotton Bollworm, Helicoverpa armigera (Lepidoptera: Noctuidae). J. Asia-Pac. Entomol. 2015, 18, 123–130. [Google Scholar] [CrossRef]
- Robledo, D.; Hernández-Urcera, J.; Cal, R.M.; Pardo, B.G.; Sánchez, L.; Martínez, P.; Viñas, A. Analysis of qPCR Reference Gene Stability Determination Methods and a Practical Approach for Efficiency Calculation on a Turbot (Scophthalmus maximus) gonad dataset. BMC Genom. 2014, 15, 648. [Google Scholar] [CrossRef] [Green Version]
- Nakayama, T.; Okada, N.; Yoshikawa, M.; Asaka, D.; Kuboki, A.; Kojima, H.; Tanaka, Y.; Haruna, S.-I. Assessment of Suitable Reference Genes for RT-qPCR Studies in Chronic Rhinosinusitis. Sci. Rep. 2018, 8, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.; Taki, F.A.; Zhang, Y.; Dobbins, D.L.; Pan, X. Evaluation and Identification of Reliable Reference Genes for Toxicological Study in Caenorhabditis elegans. Mol. Biol. Rep. 2014, 41, 3445–3455. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Xie, W.; Wang, S.; Wu, Q.; Yang, N.; Yang, X.; Pan, H.; Zhou, X.; Bai, L.; Xu, B.; et al. Reference Gene Selection for qRT-PCR Analysis in the Sweetpotato Whitefly, Bemisia tabaci (Hemiptera: Aleyrodidae). PLoS ONE 2013, 8, e53006. [Google Scholar] [CrossRef] [PubMed]
- Bansal, R.; Mittapelly, P.; Chen, Y.; Mamidala, P.; Zhao, C.; Michel, A. Quantitative RT-PCR Gene Evaluation and RNA Interference in the Brown Marmorated Stink Bug. PLoS ONE 2016, 11, e0152730. [Google Scholar] [CrossRef]
- Luo, J.; Ma, C.; Li, Z.; Zhu, B.-Q.; Zhang, J.; Lei, C.-L.; Jin, S.-X.; Hull, J.J.; Chen, L.-Z. Assessment of Suitable Reference Genes for qRT-PCR Analysis in Adelphocoris suturalis. J. Integr. Agric. 2018, 17, 2745–2757. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Primer Sequence(5′ to 3′) | Amplicon Length (bp) | E% | R2 |
|---|---|---|---|---|
| Actin | F:GCTGCCTCCTCTTCTTCCCTC R:GGAGTTGTAGGTGGTCTCGTGGATA | 156 | 99.47 | 0.9908 |
| TUB | F:TGTGATTGTCTTCAAGGCTTCC R:GATTCTGTCTGGGTATTCTTCTCG | 88 | 102.52 | 0.9936 |
| UBCE | F:ACCGCCTGGGACTTTGTATC R:TTGGACAGGAACCGAATGG | 99 | 104.67 | 0.9843 |
| PMP-2 | F:CTCTACTCGCAGATTACCAAACC R:CTGGCAGTCGCTGAAACAA | 143 | 92.86 | 0.9902 |
| HIS | F:GTGGGTCGTCTGCACCGTATCCT R:TTGTCACGAGCGGCGTTTCC | 140 | 103.66 | 0.9892 |
| EF1γ | F:GTTTCTGTCGCATTGAACCTCC R:CGATGGTCAAGAAATAACGGGTAA | 100 | 101.37 | 0.9863 |
| 18S rRNA | F:TCAGGGAACATAGGAGGC R:TCAAGCGAGGAGGAGAAT | 172 | 105.15 | 0.9961 |
| UBQ | F:AGCACGGTATAGTTACAGATTGG R:GCAGAGGATTCAGAGGAGC | 129 | 95.39 | 0.9863 |
| ΔCt | Bestkeeper | NormFinder | geNorm | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Experimental Conditions | CRGs | Stability | Rank | Stability | Rank | Stability | Rank | Stability | Rank |
| Temperature treatment | Actin | 0.459 | 5 | 1.173 | 7 | 0.326 | 5 | 0.249 | 1 |
| TUB | 0.415 | 3 | 0.985 | 3 | 0.183 | 2 | 0.328 | 5 | |
| UBCE | 0.401 | 1 | 1.080 | 4 | 0.160 | 1 | 0.310 | 4 | |
| PMP-2 | 0.402 | 4 | 1.156 | 6 | 0.189 | 3 | 0.289 | 3 | |
| HIS | 0.637 | 6 | 0.631 | 1 | 0.566 | 8 | 0.488 | 8 | |
| EF1γ | 0.439 | 2 | 1.131 | 5 | 0.274 | 4 | 0.249 | 1 | |
| 18S rRNA | 0.560 | 8 | 1.315 | 8 | 0.481 | 6 | 0.368 | 6 | |
| UBQ | 0.587 | 7 | 0.702 | 2 | 0.495 | 7 | 0.438 | 7 | |
| Developmental stages | Actin | 0.433 | 3 | 0.363 | 2 | 0.306 | 4 | 0.109 | 1 |
| TUB | 0.450 | 4 | 0.373 | 3 | 0.338 | 5 | 0.109 | 1 | |
| UBCE | 0.384 | 2 | 0.687 | 5 | 0.088 | 2 | 0.254 | 5 | |
| PMP-2 | 0.484 | 6 | 0.304 | 1 | 0.396 | 6 | 0.124 | 3 | |
| HIS | 0.450 | 5 | 0.813 | 6 | 0.211 | 3 | 0.305 | 6 | |
| EF1γ | 0.380 | 1 | 0.566 | 4 | 0.088 | 1 | 0.204 | 4 | |
| 18S rRNA | 0.851 | 8 | 0.973 | 7 | 0.793 | 8 | 0.514 | 8 | |
| UBQ | 0.670 | 7 | 1.098 | 8 | 0.579 | 7 | 0.402 | 7 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.; Zhou, J.; Liu, Y.; Gu, Y.; Zhang, H.; Ahmad, F.; Wang, G.; Ren, L. Selection and Validation of Reliable Reference Genes for qRT-PCR Normalization of Bursaphelenchus xylophilus from Different Temperature Conditions and Developmental Stages. Appl. Sci. 2022, 12, 2880. https://0-doi-org.brum.beds.ac.uk/10.3390/app12062880
Wu Y, Zhou J, Liu Y, Gu Y, Zhang H, Ahmad F, Wang G, Ren L. Selection and Validation of Reliable Reference Genes for qRT-PCR Normalization of Bursaphelenchus xylophilus from Different Temperature Conditions and Developmental Stages. Applied Sciences. 2022; 12(6):2880. https://0-doi-org.brum.beds.ac.uk/10.3390/app12062880
Chicago/Turabian StyleWu, Yajing, Jiao Zhou, Yaning Liu, Yutong Gu, Hongxia Zhang, Faheem Ahmad, Guochang Wang, and Lili Ren. 2022. "Selection and Validation of Reliable Reference Genes for qRT-PCR Normalization of Bursaphelenchus xylophilus from Different Temperature Conditions and Developmental Stages" Applied Sciences 12, no. 6: 2880. https://0-doi-org.brum.beds.ac.uk/10.3390/app12062880