
Quantitative Electroencephalogram (qEEG) as a Natural and Non-Invasive Window into Living Brain and Mind in the Functional Continuum of Healthy and Pathological Conditions
Abstract
:Featured Application
Abstract
1. Introduction
2. Why the Electroencephalogram (EEG)?
2.1. qEEG Historicism
2.2. qEEG Stability, Reliability, and Specificity
2.3. qEEG Heritability
- (a)
- qEEG beta oscillations (beta rhythm is electromagnetic oscillations in the frequency range of brain activity above 13 Hz)
- (b)
- qEEG alpha oscillations (alpha rhythm is electromagnetic oscillations in the frequency range of 8–13 Hz, arising from the synchronous and coherent electrical activity of neurons in the human brain)
- (c)
- qEEG theta oscillations (theta rhythm is electromagnetic oscillations in the frequency range of brain activity between 4 and 7.5 Hz)
2.4. qEEG and Structural Integrity of the Brain
2.5. qEEG and Functional Integrity of the Brain
2.6. qEEG and Vegetative Status/Autonomic Nervous Systems (ANS)
2.7. qEEG, Stress, and Immunity
2.8. qEEG and Cerebral Haemodynamics and Metabolism
2.9. qEEG and Neurotransmitters
2.10. qEEG and Neuropsychopathology
- Electrical changes that precede the clinical onset of a seizure by tens of seconds to minutes—the early detection of a seizure. It has been shown that patients go through a preictal transition for approximately 0.5 to 1 h before a seizure occurs [266]. On average, the prediction rate is ~81% and has an average warning time of 63 min [267];
- Whether a given seizure is epileptic or nonepileptic in origin: For example, there are groups of disorders that produce symptoms similar to an epileptic seizure: (a) cardiac arrhythmias causing syncope, episodes caused by cerebrovascular disease, movement disorders, and unusual manifestations of sleep disorders; (b) events of psychiatric origin (often referred to as psychogenic nonepileptic seizures (PNES)) [268];
- Subclinical seizures: Some seizures recorded during prolonged EEG monitoring may be asymptomatic or ‘subclinical’;
- Whether the cognitive impairments and behavioral problems in question are due to emotional, psychological, or social factors or because of brain dysfunctions or sensory deficits with quantitatively demonstrable abnormalities in brain electrical activity;
- Whether the hyperactive sensation-seeking behavior (typical for ADHD and mania) is due to hypervigilance or vigilance autostabilization behavior, which is a compensatory behavioral pattern to counter regulate a hypovigilance state, and whether withdrawal behavior (typical for depression) is due to hypovigilance or the result of a compensatory behavioral pattern that counter-regulates hypervigilance [269,270];
- Between a degenerative disorder such as AD and pseudodementia due to psychiatric illness [271];
- Between normal and abnormal maturational patterns, such as brain maturation lag (characterized by a pattern of qEEG that is typical for younger age) and brain maturational deviation (characterized by a pattern of qEEG that is not normal at any age) [253];
2.11. qEEG and Experiential Selfhood
2.12. qEEG and Disorders of Consciousness
2.13. qEEG and Aging
2.14. qEEG and Death
2.15. Causality of qEEG Oscillatory Patterns in Neuropsychopathology
3. qEEG Functional Structure and Signal Processing
- The size of oscillatory states repertoire;
- The number of functionally active oscillatory states;
- The relative incidence of change in oscillatory state types;
- Neuronal assembly disassembling;
- The speed of neuronal assemblies disassembling;
- The life-span of oscillatory states;
- The size of the neuronal ensemble;
- The probability of occurrence of a particular type of oscillatory state;
- The stability of local neuronal synchronization within a neuronal assembly;
- The sequence of oscillatory state types.
- The size of the repertoire of oscillatory states;
- The number of functionally active oscillatory states;
- The relative incidence of change in the type of oscillatory states;
- The life-span of oscillatory states;
- The size of the neuronal ensemble;
- The probability of occurrence of a particular type of oscillatory state;
- Neuronal assembly growth;
- The speed of neuronal assemblies growing;
- The sequence of oscillatory state types.

- The size of the OMs’ repertoire;
- The number of functionally active OMs;
- The relative incidence of change in the type of OM;
- The life-span of OMs;
- The probability of occurrence of a particular type of OM;
- The stability of the sequence of types of OMs.
- The size of the OMs’ repertoire;
- The number of functionally active OMs;
- The relative incidence of change in the type of OM;
- The life-span of OMs;
- The probability of occurrence of a particular type of OM;
- The sequence of types of OMs.

4. Common/Unifying Theoretical–Conceptual Framework
4.1. qEEG Phenotype and Neuropsychophysiological Type
4.2. qEEG Phenotype and Neurophysiological, Cognitive, and Behavioral Traits
- Negative valence domain;
- Positive valence systems;
- Cognitive systems;
- Systems for social processes;
- Arousal/regulatory systems.
- Tonic level of vigilance (corresponds to arousal/regulatory systems in RDoC);
- Speed of information processing (corresponds to cognitive systems in RDoC);
- Directedness of the attention (internal vs. external focus) (corresponds to cognitive systems in RDoC);
- Emotional–motivational tendency (corresponds to positive and negative valence systems in RDoC);
- Sociability (sensory stimulation and excitement tolerability) (corresponds to systems for social processes in RDoC);
- Anxiety tendency (anxious arousal vs. anxious apprehension) (corresponds to negative valence and arousal/regulatory systems in RDoC);
- Stress regulation (resistance and recovery);
- Overall brain resources (resilience).
4.2.1. Tonic Vigilance
4.2.2. Speed of Information Processing
4.2.3. Directedness of the Attention (Internal vs. External Focus)
4.2.4. Emotional–Motivational Tendency
4.2.5. Sociability (Sensory Stimulation and Excitement Tolerability)
4.2.6. Anxiety Tendency (Anxious Arousal vs. Anxious Apprehension)
4.2.7. Stress Regulation (Resistance and Recovery)
4.2.8. Overall Brain Resources (Resilience)
4.3. Why the Resting State?
4.4. qEEG Phenotype, Personality Traits, and Norm-Pathology Continuum
- Successful performance;
- Maximum efficiency;
- Minimal cost;
- Temporal adequacy.
5. Summary and Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lazarev, V.V. The relationship of theory and methodology in EEG studies of mental activity. Int. J. Psychophysiol. 2006, 62, 384–393. [Google Scholar] [CrossRef] [PubMed]
- Peled, A. Brain profiling and clinical-neuroscience. Med. Hypotheses 2006, 67, 941–946. [Google Scholar] [CrossRef] [PubMed]
- Kropotov, J.D. Quantitative EEG, Event-Related Potentials and Neurotherapy; Elsevier: Oxford, UK, 2009; p. 531. [Google Scholar]
- Bellavite, P.; Signorini, A. Pathology, complex systems, and resonance. In Fundamental Research in Ultra-High Dilution and Homoeopathy; Schulte, J., Endler, P.C., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1998; pp. 105–116. [Google Scholar]
- Basar, E. Brain-Body-Mind in the Nebulous Cartesian System: A Holistic Approach by Oscillations; Springer: New York, NY, USA, 2011; p. 523. [Google Scholar]
- Motamedi-Fakhr, S.; Moshrefi-Torbati, M.; Hill, M.; Hill, C.M.; White, P.R. Signal processing techniques applied to human sleep EEG signals—A review. Biomed. Signal Process. Control. 2014, 10, 21–33. [Google Scholar] [CrossRef]
- Da Silva, F.L. Neural mechanisms underlying brain waves: From neural membranes to networks. Electroencephalogr. Clin. Neurophysiol. 1991, 79, 81–93. [Google Scholar] [CrossRef]
- Nunez, P.L. Neocortical Dynamics and Human EEG Rhythms; Oxford University Press: New York, NY, USA, 1995; p. 730. [Google Scholar]
- Michel, C.M.; Brandeis, D. The sources and temporal dynamics of scalp electric fields. In Simultaneous EEG and fMRI. Recording, Analysis, and Application; Ullsperger, M., Debener, S.S., Eds.; Oxford University Press: New York, NY, USA, 2010; pp. 1–19. [Google Scholar]
- Freeman, W.J. Mass Action in the Nervous System. Examination of the Neurophysiological Basis of Adaptive Behavior through the EEG; Academic Press: New York, NY, USA, 1975; p. 489. [Google Scholar]
- Corsi-Cabrera, M.; Herrera, P.; Malvido, M. Correlation between EEG and cognitive abilities: Sex differences. Int. J. Neurosci. 1989, 45, 133–141. [Google Scholar] [CrossRef]
- Tsodyks, M.; Kenet, T.; Grinvald, A.; Arieli, A. Linking spontaneous activity of single cortical neurons and the underlying functional architecture. Science 1999, 286, 1943–1946. [Google Scholar] [CrossRef]
- Nunez, P.L. Toward a quantitative description of large-scale neocortical dynamic function and EEG. Behav. Brain Sci. 2000, 23, 371–437. [Google Scholar] [CrossRef]
- Bressler, S.L.; Kelso, J.A.S. Cortical coordination dynamics and cognition. Trends Cogn. Sci. 2001, 5, 26–36. [Google Scholar] [CrossRef]
- Moran, R.J.; Stephan, K.E.; Kiebel, S.J.; Rombach, N.; O’Connor, W.T.; Murphy, K.J.; Reilly, R.B.; Friston, K.J. Bayesian estimation of synaptic physiology from the spectral responses of neural masses. Neuroimage 2008, 42, 272–284. [Google Scholar] [CrossRef]
- Hadjipapas, A.; Casagrande, E.; Nevado, A.; Barnes, G.R.; Green, G.; Holliday, I.E. Can we observe collective neuronal activity from macroscopic aggregate signals? Neuroimage 2009, 44, 1290–1303. [Google Scholar] [CrossRef]
- van Albada, S.J.; Kerr, C.C.; Chiang, A.K.I.; Rennie, C.J.; Robinson, P.A. Neurophysiological changes with age probed by inverse modelling of EEG spectra. Clin. Neurophysiol. 2010, 121, 21–38. [Google Scholar] [CrossRef] [PubMed]
- Fingelkurts, A.A.; Fingelkurts, A.A. Short-term EEG spectral pattern as a single event in EEG phenomenology. Open Neuroimag. J. 2010, 4, 130–156. [Google Scholar] [CrossRef] [PubMed]
- Klimesch, W. Memory processes, brain oscillations and EEG synchronization. Int. J. Psychophysiol. 1996, 24, 61–100. [Google Scholar] [CrossRef]
- Da Silva, F.H.L. The generation of electric and magnetic signals of the brain by local networks. In Comprehensive Human Physiology; Greger, R., Windhorst, U., Eds.; Springer: Berlin/Heidelberg, Germany, 1996; Volume 1, pp. 509–528. [Google Scholar]
- Klimesch, W. EEG alpha and theta oscillations reflect cognitive and memory performance: A review and analysis. Brain Res. Rev. 1999, 29, 169–195. [Google Scholar] [CrossRef]
- Basar, E. Brain Function and Oscillations. I Vol. Brain Oscillations: Principles and Approaches; Springer: Berlin/Heidelberg, Germany, 1998; p. 396. [Google Scholar]
- Basar, E.; Basar-Eroglu, C.; Karakas, S.; Schurmann, M. Are cognitive processes manifested in event-related gamma, alpha, theta and delta oscillations in the EEG? Neurosci. Lett. 1999, 259, 165–168. [Google Scholar] [CrossRef]
- Basar, E.; Basar-Eroglu, C.; Karakas, S.; Schurmann, M. Brain oscillations in perception and memory. Int. J. Psychophysiol. 2000, 35, 95–124. [Google Scholar] [CrossRef]
- Klimesch, W. Interindividual differences in oscillatory EEG activity and cognitive performance. In The Cognitive Neuroscience of Individual Differences; Reinvang, I., Greenlee, M., Herrmann, M., Eds.; BIS: Oldenburg, Germany, 2003; pp. 87–99. [Google Scholar]
- Basar, E.; Özgören, M.; Karakas, S.; Basar-Eroglu, C. Super-synergy in the brain: The grandmother percept is manifested by multiple oscillations. Int. J. Bifurcat. Chaos 2004, 14, 453–491. [Google Scholar] [CrossRef]
- Klimesch, W.; Schack, B.; Sauseng, P. The functional significance of theta and upper alpha oscillations. Exp. Psychol. 2005, 52, 99–108. [Google Scholar] [CrossRef]
- Jurko, M.F.; Giurintano, L.P.; Giurintano, S.L.; Andy, O.J. Spontaneous awake EEG patterns in three lines of primate evolution. Behav. Biol. 1974, 10, 377–384. [Google Scholar] [CrossRef]
- Van den Bergh, B.R.; Mulder, E.J.; Mennes, M.; Glover, V. Antenatal maternal anxiety and stress and the neurobehavioural development of the fetus and child: Links and possible mechanisms. A review. Neurosci. Biobehav. Rev. 2005, 29, 237–258. [Google Scholar] [CrossRef]
- Class, Q.A.; Lichtenstein, P.; Langstrom, N.; D’Onofrio, B.M. Timing of prenatal maternal exposure to severe life events and adverse pregnancy outcomes: A population study of 2.6 million pregnancies. Psychosom. Med. 2011, 73, 234–241. [Google Scholar] [CrossRef] [PubMed]
- Grigoriadis, S.; VonderPorten, E.H.; Mamisashvili, L.; Tomlinson, G.; Dennis, C.L.; Koren, G.; Steiner, M.; Mousmanis, P.; Cheung, A.; Radford, K.; et al. The impact of maternal depression during pregnancy on perinatal outcomes: A systematic review and meta-analysis. J. Clin. Psychiatry 2013, 74, e321–e341. [Google Scholar] [CrossRef] [PubMed]
- Slykerman, R.F.; Thompson, J.; Waldie, K.; Murphy, R.; Wall, C.; Mitchell, E.A. Maternal stress during pregnancy is associated with moderate to severe depression in 11-year-old children. Acta Paediatr. 2015, 104, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulou, Z.; Vlaikou, A.M.; Theodoridou, D.; Markopoulos, G.S.; Tsoni, K.; Agakidou, E.; Drosou-Agakidou, V.; Turck, C.W.; Filiou, M.D.; Syrrou, M. Stressful newborn memories: Pre-conceptual, in utero, and postnatal events. Front. Psychiatry 2019, 10, 220. [Google Scholar] [CrossRef] [PubMed]
- Prichep, L.S.; Kowalik, S.C.; Alper, K.; de Jesus, C. Quantitative EEG characteristics of children exposed in utero to cocaine. Clin. Electroencephalogr. 1995, 26, 166–172. [Google Scholar] [CrossRef]
- Matousek, M.; Petersen, I. Frequency analysis of the EEG in normal children and adolescents. In Automation of Clinical Electroencephalography; Kellaway, P., Petersen, I., Eds.; Raven Press: New York, NY, USA, 1973; pp. 75–102. [Google Scholar]
- Cragg, L.; Kovacevic, N.; McIntosh, A.R.; Poulsen, C.; Martinu, K.; Leonard, G.; Paus, T. Maturation of EEG power spectra in early adolescence: A longitudinal study. Dev. Sci. 2011, 14, 935–943. [Google Scholar] [CrossRef]
- Scraggs, T.L. EEG maturation: Viability through adolescence. Neurodiagn. J. 2012, 52, 176–203. [Google Scholar]
- Kaminska, A.; Eisermann, M.; Plouin, P. Child EEG (and maturation). In Handbook of Clinical Neurology, Clinical Neurophysiology: Basis and Technical Aspects, 3rd ed.; Levin, K.H., Chauvel, P., Eds.; Elsevier B.V.: Amsterdam, The Netherlands, 2019; Volume 160, pp. 125–142. [Google Scholar]
- Zhadin, M.N. Rhythmic processes in the cerebral cortex. J. Theor. Biol. 1984, 108, 565–595. [Google Scholar] [CrossRef]
- Anokhin, A.P.; Birbaumer, N.; Lutzenberger, W.; Nikolaev, A.; Vogel, F. Age increases brain complexity. Electroencephalogr. Clin. Neurophysiol. 1996, 99, 63–68. [Google Scholar] [CrossRef]
- Schutter, D.J.L.G.; Leitner, C.; Kenemans, J.L.; van Honk, J. Electrophysiological correlates of cortico-subcortical interaction: A cross-frequency spectral EEG analysis. Clin. Neurophysiol. 2006, 117, 381–387. [Google Scholar] [CrossRef]
- Marosi, E.; Harmony, T.; Sánchez, L.; Becker, J.; Bernal, J.; Reyes, A.; de León, A.E.D.; Rodríguez, M.; Fernández, T. Maturation of the coherence of EEG activity in normal and learning-disabled children. EEG Clin. Neurophysiol. 1992, 83, 350–357. [Google Scholar] [CrossRef]
- Lukashevich, I.P.; Machinskaya, R.I.; Fishman, M.N. Diagnosis of the functional state of the brain in young school-age children with learning difficulties. Hum. Physiol. (Fiziol Cheloveka) 1994, 20, 34–46. [Google Scholar]
- Ulrich, G. Psychiatric Electroencephalography. In Updated and Revised Edition (2002) of the Original Textbook Psychiatrische Elektroenzephalographie (in German); Gustav Fischer Verlag: New York, NY, USA, 1994; p. 343. [Google Scholar]
- Zentner, M. Antisocial personalities. In Adult Psychopathology. A Social Work Perspective; Turner, F.J., Ed.; The Free Press: New York, NY, USA, 1984; pp. 345–363. [Google Scholar]
- Davies, R.K. Incest: Some neuropsychiatric findings. Int. J. Psychiatry Med. 1979, 9, 117–121. [Google Scholar] [CrossRef] [PubMed]
- McFarlane, A.; Clark, C.R.; Bryant, R.A.; Williams, L.M.; Niaura, R.; Paul, R.H.; Hitsman, B.L.; Stroud, L.; Alexander, D.M.; Gordon, E. The impact of early life stress on psychophysiological, personality and behavioural measures in 740 non-clinical subjects. J. Integr. Neurosci. 2005, 4, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.E. Mechanisms linking early life stress to adult health outcomes. Proc. Natl. Acad. Sci. USA 2010, 107, 8507–8512. [Google Scholar] [CrossRef]
- Harmony, T.; Alvarez, A.; Pascual, R.; Ramos, A.; Marosi, E.; De León, A.E.D.; Valdés, P.; Becker, J. EEG maturation on children with different economic and psychosocial characteristics. Int. J. Neurosci. 1988, 41, 103–113. [Google Scholar] [CrossRef]
- Otero, G.A.; Pliego-Rivero, F.B.; Fernández, T.; Ricardo, J. EEG development in children with sociocultural disadvantages: A follow-up study. Clin. Neurophysiol. 2003, 114, 1918–1925. [Google Scholar] [CrossRef]
- Marshall, P.J.; Fox, N.A. Bucharest early intervention project core group. A comparison of the electroencephalogram between institutionalized and community children in Romania. J. Cogn. Neurosci. 2004, 16, 1327–1338. [Google Scholar] [CrossRef]
- Howells, F.M.; Stein, D.J.; Russell, V.A. Childhood trauma is associated with altered cortical arousal: Insights from an EEG study. Front. Integr. Neurosci. 2012, 6, 120. [Google Scholar] [CrossRef]
- Vanderwert, R.E.; Zeanah, C.H.; Fox, N.A.; Nelson, C.A. Normalization of EEG activity among previously institutionalized children placed into foster care: A 12-year follow-up of the Bucharest Early Intervention Project. Dev. Cogn. Neurosci. 2016, 17, 68–75. [Google Scholar] [CrossRef]
- Bosch-Bayard, J.; Razzaq, F.A.; Lopez-Naranjo, C.; Wang, Y.; Li, M.; Galan-Garcia, L.; Calzada-Reyes, A.; Virues-Alba, T.; Rabinowitz, A.G.; Suarez-Murias, C.; et al. Early protein energy malnutrition impacts life-long developmental trajectories of the sources of EEG rhythmic activity. NeuroImage 2022, 254, 119144. [Google Scholar] [CrossRef] [PubMed]
- Black, L.M.; Hudspeth, W.J.; Townsend, A.L.; Bodenhamer-Davis, E. EEG Connectivity Patterns in Childhood Sexual Abuse: A Multivariate Application Considering Curvature of Brain Space. J. Neurother. 2008, 12, 141–160. [Google Scholar] [CrossRef]
- Lee, S.-H.; Park, Y.; Jin, M.J.; Lee, Y.J.; Hahn, S.W. Childhood trauma associated with enhanced high frequency band powers and induced subjective inattention of adults. Front. Behav. Neurosci. 2017, 11, 148. [Google Scholar] [CrossRef]
- Thatcher, R.W.; Walker, R.A.; Gerson, I.; Geisler, F.H. EEG discriminant analysis of mild head trauma. Electroencephalogr. Clin. Neurophysiol. 1989, 73, 10–94. [Google Scholar] [CrossRef]
- Hooshmand, H.; Beckner, E.; Radfar, R. Technical and clinical aspects of topographic brain mapping. Clin. Electroencephalogr. 1989, 20, 235–247. [Google Scholar] [CrossRef] [PubMed]
- Thornton, K.E. The electrophysiological effects of a brain injury on auditory memory functioning: The QEEG correlates of impaired memory. Arch. Clin. Neuropsychol. 2003, 18, 363–378. [Google Scholar] [CrossRef] [PubMed]
- Stassen, H.H. Computerized recognition of persons by EEG spectral patterns. Electroencephalogr. Clin. Neurophysiol. 1980, 49, 190–194. [Google Scholar] [CrossRef]
- Gasser, T.; Bacher, P.; Steinberg, H. Test–retest reliability of spectral parameters of the EEG. Electroencephalogr. Clin. Neurophysiol. 1985, 60, 312–319. [Google Scholar] [CrossRef]
- Salinsky, M.C.; Oken, B.S.; Morehead, L. Test–retest reliability in EEG frequency analysis. Electroencephalogr. Clin. Neurophysiol. 1991, 79, 382–392. [Google Scholar] [CrossRef]
- Pollock, V.E.; Schneider, L.S.; Lyness, S.A. Reliability of topographic quantitative EEG amplitude in healthy late-middle-aged and elderly subjects. Electroencephalogr. Clin. Neurophysiol. 1991, 79, 20–26. [Google Scholar] [CrossRef]
- Burgess, A.; Gruzelier, J. Individual reliability of amplitude distribution in topographical mapping of EEG. Electroencephalogr. Clin. Neurophysiol. 1993, 86, 219–223. [Google Scholar] [CrossRef]
- Harmony, T.; Fernandez, T.; Rodriguez, M.; Reyes, A.; Marosi, E.; Bernal, J. Test–retest reliability of EEG spectral parameters during cognitive tasks: II. Coherence. Int. J. Neurosci. 1993, 68, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Lund, T.R.; Sponheim, S.R.; Iacono, W.G.; Clementz, B.A. Internal consistency reliability of resting EEG power spectra in schizophrenic and normal subjects. Psychophysiology 1995, 32, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Stassen, H.H.; Bomben, G.; Hell, D. Familial brain wave patterns: Study of a 12-sib family. Psychiatr. Genet. 1998, 8, 141–153. [Google Scholar] [CrossRef]
- Dustman, R.E.; Shearer, D.E.; Emmerson, R.Y. Life-span changes in EEG spectral amplitude, amplitude variability and mean frequency. Clin. Neurophysiol. 1999, 110, 1399–1409. [Google Scholar] [CrossRef]
- Kondacs, A.; Szabo, M. Long-term intra-individual variability of the background EEG in normals. Clin. Neurophysiol. 1999, 110, 1708–1716. [Google Scholar] [CrossRef]
- Dünki, R.M.; Schmid, B.; Stassen, H.H. Intraindividual specificity and stability of human EEG: Comparing a linear vs. a onlinear approach. Methods Inf. Med. 2000, 39, 78–82. [Google Scholar]
- Poulos, M.; Rangoussi, M.; Alexandris, N.; Evangelou, A. Person identification from the EEG using nonlinear signal classification. Methods Inf. Med. 2002, 41, 64–75. [Google Scholar]
- Maltez, J.; Hyllienmark, L.; Nikulin, V.V.; Brismar, T. Time course and variability of power in different frequency bands of EEG during resting conditions. Neurophysiol. Clin. 2004, 34, 195–202. [Google Scholar] [CrossRef]
- Fingelkurts, A.A.; Fingelkurts, A.A.; Ermolaev, V.A.; Kaplan, A.Y. Stability, reliability and consistency of the compositions of brain oscillations. Int. J. Psychophysiol. 2006, 59, 116–126. [Google Scholar] [CrossRef]
- Vuga, M.; Fox, N.A.; Cohn, J.F.; George, C.J.; Levenstein, R.M.; Kovacs, M. Long-term stability of frontal electroencephalographic asymmetry in adults with a history of depression and controls. Int. J. Psychophysiol. 2006, 59, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Näpflin, M.; Wildi, M.; Sarnthein, J. Test-retest reliability of resting EEG spectra validates a statistical signature of persons. Clin. Neurophysiol. 2007, 118, 2519–2524. [Google Scholar] [CrossRef] [PubMed]
- Towers, D.N.; Allen, J.J. A better estimate of the internal consistency reliability of frontal EEG asymmetry scores. Psychophysiology 2009, 46, 132–142. [Google Scholar] [CrossRef] [PubMed]
- Cannon, R.L.; Baldwin, D.R.; Shaw, T.L.; Diloreto, D.J.; Phillips, S.M.; Scruggs, A.M.; Riehl, T.C. Reliability of quantitative EEG (qEEG) measures and LORETA current source density at 30 days. Neurosci. Lett. 2012, 518, 27–31. [Google Scholar] [CrossRef]
- Grandy, T.H.; Werkle-Bergner, M.; Chicherio, C.; Schmiedek, F.; Lövdén, M.; Lindenberger, U. Peak individual alpha frequency qualifies as a stable neurophysiological trait marker in healthy younger and older adults. Psychophysiology 2013, 50, 570–582. [Google Scholar] [CrossRef]
- Vogel, F. The genetic basis of the normal human electroencephalogram (EEG). Humangenetik 1970, 10, 91–114. [Google Scholar] [CrossRef]
- Lykken, D.T.; Tellegen, A.; Thorkelson, K. Genetic determination of EEG frequency spectra. Biol. Psychol. 1974, 1, 245–259. [Google Scholar] [CrossRef]
- Lykken, D.T.; Tellegen, A.; Iacono, W.G. EEG spectra in twins: Evidence for a neglected mechanism of genetic determination. Physiol. Psychol. 1982, 10, 60–65. [Google Scholar] [CrossRef]
- Stassen, H.H.; Bomben, G.; Propping, P. Genetic aspects of the EEG: An investigation into the within-pair similarity of monozigotic and dyzigotic twins with a new method of analysis. Electroencephalogr. Clin. Neurophysiol. 1987, 66, 489–501. [Google Scholar] [CrossRef]
- van Beijsterveldt, C.E.; Boomsma, D.I. Genetics of the human electroencephalogram (EEG) and event-related brain potentials (ERPs): A review. Hum. Genet. 1994, 94, 319–330. [Google Scholar] [CrossRef]
- Christian, J.C.; Morzorati, S.; Norton, J.A., Jr.; Williams, C.J.; O’Connor, S.; Li, T.K. Genetic analysis of the resting electroencephalographic power spectrum in human twins. Psychophysiology 1996, 33, 584–591. [Google Scholar] [CrossRef] [PubMed]
- van Baal, G.C.; De Geus, E.J.; Boomsma, D.I. Genetic architecture of EEG power spectra in early life. Electroencephalogr. Clin. Neurophysiol. 1996, 98, 502–514. [Google Scholar] [CrossRef]
- van Beijsterveldt, C.E.; Molenaar, P.C.; De Geus, E.J.; Boomsma, D.I. Heritability of human brain functioning as assessed by electroencephalography. Am. J. Hum. Genet. 1996, 58, 562–573. [Google Scholar] [PubMed]
- Posthuma, D.; Neale, M.C.; Boomsma, D.I.; De Geus, E.J.C. Are smarter brains running faster? Heritability of alpha peak frequency, IQ, and their interrelation. Behav. Genet. 2001, 31, 567–579. [Google Scholar] [CrossRef]
- van Baal, G.; van Beijsterveldt, C.; Molenaar, P.; Boomsma, D.; De Geus, E. A genetic perspective on the developing brain: Electrophysiological indices of neural functioning in young and adolescent twins. Eur. Psychol. 2001, 6, 254–263. [Google Scholar] [CrossRef]
- van Beijsterveldt, C.E.M.; van Baal, G. Twin and family studies of the human electroencephalogram: A review and a meta-analysis. Biol. Psychol. 2002, 61, 111–138. [Google Scholar] [CrossRef]
- Smit, D.J.A.; Posthuma, D.; Boomsma, D.I.; De Geus, E.J.C. Heritability of background EEG across the power spectrum. Psychophysiology 2005, 42, 691–697. [Google Scholar] [CrossRef]
- Anokhin, A.P.; Müller, V.; Lindenberger, U.; Heath, A.C.; Myers, E. Genetic influences on dynamic complexity of brain oscillations. Neurosci. Lett. 2006, 397, 93–98. [Google Scholar] [CrossRef]
- Smit, C.M.; Wright, M.J.; Hansell, N.K.; Geffen, G.M.; Martin, N.G. Genetic variation of individual alpha frequency (IAF) and alpha power in a large adolescent twin sample. Int. J. Psychophysiol. 2006, 61, 235–243. [Google Scholar] [CrossRef]
- Tang, Y.; Chorlian, D.B.; Rangaswamy, M.; O’Connor, S.; Taylor, R.; Rohrbaugh, J.; Porjesz, B.; Begleiter, H. Heritability of bipolar EEG spectra in a large sib-pair population. Behav. Genet. 2007, 37, 302–313. [Google Scholar] [CrossRef]
- Eischen, S.E.; Luckritz, J.Y.; Polich, J. Spectral analysis of EEG from families. Biol. Psychol. 1995, 41, 61–68. [Google Scholar] [CrossRef]
- Begleiter, H.; Porjesz, B. Genetics of human brain oscillations. Int. J. Psychophysiol. 2006, 60, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Winterer, G.; Smolka, M.; Samochowiec, J.; Ziller, M.; Mahlberg, R.; Gallinat, J.; Rommelspacher, H.P.; Herrmann, W.M.; Sander, T. Association of EEG coherence and an exonic GABA(B)R1 gene polymorphism. Am. J. Med. Genet. 2003, 117B, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Porjesz, B.; Almasy, L.; Edenberg, H.; Wang, K.; Chorlian, D.B.; Foroud, T.; Goate, A.; Rice, J.P.; O’Connor, S.; Rohrbaugh, J.; et al. Linkage disequilibrium between the beta frequency of the human EEG and a GABAa receptor gene locus. Proc. Natl. Acad. Sci. USA 2002, 99, 3729–3733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edenberg, H.J.; Dick, D.M.; Xuei, X.; Tian, H.; Almasy, L.; Bauer, L.O.; Crowe, R.R.; Goate, A.; Hesselbrock, V.; Jones, K.; et al. Variations in GABRA2, encoding the alpha 2 subunit of the GABA(A) receptor, are associated with alcohol dependence and with brain oscillations. Am. J. Hum. Genet. 2004, 74, 705–714. [Google Scholar] [CrossRef] [PubMed]
- Winterer, G.; Mahlberg, R.; Smolka, M.N.; Samochowiec, J.; Ziller, M.; Rommelspacher, H.P.; Herrmann, W.M.; Schmidt, L.G.; Sander, T. Association analysis of exonic variants of the GABA(B)-receptor gene and alpha electroencephalogram voltage in normal subjects and alcohol-dependent patients. Behav. Genet. 2003, 33, 7–15. [Google Scholar] [CrossRef]
- Ducci, F.; Enoch, M.A.; Yuan, Q.; Shen, P.H.; White, K.V.; Hodgkinson, C.; Goldman, D. HTR3B is associated with alcoholism with antisocial behavior and alpha EEG power--an intermediate phenotype for alcoholism and co-morbid behaviors. Alcohol 2009, 43, 73–84. [Google Scholar] [CrossRef]
- Enoch, M.A.; White, K.V.; Waheed, J.; Goldman, D. Neurophysiological and genetic distinctions between pure and comorbid anxiety disorders. Depress. Anxiety 2008, 25, 383–392. [Google Scholar] [CrossRef]
- Enoch, M.A.; Shen, P.H.; Ducci, F.; Yuan, Q.; Liu, J.; White, K.V.; Albaugh, B.; Hodgkinson, C.A.; Goldman, D. Common genetic origins for EEG, alcoholism and anxiety: The role of CRH-BP. PLoS ONE 2008, 3, e3620. [Google Scholar] [CrossRef]
- Enoch, M.A.; Xu, K.; Ferro, E.; Harris, C.R.; Goldman, D. Genetic origins of anxiety in women: A role for a functional catechol-O-methyltransferase polymorphism. Psychiatr. Genet. 2003, 13, 33–41. [Google Scholar] [CrossRef]
- Enoch, M.A.; Rohrbaugh, J.W.; Davis, E.Z.; Harris, C.R.; Ellingson, R.J.; Andreason, P.; Moore, V.; Varner, J.L.; Brown, G.L.; Eckardt, M.J. Relationship of genetically transmitted alpha EEG traits to anxiety disorders and alcoholism. Am. J. Med. Genet. 1995, 60, 400–408. [Google Scholar] [CrossRef] [PubMed]
- Enoch, M.A.; White, K.V.; Harris, C.R.; Robin, R.W.; Ross, J.; Rohrbaugh, J.W.; Goldman, D. Association of low-voltage alpha EEG with a subtype of alcohol use disorders. Alcohol. Clin. Exp. Res. 1999, 23, 1312–1319. [Google Scholar] [CrossRef] [PubMed]
- Zoon, H.F.; Veth, C.P.; Arns, M.; Drinkenburg, W.H.; Talloen, W.; Peeters, P.J.; Kenemans, J.L. EEG alpha power as an intermediate measure between brain-derived neurotrophic factor Val66Met and depression severity in patients with major depressive disorder. J. Clin. Neurophysiol. 2013, 30, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Bodenmann, S.; Rusterholz, T.; Dürr, R.; Stoll, C.; Bachmann, V.; Geissler, E.; JaggiSchwarz, K.; Landolt, H.P. The functional val158met polymorphism of COMT predicts interindividual differences in brain alpha oscillations in young men. J. Neurosci. 2009, 29, 10855–10862. [Google Scholar] [CrossRef] [Green Version]
- Meyers, J.L.; Zhang, J.; Chorlian, D.B.; Pandey, A.K.; Kamarajan, C.; Wang, J.-C.; Wetherill, L.; Lai, D.; Chao, M.; Chan, G.; et al. A genome-wide association study of interhemispheric theta EEG coherence: Implications for neural connectivity and alcohol use behavior. Mol. Psychiatry 2021, 26, 5040–5052. [Google Scholar] [CrossRef]
- Venables, N.C.; Bernat, E.M.; Sponheim, S.R. Genetic and disorder-specific aspects of resting state EEG abnormalities in schizophrenia. Schizophr. Bull. 2009, 35, 826–839. [Google Scholar] [CrossRef]
- da Silva, F.H.L.; van Rotterdam, A.; Barts, P.; van Heusden, E.; Burr, W. Models of neuronal populations: The basic mechanism of rhythmicity. In Perspectives of Brain Research. Progress in Brain Research; Corner, M.A., Swaab, D.F., Eds.; Elsevier: Amsterdam, The Netherlands, 1976; Volume 45, pp. 281–308. [Google Scholar]
- Hughes, J.R.; Cayaffa, J.J. The EEG in patients at different ages without organic cerebral disease. Electroencephalogr. Clin. Neurophysiol. 1977, 42, 776–784. [Google Scholar] [CrossRef]
- Goldensohn, E.S. Use of EEG for evaluation of focal intracranial lesions. In Current Practice of Clinical Electroencephalography; Klass, D.W., Daly, D.D., Eds.; Raven: New York, NY, USA, 1979; pp. 307–341. [Google Scholar]
- da Silva, F.H.L.; Vos, J.E.; Mooibroek, J.; van Rotterdam, A. Relative contributions of intracortical and thalamo-cortical processes in the generation of alpha rhythms, revealed by partial coherence analysis. Electroencephalogr. Clin. Neurophysiol. 1980, 50, 449–456. [Google Scholar] [CrossRef]
- Steriade, M.; Llinas, R.R. The functional states of the thalamus and the associated neuronal interplay. Physiol. Rev. 1988, 68, 649–742. [Google Scholar] [CrossRef]
- Anokhin, A.; Vogel, F. EEG alpha rhythm frequency and intelligence in normal adults. Intelligence 1996, 23, 1–14. [Google Scholar] [CrossRef]
- Leocani, L.; Locatelli, T.; Martinelli, V.; Rovaris, M.; Falautano, M.; Filippi, M.; Magnani, G.; Comi, G. Electroencephalographic coherence analysis in multiple sclerosis: Correlation with clinical, neuropsychological, and MRI findings. J. Neurol. Neurosurg. Psychiatry 2000, 69, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Moretti, D.V.; Babiloni, C.; Binetti, G.; Cassetta, E.; Dal Forno, G.; Ferreric, F.; Ferri, R.; Lanuzza, B.; Miniussi, C.; Nobili, F.; et al. Individual analysis of EEG frequency and band power in mild Alzheimer’s disease. Clin. Neurophysiol. 2004, 115, 299–308. [Google Scholar] [CrossRef]
- Nunez, P.L.; Srinivasan, R. A theoretical basis for standing and traveling brain waves. Clin. Neurophysiol. 2006, 117, 2425–2435. [Google Scholar] [CrossRef] [PubMed]
- Moretti, D.V.; Miniussi, C.; Frisoni, G.; Zanetti, O.; Binetti, G.; Geroldi, C.; Galluzzi, S.; Rossini, P.M. Vascular damage and EEG markers in subjects with mild cognitive impairment. Neurophysiol. Clin. 2007, 118, 1866–1876. [Google Scholar] [CrossRef] [Green Version]
- Babiloni, C.; Frisoni, G.B.; Pievani, M.; Vecchio, F.; Lizio, R.; Buttiglione, M.; Geroldi, C.; Fracassi, C.; Eusebi, F.; Ferri, R.; et al. Hippocampal volume and cortical sources of EEG alpha rhythms in mild cognitive impairment and Alzheimer disease. Neuroimage 2009, 44, 123–135. [Google Scholar] [CrossRef]
- Valdés-Hernández, P.A.; Ojeda-González, A.; Martínez-Montes, E.; Lage-Castellanos, A.; Virués-Alba, T.; Valdés-Urrutia, L.; Valdes-Sosa, P.A. White matter architecture rather than cortical surface area correlates with the EEG alpha rhythm. NeuroImage 2010, 49, 2328–2339. [Google Scholar] [CrossRef]
- Bhattacharya, B.S.; Coyle, D.; Maguire, L.P. A thalamo-cortico-thalamic neural mass model to study alpha rhythms in Alzheimer’s disease. Neural Netw. 2011, 24, 631–645. [Google Scholar] [CrossRef]
- Jann, K.; Federspiel, A.; Giezendanner, S.; Andreotti, J.; Kottlow, M.; Dierks, T.; Koenig, T. Linking brain connectivity across different time scales with electroencephalogram, functional magnetic resonance imageing, and diffusion tensor imageing. Brain Connect 2012, 2, 11–20. [Google Scholar] [CrossRef]
- Garcés, P.; Vicente, R.; Wibral, M.; Pineda-Pardo, J.Á.; López, M.E.; Aurtenetxe, S.; Marcos, A.; de Andrés, M.E.; Yus, M.; Sancho, M.; et al. Brain-wide slowing of spontaneous alpha rhythms in mild cognitive impairment. Front. Ageing Neurosci. 2013, 5, 100. [Google Scholar] [CrossRef]
- Babiloni, C.; Carducci, F.; Lizio, R.; Vecchio, F.; Baglieri, A.; Bernardini, S.; Cavedo, E.; Bozzao, A.; Buttinelli, C.; Esposito, F.; et al. Resting state cortical electroencephalographic rhythms are related to gray matter volume in subjects with mild cognitive impairment and Alzheimer’s disease. Hum. Brain Mapp. 2013, 34, 1427–1446. [Google Scholar] [CrossRef]
- Hindriks, R.; van Putten, M.J.A.M. Thalamo-cortical mechanisms underlying changes in amplitude and frequency of human alpha oscillations. NeuroImage 2013, 70, 150–163. [Google Scholar] [CrossRef] [PubMed]
- Thatcher, R.W.; Biver, C.; McAlaster, R.; Salazar, A.M. Biophysical linkage between MRI and EEG coherence in traumatic brain injury. NeuroImage 1998, 8, 307–326. [Google Scholar] [CrossRef] [PubMed]
- Thatcher, R.W.; Biver, C.L.; Gomez-Molina, J.F.; North, D.; Curtin, R.; Walker, R.W.; Salazar, A. Estimation of the EEG power spectrum by MRI T2 relaxation time in traumatic brain injury. Clin. Neurophysiol. 2001, 112, 1729–1745. [Google Scholar] [CrossRef]
- Ray, W.; Cole, H. EEG alpha activity reflects attentional demands and beta activity reflects emotional and cognitive processes. Science 1985, 228, 750–752. [Google Scholar] [CrossRef]
- Lazarev, V.V. Factorial structure of the principal EEG parameters during intellectual activity. I. Local characteristics of nonhomogeneity of functional states. Hum. Physiol. 1986, 12, 375–382, (A translation of Fiziol. Cheloveka). [Google Scholar]
- Lazarev, V.V. Factorial structure of the principal EEG parameters during intellectual activity. II. Topography of functional states. Hum. Physiol. 1987, 13, 9–12, (A translation of Fiziol. Cheloveka). [Google Scholar]
- Lazarev, V.V. On the intercorrelation of some frequency and amplitude parameters of the human EEG and its functional significance. Com. I. Multidimensional neurodynamic organization of functional states of the brain during intellectual, perceptive and motor activity in normal subjects. Int. J. Psychophysiol. 1998, 28, 77–98. [Google Scholar]
- Lazarev, V.V. On the intercorrelation of some frequency and amplitude parameters of the human EEG and its functional significance. Com. II. Neurodynamic imbalance in endogenous asthenic-like disorders. Int. J. Psychophysiol. 1998, 29, 277–289. [Google Scholar] [CrossRef]
- Mizuki, Y. Frontal lobe: Mental function and EEG. Am. J. EEG Technol. 1987, 27, 91–101. [Google Scholar] [CrossRef]
- Klimesch, W.; Schimke, H.; Ladurner, G.; Pfurtscheller, G. Alpha frequency and memory performance. J. Psychophysiol. 1990, 4, 381–390. [Google Scholar]
- Klimesch, W.; Schimke, H.; Pfurtscheller, G. Alpha frequency, cognitive load and memory performance. Brain Topogr. 1993, 5, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Harmony, T.; Fernandez, T.; Silva, J.; Bernal, J.; Díaz-Comas, L.; Reyes, A.; Marosi, E.; Rodríguez, M.; Rodríguez, M. EEG delta activity: An indicator of attention to internal processing during performance of mental tasks. Int. J. Psychophysiol. 1996, 24, 161–171. [Google Scholar] [CrossRef]
- Doppelmayr, M.; Klimesch, W.; Schwaiger, J.; Auinger, P.; Winkler, T. Theta synchronization in the human EEG and episodic retrieval. Neurosci. Lett. 1998, 257, 41–44. [Google Scholar] [CrossRef]
- Basar, E. Brain Function and Oscillations. II Vol. Integrative Brain Function. Neurophysiology and Cognitive Processes; Springer: Berlin/Heidelberg, Germany, 1999; p. 515. [Google Scholar]
- Basar, E.; Basar-Eroglu, C.; Karakas, S.; Schurmann, M. Gamma, alpha, delta, and theta oscillations govern cognitive processes. Int. J. Psychophysiol. 2001, 39, 241–248. [Google Scholar] [CrossRef]
- Basar, E.; Schurmann, M.; Sakowitz, O. The selectively distributed theta system: Functions. Int. J. Psychophysiol. 2001, 39, 197–212. [Google Scholar] [CrossRef]
- Angelakis, E.; Lubar, J.F.; Stathopoulou, S. Electroencephalographic peak alpha frequency correlates of cognitive traits. Neurosci. Lett. 2004, 371, 60–63. [Google Scholar] [CrossRef] [PubMed]
- Angelakis, E.; Lubar, J.F.; Stathopoulou, S.; Kounios, J. Peak alpha frequency: An electroencephalographic measure of cognitive preparedness. Clin. Neurophysiol. 2004, 115, 887–897. [Google Scholar] [CrossRef]
- Clark, C.R.; Veltmeyer, M.D.; Hamilton, R.J.; Simms, E.; Paul, R.; Hermens, D.; Gordon, E. Spontaneous alpha peak frequency predicts working memory performance across the age span. Int. J. Psychophysiol. 2004, 53, 1–9. [Google Scholar] [CrossRef]
- Knyazev, G.G. Motivation, emotion, and their inhibitory control mirrored in brain oscillations. Neurosci. Biobehav. Rev. 2007, 31, 377–395. [Google Scholar] [CrossRef]
- Basar, E. A review of alpha activity in integrative brain function: Fundamental physiology, sensory coding, cognition and pathology. Int. J. Psychophysiol. 2012, 86, 1–24. [Google Scholar] [CrossRef]
- Grandy, T.H.; Werkle-Bergner, M.; Chicherio, C.; Lövdén, M.; Schmiedek, F.; Lindenberger, U. Individual alpha peak frequency is related to latent factors of general cognitive abilities. NeuroImage 2013, 79, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Mierau, A.; Klimesch, W.; Lefebvre, J. State-dependent alpha peak frequency shifts: Experimental evidence, potential mechanisms and functional implications. Neuroscience 2017, 360, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Engel, A.K.; Fries, P.; Singer, W. Dynamic predictions: Oscillations and synchrony in top-down processing. Nat. Rev. Neurosci. 2001, 2, 704–716. [Google Scholar] [CrossRef]
- Buzsáki, G.; Draguhn, A. Neuronal oscillations in cortical networks. Science 2004, 304, 1926–1929. [Google Scholar] [CrossRef] [PubMed]
- Gazzaniga, M.S.; Ivry, R.B.; Mangun, G.R. Cognitive Neuroscience: The Biology of The Mind, 2nd ed.; W.W. Norton &Company: New York, NY, USA, 2002; p. 681. [Google Scholar]
- Fingelkurts, A.A.; Fingelkurts, A.A. EEG oscillatory states: Universality, uniqueness and specificity across healthy-normal, altered and pathological brain conditions. PLoS ONE 2014, 9, e87507. [Google Scholar] [CrossRef]
- John, E.R.; Karmel, B.Z.; Corning, W.C.; Easton, P.; Brown, D.; Ahn, H.; John, M.; Harmony, T.; Prichep, L.; Toro, A.; et al. Neurometrics: Numerical taxonomy identifies different profiles of brain functions within groups of behaviourally similar people. Science 1977, 196, 1393–1410. [Google Scholar] [CrossRef]
- Gevins, A. Electrophysiological imaging of brain function. In Brain Mapping. The Methods, 2nd ed.; Toga, A.W., Mazzoitta, J.C., Eds.; Elsevier Science: New York, NY, USA, 2002; pp. 175–188. [Google Scholar]
- Gebber, G.L.; Zhong, S.; Barman, S.M. The functional significance of the 10-Hz sympathetic rhythm: A hypothesis. Clin. Exp. Hypertens. 1995, 17, 181–195. [Google Scholar] [CrossRef]
- Osintseva, Y.V.; Nadezhdina, M.V.; Zhezher, M.N.; Kurus, O.S.; Skulskaya, N.I. The vegetative status and bioelectric activity of the brain in different terms of the remote period of a fighting craniocereberal trauma. Bull. Sib. Med. 2010, 4, 84–88. [Google Scholar] [CrossRef]
- Olbrich, S.; Sander, C.; Matschinger, H.; Mergl, R.; Trenner, M.; Schönknecht, P.; Hegerl, U. Brain and body. Associations between EEG-vigilance and the autonomous nervous system activity during rest. J. Psychophysiol. 2011, 25, 190–200. [Google Scholar] [CrossRef]
- Duschek, S.; Wörsching, J.; del Paso, G.A.R. Autonomic cardiovascular regulation and cortical tone. Clin. Physiol. Funct. Imaging 2014, 35, 383–392. [Google Scholar] [CrossRef]
- Langhorst, P.; Stroh-Werz, M.; Dittmar, K.; Camerer, H. Facultative coupling of reticular neuronal activity with peripheral cardiovascular and central cortical rhythms. Brain Res. 1975, 87, 407–418. [Google Scholar] [CrossRef]
- Langhorst, P.; Schulz, P.; Lambertz, M.; Schulz, G.; Camerer, H. Dynamic characteristics of the “unspecific brain stem system”. In Central Interaction between Respiratory and Cardiovascular Control System; Koepchen, H.P., Hilton, S.M., Trzebski, A., Eds.; Springer: New York, NY, USA, 1980; pp. 30–41. [Google Scholar]
- Achimowicz, J.Z. Evaluation of pilot psychophysiological state in real time by analysis of spectral dynamics in EEG and ERP correlates of sensory and cognitive brain functions and its possible coupling with autonomic nervous system. In Human System Division, Research Proposal Draft Version 10.5.; H.G. Armstrong Aero-Space Medical Research Laboratory, Wright–Petterson Air Force Base: Dayton, OH, USA, 1992. [Google Scholar]
- Jennings, J.R.; Coles, M.G.H. Handbook of Cognitive Psychophysiology, Central and Autonomic Nervous System Approaches; Wiley Psychophysiology Handbooks; Wiley: Chichester, UK, 1991; p. 762. [Google Scholar]
- Walker, B.B.; Walker, J.M. Phase relationship between cariotid pressure and ongoing electrocortical activity. Int. J. Psychophysiol. 1983, 1, 65–73. [Google Scholar] [CrossRef]
- Sarà, M.; Pistoia, F. Complexity loss in physiological time series of patients in a vegetative state. Nonlinear Dyn. Psychol. Life Sci. 2010, 14, 1–13. [Google Scholar]
- Fingelkurts, A.A.; Fingelkurts, A.A.; Bagnato, S.; Boccagni, C.; Galardi, G. Life or death: Prognostic value of a resting EEG with regards to survival in patients in vegetative and minimally conscious states. PLoS ONE 2011, 6, e25967. [Google Scholar] [CrossRef] [PubMed]
- Fingelkurts, A.A.; Fingelkurts, A.A.; Bagnato, S.; Boccagni, C.; Galardi, G. Toward operational architectonics of consciousness: Basic evidence from patients with severe cerebral injuries. Cogn. Process. 2012, 13, 111–131. [Google Scholar] [CrossRef] [PubMed]
- Sarà, M.; Sebastiano, F.; Sacco, S.; Pistoia, F.; Onorati, P.; Albertini, G.; Carolei, A. Heart rate nonlinear dynamics in patients with persistent vegetative state: A preliminary report. Brain Inj. 2008, 22, 33–37. [Google Scholar] [CrossRef]
- Wittling, W. The right hemisphere and the human stress response. Acta Physiol. Scand. Suppl. 1997, 640, 55–59. [Google Scholar]
- Hewig, J.; Schlotz, W.; Gerhards, F.; Breitenstein, C.; Lürken, A.; Naumann, E. Associations of the cortisol awakening response (CAR) with cortical activation asymmetry during the course of an exam stress period. Psychoneuroendocrinology 2008, 33, 83–91. [Google Scholar] [CrossRef]
- Tops, M.; Wijers, A.A.; van Staveren, A.S.; Bruin, K.J.; Den Boer, J.A.; Meijman, T.F.; Korf, J. Acute cortisol administration modulates EEG alpha asymmetry in volunteers: Relevance to depression. Biol. Psychol. 2005, 69, 181–193. [Google Scholar] [CrossRef]
- Buss, K.A.; Malmstadt, J.R.; Dolski, I.; Kalin, N.H.; Goldsmith, H.H.; Davidson, R.J. Right frontal brain activity, cortisol, and withdrawal behavior in 6-month-old infants. Behav. Neurosci. 2003, 117, 11–20. [Google Scholar] [CrossRef]
- Schutter, D.J.L.G.; Van Honk, J.; Koppeschaar, H.P.F.; Kahn, R.S. Cortisol and reduced interhemispheric coupling between the left prefrontal and the right parietal cortex. J. Neuropsychiatry Clin. Neurosci. 2002, 14, 89–90. [Google Scholar] [CrossRef] [PubMed]
- Birmanns, B.; Saphier, D.; Abramsky, O. a-Interferon modifies cortical EEG activity: Dose-dependence and antagonism by naloxone. J. Neurol. Sci. 1990, 100, 22–26. [Google Scholar] [CrossRef]
- Saphier, D.; Ovadia, H.; Abramsky, O. Neural responses to antigenic challenges and immunomodulatory factors. Yale J. Biol. Med. 1990, 63, 109–119. [Google Scholar]
- Kang, D.H.; Davidson, R.J.; Coe, C.L.; Wheeler, R.F.; Tomarken, A.J.; Ershler, W. Frontal brain asymmetry and immune function. Behav. Neurosci. 1991, 105, 860–869. [Google Scholar] [CrossRef] [PubMed]
- Rosenkranz, M.A.; Jackson, D.C.; Dalton, K.M.; Dolski, I.; Ryff, C.D.; Singer, B.H.; Muller, D.; Kalin, N.H.; Davidson, R.J. Affective style and in vivo immune response: Neurobehavioral mechanisms. Proc. Natl. Acad. Sci. USA 2003, 100, 11148–11152. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.-H.; Lee, J.-T. Stress and EEG. In Convergence and Hybrid Information Technologies; Crisan, M., Ed.; INTECH: Rijeka, Croatia, 2010; pp. 413–426. [Google Scholar]
- Vanhollebeke, G.; De Smet, S.; De Raedt, R.; Baeken, C.; van Mierlo, P.; Vanderhasselt, M.A. The neural correlates of psychosocial stress: A systematic review and meta-analysis of spectral analysis EEG studies. Neurobiol. Stress 2022, 18, 100452. [Google Scholar] [CrossRef] [PubMed]
- Davidson, R.J.; Coe, C.C.; Dolski, I.; Donzella, B. Individual differences in prefrontal activation asymmetry predict natural killer cell activity at rest and in response to challenge. Brain Behav. Immun. 1999, 13, 93–108. [Google Scholar] [CrossRef]
- Hecht, D. Depression and the hyperactive right-hemisphere. Neurosci. Res. 2010, 68, 77–87. [Google Scholar] [CrossRef]
- Davis, P.A. Effect on the EEG of changing the blood sugar level. Arch. Neurol. Psychiatry 1943, 49, 186–194. [Google Scholar] [CrossRef]
- Sulg, I.A.; Sotaniemi, K.A.; Tolonen, U.; Hokkanen, E. Dependence between cerebral metabolism and blood flow as reflected in the quantitative EEG. Adv. Biol. Psychiatry 1981, 6, 102–108. [Google Scholar]
- Köpruner, V.; Pfurtscheller, G.; Auer, L.M. Quantitative EEG in normals and in patients with cerebral ischemia. Prog. Brain Res. 1984, 62, 29–50. [Google Scholar] [PubMed]
- Knyazeva, M.G.; Vil’davskii, V.U. Correspondence of spectral characteristics of EEG and regional blood circulation in 9-14 years old children. Hum. Physiol. (Physiol. Cheloveka) 1986, 12, 387–394. [Google Scholar]
- Passero, S.; Rocchi, R.; Vatti, G.; Burgalassi, L.; Battistini, N. Quantitative EEG mapping, regional cerebral blood flow and neuropsychological function in Alzheimer’s disease. Dementia 1995, 6, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Kraaier, V.; van Huffelen, A.C.; Wieneke, G.H. Changes in quantitative EEG and blood flow velocity due to standardized hyperventilation; a model of transient ischaemia in young human subjects. Electroencephalogr. Clin. Neurophysiol. 1988, 70, 377–387. [Google Scholar] [CrossRef]
- Szelies, B.; Mielke, R.; Kessler, J.; Heiss, W.D. EEG power changes are related with regional cerebral glucose metbolism in vascular dementia. Clin. Neurophysiol. 1999, 110, 615–620. [Google Scholar] [CrossRef]
- Alper, K.R.; John, E.R.; Brodie, J.; Günther, W.; Daruwala, R.; Prichep, L.S. Correlation of PET and qEEG in normal subjects. Psychiatry Res. 2006, 146, 271–282. [Google Scholar] [CrossRef]
- Jann, K.; Koenig, T.; Dierks, T.; Boesch, C.; Federspiel, A. Association of individual resting state EEG alpha frequency and cerebral blood flow. NeuroImage 2010, 51, 365–372. [Google Scholar] [CrossRef]
- O’Gorman, R.L.; Poil, S.S.; Brandeis, D.; Klaver, P.; Bollmann, S.; Ghisleni, C.; Lüchinger, R.; Martin, E.; Shankaranarayanan, A.; Alsop, D.C.; et al. Coupling between resting cerebral perfusion and EEG. Brain Topogr. 2013, 26, 442–457. [Google Scholar] [CrossRef]
- Babiloni, C.; Del Percio, C.; Caroli, A.; Salvatore, E.; Nicolai, E.; Marzano, N.; Lizio, R.; Cavedo, E.; Landau, S.; Chen, K.; et al. Cortical sources of resting state EEG rhythms are related to brain hypometabolism in subjects with alzheimer’s disease: An EEG-Pet study. Neurobiol. Aging 2016, 48, 122–134. [Google Scholar] [CrossRef]
- Cohn, R.; Raines, G. Cerebral vascular lesions: Electroencephalographic and neuropathologic correlations. Arch. Neurol. Psychiatry 1948, 60, 165–181. [Google Scholar] [CrossRef]
- Ingvar, D.H.; Sjolund, B.; Ardo, A. Correlation between dominant EEG frequency, cerebral oxygen uptake and blood flow. Electroencephalogr. Clin. Neurophysiol. 1976, 41, 268–276. [Google Scholar] [CrossRef]
- Blume, W.T.; Ferguson, G.G.; McNeill, D.K. Significance of EEG changes at carotid endarterectomy. Stroke 1985, 17, 891–897. [Google Scholar] [CrossRef]
- Jonkman, E.J.; Poortvliet, D.C.J.; Veering, M.M.; De Weerd, A.W.; John, E.R. The use of neurometrics in the study of patients with cerebral ischemia. Electroencephalogr. Clin. Neurophysiol. 1985, 61, 333–341. [Google Scholar] [CrossRef]
- Nagata, K. Topographic EEG in brain ischemia: Correlation with blood flow and metabolism. Brain Topogr. 1988, 1, 97–106. [Google Scholar] [CrossRef]
- Nagata, K.; Tagwa, K.; Hiroi, S.; Shishido, F.; Uemura, K. Electroencephalographic correlates of blood flow and oxygen metabolism provided by positron emission tomography in patients with cerebral infarction. Electroencephalogr. Clin. Neurophysiol. 1989, 72, 16–30. [Google Scholar] [CrossRef]
- Claassen, J.; Hirsch, L.J.; Kreiter, K.T.; Du, E.Y.; Connolly, S.E.; Emerson, R.G.; Mayer, S.A. Quantitative continuous EEG for detecting delayed cerebral ischemia in patients with poor-grade subarachnoid hemorrhage. Clin. Neurophysiol. 2004, 115, 2699–2710. [Google Scholar] [CrossRef]
- Hughes, J.R.; John, E.R. Conventional and quantitative electroencephalography in psychiatry. J. Neuropsychiatry Clin. Neurosci. 1999, 11, 190–208. [Google Scholar] [CrossRef]
- Mueller, T.M.; Gollwitzer, S.; Hopfengärtner, R.; Rampp, S.; Lang, J.D.; Stritzelberger, J.; Madžar, D.; Reindl, C.; Sprügel, M.I.; Onugoren, M.D.; et al. Alpha power decrease in quantitative EEG detects development of cerebral infarction after subarachnoid hemorrhage early. Clin. Neurophysiol. 2021, 132, 1283–1289. [Google Scholar] [CrossRef]
- Vatinno, A.A.; Simpson, A.; Ramakrishnan, V.; Bonilha, H.S.; Bonilha, L.; Seo, N.J. The prognostic utility of electroencephalography in stroke recovery: A systematic review and meta-analysis. Neurorehabil. Neural Repair 2022, 36, 255–268. [Google Scholar] [CrossRef]
- Gollwitzer, S.; Groemer, T.; Rampp, S.; Hagge, M.; Olmes, D.; Huttner, H.B.; Schwab, S.; Madžar, D.; Hopfengaertner, R.; Hamer, H.M. Early prediction of delayed cerebral ischemia in subarachnoid hemorrhage based on quantitative EEG: A prospective study in adults. Clin. Neurophysiol. 2015, 126, 1514–1523. [Google Scholar] [CrossRef]
- Rots, M.L.; van Putten, M.J.; Hoedemaekers, C.W.; Horn, J. Continuous EEG monitoring for early detection of delayed cerebral ischemia in subarachnoid hemorrhage: A pilot study. Neurocrit. Care 2016, 24, 207–216. [Google Scholar] [CrossRef]
- Balança, B.; Dailler, F.; Boulogne, S.; Ritzenthaler, T.; Gobert, F.; Rheims, S.; Andre-Obadia, N. Diagnostic accuracy of quantitative EEG to detect delayed cerebral ischemia after subarachnoid hemorrhage: A preliminary study. Clin. Neurophysiol. 2018, 129, 1926–1936. [Google Scholar] [CrossRef]
- Vakalopoulos, C. The EEG as an index of neuromodulator balance in memory and mental illness. Front. Neurosci. 2014, 8, 63. [Google Scholar] [CrossRef] [Green Version]
- Lubar, J.F. Neocortical dynamics: Implication for understanding the role of neurofeedback and related techniques for the enhancement of attention. Appl. Psychophysiol. Biofeedback 1997, 22, 111–126. [Google Scholar] [CrossRef]
- Nistico, G.; Nappy, G. Locus coeruleus, an integrative station involved in the control of several vital functions. Funct. Neurol. 1993, 8, 5–25. [Google Scholar]
- Panyushkina, S.V.; Kurova, N.S.; Egorov, S.F.; Koshelev, V.V. Individual EEG reactions of healthy humans to mutually antagonistic noradrenotropic influences. Zh Vyss. Nerv Deyat 1994, 44, 457–469. [Google Scholar]
- Sadato, N.; Nakamura, S.; Oohashi, T. Neural networks for generation and suppression of alpha rhythm: A PET study. NeuroReport 1998, 9, 893–897. [Google Scholar] [CrossRef]
- Chavanon, M.-L.; Wacker, J.; Stemmler, G. Paradoxical dopaminergic drug effects in extraversion: Dose- and time-dependent effects of sulpiride on EEG theta activity. Front. Hum. Neurosci. 2013, 7, 117. [Google Scholar] [CrossRef]
- Knott, V.J.; Hovson, A.L.; Perugimi, M. The effect of acute tryptophan depletion and fenfluramine on quantitative EEG and mood in healthy male subjects. Biol. Psychiatry 1999, 46, 229–238. [Google Scholar] [CrossRef]
- Steriade, M.; Gloor, P.; Llinas, R.R.; da Silva, F.H.L.; Mesulam, M.-M. Basic mechanisms of cerebral rhythmic activities. Report of IFCN Committee on Basic Mechanisms. Electroencepahlogr. Clin. Neurophysiol. 1990, 76, 481–508. [Google Scholar] [CrossRef]
- John, E.R.; Prichep, L.S.; Winterer, G.; Herrmann, W.M.; diMichele, F.; Halper, J.; Bolwig, T.G.; Cancro, R. Electrophysiological subtypes of psychotic states. Acta Psychiatr. Scand. 2007, 116, 17–35. [Google Scholar] [CrossRef]
- Small, J.G. Psychiatric disorders and EEG. In Electroencephalography: Basic Principles, Clinical Applications, and Related Fields; Niedermeyer, E., da Silva, F.L., Eds.; Williams and Wilkins: Baltimore, MD, USA, 1993; pp. 581–596. [Google Scholar]
- Hughes, J.R. The EEG in psychiatry: An outline with summarized points and references. Clin. Electroencephalogr. 1995, 26, 92–101. [Google Scholar] [CrossRef]
- Sam, M.C.; So, E.L. Significance of epileptiform discharges in patients without epilepsy in the community. Epilepsia 2001, 42, 1273–1278. [Google Scholar] [CrossRef]
- Zivin, L.; Marsan, C.A. Incidence and prognostic significance of “epileptiform” activity in the EEG of nonepileptic subjects. Brain 1968, 91, 751–777. [Google Scholar] [CrossRef]
- Standage, K.F. The etiology of hysterical seizures. Can. Psychiatr. Assoc. J. 1975, 20, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Cohen, R.J.; Suter, C. Hysterical seizures: Suggestion as a provocative EEG test. Ann. Neurol. 1982, 11, 391–395. [Google Scholar] [CrossRef]
- King, D.W.; Gallagher, B.B.; Murvin, A.J.; Smith, D.B.; Marcus, D.J.; Hartlage, L.C.; Ward, L.C., 3rd. Pseudoseizures: Diagnostic evaluation. Neurology 1982, 32, 18–23. [Google Scholar] [CrossRef]
- Luther, J.S.; McNamara, J.O.; Carwile, S.; Miller, P.; Hope, V. Pseudoepileptic seizures: Methods and video analysis to aid diagnosis. Ann. Neurol. 1982, 12, 458–462. [Google Scholar] [CrossRef]
- Wilkus, R.J.; Dodrill, C.B.; Thompson, P.M. Intensive EEG monitoring and psychological studies of patients with pseudoepileptic seizures. Epilepsia 1984, 25, 100–107. [Google Scholar] [CrossRef]
- Wilkes, R.J.; Thompson, P.M.; Vossler, D.G. Bizarre ictal automatisms: Frontal lobe epileptic or psychogenic seizures. J. Epilepsy 1990, 3, 297–313. [Google Scholar] [CrossRef]
- Lelliott, P.T.; Fenwick, P. Cerebral pathology in pseudoseizures. Acta Neurol. Scand. 1991, 83, 29–132. [Google Scholar] [CrossRef]
- Bowman, E.S. Etiology and clinical course of pseudoseizures: Relationship to trauma, depression, and dissociation. Psychosomatics 1993, 34, 333–342. [Google Scholar] [CrossRef]
- Devinsky, O.; Sanchez-Villasenor, F.; Vazquez, B.; Kothari, M.; Alper, K.; Luciano, D. Clinical profile of patients with epileptic and nonepileptic seizures. Neurology 1996, 46, 1530–1533. [Google Scholar] [CrossRef] [PubMed]
- Shelley, B.P.; Trimble, M.R.; Boutros, N.N. Electroencephalographic cerebral dysrhythmic abnormalities in the trinity of nonepileptic general population, neuropsychiatric, and neurobehavioral disorders. J. Neuropsychiatry Clin. Neurosci. 2008, 20, 7–22. [Google Scholar] [CrossRef]
- Pillmann, F.; Rohde, A.; Ullrich, S.; Draba, S.; Sannemüller, U.; Marneros, A. Violence, criminal behavior, and the EEG: Significance of left hemispheric focal abnormalities. J. Neuropsychiatry Clin. Neurosci. 1999, 11, 454–457. [Google Scholar] [CrossRef]
- Hughes, J.; Leander, R.; Ketchum, G. Electroencephalographic study of specific reading disabilities. EEG. Clin. Neurophysiol. 1949, 1, 377. [Google Scholar]
- Ribas, J.C.; Baptistete, E.; Fonseca, C.A.; Tiba, I.; Filho, H.S.C. Behavior disorders with predominance of aggressiveness, irritability, impulsiveness, and instability: Clinical electroencephalographic study of 100 cases. Arq. De Neuro-Psiquiatr. 1974, 32, 187–194. [Google Scholar] [CrossRef]
- Harty, J.E.; Gibbs, E.L.; Gibbs, F.A. Electroencephalographic study of two hundred and seventy-five candidates for military service. War Med. 1942, 2, 923–930. [Google Scholar]
- Socanski, D.; Herigstad, A.; Thomsen, P.H.; Dag, A.; Larsen, T.K. Epileptiform abnormalities in children diagnosed with attention deficit/hyperactivity disorder. Epilepsy Behav. 2010, 19, 483–486. [Google Scholar] [CrossRef]
- Dierks, T.; Ihl, R.; Frolich, L.; Maurer, K. Dementia of the Alzheimer type: Effects on the spontaneous EEG described by dipole sources. Psychiatry Res. 1993, 50, 51–162. [Google Scholar] [CrossRef]
- Prichep, L.S.; Mas, F.; Hollander, E.; Liebowitz, M.; John, E.R.; Almas, M.; DeCaria, C.M.; Levine, R.H. Quantitative electroencephalographic (QEEG) subtyping of obsessive-compulsive disorder. Psychiatry Res. 1993, 50, 25–32. [Google Scholar] [CrossRef]
- Inui, K.; Motomura, E.; Okushima, R.; Kaige, H.; Inoue, K.; Nomura, J. Electroencephalographic findings in patients with DSM-IV mood disorder, schizophrenia, and other psychotic disorders. Biol. Psychiatry 1998, 43, 69–75. [Google Scholar] [CrossRef]
- Huang, C.; Wahlund, L.O.; Dierks, T.; Julin, P.; Winblad, B.; Jelic, V. Discrimination of Alzheimer’s disease and mild cognitive impairment by equivalent EEG sources: A cross-sectional and longitudinal study. Clin. Neurophysiol. 2000, 11, 1961–1967. [Google Scholar] [CrossRef]
- Monastra, V.J.; Lubar, J.F.; Linden, M. The development of a quantitative electroencephalographic scanning process for attention deficit-hyperactivity disorder: Reliability and validity studies. Neuropsychology 2001, 15, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Thatcher, R.W.; North, D.M.; Curtin, R.T.; Walker, R.A.; Biver, C.J.; Gomez, J.F.; Salazar, A.M. An EEG severity index of traumatic brain injury. J. Neuropsychiatry Clin. Neurosci. 2001, 13, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Karadag, F.; Oguzhanoglu, N.K.; Kurt, T.; Oguzhanoglu, A.; Atesci, F.; Ozdel, O. Quantitative EEG analysis in obsessive compulsive disorder. Int. J. Neurosci. 2003, 113, 833–847. [Google Scholar] [CrossRef]
- Boutros, N.N.; Torello, M.; McGlashan, T.H. Electrophysiological aberrations in borderline personality disorder: State of the evidence. J. Neuropsychiatry Clin. Neurosci. 2003, 15, 145–154. [Google Scholar] [CrossRef]
- Rowe, D.L. Biophysical modeling of tonic cortical electrical activity in attention deficit hyperactivity disorder. Int. J. Neurosci. 2005, 115, 1273–1305. [Google Scholar] [CrossRef]
- Babiloni, C.; Benussi, L.; Binetti, G.; Cassetta, E.; Dal Forno, G.; Del Percio, C.; Ferreri, F.; Ferri, R.; Frisoni, G.; Ghidoni, R.; et al. Apolipoprotein E and alpha brain rhythms in mild cognitive impairment: A multicentric electroencephalogram study. Ann. Neurol. 2006, 59, 323–334. [Google Scholar] [CrossRef]
- Başar, E.; Güntekin, B. A review of brain oscillations in cognitive disorders and the role of neurotransmitters. Brain Res. 2008, 1235, 172–193. [Google Scholar] [CrossRef]
- Fingelkurts, A.A.; Fingelkurts, A.A. Alpha rhythm operational architectonics in the continuum of normal and pathological brain states: Current state of research. Int. J. Psychophysiol. 2010, 76, 93–106. [Google Scholar] [CrossRef] [PubMed]
- Schultz, E.V.; Baburin, I.N.; Karavaeva, T.A.; Karvasarsky, B.D.; Slezin, V.B. Bioelectric brain activity in patients with neurotic and neurosis-like disorders (according to a spectral analysis). Bekhterev. Rev. Psychiatry Med. Psychol. 2010, 3, 26–31. [Google Scholar]
- Lee, S.M.; Jang, K.-I.; Chae, G.-H. Electroencephalographic correlates of suicidal ideation in the theta band. Clin. EEG Neurosci. 2017, 48, 316–321. [Google Scholar] [CrossRef]
- Kanda, P.A.M.; Anghinah, R.; Smidth, M.T.; Silva, J.M. The clinical use of quantitative EEG in cognitive disorders. Dement. Neuropsychol. 2009, 3, 195–203. [Google Scholar] [CrossRef] [PubMed]
- John, E.R. The role of quantitative EEG topographic mapping or ‘neurometrics’ in the diagnosis of psychiatric and neurological disorders: The pros. Electroencephalogr. Clin. Neurophysiol. 1989, 73, 2–4. [Google Scholar] [CrossRef]
- Abrams, R.; Taylor, M.A. Differential EEG patterns in affective disorder and schizophrenia. Arch. Gen. Psychiatry 1979, 36, 1355–1358. [Google Scholar] [CrossRef]
- Giannitrapani, D.; Collins, J. EEG differentiation between Alzheimer’s and non-Alzheimer’s dementias. In The EEG of Mental Activities; Giannitrapani, D., Murri, L., Eds.; Karger: New York, NY, USA, 1988; pp. 26–41. [Google Scholar]
- Goodin, D.S.; Aminoff, M.J. Electrophysiological differences between subtypes of dementia. Brain 1986, 109, 1102–1113. [Google Scholar] [CrossRef]
- Chabot, R.J.; Serfontein, G. Quantitative EEG profiles of children with attention deficit disorder. Biol. Psychiatry 1996, 40, 951–963. [Google Scholar] [CrossRef]
- John, E.R.; Prichep, L.; Ahn, H.; Easton, P.; Fridman, J.; Kaye, H. Neurometric evaluation of cognitive dysfunctions and neurological disorders in children. Prog. Neurobiol. 1983, 21, 239–290. [Google Scholar] [CrossRef]
- Coburn, K.L.; Lauterbach, E.C.; Boutros, N.N.; Black, K.J.; Arciniegas, D.B.; Coffey, C.E. The value of quantitative electroencephalography in clinical psychiatry: A report by the Committee on Research of the American Neuropsychiatric Association. J. Neuropsychiatry Clin. Neurosci. 2006, 18, 460–500. [Google Scholar] [CrossRef]
- Knott, V.; Bakish, D.; Lusk, S.; Barkely, J.; Perugini, M. Quantitative EEG correlates of panic disorder. Psychiatry Res. 1996, 68, 31–39. [Google Scholar] [CrossRef]
- Deslandes, A.; Veiga, H.; Cagy, M.; Fiszman, A.; Piedade, R.; Ribeiro, P. Quantitative electroencephalography (qEEG) to discriminate primary degenerative dementia from major depression disorder (depression). Arq. Neuropsiquiatr. 2004, 62, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Pucci, E.; Belardinelli, N.; Cacchiò, G.; Signorino, M.; Angeleri, F. EEG power spectrum differences in early and late onset forms of Alzheimer’s disease. Clin. Neurophysiol. 1999, 110, 621–631. [Google Scholar] [CrossRef]
- Dierks, T.; Perisic, I.; Frölich, L.; Ihl, R.; Maurer, K. Topography of the qEEG in dementia of Alzheimer type: Relation to severity of dementia. Psychol. Res. 1991, 40, 181–194. [Google Scholar]
- Prichep, L.S.; John, E.R.; Ferris, S.H.; Rausch, L.; Fang, Z.; Cancro, R.; Torossian, C.; Reisberg, B. Prediction of longitudinal cognitive decline in normal elderly with subjective complaints using electrophysiological imaging. Neurobiol. Aging 2006, 27, 471–481. [Google Scholar] [CrossRef]
- Bauer, L.O. Predicting relapse to alcohol and drug abuse via quantitative electroencephalography. Neuropsychopharmacology 2001, 25, 332–340. [Google Scholar] [CrossRef]
- Naunheim, R.S.; Treaster, M.; English, J.; Casner, T.; Chabot, R. Use of brain electrical activity to quantify traumatic brain injury in the emergency department. Brain Inj. 2010, 24, 1324–1329. [Google Scholar] [CrossRef]
- Ritchlin, C.T.; Chabot, R.J.; Alper, K.; Buyon, J.; Belmont, H.M.; Roubey, R.; Abramson, S.B. Quantitative electroencephalography: A new approach to the diagnosis of cerebral dysfunction in systemic lupus erythematosus. Arth. Rheumat. 1992, 35, 1330–1342. [Google Scholar] [CrossRef]
- Sloan, E.P.; Fenton, G.W.; Kennedy, J.S.J.; MacLennan, J.M. Electroencephalography and single photon emission computed tomography in dementia: A comparative study. Psychol. Med. 1995, 25, 631–638. [Google Scholar] [CrossRef]
- Kropotov, J.D.; Pąchalska, M.; Mueller, A. New neurotechnologies for the diagnosis and modulation of brain dysfunctions. Health Psychol. Rep. 2014, 2, 73–82. [Google Scholar] [CrossRef]
- Kropotov, J.D.; Müller, A.; Candrian, G.; Valery, P. Neurobiology of ADHD: A New Diagnostic Approach Based on Electrophysiological Endophenotypes; Springer: London, UK, 2013; p. 300. [Google Scholar]
- Fisher, N.K.; Talathi, S.S.; Cadotte, A.; Carney, P.R. Epilepsy detection and monitoring. In Quantitative EEG Analysis Methods and Clinical Applications; Tong, S., Thakor, N.V., Eds.; Artech House: Norwood, MA, USA, 2009; pp. 141–167. [Google Scholar]
- Pardalos, P.M. Seizure warning algorithm based on optimization and nonlinear dynamics. Math. Program 2004, 101, 365–385. [Google Scholar] [CrossRef]
- Drislane, F.W. The clinical use of ambulatory EEG. In Atlas of Ambulatory EEG; Chang, B.S., Schachter, S.C., Schomer, D.L., Eds.; Elsevier: Amsterdam, The Netherlands, 2005; pp. 17–25. [Google Scholar]
- Hegerl, U.; Hensch, T. The vigilance regulation model of affective disorders and ADHD. Neurosci. Biobehav. Rev. 2014, 44, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Wittekind, D.A.; Spada, J.; Gross, A.; Hensch, T.; Jawinski, P.; Ulke, C.; Sander, C.; Hegerl, U. Early report on brain arousal regulation in manic vs. depressive episodes in bipolar disorder. Bipolar Disord. 2016, 18, 502–510. [Google Scholar] [CrossRef]
- Brenner, R.P. EEG and dementia, Chapter 19. In Electroencephalography, Basic Principles, Clinical Applications, and Related Fields, 4th ed.; Niedermeyer, E., da Silva, F.L., Eds.; Williams and Wilkins: Baltimore, MD, USA, 1999; pp. 349–359. [Google Scholar]
- Fingelkurts, A.A.; Fingelkurts, A.A.; Bagnato, S.; Boccagni, C.; Galardi, G. EEG oscillatory states as neuro-phenomenology of consciousness as revealed from patients in vegetative and minimally conscious states. Conscious Cogn. 2012, 21, 149–169. [Google Scholar] [CrossRef]
- Fingelkurts, A.A.; Fingelkurts, A.A.; Bagnato, S.; Boccagni, C.; Galardi, G. Dissociation of vegetative and minimally conscious patients based on brain operational architectonics: Factor of etiology. Clin. EEG Neurosci. 2013, 44, 209–220. [Google Scholar] [CrossRef]
- Fingelkurts, A.A.; Fingelkurts, A.A.; Bagnato, S.; Boccagni, C.; Galardi, G. Prognostic value of resting-state electroencephalography structure in disentangling vegetative and minimally conscious states: A preliminary study. Neurorehabil. Neural Repair 2013, 27, 345–354. [Google Scholar] [CrossRef]
- Fingelkurts, A.A.; Fingelkurts, A.A.; Bagnato, S.; Boccagni, C.; Galardi, G. The value of spontaneous EEG oscillations in distinguishing patients in vegetative and minimally conscious states, chapter 5. In Application of Brain Oscillations in Neuropsychiatric Diseases (Supplements to Clinical Neurophysiology); Basar, E., Basar-Eroglu, C., Ozerdem, A., Rossini, P.M., Yener, G.G., Eds.; Elsevier B.V.: Amsterdam, The Netherlands, 2013; Volume 62, pp. 81–99. [Google Scholar]
- Fingelkurts, A.A.; Fingelkurts, A.A. Brain space and time in mental disorders: Paradigm shift in biological psychiatry. Int. J. Psychiatry Med. 2019, 54, 53–63. [Google Scholar] [CrossRef]
- Dittrich, A. The standardized psychometric assessment of altered states of consciousness (ASCs) in humans. Pharmacopsychiatry 1998, 31, 80–84. [Google Scholar] [CrossRef]
- Parnas, J.; Møller, P.; Kircher, T.; Thalbitzer, J.; Jansson, L.; Handest, P.; Zahavi, D. EASE: Examination of anomalous self-experience. Psychopathology 2005, 38, 236–258. [Google Scholar] [CrossRef]
- Beck, A.T. The evolution of the cognitive model of depression and its neurobiological correlates. Am. J. Psychiatry 2008, 165, 969–977. [Google Scholar] [CrossRef]
- Musholt, K. Thinking about Oneself: From Nonconceptual Content to the Concept of a Self; MIT Press: Cambridge, UK, 2015; p. 232. [Google Scholar]
- Northoff, G.; Heinzel, A.; de Greck, M.; Bermpohl, F.; Dobrowolny, H.; Panksepp, J. Self-referential processing in our brain. A meta-analysis of imaging studies on the self. NeuroImage 2006, 31, 440–457. [Google Scholar] [CrossRef] [PubMed]
- Northoff, G. Is the self a higher-order or fundamental function of the brain? The ‘basis model of self-specificity’ and its encoding by the brain’s spontaneous activity. Cogn. Neurosci. 2016, 7, 203–222. [Google Scholar] [CrossRef] [PubMed]
- Raichle, M.E.; MacLeod, A.M.; Snyder, A.Z.; Powers, W.J.; Gusnard, D.A.; Shulman, G.L. A default mode of brain function. Proc. Natl. Acad. Sci. USA 2001, 98, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Gusnard, D.A. Being a self: Considerations from functional imaging. Conscious Cogn. 2005, 14, 679–697. [Google Scholar] [CrossRef]
- Schilbach, L.; Eickhoff, S.B.; Rotarska-Jagiela, A.; Fink, G.R.; Vogeley, K. Minds at rest? Social cognition as the default mode of cognizing and its putative relationship to the “default system” of the brain. Conscious Cogn. 2008, 17, 457–467. [Google Scholar] [CrossRef]
- Fingelkurts, A.A.; Fingelkurts, A.A. Persistent operational synchrony within brain default-mode network and self-processing operations in healthy subjects. Brain Cogn. 2011, 75, 79–90. [Google Scholar] [CrossRef]
- Fingelkurts, A.A.; Fingelkurts, A.A.; Bagnato, S.; Boccagni, C.; Galardi, G. DMN Operational Synchrony Relates to Self-Consciousness: Evidence from Patients in Vegetative and Minimally Conscious States. Open Neuroimag. J. 2012, 6, 55–68. [Google Scholar] [CrossRef]
- Laufs, H.; Kleinschmidt, A.; Beyerle, A.; Eger, E.; Salek-Haddadi, A.; Preibisch, C.; Krakow, K. EEG-correlated fMRI of human alpha activity. Neuroimage 2003, 19, 1463–1476. [Google Scholar] [CrossRef]
- Mantini, D.; Perrucci, M.G.; Del Gratta, C.; Romani, G.L.; Corbetta, M. Electrophysiological signatures of resting state networks in the human brain. Proc. Natl. Acad. Sci. USA 2007, 104, 13170–13175. [Google Scholar] [CrossRef] [Green Version]
- Jann, K.; Dierks, T.; Boesch, C.; Kottlow, M.; Strik, W.; Koenig, T. BOLD correlates of EEG alpha phase-locking and the fMRI default mode network. Neuroimage 2009, 45, 903–916. [Google Scholar] [CrossRef]
- Knyazev, G.G.; Slobodskoj-Plusnin, J.Y.; Bocharov, A.V.; Pylkova, L.V. The default mode network and EEG α oscillations: An independent component analysis. Brain Res. 2011, 1402, 67–79. [Google Scholar] [CrossRef] [PubMed]
- Knyazev, G.G.; Savostyanov, A.N.; Volf, N.V.; Liou, M.; Bocharov, A.V. EEG correlates of spontaneous self-referential thoughts: A cross-cultural study. Int. J. Psychophysiol. 2012, 86, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Fingelkurts, A.A.; Fingelkurts, A.A.; Kallio-Tamminen, T. Long-term meditation training induced changes in the operational synchrony of default mode network modules during a resting state. Cogn. Process. 2016, 17, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Fingelkurts, A.A.; Fingelkurts, A.A.; Kallio-Tamminen, T. Trait lasting alteration of the brain default mode network in experienced meditators and the experiential selfhood. Self Identity 2016, 15, 381–393. [Google Scholar] [CrossRef]
- Fingelkurts, A.A.; Fingelkurts, A.A.; Kallio-Tamminen, T. Selfhood triumvirate: From phenomenology to brain activity and back again. Conscious Cogn. 2020, 86, 103031. [Google Scholar] [CrossRef]
- Gallagher, S. A pattern theory of self. Front. Hum. Neurosci. 2013, 7, 443. [Google Scholar] [CrossRef]
- Gallagher, S.; Daly, A. Dynamical relations in the self-pattern. Front. Psychol. 2018, 9, 664. [Google Scholar] [CrossRef]
- Fingelkurts, A.A.; Fingelkurts, A.A.; Kallio-Tamminen, T. Self, Me and I in the repertoire of spontaneously occurring altered states of Selfhood: Eight neurophenomenological case study reports. Cogn. Neurodyn. 2022, 16, 255–282. [Google Scholar] [CrossRef]
- Fingelkurts, A.A.; Fingelkurts, A.A. Longitudinal dynamics of 3-dimensional components of selfhood after severe traumatic brain injury: A qEEG case study. Clin. EEG Neurosci. 2017, 48, 327–337. [Google Scholar] [CrossRef]
- Fingelkurts, A.A.; Fingelkurts, A.A. Three-dimensional components of selfhood in treatment-naive patients with major depressive disorder: A resting-state qEEG imaging study. Neuropsychologia 2017, 99, 30–36. [Google Scholar] [CrossRef]
- Fingelkurts, A.A.; Fingelkurts, A.A. Alterations in the three components of selfhood in persons with post-traumatic stress disorder symptoms: A pilot qEEG neuroimaging study. Open Neuroimag. J. 2018, 12, 42–54. [Google Scholar] [CrossRef] [PubMed]
- Beck, A.T. Cognitive models of depression. J. Cogn. Psychother. 1987, 1, 5–37. [Google Scholar]
- Damasio, A.R. The Feeling of What Happens: Body and Emotion in the Making of Consciousness; Harcourt Brace: San Diego, CA, USA, 1999; p. 400. [Google Scholar]
- Rimes, K.A.; Watkins, E. The effects of self-focused rumination on global negative self-judgements in depression. Behav. Res. Ther. 2005, 43, 1673–1681. [Google Scholar] [CrossRef]
- Northoff, G. Psychopathology and pathophysiology of the self in depression-neuropsychiatric hypothesis. J. Affect. Disord. 2007, 104, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Nolen-Hoeksema, S.; Wisco, B.E.; Lyubomirsky, S. Rethinking rumination. Perspect. Psychol. Sci. 2008, 3, 400–424. [Google Scholar] [CrossRef] [PubMed]
- Paulus, M.P.; Stein, M.B. Interoception in anxiety and depression. Brain Struct. Funct. 2010, 214, 451–463. [Google Scholar]
- Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; The American Psychiatric Association: Arlington, VA, USA, 2013.
- Zepinic, V. Understanding and Treating Complex Trauma; Xlibris: London, UK, 2011. [Google Scholar]
- van der Kolk, B.A. The body keeps the score: Memory and the evolving psychobiology of posttraumatic stress. Harv. Rev. Psychiatry 1994, 1, 253–265. [Google Scholar] [CrossRef]
- McNally, R.J. Remembering Trauma; Belknap Press/Harvard University Press: Cambridge, MA, USA, 2003; p. 448. [Google Scholar]
- Ataria, Y. Traumatic memories as black holes: A qualitative-phenomenological approach. Qual. Psychol. 2014, 1, 123–140. [Google Scholar] [CrossRef]
- van der Kolk, B.A.; Fisler, R. Dissociation and the Fragmentary Nature of Traumatic Memories: Overview and Exploratory Study. 1995. Available online: http://www.trauma-pages.com/a/vanderk2.php (accessed on 1 May 2022).
- Kullberg-Turtiainen, M.; Vuorela, K.; Huttula, L.; Turtiainen, P.; Koskinen, S. Individualized goal directed dance rehabilitation in chronic state of severe traumatic brain injury: A case study. Heliyon 2019, 5, e01184. [Google Scholar] [CrossRef]
- Fingelkurts, A.A.; Fingelkurts, A.A.; Bagnato, S.; Boccagni, C.; Galardi, G. The chief role of frontal operational module of the brain default mode network in the potential recovery of consciousness from the vegetative state: A preliminary comparison of three case reports. Open Neuroimag. J. 2016, 10 (Suppl. S1, M4), 41–51. [Google Scholar] [CrossRef]
- Laureys, S.; Celesia, G.G.; Cohadon, F.; Lavrijsen, J.; León-Carrión, J.; Sannita, W.G.; Sazbon, L.; Schmutzhard, E.; von Wild, K.R.; Zeman, A.; et al. European Task Force on disorders of consciousness, unresponsive wakefulness syndrome: A new name for the vegetative state or apallic syndrome. BMC Med. 2010, 8, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterson, A.; Bayne, T. Post-comatose disorders of consciousness. In The Routledge Handbook of Consciousness; Gennaro, R., Ed.; Routledge: Abingdon, UK, 2018; pp. 351–365. [Google Scholar]
- Jennett, B.; Plum, F. Persistent vegetative state after brain damage. A syndrome in search of a name. Lancet 1972, 1, 734–737. [Google Scholar] [CrossRef]
- Giacino, J.T.; Ashwal, S.; Childs, N.; Cranford, R.; Jennett, B.; Katz, D.I.; Kelly, J.P.; Rosenberg, J.H.; Whyte, J.; Zafonte, R.D.; et al. The minimally conscious state: Definition and diagnostic criteria. Neurology 2002, 58, 349–353. [Google Scholar] [CrossRef] [PubMed]
- Naccache, L. Minimally conscious state or cortically mediated state? Brain 2018, 141, 949–960. [Google Scholar] [CrossRef]
- Bagnato, S.; Boccagni, C.; Sant’Angelo, A.; Fingelkurts, A.A.; Fingelkurts, A.A.; Galardi, G. Emerging from an unresponsive wakefulness syndrome: Brain plasticity has to cross a threshold level. Neurosci. Biobehav. Rev. 2013, 37, 2721–2736. [Google Scholar] [CrossRef]
- Porcaro, C.; Nemirovsky, I.E.; Riganello, F.; Mansour, Z.; Cerasa, A.; Tonin, P.; Stojanoski, B.; Soddu, A. Diagnostic developments in differentiating unresponsive wakefulness syndrome and the minimally conscious state. Front. Neurol. 2022, 12, 778951. [Google Scholar] [CrossRef]
- Bagnato, S.; Boccagni, C.; Prestandrea, C.; Sant’Angelo, A.; Castiglione, A.; Galardi, G. Prognostic value of standard EEG in traumatic and non-traumatic disorders of consciousness following coma. Clin. Neurophysiol. 2010, 121, 274–280. [Google Scholar] [CrossRef]
- Brenner, R.P. The interpretation of the EEG of stupor and coma. Neurologist 2005, 11, 271–284. [Google Scholar] [CrossRef]
- Gosseries, O.; Schnakers, C.; Ledoux, D.; Vanhaudenhuyse, A.; Bruno, M.A.; Demertzi, A.; Noirhomme, Q.; Lehembre, R.; Damas, P.; Goldman, S.; et al. Automated EEG entropy measurements in coma, vegetative state/unresponsive wakefulness syndrome and minimally conscious state. Funct. Neurol. 2011, 26, 25–30. [Google Scholar]
- Lehembre, R.; Gosseries, O.; Lugo, Z.; Jedidi, Z.; Chatelle, C.; Sadzot, B.; Laureys, S.; Noirhomme, Q. Electrophysiological investigations of brain function in coma, vegetative and minimally conscious patients. Arch. Ital. Biol. 2012, 150, 122–139. [Google Scholar] [CrossRef]
- Fingelkurts, A.A.; Fingelkurts, A.A.; Bagnato, S.; Boccagni, C.; Galardi, G. Long-term (six years) clinical outcome discrimination of patients in the vegetative state could be achieved based on the operational architectonics EEG analysis: A pilot feasibility study. Open Neuroimag. J. 2016, 10 (Suppl. S1, M6), 69–79. [Google Scholar] [CrossRef] [PubMed]
- Sarà, M.; Pistoia, F.; Pasqualetti, P.; Sebastiano, F.; Onorati, P.; Rossini, P.M. Functional isolation within the cerebral cortex in the vegetative state: A nonlinear method to predict clinical outcomes. Neurorehabil. Neural Repair 2011, 25, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Fingelkurts, A.A.; Fingelkurts, A.A. Operational architectonics methodology for EEG analysis: Theory and results. Neuromethods 2015, 91, 1–59. [Google Scholar] [CrossRef]
- Fingelkurts, A.A.; Fingelkurts, A.A.; Neves, C.F.H. Natural world physical, brain operational, and mind phenomenal space–time. Phys. Life Rev. 2010, 7, 195–249. [Google Scholar] [CrossRef]
- Cacciola, A.; Naro, A.; Milardi, D.; Bramanti, A.; Malatacca, L.; Spitaleri, M.; Leo, A.; Muscoloni, A.; Cannistraci, C.V.; Bramanti, P.; et al. Functional brain network topology discriminates between patients with minimally conscious state and unresponsive wakefulness syndrome. J. Clin. Med. 2019, 8, 306. [Google Scholar] [CrossRef] [PubMed]
- Tononi, G.; Boly, M.; Massimini, M.; Koch, C. Integrated information theory: From consciousness to its physical substrate. Nat. Rev. Neurosci. 2016, 17, 450–461. [Google Scholar] [CrossRef]
- Hubbard, O.; Sunde, D.; Goldensohn, E.S. The EEG in centenarians. Electroencephalogr. Clin. Neurophysiol. 1976, 40, 407–417. [Google Scholar] [CrossRef]
- Matejcek, M. Some relationships between occipital EEG activity and age. A spectral analytic study. Rev. Electroencephalogr. Neurophysiol. Clin. 1980, 10, 122–130. [Google Scholar] [CrossRef]
- Matthis, P.; Scheffner, D.; Benninger, C.; Lipinski, C.; Stolzis, L. Changes in the background activity of the electroencephalogram according to age. Electroencephalogr. Clin. Neurophysiol. 1980, 49, 626–635. [Google Scholar] [CrossRef]
- Marciani, M.G.; Maschio, M.; Spanedda, F.; Caltagirone, C.; Gigli, G.; Bernardi, G. Quantitative EEG evaluation in normal elderly subjects during mental processes: Age-related changes. Int. J. Neurosci. 1994, 76, 131–140. [Google Scholar] [CrossRef]
- Shigeta, M.; Julin, P.; Almkvist, O.; Basun, H.; Rudberg, U.; Wahlund, L.-O. EEG in successful ageing; a 5-year follow-up study from the eighth to ninth decade of life. Electroencephalogr. Clin. Neurophysiol. 1995, 95, 77–83. [Google Scholar] [CrossRef]
- Li, D.; Sun, F.; Jiao, Y. Frontal EEG characters in ageing and the correlativity with some cognitive abilities. Acta Psychol. Sin. 1996, 28, 76–81. [Google Scholar]
- Widagdo, M.; Pierson, J.; Helme, R. Age-related changes in qEEG during cognitive tasks. Int. J. Neurosci. 1998, 95, 63–75. [Google Scholar] [CrossRef]
- Van Sweden, B.; Wauquier, A.; Niedermeyer, E. Normal ageing and transient cognitive disorders in the elderly. In Electroencephalography: Basic Principles, Clinical Applications, and Related Fields; Niedermeyer, E., da Silva, F.H.L., Eds.; Williams and Wilkins: Baltimore, MD, USA, 1999; pp. 340–348. [Google Scholar]
- Kikuchi, M.; Wada, Y.; Koshino, Y.; Nanbu, Y.; Hashimoto, T. Effect of normal ageing upon interhemispheric EEG coherence: Analysis during rest and photic stimulation. Clin. Electroencephalogr. 2000, 31, 170–174. [Google Scholar] [CrossRef] [PubMed]
- Babiloni, C.; Binetti, G.; Cassarino, A.; Dal Forno, G.; Del Percio, C.; Ferreri, F.; Ferri, R.; Frisoni, G.; Galderisi, S.; Hirata, K.; et al. Sources of cortical rhythms in adults during physiological ageing: A multicentric EEG study. Hum. Brain Mapp. 2006, 27, 162–172. [Google Scholar] [CrossRef]
- Boha, R.; Stam, C.J.; Molnár, M. Age-dependent features of EEG-reactivity-spectral, complexity, and network characteristics. Neurosci. Lett. 2010, 479, 79–84. [Google Scholar]
- Peltz, C.B.; Kim, H.L.; Kawas, C.H. Abnormal EEGs in cognitively and physically healthy oldest-old: Findings from the 90þ study. J. Clin. Neurophysiol. 2010, 27, 292–295. [Google Scholar] [CrossRef]
- Knyazeva, M.G.; Barzegaran, E.; Vildavski, V.Y.; Demonet, J.-F. Ageing of human alpha rhythm. Neurobiol. Ageing 2018, 69, 261–273. [Google Scholar]
- Markand, O.N. Electroencephalogram in dementia. Am. J. EEG Technol. 1986, 26, 3–17. [Google Scholar] [CrossRef]
- Samson-Dollfus, D.; Delapierre, G.; Do Marcolino, C.; Blondeau, C. Normal and pathological changes in alpha rhythms. Int. J. Psychophysiol. 1997, 26, 395–409. [Google Scholar] [CrossRef]
- Fernández, A.; Hornero, R.; Mayo, A.; Poza, J.; Gil-Gregorio, P.; Ortiz, T. EEG spectral profile in Alzheimer’s disease and mild cognitive impairment. Clin. Neurophysiol. 2006, 117, 306–314. [Google Scholar] [CrossRef] [PubMed]
- Varela, F. Neurophenomenology: A methodological remedy for the hard problem. J. Conscious Stud. 1996, 3, 330–349. [Google Scholar]
- Borjigin, J.; Lee, U.; Liu, T.; Pal, D.; Huff, S.; Klarr, D.; Sloboda, J.; Hernandez, J.; Wang, M.M.; Mashour, G.A. Surge of neurophysiological coherence and connectivity in the dying brain. Proc. Natl. Acad. Sci. USA 2013, 110, 14432–14437. [Google Scholar] [CrossRef] [PubMed]
- Vicente, R.; Rizzuto, M.; Sarica, C.; Yamamoto, K.; Sadr, M.; Khajuria, T.; Fatehi, M.; Moien-Afshari, F.; Haw, C.S.; Llinas, R.R.; et al. Enhanced interplay of neuronal coherence and coupling in the dying human brain. Front. Aging Neurosci. 2022, 14, 813531. [Google Scholar] [CrossRef]
- Chawla, L.S.; Akst, S.; Junker, C.; Jacobs, B.; Seneff, M.G. Surges of electroencephalogram activity at the time of death: A case series. J. Palliat. Med. 2009, 12, 1095–1100. [Google Scholar] [CrossRef] [PubMed]
- Persinger, M.A.; Rouleau, N.; Murugan, N.J.; Tessaro, L.W.E.; Costa, J.N. When is the brain dead? Living-like electrophysiological responses and photon emissions from applications of neurotransmitters in fixed post-mortem human brains. PLoS ONE 2016, 11, e0167231. [Google Scholar] [CrossRef]
- Blundon, E.G.; Gallagher, R.E.; Ward, L.M. Electrophysiological evidence of preserved hearing at the end of life. Sci. Rep. 2020, 10, 10336. [Google Scholar] [CrossRef]
- Blundon, E.G.; Gallagher, R.E.; Ward, L.M. Resting state network activation and functional connectivity in the dying brain. Clin. Neurophysiol. 2022, 135, 166–178. [Google Scholar] [CrossRef]
- Blundon, E.G.; Gallagher, R.E.; Ward, L.M. Electrophysiological evidence of sustained attention to music among conscious participants and unresponsive hospice patients at the end of life. Clin. Neurophysiol. 2022, 139, 9–22. [Google Scholar] [CrossRef]
- Fernández-Torre, J.L.; Hernández-Hernández, M.A.; Muñoz-Esteban, C. Non confirmatory electroencephalography in patients meeting clinical criteria for brain death: Scenario and impact on organ donation. Clin. Neurophysiol. 2013, 124, 2362–2367. [Google Scholar] [CrossRef]
- Grigg, M.M.; Kelly, M.A.; Celesia, G.G.; Ghobrial, M.W.; Ross, E.R. Electroencephalographic activity after brain death. Arch. Neurol. 1987, 44, 948–954. [Google Scholar] [CrossRef] [PubMed]
- Sutter, R.; Stevens, R.D.; Kaplan, P.W. Significance of triphasic waves in patients with acute encephalopathy: A nine-year cohort study. Clin. Neurophysiol. 2013, 124, 1952–1958. [Google Scholar] [CrossRef] [PubMed]
- Itil, T.M. Quantitative pharmacoelectroencephalography. In Psychotropic Drugs and the Human EEG: Modern Problems in Pharmachopsychiatry; Itil, T.M., Ed.; Karger: New York, NY, USA, 1974; Volume 8, pp. 43–75. [Google Scholar]
- Herrmann, W.M.; Schaerer, E. Pharmaco-EEG: Computer EEG analysis to describe the projection of drug effects on a functional cerebral level in humans. In Handbook of Electroencephalography and Clinical Neurophysiology; Silva, F.H.L., Leeuwen, W.S., Rémond, A., Eds.; Elsevier: Amsterdam, The Netherlands, 1986; Volume 2, pp. 386–445. [Google Scholar]
- Saletu, B. The use of pharmaco-EEG in drug profiling. In Human Psychopharmacology Measures and Methods; Hindmarch, I., Stonier, P.D., Eds.; John Wiley: New York, NY, USA, 1987; Volume 1, pp. 173–200. [Google Scholar]
- Mandema, J.W.; Danhof, M. Electroencephalogram effect measures and relationships between pharmacokinetics and pharmacodynamics of centrally acting drugs. Clin. Pharmacokinet. 1992, 23, 191–215. [Google Scholar] [CrossRef]
- Bruder, G.E.; Stewart, J.W.; Tenke, C.E.; McGrath, P.J.; Leite, P.; Bhattacharya, N.; Quitkin, F.M. Electroencephalographic and perceptual asymmetry differences between responders and nonresponders to an SSRI antidepressant. Biol. Psychiatry 2001, 49, 416–425. [Google Scholar] [CrossRef]
- Hermens, D.F.; Cooper, N.J.; Kohn, M.; Clarke, S.; Gordon, E. Predicting stimulant medication response in ADHD: Evidence from an integrated profile of neuropsychological, psychophysiological and clinical factors. J. Integr. Neurosci. 2005, 4, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Arns, M.; Gunkelman, J.; Breteler, M.; Spronk, D. EEG phenotypes predict treatment outcome to stimulants in children with ADHD. J. Integr. Neurosci. 2008, 7, 421–438. [Google Scholar] [CrossRef]
- Bruder, G.E.; Sedoruk, J.P.; Stewart, J.W.; McGrath, P.J.; Quitkin, F.M.; Tenke, C.E. Electroencephalographic alpha measures predict therapeutic response to a selective serotonin reuptake inhibitor antidepressant: Pre- and post-treatment findings. Biol. Psychiatry 2008, 63, 1171–1177. [Google Scholar] [CrossRef]
- Iosifescu, D.V.; Greenwald, S.; Devlin, P.; Perlis, R.H.; Denninger, J.W.; Alpert, J.E.; Fava, M. Pretreatment frontal EEG and changes in suicidal ideation during SSRI treatment in major depressive disorder. Acta Psychiatr. Scand. 2008, 117, 271–276. [Google Scholar] [CrossRef]
- Leuchter, A.F.; Cook, I.A.; Gilmer, W.S.; Marangell, L.B.; Burgoyne, K.S.; Howland, R.H.; Trivedi, M.H.; Zisook, S.; Jain, R.; Fava, M.; et al. Effectiveness of a quantitative electroencephalographic biomarker for predicting differential response or remission with escitalopram and bupropion in major depressive disorder. Psychiatry Res. 2009, 169, 132–138. [Google Scholar] [CrossRef]
- Leuchter, A.F.; Cook, I.A.; Marangell, L.B.; Gilmer, W.S.; Burgoyne, K.S.; Howland, R.H.; Trivedi, M.H.; Zisook, S.; Jain, R.; McCracken, J.T.; et al. Comparative effectiveness of biomarkers and clinical indicators for predicting outcomes of SSRI treatment in Major Depressive Disorder: Results of the BRITE-MD study. Psychiatry Res. 2009, 169, 124–131. [Google Scholar] [CrossRef]
- Iznak, A.F.; Iznak, E.V. EEG predictors of therapeutic responses in psychiatry. Neurosci. Behav. Physiol. 2022, 52, 207–212. [Google Scholar] [CrossRef]
- Cook, I.A.; Hunter, A.M.; Korb, A.; Farahbod, H.; Leuchter, A.F. EEG signals in psychiatry: Biomarkers for depression management. In Quantitative EEG Analysis Methods and Clinical Applications; Tong, S., Thakor, N.V., Eds.; Artech House: Norwood, MA, USA, 2009; pp. 289–315. [Google Scholar]
- Mednick, S.A.; Vka, J.V.; Gabrielli, J.W.F.; Itil, T.M. EEG as a predictor of antisocial behavior. Criminology 1981, 19, 219–230. [Google Scholar] [CrossRef]
- Fingelkurts, A.A.; Fingelkurts, A.A.; Neves, C.F.H. The structure of brain electromagnetic field relates to subjective experience: Exogenous magnetic field stimulation study. In Proceedings of the Neuroscience Finland 2013 Meeting: Optogenetics and Brain Stimulation, Helsinki, Finland, 22 March 2013. [Google Scholar]
- Daskalakis, Z.J.; Levinson, A.J.; Fitzgerald, P.B. Repetitive transcranial magnetic stimulation for major depressive disorder: A review. Can. J. Psychiatry 2008, 53, 555–566. [Google Scholar] [CrossRef]
- Lam, R.W.; Chan, P.; Wilkins-Ho, M.; Yatham, L.N. Repetitive transcranial magnetic stimulation for treatment-resistant depression: A systematic review and meta-analysis. Can. J. Psychiatry 2008, 53, 621–631. [Google Scholar] [CrossRef]
- Walker, J.E.; Kozlowski, G.P. Neurofeedback treatment of epilepsy. Child Adolesc. Psychiatr. Clin. N. Am. 2005, 14, 163–176. [Google Scholar] [CrossRef]
- Tan, G.; Thornby, J.; Hammond, D.C.; Strehl, U.; Canady, B.; Arnemann, K.; Kaiser, D.A. Meta-analysis of EEG biofeedback in treating epilepsy. Clin. EEG Neurosci. 2009, 40, 173–179. [Google Scholar] [CrossRef]
- Hammond, D.C. Neurofeedback with anxiety and affective disorders. Child Adolesc. Psychiatr. Clin. N. Am. 2005, 14, 105–123. [Google Scholar] [CrossRef]
- Surmeli, T.; Ertem, A.; Eralp, E.; Kos, I.H. Schizophrenia and the efficacy of qEEG-guided neurofeedback treatment: A clinical case series. Neurosci. Lett. 2011, 500S, e16. [Google Scholar] [CrossRef]
- Dehghani-Arani, F.; Rostami, R.; Nadali, H. Neurofeedback training for opiate addiction: Improvement of mental health and craving. Appl. Psychophysiol. Biofeedback 2013, 38, 133–141. [Google Scholar] [CrossRef]
- Arns, M.; De Ridder, S.; Strehl, U.; Breteler, M.; Coenen, A. Efficacy of neurofeedback treatment in ADHD: The effects on inattention, impulsivity and hyperactivity: A meta-analysis. Clin. EEG Neurosci. 2009, 40, 180–189. [Google Scholar] [CrossRef]
- Gevensleben, H.; Holl, B.; Albrecht, B.; Schlamp, D.; Kratz, O.; Studer, P.; Wangler, S.; Rothenberger, A.; Moll, G.H.; Heinrich, H. Distinct EEG effects related to neurofeedback training in children with ADHD: A randomized controlled trial. Int. J. Psychophysiol. 2009, 74, 149–157. [Google Scholar] [CrossRef]
- Hammer, B.U.; Colbert, A.P.; Brown, K.A.; Ilioi, E.C. Neurofeedback for insomnia: A pilot study of Z-score SMR and individualized protocols. Appl. Psychophysiol. Biofeedback 2011, 36, 251–264. [Google Scholar] [CrossRef]
- Kouijzer, M.E.J.; van Schie, H.T.; de Moor, J.M.H.; Gerrits, B.J.L.; Buitelaar, J.K. Neurofeedback treatment in autism. Preliminary findings in behavioral, cognitive, and neurophysiological functioning. Res. Autism Spectr. Disord. 2010, 4, 386–399. [Google Scholar] [CrossRef]
- Ibric, V.L.; Dragomirescu, L.G. Neurofeedback in pain management. In Introduction to Quantitative EEG Neurofeedback Advanced Theory and Application, 2nd ed.; Budzynski, T.H., Budzynski, H.K., Evans, J.R., Abarbanel, A., Eds.; Elsevier: New York, NY, USA, 2009; pp. 417–451. [Google Scholar] [CrossRef]
- Orlando, P.C.; Rivera, R.O. Neurofeedback for elementary students with identified learning problems. J. Neurother. 2004, 8, 5–19. [Google Scholar] [CrossRef]
- Breteler, M.H.; Arns, M.; Peters, S.; Giepmans, I.; Verhoeven, L. Improvements in spelling after QEEG-based neurofeedback in dyslexia: A randomized controlled treatment study. Appl. Psychophysiol. Biofeedback 2010, 35, 5–11. [Google Scholar] [CrossRef] [Green Version]
- Hanslmayr, S.; Sauseng, P.; Doppelmayr, M.; Schabus, M.; Klimesch, W. Increasing individual upper alpha power by neurofeedback improves cognitive performance in human subjects. Appl. Psychophysiol. Biofeedback 2005, 30, 1–10. [Google Scholar] [CrossRef]
- Angelakis, E.; Stathopoulou, S.; Frymiare, J.L. EEG neurofeedback: A brief overview and an example of peak alpha frequency training for cognitive enhancement in the elderly. Clin. Neuropsychol. 2007, 21, 110–129. [Google Scholar] [CrossRef]
- Fingelkurts, A.A.; Fingelkurts, A.A.; Neves, C.F.H. Consciousness as a phenomenon in the operational architectonics of brain organization: Criticality and self-organization considerations. Chaos Solitons Fractals 2013, 55, 13–31. [Google Scholar] [CrossRef]
- Thatcher, R.W.; John, E.R. Functional Neuroscience: I. Foundations of Cognitive Processes; Lawrence Erlbaum: Boca Raton, NJ, USA, 1977; p. 370. [Google Scholar]
- Bodunov, M.V. The EEG “alphabet”: The typology of human EEG stationary segments. In Individual and Psychological Differences and Bioelectrical Activity of Human Brain; Rusalov, V.M., Ed.; Nauka: Moscow, Russia, 1988; pp. 56–70. (In Russian) [Google Scholar]
- Jansen, B.H.; Cheng, W.-K. Structural EEG analysis: An explorative study. Int. J. Biomed. Comput. 1988, 23, 221–237. [Google Scholar] [CrossRef]
- Fingelkurts, A.A.; Fingelkurts, A.A.; Kaplan, A.Y. The regularities of the discrete nature of multi-variability of EEG spectral patterns. Int. J. Psychophysiol. 2003, 47, 23–41. [Google Scholar] [CrossRef]
- Fingelkurts, A.A.; Fingelkurts, A.A.; Krause, C.M.; Kaplan, A.Y. Systematic rules underlying spectral pattern variability: Experimental results and a review of the evidences. Int. J. Neurosci. 2003, 113, 1447–1473. [Google Scholar] [CrossRef] [PubMed]
- Fingelkurts, A.A.; Fingelkurts, A.A. Operational Architectonics of the human brain biopotential field: Towards solving the mind-brain problem. BrainMind 2001, 2, 261–296. Available online: http://www.bm-science.com/team/art18.pdf (accessed on 1 May 2022).
- Kaplan, A.Y.; Fingelkurts, A.A.; Fingelkurts, A.A.; Borisov, S.V.; Darkhovsky, B.S. Nonstationary nature of the brain activity as revealed by EEG/MEG: Methodological, practical and conceptual challenges. Signal Process. 2005, 85, 2190–2212. [Google Scholar] [CrossRef]
- Fingelkurts, A.A.; Fingelkurts, A.A. Brain-mind Operational Architectonics imaging: Technical and methodological aspects. Open Neuroimag. J. 2008, 2, 73–93. [Google Scholar] [CrossRef]
- Fingelkurts, A.A.; Fingelkurts, A.A. Editorial: EEG Phenomenology and Multiple Faces of Short-term EEG Spectral Pattern. Open Neuroimag. J. 2010, 4, 111–113. [Google Scholar] [PubMed]
- Fingelkurts, A.A.; Fingelkurts, A.A. Making complexity simpler: Multivariability and metastability in the brain. Int. J. Neurosci. 2004, 114, 843–862. [Google Scholar] [CrossRef]
- Barlow, J.S. Methods of analysis of nonstationary EEGs, with emphasis on segmentation techniques: A comparative review. J. Clin. Neurophysiol. 1985, 2, 267–304. [Google Scholar] [CrossRef]
- Fingelkurts, A.A.; Fingelkurts, A.A. Mapping of the brain operational architectonics. In Focus on Brain Mapping; Chen, F.J., Ed.; Research Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2005; pp. 59–98. Available online: http://www.bm-science.com/team/chapt3.pdf (accessed on 1 May 2022).
- Friston, K. The labile brain. I. Neuronal transients and nonlinear coupling. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2000, 355, 215–236. [Google Scholar] [CrossRef]
- Triesch, J.; von der Malsburg, C. Democratic integration: Self-organized integration of adaptive cues. Neural Comput. 2001, 13, 2049–2074. [Google Scholar] [CrossRef]
- Buzsaki, G. Large-scale recording of neuronal ensembles. Nat. Neurosci. 2004, 7, 446–451. [Google Scholar] [CrossRef]
- Averbeck, B.B.; Lee, D. Coding and transmission of information by neural ensembles. Trends Neurosci. 2004, 27, 225–230. [Google Scholar] [CrossRef]
- Freeman, W.J.; Vitiello, G. Nonlinear brain dynamics and many-body field dynamics. Electromagn. Biol. Med. 2005, 24, 233–241. [Google Scholar] [CrossRef]
- Stam, C.J. Nonlinear Brain Dynamics, 1st ed.; Nova Science Publishers, Inc.: New York, NY, USA, 2006; p. 148. [Google Scholar]
- Freeman, W.J. Indirect biological measures of consciousness from field studies of brains as dynamical systems. Neural Netw. 2007, 20, 1021–1031. [Google Scholar] [CrossRef]
- Lehmann, D. Brain electric microstates and cognition: The atoms of thought. In Machinery of the Mind; John, E.R., Ed.; Birkhauser: Boston, MA, USA, 1990; pp. 209–224. [Google Scholar]
- Freeman, W.J. Tutorial on neurobiology: From single neurons to brain chaos. Int. J. Bifurcat. Chaos 1992, 2, 451–482. [Google Scholar] [CrossRef]
- John, R.; Easton, P.; Isenhart, R. Consciousness and cognition may be mediated by multiple independent coherent ensembles. Con Cogn. 1997, 6, 3–39. [Google Scholar] [CrossRef]
- Wright, J.J.; Kydd, R.R.; Lees, G.J. State-changes in the brain viewed as linear steady-states and non-linear transitions between steady-states. Biol. Cybern. 1985, 53, 11–17. [Google Scholar] [CrossRef]
- Lehmann, D.; Strik, W.K.; Henggeler, B.; Koenig, T.; Koukkou, M. Brain electric microstates and momentary conscious mind states as building blocks of spontaneous thinking: I. Visual imagery and abstract thoughts. Int. J. Psychophysiol. 1998, 29, 1–11. [Google Scholar] [CrossRef]
- Fingelkurts, A.A.; Fingelkurts, A.A. Topographic mapping of rapid transitions in EEG multiple frequencies: EEG frequency domain of operational synchrony. Neurosci. Res. 2010, 68, 207–224. [Google Scholar] [CrossRef]
- Freeman, W.J. On the problem of anomalous dispersion in chaoto-chaotic phase transitions of neural masses, and its significance for the management of perceptual information in brains. In Synergetics of Cognition; Haken, H., Stadler, M., Eds.; Springer: Berlin/Heidelberg, Germany, 1990; Volume 45, pp. 126–143. [Google Scholar]
- Palm, G. Cell assemblies as a guideline for brain research. Concepts Neurosci. 1990, 1, 133–147. [Google Scholar]
- Eichenbaum, H. Thinking about brain cell assemblies. Science 1993, 261, 993–994. [Google Scholar] [CrossRef]
- von der Malsburg, C. The what and why of binding: The modeler’s perspective. Neuron 1999, 24, 95–104. [Google Scholar] [CrossRef]
- Buzsáki, G. Rhythms of the Brain; Oxford University Press: Oxford, UK, 2006; p. 448. [Google Scholar]
- Lehmann, D. Multichannel topography of human alpha EEG fields. Electroencephalogr. Clin. Neurophysiol. 1971, 31, 439–449. [Google Scholar] [CrossRef]
- Stam, C.J. Nonlinear dynamical analysis of EEG and MEG: Review of an emerging field. Clin. Neurophysiol. 2005, 116, 2266–2301. [Google Scholar] [CrossRef]
- Wackermann, J.; Allefeld, C. On the meaning and interpretation of global descriptors of brain electrical activity. Including a reply to X. Pei et al. Int. J. Psychophysiol. 2007, 64, 199–210. [Google Scholar] [CrossRef]
- Freeman, W.J. The wave packet: An action potential for the 21st Century. J. Integr. Neurosci. 2003, 2, 3–30. [Google Scholar] [CrossRef]
- Fingelkurts, A.A.; Fingelkurts, A.A. Morphology and dynamic repertoire of EEG short-term spectral patterns in rest: Explorative study. Neurosci. Res. 2010, 66, 299–312. [Google Scholar] [CrossRef]
- Freeman, W.J.; Holmes, M.D. Metastability, instability, and state transition in neocortex. Neural Netw. 2005, 18, 497–504. [Google Scholar] [CrossRef] [Green Version]
- Penttonen, M.; Buzsaki, G. Natural logarithmic relationship between brain oscillators. Thalamus Relat. Syst. 2003, 2, 145–152. [Google Scholar] [CrossRef]
- Hoppensteadt, F.C.; Izhikevich, E.M. Weakly Connected Neural Networks; Springer: New York, NY, USA, 1997; p. 402. [Google Scholar]
- Izhikevich, E.M. Weakly connected quasi-periodic oscillators, FM interactions, and multiplexing in the brain. SIAM J. Appl. Math. 1999, 59, 2193–2223. [Google Scholar] [CrossRef]
- Izhikevich, E.M.; Desai, N.S.; Walcott, E.C.; Hoppensteadt, F.C. Bursts as a unit of neural information: Selective communication via resonance. Trends Neurosci. 2003, 26, 161–167. [Google Scholar] [CrossRef]
- Freeman, W.J.; Rogers, L.J. Fine temporal resolution of analytic phase reveals episodic synchronization by state transitions in gamma EEGs. J. Neurophysiol. 2002, 87, 937–945. [Google Scholar] [CrossRef]
- Kozma, R.; Puljic, M.; Balister, P.; Bollobas, B.; Freeman, W.J. Phase transitions in the neuropercolation model of neural populations with mixed local and nonlocal interactions. Biol. Cyber. 2005, 92, 367–379. [Google Scholar] [CrossRef]
- Friston, K.J.; Frith, C.D.; Liddle, P.F.; Frackowiak, R.S.J. Functional connectivity: The principal component analysis of large (PET) data sets. J. Cereb. Blood Flow Metab. 1993, 13, 5–14. [Google Scholar] [CrossRef]
- Friston, K.J.; Frith, C.D.; Fletcher, P.; Liddle, P.F.; Frackowiak, R.S.J. Functional topography: Multidimensional scaling and functional connectivity in the brain. Cereb. Cortex 1996, 6, 156–164. [Google Scholar] [CrossRef]
- Nunez, P.L. Generation of human EEG by a combination of long- and short-range neocortical interactions. Brain Topogr. 1989, 1, 199–215. [Google Scholar] [CrossRef]
- Werner, G. Metastability, criticality and phase transitions in brain and its models. Biosystems 2007, 90, 496–508. [Google Scholar] [CrossRef]
- Kelso, J.A.S. Dynamic Patterns: The Self-Organization of Brain and Behavior; MIT Press: Cambridge, MA, USA, 1995; p. 334. [Google Scholar]
- Fingelkurts, A.A.; Fingelkurts, A.A.; Neves, C.F.H. Phenomenological architecture of mind and operational architectonics of the brain: The unified metastable continuum. New Math. Nat. Comput. 2009, 5, 221–244. [Google Scholar] [CrossRef] [Green Version]
- Ukhtomsky, A.A. Selected Works; Nauka: Lenengrad, Russia, 1978; p. 358. (In Russian) [Google Scholar]
- Breakspear, M.; Terry, J.R.; Friston, K.J.; Harris, A.W.F.; Williams, L.M.; Brown, K.; Brennan, J.; Gordona, E. A disturbance of nonlinear interdependence in scalp EEG of subjects with first episode schizophrenia. NeuroImage 2003, 20, 466–478. [Google Scholar] [CrossRef]
- Tononi, G.; Edelman, G.M. Schizophrenia and the mechanisms of conscious integration. Brain Res. Rev. 2000, 31, 391–400. [Google Scholar] [CrossRef]
- Andreasen, N.C. A unitary model of schizophrenia: Bleuler’s “fragmented phrene” as schizencephaly. Arch. Gen. Psychol. 1999, 56, 781–787. [Google Scholar] [CrossRef]
- Schulman, J.J.; Cancro, R.; Lowe, S.; Lu, F.; Walton, K.D.; Llinás, R.R. Imaging of thalamocortical dysrhythmia in neuropsychiatry. Front. Hum. Neurosci. 2011, 5, 69. [Google Scholar] [CrossRef]
- Buckholtz, J.W.; Meyer-Lindenberg, A. Psychopathology and the human connectome: Toward a transdiagnostic model of risk for mental illness. Neuron 2012, 74, 990–1004. [Google Scholar] [CrossRef]
- Dawson, K.A. Temporal organization of the brain: Neurocognitive mechanisms and clinical implications. Brain Cogn. 2004, 54, 75–94. [Google Scholar] [CrossRef]
- Gasser, T.; Bacher, P.; Mochs, J. Transformation towards the normal distribution of broad band spectral parameters of the EEG. Electroencephalogr. Clin. Neurophysiol. 1982, 53, 119–124. [Google Scholar] [CrossRef]
- John, E.R.; Prichep, L.S.; Easton, P. Normative data banks and neurometrics: Basic concepts, methods and results of norm construction. In Handbook of Electroencephalography and Clinical Neurophysiology; Gevins, A.S., Remond, A., Eds.; Elsevier: Amsterdam, The Netherlands, 1987; Volume I, pp. 449–495. [Google Scholar]
- John, E.R.; Prichep, L.S. Principles of neurometrics and neurometric analysis of EEG and evoked potentials. In EEG: Basic Principles, Clinical Applications and Related Fields; Niedermeyer, E., Da Silva, F.L., Eds.; Williams & Wilkins: Baltimore, MD, USA, 1993; pp. 989–1003. [Google Scholar]
- Fingelkurts, A.A.; Fingelkurts, A.A. Altered structure of dynamic electroencephalogram oscillatory pattern in major depression. Biol. Psychiatry 2015, 77, 1050–1060. [Google Scholar] [CrossRef]
- Johnstone, J.; Gunkelman, J.; Lunt, J. Clinical database development: Characterization of EEG phenotypes. Clin. EEG Neurosci. 2005, 36, 99–107. [Google Scholar] [CrossRef]
- Zhirmunskaya, E.K.; Losev, B.C.; Maslov, V.K. Mathematical analysis of EEG type and interhemispheric EEG asymmetry. Hum. Physiol. 1978, 5, 791–799. [Google Scholar]
- Zhirmunskaya, E.A.; Losev, V.S. The concept of type in the classification of electroencephalograms. Hum. Physiol. 1980, 6, 1039–1045. [Google Scholar]
- Zhirmunskaya, E.A.; Losev, B.C. Description Systems and Classification of Human Electroencephalograms; Nauka: Moscow, Russia, 1984; p. 81. [Google Scholar]
- Zhirmunskaya, E.A. In Search of an Explanation of EEG Phenomena; Biola: Moscow, Russia, 1996; p. 117. [Google Scholar]
- Misyuk, N.N. Diagnostic efficiency and basic principles of classification of electroencephalograms. Med. News 2006, 1, 24–33. [Google Scholar]
- Bochkarev, V.C.; Panyushkin, S.V. Electroencephalographic studies in borderline conditions. In Borderline Mental Disorders, 3rd ed.; Aleksandrovsky, Y.A., Ed.; Moscow, Russia, 2000; pp. 120–133. [Google Scholar]
- Chemiy, V.I.; Ostrovaya, T.V. The diagnostic algorithm of assessment of EEG for estimation of brain in vestigation of central nervous system reactivity in response to photostimulation and pharmacological influence. Neurosci. Theor. Clin. Asp. 2005, 1, 12–51. [Google Scholar]
- Koroleva, N.V.; Kolesnikov, S.I.; Dolgih, V.V. Dynamic of electroencephalogram’s descriptors in children with different EEG types. Bull. RAMH 2007, 2, 49–51. [Google Scholar]
- Gelda, A.P.; Dokukina, T.V.; Misyuk, N.N.; Cosmidiadi, A.O. Registration of electroencephalograms during psychopharmacotherapy. Med. J. 2008, 4, 16–18. [Google Scholar]
- Misyuk, N.N.; Gelda, A.P.; Dokukina, T.V.; Cosmidiadi, A.O. Types of electroencephalograms in schizophrenia. Med. J. 2008, 4, 41–43. [Google Scholar]
- Zvereva, Z.F.; Vanchakova, N.P.; Zolotaryova, N.N. Clinical and neurophysiological parameters in patients with discirculatory encephalopathy. Zh. Nevrol. Psikhiatr. 2010, 110, 2–15. [Google Scholar]
- Koichubekov, B.K.; Sorokina, M.A.; Pashev, V.I.; Shaikhin, A.M. Individually-typological of CNS’s regulatory processes in persons with initial signs of neurocirculatory dystonia. Fundam. Res. 2012, 5, 300–304. [Google Scholar]
- Gibson, E.; Lobaugh, N.J.; Joordens, J.; McIntosh, A.R. EEG variability: Task-driven or subject-driven signal of interest? NeuroImage 2022, 252, 119034. [Google Scholar] [CrossRef]
- Danilova, N.N. On individual peculiarities of the electrical activity of the cerebral cortex of humans. In Typological Peculiarities of the Higher Nervous Activity of Humans; Academic Psychological Science: Moscow, Russia, 1963; Volume 3, pp. 262–274. [Google Scholar]
- Nebylitsyn, V.D. An electroencephalographic study of the properties of the strength of the nervous system and the balance of the nerve processes in humans using factorial analysis. In Typological Peculiarities of Higher Nervous Activity in Humans; Academic Psychological Science: Moscow, Russia, 1963; pp. 47–80. [Google Scholar]
- Egorova, I.S. Electroencephalography; Meditsina: Moscow, Russia, 1973. [Google Scholar]
- Soroko, S.I.; Bekshaev, S.S. Statistical structure of EEG rhythms and individual properties of brain self-regulatory mechanisms. Physiol. J. 1981, 67, 1765–1773. [Google Scholar]
- Boldyreva, G.N.; Sharova, E.V.; Dobronravova, I.S. Role of cerebral regulatory structures in the formation of EEG in humans. Hum. Physiol. (Fiziol. Cheloveka) 2000, 26, 19–34. [Google Scholar]
- Maryutina, T.M. Intermediate phenotypes of intelligence in the context of genetic psychophysiology. Psychology. J. High. Sch. Econ. 2007, 4, 22–47. [Google Scholar]
- Clarke, A.R.; Barry, R.J.; McCarthy, R.; Selikowitz, M.; Johnstone, S.J.; Hsu, C.-I.M.; Magee, C.A.; Lawrence, C.A.; Croft, R.J. Coherence in children with attention-deficit/hyperactivity disorder and excess beta activity in their EEG. Clin. Neurophysiol. 2007, 118, 1472–1479. [Google Scholar] [CrossRef]
- Suffin, S.C.; Emory, W.H.; Gutierrez, N.; Arora, G.S.; Schiller, M.J.; Kling, A. A QEEG database method for predicting pharmacotherapeutic outcome in refractory major depressive disorders. J. Am. Phys. Surg. 2007, 12, 104–108. [Google Scholar]
- Almasy, L. Quantitative risk factors as indices of alcoholism susceptibility. Ann. Med. 2003, 35, 337–343. [Google Scholar] [CrossRef]
- Porjesz, B.; Begleiter, H. Alcoholism and human electrophysiology. Alcohol. Res. Health 2003, 27, 53–160. [Google Scholar]
- Vasilevskii, N.N.; Suvorov, N.B.; Sidorov, Y.A.; Bovtyushko, V.G. Risk factors and some features of the pathology depending on the organization type of cerebral neurodynamyics. Vestn. Ross Akad. Med. Nauk. 1996, 9, 14–18. [Google Scholar]
- Kendler, K.; Neale, M. Endophenotype: A conceptual analysis. Mol. Psychiatry 2010, 15, 789–797. [Google Scholar] [CrossRef]
- Bodunov, M.V. Individual and typological characteristics of EEG structure. J. High Nerve Act. 1985, 35, 1045–1052. [Google Scholar]
- Vasilevskii, N.N.; Soroko, S.I.; Zingerman, A.M. Psychophysiological principles of individual typological features of humans. In Mechanisms of Brain Activity in Humans; Bekhtereva, N.P., Ed.; Nauka: Leningrad, Russia, 1988; pp. 455–490. [Google Scholar]
- Soroko, S.I.; Bekshaev, S.S.; Sidorov, Y.A. The Main Types of The Brain Self-Regulation Mechanisms; Nauka: Leningrad, Russia, 1990; p. 205. [Google Scholar]
- Suvorov, N.B.; Zueva, N.G.; Guseva, N.L. Reflection of individual typological features in the structure of spatial interaction of EEG waves of various frequency ranges. Hum. Physiol. 2000, 26, 301–306, (Translated from Fiziologiya Cheloveka 2000, 26, 60–66). [Google Scholar] [CrossRef]
- Roth, N.; Sask, G. Relations between slow (4 cps) EEG activity, sensor motor speed, and psychopathology. Int. J. Psychophysiol. 1990, 9, 121–127. [Google Scholar] [CrossRef]
- Markina, A.V.; Pashina, A.K.; Rumanova, N.B. Correlation of the electroencephalogram rhythms with cognitive/personality-related peculiarities of the subject. Physiol. J. 2000, 21, 47–55. [Google Scholar]
- Chorayan, O.G.; Aidarkin, E.K.; Chorayan, I.O. Individual-typological features of regulation and interaction of functional systems in different modes of activity: Review. Valeology 2001, 2, 5–15. [Google Scholar]
- Svyatogor, I.A.; Mokhovikova, I.A.; Bekshayev, S.S.; Nozdrachev, A.D. EEG pattern as an instrument for evaluation of neurophysiological mechanisms underlying adaptation disorders. J. High. Nerve Act. 2005, 55, 178–188. [Google Scholar]
- Konareva, I.N. Modulation of low-frequency EEG rhythms under conditions of an activation reaction: Dependence on psychological characteristics of personality. Neurophysiology 2011, 42, 42–52. [Google Scholar] [CrossRef]
- Knott, J.R.; Gottlieb, J.S. Electroencephalographic evaluation of psychopathic personality: Correlations with age, sex, family history and antecedent illness or injury. Arch. Neurol. Psychiatry 1944, 52, 515–519. [Google Scholar] [CrossRef]
- Hodge, R.S. The impulsive psychopath: A clinical and electrophysiological study. J. Ment. Sci. 1945, 91, 472–476. [Google Scholar] [CrossRef]
- Diethelm, O.; Simons, D.J. Electroencephalographic changes associated with psychopathic personalities. Arch. Neurol. Psychiatry 1946, 55, 410–413. [Google Scholar]
- Hill, D. EEG in episodic psychotic and psychopathic behaviour: A classification of data. Electroencephalogr. Clin. Neurophysiol. 1952, 4, 419–442. [Google Scholar] [CrossRef]
- Mizuki, Y.; Kajimura, N.; Nishikori, S.; Imaizumi, J.; Yamada, M. Appearance of frontal midline theta rhythm and personality traits. Folia Psychiatr. Neurol. Jpn. 1984, 38, 451–458. [Google Scholar] [CrossRef]
- Rusalov, V.M.; Rusalova, M.N.; Kalashnikova, I.G.; Stepanov, V.G.; Strel’nikova, T.N. The bioelectrical activity of the human brain in representatives of different temperamental types. Zh. Vyss. Nerv. Deiat. Im. I P Pavlov. 1993, 43, 530–542. (In Russian) [Google Scholar] [PubMed]
- Golan, Z.; Neufield, M.Y. Individual differences in alpha rhythm as characterizing temperament related to cognitive performances. Personal. Individ. Differ. 1996, 21, 775–784. [Google Scholar] [CrossRef]
- Sviderskaia, N.E.; Korol’kova, T.A. The effect of the properties of the nervous system and the temperament on the spatial organization of the EEG. Zh. Vyss. Nerv. Deiat. Im. I P Pavlov. 1996, 46, 849–858. [Google Scholar]
- Tran, Y.; Craig, A.; McIsaac, P. Extraversion–introversion and 8–13 Hz waves in frontal cortical regions. Persinal. Individ. Differ. 2001, 30, 205–215. [Google Scholar] [CrossRef]
- Knyazev, G.G.; Slobodskaya, H.R. Personality trait of behavioural inhibition is associated with oscillatory systems reciprocal relationships. Int. J. Psychophysiol. 2003, 48, 247–261. [Google Scholar] [CrossRef]
- Chi, S.E.; Park, C.B.; Lim, S.L.; Park, E.H.; Lee, Y.H.; Lee, K.H.; Kim, E.J.; Kim, H.T. EEG and personality dimensions: A consideration based on the brain oscillatory systems. Personal. Individ. Differ. 2005, 39, 669–681. [Google Scholar] [CrossRef]
- Knyazev, G.G. EEG correlates of personality types. Neth. J. Psychol. 2006, 62, 78–87. [Google Scholar] [CrossRef]
- Konareva, I.N. Correlation between level of aggressiveness of personality and characteristics of EEG frequency components. Neurophysiology 2006, 38, 380–388. [Google Scholar] [CrossRef]
- Knyazev, G.G. Antero-posterior EEG spectral power gradient as a correlate of extraversion and behavioral inhibition. Open Neuroimag. J. 2010, 4, 114–120. [Google Scholar] [CrossRef]
- Jach, H.K.; Feuerriegel, D.; Smillie, L.D. Decoding personality trait measures from resting EEG: An exploratory report. Cortex 2020, 130, 158–171. [Google Scholar] [CrossRef]
- Jawinski, P.; Markett, S.; Sander, C.; Huang, J.; Ulke, C.; Hegerl, U.; Hensch, T. The Big Five personality traits and brain arousal in the resting state. Brain Sci. 2021, 11, 1272. [Google Scholar] [CrossRef]
- Gallagher, J.R.; Gibbs, E.L.; Gibbs, F.A. Relation between the electrical activity of the cortex and the personality in adolescent boys. Psychosom. Med. 1942, 4, 134–139. [Google Scholar] [CrossRef]
- Kennard, M.K. The electroencephalogram in psychological disorders: A review. Psychosom. Med. 1953, 15, 95–115. [Google Scholar] [CrossRef]
- Williams, D. The significance of an abnormal electroencephalogram. J. Neurol. Psychiatry 1941, 4, 257–268. [Google Scholar] [CrossRef]
- Hill, D.; Watterson, D. Electroencephalographic studies of psychopathic personalities. J. Neurol. Psychiatry 1942, 5, 47–65. [Google Scholar] [CrossRef]
- Williams, D. Neural factors related to habitual aggression: Consideration of differences between those habitual aggressives and others who have committed crimes of violence. Brain 1969, 92, 503–520. [Google Scholar] [CrossRef]
- Akiskal, H.S.; Hirschfeld, R.M.A.; Yerevanian, B.I. The relationship of personality to affective disorders: A critical review. Arch. Gen. Psychiatry 1983, 40, 801–810. [Google Scholar] [CrossRef]
- Hensch, T.; Herold, U.; Brocke, B. An electrophysiological endophenotype of hypomanic and hyperthymic personality. J. Affect. Disord. 2007, 101, 13–26. [Google Scholar] [CrossRef]
- Barnett, J.H.; Huang, J.; Perlis, R.H.; Young, M.M.; Rosenbaum, J.F.; Nierenberg, A.A.; Sachs, G.; Nimgaonkar, V.L.; Miklowitz, D.J.; Smoller, J.W. Personality and bipolar disorder: Dissecting state and trait associations between mood and personality. Psychol. Med. 2011, 41, 1593–1604. [Google Scholar] [CrossRef]
- Klein, D.N.; Kotov, R.; Bufferd, S.J. Personality and depression: Explanatory models and review of the evidence. Annu. Rev. Clin. Psychol. 2011, 7, 269–295. [Google Scholar] [CrossRef]
- Jeronimus, B.F.; Kotov, R.; Riese, H.; Ormel, J. Neuroticism’s prospective association with mental disorders halves after adjustment for baseline symptoms and psychiatric history, but the adjusted association hardly decays with time: A meta-analysis on 59 longitudinal/prospective studies with 443 313 participants. Psychol. Med. 2016, 46, 2883–2906. [Google Scholar]
- Widiger, T.A. Personality and psychopathology. World Psychiatry 2011, 10, 103–106. [Google Scholar] [CrossRef]
- Robinson, D.L. How brain arousal systems determine different temperament types and the major dimensions of personality. Personal. Individ. Differ. 2001, 31, 1233–1259. [Google Scholar] [CrossRef]
- Knyazev, G.G.; Slobodskaya, H.R.; Safronova, M.V.; Sorokin, O.V.; Goodman, R.; Wilson, G.D. Personality, psychopathology and brain oscillations. Personal. Individ. Differ. 2003, 53, 1331–1349. [Google Scholar] [CrossRef]
- Cuthbert, B.N.; Insel, T.R. Toward the future of psychiatric diagnosis: The seven pillars of RDoC. BMC Med. 2013, 11, 126. [Google Scholar] [CrossRef] [PubMed]
- Trofimova, I.; Bajaj, S.; Bashkatov, S.A.; Blair, J.; Brandt, A.; Chan, R.C.K.; Clemens, B.; Corr, P.J.; Cyniak-Cieciura, M.; Demidova, L.; et al. What is next for the neurobiology of temperament, personality and psychopathology? Curr. Opin. Behav. Sci. 2022, 45, 101143. [Google Scholar] [CrossRef]
- de Rooij, S.R. Are brain and cognitive reserve shaped by early life circumstances? Front. Neurosci. 2022, 16, 825811. [Google Scholar] [CrossRef]
- Kent, L.; Nelson, B.; Northoff, G. Can disorders of subjective time inform the differential diagnosis of psychiatric disorders? A transdiagnostic taxonomy of time. Early Interv. Psychiatry 2022, 2022, 1–13. [Google Scholar] [CrossRef]
- Danilova, N.N. Psychophysiological Diagnostics of Functional States; Golubeva, E.A., Tushmalova, N.A., Eds.; Publishing House of Moscow State University: Moscow, Russia, 1992; p. 192. [Google Scholar]
- Bechtereva, N.P. Human Brain in Health and Disease; ACT Press: St. Petersburg, Russia, 2010. [Google Scholar]
- Kahneman, D. Attention and Effort; Englewood Cliffs: Prentice Hall, NJ, USA, 1973; p. 246. [Google Scholar]
- Hegerl, U.; Wilk, K.; Olbrich, S.; Schoenknecht, P.; Sander, C. Hyperstable regulation of vigilance in patients with major depressive disorder. World J. Biol. Psychiatry 2012, 13, 436–446. [Google Scholar] [CrossRef]
- Wahbeh, H.; Oken, B.S. Peak high-frequency HRV and peak alpha frequency higher in PTSD. Appl. Psychophys. Biof. 2013, 38, 57–69. [Google Scholar] [CrossRef]
- Schmidt, F.M.; Pschiebl, A.; Sander, C.; Kirkby, K.C.; Thormann, J.; Minkwitz, J.; Chittka, T.; Weschenfelder, J.; Holdt, L.M.; Teupser, D.; et al. Impact of serum cytokine levels on EEG-measured arousal regulation in patients with major depressive disorder and healthy controls. Neuropsychobiology 2016, 73, 1–9. [Google Scholar] [CrossRef]
- Ulke, C.; Tenke, C.E.; Kayser, J.; Sander, C.; Böttger, D.; Wong, L.Y.X.; Alvarenga, J.E.; Fava, M.; McGrath, P.J.; Deldin, P.J.; et al. Resting EEG measures of brain arousal in a multisite study of major depression. Clin. EEG Neurosci. 2019, 50, 3–12. [Google Scholar] [CrossRef]
- Hegerl, U.; Lam, R.W.; Malhi, G.S.; McIntyre, R.S.; Demyttenaere, K.; Mergl, R.; Gorwood, P. Conceptualising the neurobiology of fatigue. Aust. N. Z. J. Psychiatry 2013, 47, 312–316. [Google Scholar] [CrossRef]
- Hegerl, U.; Ulke, C. Fatigue with up-vs downregulated brain arousal should not be confused. Prog. Brain Res. 2016, 229, 239–254. [Google Scholar]
- Ulke, C.; Surova, G.; Sander, C.; Engel, C.; Wirkner, K.; Jawinski, P.; Hensch, T.; Hegerl, U. Fatigue in cancer and neuroinflammatory and autoimmune disease: CNS arousal matters. Brain Sci. 2020, 10, 569. [Google Scholar] [CrossRef]
- Okogbaa, O.G.; Shell, R.L.; Filipusic, D. On the investigation of the neurophysiological correlates of knowledge worker mental fatigue using the EEG signal. Appl. Ergon. 1994, 25, 355–365. [Google Scholar] [CrossRef]
- Billiot, K.M.; Budzynski, T.H.; Andrasik, F. EEG patterns and chronic fatigue syndrome. J. Neurother. 1997, 2, 20–30. [Google Scholar] [CrossRef]
- Zinn, M.A.; Zinn, M.L.; Valencia, I.; Jason, L.A.; Montoya, J.G. Cortical hypoactivation during resting EEG suggests central nervous system pathology in patients with chronic fatigue syndrome. Biol. Psychol. 2018, 136, 87–99. [Google Scholar] [CrossRef]
- Van Luijtelaar, G.; Verbraak, M.; van den Bunt, M.; Keijsers, G.; Arns, M. EEG findings in burnout patients. J. Neuropsychiatry Clin. Neurosci. 2010, 22, 208–217. [Google Scholar] [CrossRef]
- Stenberg, G. Personality and the EEG: Arousal and emotional arousability. Personal. Individ. Differ. 1992, 13, 1097–1113. [Google Scholar] [CrossRef]
- Clarke, A.R.; Barry, R.J.; McCarthy, R.; Selikowitz, M.; Brown, C. EEG evidence for a new conceptualisation of attention deficit hyperactivity disorder. Clin. Neurophysiol. 2002, 113, 1036–1044. [Google Scholar] [CrossRef]
- Zhang, D.-W.; Johnstone, S.J.; Roodenrys, S.; Luo, X.; Li, H.; Wang, E.; Zhao, Q.; Song, Y.; Liu, L.; Qian, Q.; et al. The role of EEG localized activation and central nervous system arousal in executive function performance in children with Attention-Deficit/Hyperactivity Disorder. Clin. Neurophysiol. 2018, 129, 1192–1200. [Google Scholar] [CrossRef]
- Ulke, C.; Sander, C.; Jawinski, P.; Mauche, N.; Huang, J.; Spada, J.; Wittekind, D.; Mergl, R.; Luck, T.; Riedel-Heller, S.; et al. Sleep disturbances and upregulation of brain arousal during daytime in depressed versus non-depressed elderly subjects. World J. Biol. Psychiatry 2017, 18, 633–640. [Google Scholar] [CrossRef]
- Eysenck, H.J. The Biological Basis of Personality; Thomas: Springfield, IL, USA, 1967; p. 420. [Google Scholar]
- Lo, M.T.; Hinds, D.A.; Tung, J.Y.; Franz, C.; Fan, C.C.; Wang, Y.; Smeland, O.B.; Schork, A.; Holland, D.; Kauppi, K.; et al. Genome-wide analyses for personality traits identify six genomic loci and show correlations with psychiatric disorders. Nat. Genet. 2017, 49, 152–156. [Google Scholar] [CrossRef]
- Baselmans, B.M.L.; Jansen, R.; Ip, H.F.; van Dongen, J.; Abdellaoui, A.; van de Weijer, M.P.; Bao, Y.; Smart, M.; Kumari, M.; Willemsen, G.; et al. Multivariate genome-wide analyses of the well-being spectrum. Nat. Genet. 2019, 51, 445–451. [Google Scholar] [CrossRef]
- Moruzzi, G.; Magoun, H.W. Brain stem reticular formation and activating of the EEG. Electroencephalogr. Clin. Neurophysiol. 1949, 1, 455–473. [Google Scholar] [CrossRef]
- Kubicki, S.; Herrmann, W.M.; Fichte, K.; Freund, G. Reflections on the topics—EEG Frequency bands and regulation of vigilance. Pharmakopsychiatr. Neuropsychopharmakol. 1979, 12, 237–245. [Google Scholar] [CrossRef]
- Cantero, J.L.; Atienza, M.; Gómez, C.M.; Salas, R.M. Spectral structure and brain mapping of human alpha activities in different arousal states. Neuropsychobiology 1999, 39, 110–116. [Google Scholar] [CrossRef]
- Steriade, M. Brainstem activation of thalamocortical systems. Brain Res. Bull. 1999, 50, 391–392. [Google Scholar] [CrossRef]
- Ng, S.C.; Raveendran, P. EEG peak alpha frequency as an indicator for physical fatigue. IFMBE Proc. 2007, 16, 517–520. [Google Scholar]
- Huang, J.; Sander, C.; Jawinski, P.; Ulke, C.; Spada, J.; Hegerl, U.; Hensch, T. Test-retest reliability of brain arousal regulation as assessed with VIGALL 2.0. Neuropsychiatr. Electrophysiol. 2015, 1, 13. [Google Scholar] [CrossRef]
- Stoppe, M.; Meyer, K.; Schlingmann, M.; Olbrich, S.; Bergh, F.T. Hyperstable arousal regulation in multiple sclerosis. Psychoneuroendocrinology 2019, 110, 104417. [Google Scholar] [CrossRef]
- Barry, R.J.; De Blasio, F.M.; Fogarty, J.S.; Clarke, A.R. Natural alpha frequency components in resting EEG and their relation to arousal. Clin. Neurophysiol. 2020, 131, 205–212. [Google Scholar] [CrossRef]
- Malone, S.M.; Burwell, S.J.; Vaidyanathan, U.; Miller, M.B.; McGue, M.; Iacono, W.G. Heritability and molecular-genetic basis of resting EEG activity: A genome-wide association study. Psychophysiology 2014, 51, 1225–1245. [Google Scholar] [CrossRef] [Green Version]
- Valera, F.J.; Toro, A.; John, E.R.; Schwartz, E.L. Perceptual framing and cortical alpha rhythm. Neuropsychologia 1981, 19, 675–686. [Google Scholar] [CrossRef]
- Busch, N.A.; Dubois, J.; VanRullen, R. The phase of ongoing EEG oscillations predicts visual perception. J. Neurosci. 2009, 29, 7869–7876. [Google Scholar] [CrossRef] [PubMed]
- Lorincz, M.L.; Kékesi, K.A.; Juhász, G.; Crunelli, V.; Hughes, S.W. Temporal framing of thalamic relay-mode firing by phasic inhibition during the alpha rhythm. Neuron 2009, 63, 683–696. [Google Scholar] [CrossRef] [PubMed]
- Cecere, R.; Rees, G.; Romei, V. Individual differences in alpha frequency drive crossmodal illusory perception. Curr. Biol. 2015, 25, 231–235. [Google Scholar] [CrossRef]
- Samaha, J.; Postle, B.R. The speed of alpha-band oscillations predicts the temporal resolution of visual perception. Curr. Biol. 2015, 25, 2985–2990. [Google Scholar] [CrossRef]
- Callaway, E.; Yeager, C.L. Relationship between reaction time and electroencephalographic alpha phase. Science 1960, 132, 1765–1766. [Google Scholar] [CrossRef]
- Surwillo, W.W. Frequency of the “alpha” rhythm, reaction time and age. Nature 1961, 191, 823–824. [Google Scholar] [CrossRef]
- Surwillo, W.W. The relation of simple response time to brain wave frequency and the effects of age. Electroencephalogr. Clin. Neurophysiol. 1963, 15, 105–114. [Google Scholar] [CrossRef]
- Surwillo, W.W. The relation of decision time to brain wave frequency and to age. Electroencephalogr. Clin. Neurophysiol. 1964, 16, 510–514. [Google Scholar] [CrossRef]
- Surwillo, W.W. The electroencephalogram in the prediction of human reaction time during growth and development. Biol. Psychol. 1975, 3, 79–90. [Google Scholar] [CrossRef]
- Woodruff, D.S. Relationships among EEG alpha frequency, reaction time, and age: A biofeedback study. Psychophysiology 1975, 12, 673–681. [Google Scholar] [CrossRef] [PubMed]
- Klimesch, W.; Doppelmayr, M.; Schimke, H.; Pachinger, T. Alpha frequency, reaction time, and the speed of processing information. J. Clin. Neurophysiol. 1996, 13, 511–518. [Google Scholar] [CrossRef]
- Jin, Y.; O’Halloran, J.; Plon, L.; Sandman, C.; Potkin, S. Alpha EEG predicts visual reaction time. Int. J. Neurosci. 2006, 116, 1035–1044. [Google Scholar] [CrossRef] [PubMed]
- Kostyunina, M.B.; Kulikov, M.A. Frequency characteristics of EEG spectra in the emotions. Neurosci. Behav. Physiol. 1996, 26, 340–343. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, G.; Copello, F.; Vitali, P.; Perego, G.; Nobili, F. EEG spectral profile to stage Alzheimer’s disease. Clin. Neurophysiol. 1999, 110, 1831–1837. [Google Scholar] [CrossRef]
- Lansbergen, M.M.; Arns, M.; van Dongen-Boomsma, M.; Spronk, D.; Buitelaar, J.K. The increase in theta/beta ratio on resting-state EEG in boys with attention deficit/hyperactivity disorder is mediated by slow alpha peak frequency. Prog. Neuropsychopharmacol. Biol. Psychiatry 2011, 35, 47–52. [Google Scholar] [CrossRef]
- Northoff, G.; Duncan, N.W. How do abnormalities in the brain’s spontaneous activity translate into symptoms in schizophrenia? From an overview of resting state activity findings to a proposed spatiotemporal psychopathology. Prog. Neurobiol. 2016, 145–146, 26–45. [Google Scholar] [CrossRef]
- Yeum, T.-S.; Kang, U.G. Reduction in alpha peak frequency and coherence on quantitative electroencephalography in patients with schizophrenia. J. Korean Med. Sci. 2018, 33, e179. [Google Scholar] [CrossRef]
- Murphy, M.; Öngür, D. Decreased peak alpha frequency and impaired visual evoked potentials in first episode psychosis. Neuroimage Clin. 2019, 22, 101693. [Google Scholar] [CrossRef]
- Fuchs, T. Temporality and psychopathology. Phenom. Cogn. Sci. 2013, 12, 75–104. [Google Scholar] [CrossRef]
- Stanghellini, G.; Ballerini, M.; Presenza, S.; Mancini, M.; Northoff, G.; Cutting, J. Abnormal time experiences in major depression: An empirical qualitative study. Psychopathology 2017, 50, 125–140. [Google Scholar] [CrossRef] [PubMed]
- Northoff, G.; Magioncalda, P.; Martino, M.; Lee, H.-C.; Tseng, Y.-C.; Lane, T. Too fast or too slow? Time and neuronal variability in bipolar disorder-A combined theoretical and empirical investigation. Schizophr. Bull. 2018, 44, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Ronconi, L.; Busch, N.A.; Melcher, D. Alpha-band sensory entrainment alters the duration of temporal windows in visual perception. Sci. Rep. 2018, 8, 11810. [Google Scholar] [CrossRef] [PubMed]
- Chota, S.; VanRullen, R. Visual entrainment at 10 Hz causes periodic modulation of the flash lag illusion. Front. Neurosci. 2019, 13, 232. [Google Scholar] [CrossRef] [PubMed]
- Surwillo, W.W. Timing of behavior in senescence and the role of the central nervous system. In Human Aging and Behavior; Talland, G.A., Ed.; Academic: New York, NY, USA, 1968; pp. 1–35. [Google Scholar]
- Surwillo, W.W.; Titus, T.G. Reaction time and the psychological refractory period in children and adults. Dev. Psychobiol. 1976, 9, 517–527. [Google Scholar] [CrossRef]
- Wedensky, N.E. Excitation, inhibition and narcosis. In Collected Works; Rusinov, V.S., Ed.; LGU: Leningrad, Russia, 1953; Volume 4, pp. 517–679. [Google Scholar]
- Golikov, N.V. Physiological Lability and Its Changes in Basic Nervous Processes; LGU: Leningrad, Russia, 1950. [Google Scholar]
- Posner, M. The attention system of the human brain. Ann. Neurosci. 1989, 13, 25–42. [Google Scholar] [CrossRef]
- Cisler, J.M.; Koster, E.H.W. Mechanisms of attentional biases towards threat in the anxiety disorders: An integrative review. Clin. Psychol. Rev. 2010, 30, 203–216. [Google Scholar] [CrossRef]
- Inanaga, K. Frontal midline theta rhythm and mental activity. Psychiatry Clin. Eurosc. 1998, 52, 555–566. [Google Scholar] [CrossRef]
- Martino, M.; Magioncalda, P.; Huang, Z.; Conio, B.; Piaggio, N.; Duncan, N.W.; Rocchi, G.; Escelsior, A.; Marozzi, V.; Wolff, A.; et al. Contrasting variability patterns in the default mode and sensorimotor networks balance in bipolar depression and mania. Proc. Natl. Acad. Sci. USA 2016, 113, 4824–4829. [Google Scholar] [CrossRef]
- Northoff, G. The brain’s spontaneous activity and its psychopathological symptoms—“Spatiotemporal binding and integration”. Prog. Neuropsychopharmacol. Biol. Psychiatry 2018, 80 Pt B, 81–90. [Google Scholar] [CrossRef]
- Northoff, G.; Tumati, S. ‘Average is good, Extremes are bad’-Non-linear inverted U-shaped relationship between neural mechanisms and functionality of mental features. Neurosci. Biobehav. Rev. 2019, 104, 11–25. [Google Scholar] [CrossRef] [PubMed]
- Kao, S.-C.; Huang, C.-J.; Hung, T.-M. Frontal midline theta is a specific indicator of optimal Attentional engagement during skilled putting performance. J. Sport Exerc. Psychol. 2013, 35, 470–478. [Google Scholar] [CrossRef] [PubMed]
- Marchand, W.R. Self-referential thinking, suicide, and function of the cortical midline structures and striatum in mood disorders: Possible implications for treatment studies of mindfulness-based interventions for bipolar depression. Depress. Res. Treat. 2012, 2012, 246725. [Google Scholar] [CrossRef]
- Northoff, G. Spatiotemporal psychopathology I: No rest for the brain’s resting state activity in depression? Spatiotemporal psychopathology of depressive symptoms J. Affect. Disord. 2016, 190, 854–866. [Google Scholar] [CrossRef] [PubMed]
- Aftanas, L.l.; Golocheikine, S.A. Human anterior and frontal midline theta and lower alpha refect emotionally positive state and internalized attention: High-resolution EEG investigation of meditation. Neurosci. Lett. 2001, 310, 57–60. [Google Scholar] [CrossRef]
- Kubota, Y.; Sato, W.; Toichi, M.; Murai, T.; Okada, T.; Hayashi, A.; Sengoku, A. Frontal midline theta rhythm is correlated with cardiac autonomic activities during the performance of an attention demanding meditation procedure. Brain Res. Cogn. Brain Res. 2001, 11, 281–287. [Google Scholar] [CrossRef]
- Cahn, B.R.; Polich, J. Meditation states and traits: EEG, ERP, and neuroimaging studies. Psychol. Bull. 2006, 132, 180–211. [Google Scholar] [CrossRef]
- Klimesch, W.; Sauseng, P.; Hanslmayr, S. EEG alpha oscillations: The inhibition-timing hypothesis. Brain Res. Rev. 2007, 53, 63–88. [Google Scholar] [CrossRef]
- Doppelmayr, M.; Finkenzeller, T.; Sauseng, P. Frontal midline theta in the pre shot phase of rifle shooting: Differences between experts and novice. Neuropsychologia 2008, 46, 1463–1467. [Google Scholar] [CrossRef]
- Fox, J.J.; Snyder, A.C. The role of alpha-band brain oscillations as a sensory suppression mechanism during selective attention. Front. Psychol. 2011, 2, 154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konareva, I.N. Locus of psychological control and characteristics of the EEG frequency components. Neurophysiology 2011, 43, 534–542. [Google Scholar] [CrossRef]
- Arns, M.; Etkin, A.; Hegerl, U.; Williams, L.M.; DeBattista, C.; Palmer, D.M.; Fitzgerald, P.B.; Harris, A.; deBeuss, R.; Gordon, E. Frontal and rostral anterior cingulate (rACC) theta EEG in depression: Implications for treatment outcome? Eur. Neuropsychopharmacol. 2015, 25, 1190–1200. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.E.; Reznik, S.J.; Stewart, J.L.; Allen, J.J.B. Assessing and conceptualizing frontal EEG asymmetry: An updated primer on recording, processing, analyzing, and interpreting frontal alpha asymmetry. Int. J. Psychophysiol. 2017, 111, 98–114. [Google Scholar] [CrossRef]
- Wheeler, R.E.; Davidson, R.J.; Tomarken, A.J. Frontal brain asymmetry and emotional reactivity: A biological substrate of affective style. Psychophysiology 1993, 30, 82–89. [Google Scholar] [CrossRef]
- Davidson, R.J. Affective style and affective disorders: Perspectives from affective neuroscience. Cogn. Emot. 1998, 12, 307–330. [Google Scholar] [CrossRef]
- Blackhart, G.C.; Minnix, J.A.; Kline, J.P. Can EEG asymmetry patterns predict future development of anxiety and depression? Biol. Psychol. 2006, 72, 46–50. [Google Scholar] [CrossRef]
- Nusslock, R.; Shackman, A.J.; Harmon-jones, E.; Alloy, L.B.; Coan, J.A.; Abramson, L.Y. Cognitive vulnerability and frontal brain asymmetry: Common predictors of first prospective depressive episode. J. Abnorm. Psychol. 2011, 120, 497–503. [Google Scholar] [CrossRef]
- Gollan, J.K.; Hoxha, D.; Chihade, D.; Pflieger, M.E.; Rosebrock, L.; Cacioppo, J. Frontal alpha EEG asymmetry before and after behavioral activation treatment for depression. Biol. Psychol. 2014, 99, 198–208. [Google Scholar] [CrossRef]
- Lahey, B.B.; Krueger, R.F.; Rathouz, P.J.; Waldman, I.D.; Zald, D.H. A hierarchical causal taxonomy of psychopathology across the life span. Psychol. Bull. 2017, 143, 142–186. [Google Scholar] [CrossRef]
- Heller, W. Neuropsychological mechanisms of individual differences in emotion, personality, and arousal. Neuropsychology 1993, 7, 476–489. [Google Scholar] [CrossRef]
- Quaedflieg, C.W.E.M.; Meyer, T.; Smulders, F.T.Y.; Smeets, T. The functional role of individual-alpha based frontal asymmetry in stress responding. Biol. Psychol. 2015, 104, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Coan, J.A.; Allen, J.J.; Harmon-Jones, E. Voluntary facial expression and hemispheric asymmetry over the frontal cortex. Psychophysiology 2001, 38, 912–925. [Google Scholar] [CrossRef] [PubMed]
- Thibodeau, R.; Jorgensen, R.S.; Kim, S. Depression, anxiety, and resting frontal EEG asymmetry: A meta-analytic review. J. Abnorm. Psychol. 2006, 115, 715–729. [Google Scholar] [CrossRef] [PubMed]
- White, J.; Kivimaki, M.; Jokela, M.; Batty, G.D. Association of inflammation with specific symptoms of depression in a general population of older people: The English Longitudinal Study of Ageing. Brain Behav. Immun. 2017, 61, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Dawson, G.; Frey, K.; Panagiotides, H.; Osterling, J.; Hessl, D. Infants of depressed mothers exhibit atypical frontal brain activity: A replication and extension of previous findings. J. Child Psychol. Psychiatr. 1997, 38, 179–186. [Google Scholar] [CrossRef]
- Robinson, R.G.; Downhill, J.E. Lateralization of psychopathology in response to focal brain injury. In Brain Asymmetry; Davidson, R.J., Hugdahl, K., Eds.; The MIT Press: Cambridge, MA, USA, 1995; pp. 693–711. [Google Scholar]
- Davidson, R.J. Affective style, psychopathology, and resilience: Brain mechanisms and plasticity. Am. Psychol. 2000, 55, 1196–1214. [Google Scholar] [CrossRef]
- Harmon-Jones, E.; Sigelman, J. State anger and prefrontal brain activity: Evidence that insult-related relative left-prefrontal activation is associated with experienced anger and aggression. J. Personal. Soc. Psychol. 2001, 80, 797–803. [Google Scholar] [CrossRef]
- Harmon-Jones, E.; Peterson, C.K.; Harris, C.R. Jealousy: Novel methods and neural correlates. Emotion 2009, 9, 113–117. [Google Scholar] [CrossRef]
- Drake, R.A.; Ulrich, G. Line bisecting as a predictor of personal optimism and desirability of risky behaviors. Acta Psychol. 1992, 79, 219–226. [Google Scholar] [CrossRef]
- De Pascalis, V.; Cozzuto, G.; Caprara, G.V.; Alessandri, G. Relations among EEG alpha asymmetry, BIS/BAS, and dispositional optimism. Biol. Psychol. 2013, 94, 198–209. [Google Scholar] [CrossRef] [PubMed]
- Terzian, H. Behavioural and EEG effects of intracarotid sodium amytal injection. Acta Neurochir. 1964, 12, 230–239. [Google Scholar] [CrossRef] [PubMed]
- Gainotti, G. Emotional behavior and hemispheric side of the lesion. Cortex 1972, 8, 41–55. [Google Scholar] [CrossRef]
- Henriques, J.B.; Davidson, R.J. Left Frontal Hypoactivation in Depression. J. Abnorm. Psychol. 1991, 100, 535–545. [Google Scholar] [CrossRef]
- Davidson, R.J. Asymmetric brain function, affective style, and psychopathology: The role of early experience and plasticity. Dev. Psychopathol. 1994, 6, 741–758. [Google Scholar] [CrossRef]
- Coan, J.A.; Allen, J.J.B. Frontal EEG asymmetry as a moderator and mediator of emotion. Biol. Psychol. 2004, 67, 7–49. [Google Scholar] [CrossRef]
- Schutter, D.J.L.G. Antidepressant efficacy of high-frequency transcranial magnetic stimulation over the left dorsolateral prefrontal cortex in double-blind sham-controlled designs: A meta-analysis. Psychol. Med. 2009, 39, 65–75. [Google Scholar] [CrossRef]
- Choi, S.W.; Chi, S.E.; Chung, S.Y.; Kim, J.W.; Ahn, C.Y.; Kim, H.T. Is alpha wave neurofeedback effective with randomized clinical trials in depression? A pilot study. Neuropsychobiology 2010, 63, 43–51. [Google Scholar] [CrossRef]
- Schutter, D.J.L.G. Quantitative review of the efficacy of slow-frequency magnetic brain stimulation in major depressive disorder. Psychol. Med. 2010, 40, 1789–1795. [Google Scholar] [CrossRef]
- Stewart, J.L.; Bismark, A.W.; Towers, D.N.; Coan, J.A.; Allen, J.J. Resting frontal EEG asymmetry as an endophenotype for depression risk: Sex-specific patterns of frontal brain asymmetry. J. Abnorm. Psychol. 2010, 119, 502–512. [Google Scholar] [CrossRef]
- Hortensius, R.; Schutter, D.J.L.G.; Harmon-Jones, E. When anger leads to aggression: Induction of relative left frontal cortical activity with transcranial direct current stimulation increases the anger–aggression relationship. Soc. Cogn. Affect. Neurosci. 2012, 7, 342–347. [Google Scholar] [CrossRef] [PubMed]
- Smit, D.J.A.; Posthuma, D.; Boomsma, D.I.; De Geus, E.J. The relation between frontal EEG asymmetry and the risk for anxiety and depression. Biol. Psychol. 2007, 74, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Fishman, I.; Ng, R.; Bellugi, U. Do extraverts process social stimuli differently from introverts? Cogn. Neurosci. 2011, 2, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.A. The psychophysiological basis of introversion-extraversion. Behav. Res. Ther. 1970, 8, 249–266. [Google Scholar] [CrossRef]
- Carver, C.S.; White, T.L. Behavioral inhibition behavioral activation and affective responses to impending reward and punishment: The BIS/BAS scales. J. Personal. Soc. Psychol. 1994, 67, 319–333. [Google Scholar] [CrossRef]
- Matthews, G.; Gilliland, K. The personality theories of H.J. Eysenck and J.A. Gray: A comparative review. Personal. Individ. Differ. 1999, 26, 583–626. [Google Scholar] [CrossRef]
- Brocke, B.; Battmann, W. The arousal-activation theory of extraversion and neuroticism: A systematic analysis and principal conclusions. Adv. Behav. Res. Ther. 1992, 14, 211–246. [Google Scholar] [CrossRef]
- McAdams, D.P.; Pals, J.L. A new big five. Am. Psychol. 2006, 61, 204–217. [Google Scholar] [CrossRef]
- Yamasue, H.; Abe, O.; Suga, M.; Yamada, H.; Inoue, H.; Tochigi, M.; Rogers, M.; Aoki, S.; Kato, N.; Kasai, K. Gender-common and –specific neuroanatomical basis of human anxiety-related personality traits. Cereb. Cortex 2008, 18, 46–52. [Google Scholar] [CrossRef]
- Zuckerman, M. Sensation Seeking: Beyond the Optimal Level of Arousal; Distributed by the Halsted Press Division of Wiley; L. Erlbaum Associates: Hillsdale, NJ, USA, 1979; p. 449. [Google Scholar]
- Samuel, D.B.; Widiger, T.A. A meta-analytic review of the relationships between the five-factor model and DSM-IV-TR personality disorders: A facet level analysis. Clin. Psychol. Rev. 2008, 28, 1326–1342. [Google Scholar] [CrossRef] [PubMed]
- Watson, D.; Stasik, S.M.; Ellickson-Larew, S.; Stanton, K. Extraversion and psychopathology: A facet-level analysis. J. Abnorm. Psychol. 2015, 124, 432–446. [Google Scholar] [CrossRef] [PubMed]
- Watson, D.; Ellickson-Larew, S.; Stanton, K.; Levin-Aspenson, H.F.; Khoo, S.; Stasik-O’Brien, S.M.; Clark, L.A. Aspects of extraversion and their associations with psychopathology. J. Abnorm. Psychol. 2019, 128, 777–794. [Google Scholar] [CrossRef] [PubMed]
- Watson, D.; Stanton, K.; Khoo, S.; Ellickson-Larew, S.; Stasik-O’Brien, S.M. Extraversion and psychopathology: A multilevel hierarchical review. J. Res. Personal. 2019, 81, 1–10. [Google Scholar] [CrossRef]
- O’Gorman, J.G.; Lloyd, J.E.M. Extraversion, impulsiveness, and EEG alpha activity. Personal. Individ. Differ. 1987, 8, 169–174. [Google Scholar] [CrossRef]
- Wall, T.L.; Schuckit, M.A.; Mungas, D.; Ehlers, C.L. EEG alpha activity and personality traits. Alcohol 1990, 7, 461–464. [Google Scholar] [CrossRef]
- Knyazev, G.G. Is cortical distribution of spectral power a stable individual characteristic? Int. J. Psychophysiol. 2009, 72, 123–133. [Google Scholar] [CrossRef]
- Sampaio, A.; Soares, J.M.; Coutinho, J.; Sousa, N.; Goncalves, O.F. The Big Five default brain: Functional evidence. Brain Struct. Funct. 2014, 219, 1913–1922. [Google Scholar] [CrossRef]
- Ikeda, S.; Takeuchi, H.; Taki, Y.; Nouchi, R.; Yokoyama, R.; Kotozaki, Y.; Nakagawa, S.; Sekiguchi, A.; Iizuka, K.; Yamamoto, Y.; et al. A Comprehensive analysis of the correlations between resting-state oscillations in multiple-frequency bands and Big Five traits. Front. Hum. Neurosci. 2017, 11, 321. [Google Scholar] [CrossRef]
- Gianotti, L.R.R.; Dahinden, F.M.; Baumgartner, T.; Knoch, D. Understanding individual differences in domain-general prosociality: A resting EEG study. Brain Topogr. 2019, 32, 118–126. [Google Scholar] [CrossRef]
- Roslan, N.S.; Izhar, L.I.; Faye, I.; Amin, H.U.; Saad, M.N.M.; Sivapalan, S.; Karim, S.A.A.; Rahman, M.A. Neural correlates of eye contact in face-to-face verbal interaction: An EEG-based study of the extraversion personality trait. PLoS ONE 2019, 14, e0219839. [Google Scholar] [CrossRef]
- Martens, R.; Burton, D.; Vealey, R.; Bump, L.; Smith, D. The development of the competitive state anxiety inventory-2 (CSAI-2). In Competitive Anxiety in Sport; Martens, R., Vealey, R.S., Burton, D., Eds.; Human Kinetics: Chapaign, IL, USA, 1990; pp. 117–190. [Google Scholar]
- Clark, L.A.; Watson, D. Tripartite model of anxiety and depression: Psychometric evidence and taxonomic implications. J. Abnorm. Psychol. 1991, 100, 316–336. [Google Scholar] [CrossRef] [PubMed]
- Mathersul, D.; Williams, L.; Hopkinson, P.; Kemp, A. Investigating models of affect: Relationships among EEG alpha asymmetry, depression, and anxiety. Emotion 2008, 8, 560–572. [Google Scholar] [CrossRef] [PubMed]
- Barlow, D.H. Disorders of emotion. Psychol. Inq. 1991, 2, 58–71. [Google Scholar] [CrossRef]
- Watson, D. Rethinking the mood and anxiety disorders: A quantitative hierarchical model for DSM-V. J. Abnorm. Psychol. 2005, 114, 522–536. [Google Scholar] [CrossRef] [PubMed]
- Nusslock, R.; Shackman, A.; McMenamin, B.; Greischar, L.; Davidson, R.; Kovacs, M. Comorbid anxiety moderates the relationship between depression history and prefrontal EEG asymmetry. Psychophysiology 2018, 55, 12953. [Google Scholar] [CrossRef]
- Heller, W.; Nitschke, J.B.; Etienne, M.A.; Miller, G.A. Patterns of regional brain activity differentiate types of anxiety. J. Abnorm. Psychol. 1997, 106, 376. [Google Scholar] [CrossRef]
- Wiedemann, G.; Pauli, P.; Dengler, W.; Lutzenberger, W.; Birbaumer, N.; Buchkremer, G. Frontal brain asymmetry as a biological substrate of emotions in patients with panic disorders. Arch. Gen. Psychiatry 1999, 56, 78–84. [Google Scholar] [CrossRef]
- Knyazev, G.G.; Savostyanov, A.N.; Levin, E.A. Alpha oscillations as a correlate of trait anxiety. Int. J. Psychophysiol. 2004, 53, 147–160. [Google Scholar] [CrossRef]
- Knyazev, G.G.; Savostyanov, A.N.; Levin, E.A. Alpha synchronization and anxiety: Implications for inhibition vs. alertness hypotheses. Int. J. Psychophysiol. 2006, 59, 151–158. [Google Scholar] [CrossRef]
- Wise, V.; McFarlane, A.C.; Clark, C.R.; Battersby, M. An integrative assessment of brain and body function “at rest” in panic disorder: A combined quantitative EEG/autonomic function study. Int. J. Psychophysiol. 2011, 79, 155–165. [Google Scholar] [CrossRef]
- Yehuda, R.; Flory, J.D.; Pratchett, L.C.; Buxbaum, J.; Ising, M.; Holsboer, F. Putative biological mechanisms for the association between early life adversity and the subsequent development of PTSD. Psychopharmacology 2010, 212, 405–417. [Google Scholar] [CrossRef] [PubMed]
- Mehta, D.; Klengel, T.; Conneely, K.N.; Smith, A.K.; Altmann, A.; Pace, T.W.; Rex-Haffner, M.; Loeschner, A.; Gonik, M.; Mercer, K.B.; et al. Childhood maltreatment is associated with distinct genomic and epigenetic profiles in posttraumatic stress disorder. Proc. Natl. Acad. Sci. USA 2013, 110, 8302–8307. [Google Scholar] [CrossRef] [PubMed]
- Miller, B.F.; Seals, D.R.; Hamilton, K.L. A viewpoint on considering physiological principles to study stress resistance and resilience with aging. Ageing Res. Rev. 2017, 38, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Fleshner, M.; Maier, S.F.; Lyons, D.M.; Raskind, M.A. The neurobiology of the stress-resistant brain. Stress 2011, 14, 498–502. [Google Scholar] [CrossRef]
- McCrory, E.J.; De Brito, S.A.; Sebastian, C.L.; Mechelli, A.; Bird, G.; Kelly, P.A.; Viding, E. Heightened neural reactivity to threat in child victims of family violence. Curr. Biol. 2011, 21, R947–R948. [Google Scholar] [CrossRef]
- Zenkov, L.R.; Ronkin, M.A. Functional Diagnosis of Nervous Diseases; Medicine: Moscow, Russia, 1982; p. 432. [Google Scholar]
- Alonso, J.F.; Romero, S.; Ballester, M.R.; Antonijoan, R.M.; Mañanas, M.A. Stress assessment based on EEG univariate features and functional connectivity measures. Physiol. Meas. 2015, 36, 1351–1365. [Google Scholar] [CrossRef]
- Choi, Y.; Kim, M.; Chun, C. Measurement of occupants’ stress based on electroencephalograms (EEG) in twelve combined environments. Build. Environ. 2015, 88, 65–72. [Google Scholar] [CrossRef]
- Marshall, A.C.; Cooper, N.R.; Segrave, R.; Geeraert, N. The effects of long-term stress exposure on aging cognition: A behavioral and EEG investigation. Neurobiol. Aging 2015, 36, 2136–2144. [Google Scholar] [CrossRef]
- Al-Shargie, F.; Kiguchi, M.; Badruddin, N.; Dass, S.C.; Hani, A.F.M.; Tang, T.B. Mental stress assessment using simultaneous measurement of EEG and fNIRS. Biomed. Opt. Express 2016, 7, 3882–3898. [Google Scholar] [CrossRef]
- Stern, Y. What is cognitive reserve? Theory and research application of the reserve concept. J. Int. Neuropsychol. Soc. 2002, 8, 448–460. [Google Scholar] [CrossRef]
- Bartrés-Faz, D.; Arenaza-Urquijo, E.M. Structural and functional imaging correlates of cognitive and brain reserve hypotheses in healthy and pathological aging. Brain Topogr. 2011, 24, 340–357. [Google Scholar] [CrossRef] [PubMed]
- Medaglia, J.D.; Pasqualetti, F.; Hamilton, R.H.; Thompson-Schill, S.L.; Bassett, D.S. Brain and cognitive reserve: Translation via network control theory. Neurosci. Biobehav. Rev. 2017, 75, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Satz, P. Brain reserve capacity on symptom onset after brain injury: A formulation and review of evidence for threshold theory. Neuropsychology 1993, 7, 273–295. [Google Scholar] [CrossRef]
- Perneczky, R.; Green, R.C.; Kurz, A. Head circumference, atrophy, and cognition: Implications for brain reserve in Alzheimer disease. Neurology 2010, 75, 137–142. [Google Scholar] [CrossRef]
- López, M.E.; Aurtenetxe, S.; Pereda, E.; Cuesta, P.; Castellanos, N.P.; Bruña, R.; Niso, G.; Maestú, F.; Bajo, R. Cognitive reserve is associated with the functional organization of the brain in healthy aging: A MEG study. Front. Aging Neurosci. 2014, 6, 125. [Google Scholar]
- Stern, Y.; Habeck, C.; Moeller, J.; Scarmeas, N.; Anderson, K.E.; Hilton, H.J.; Flynn, J.; Sackeim, H.; van Heertum, R. Brain networks associated with cognitive reserve in healthy young and old adults. Cereb. Cortex 2005, 15, 394–402. [Google Scholar] [CrossRef]
- Fingelkurts, A.A.; Fingelkurts, A.A. Turning back the clock: A retrospective study on brain age change in response to nutraceuticals supplementation vs. lifestyle modifications. Mediterr. J. Nutr. Metab. 2022. [Google Scholar] [CrossRef]
- Fratiglioni, L.; Wang, H. Brain reserve hypothesis in dementia. J. Alzheimers. Dis. 2007, 12, 11–22. [Google Scholar] [CrossRef]
- Stern, Y. Cognitive reserve. Neuropsychologia 2009, 47, 2015–2028. [Google Scholar] [CrossRef]
- Cespón, J.; Miniussi, C.; Pellicciari, M.C. Interventional programs to improve cognition during healthy and pathological ageing: Cortical modulations and evidence for brain plasticity. Ageing Res. Rev. 2018, 43, 81–98. [Google Scholar] [CrossRef]
- Nunez, P.L.; Wingeier, B.M.; Silberstein, R.B. Spatial–temporal structures of human alpha rhythms: Theory, microcurrent sources, multiscale measurements, and global binding of local networks. Hum. Brain Mapp. 2001, 13, 125–164. [Google Scholar] [CrossRef] [PubMed]
- Jellinger, K.A. The pathology of ischemic-vascular dementia: An update. J. Neurol. Sci. 2002, 203–204, 153–157. [Google Scholar] [CrossRef]
- Dockree, P.M.; Kelly, S.P.; Foxe, J.J.; Reilly, R.B.; Robertson, I.H. Optimal sustained attention is linked to the spectral content of background EEG activity: Greater ongoing tonic alpha (10 Hz) power supports successful phasic goal activation. Eur. J. Neurosci. 2007, 25, 900–907. [Google Scholar] [PubMed]
- Arnau, S.; Möckel, T.; Rinkenauer, G.; Wascher, E. The interconnection of mental fatigue and aging: An EEG study. Int. J. Psychophysiol. 2017, 117, 17–25. [Google Scholar] [CrossRef]
- Zhirmunskaya, E.A.; Makarova, G.V. Relation of separate waves’ mean level of fronts’ asymmetry to structure of human EEG. In Functional States of the Brain; Sokolov, E.N., Danilova, N.N., Khomskaya, E.D., Eds.; Moscow University Press: Moscow, Russia, 1975; pp. 113–118. (In Russian) [Google Scholar]
- Raichle, M.E.; Snyder, A.Z. A default mode of brain function: A brief history of an evolving idea. NeuroImage 2007, 37, 1083–1090. [Google Scholar] [CrossRef]
- Deco, G.; Jirsa, V.K.; McIntosh, A.R. Emerging concepts for the dynamical organization of resting-state activity in the brain. Nat. Rev. Neurosci. 2011, 12, 43–56. [Google Scholar] [CrossRef]
- Allen, T.A.; DeYoung, C.G. Personality neuroscience and the five-factor model. In The Oxford Handbook of the Five-Factor Model; Widiger, T.A., Ed.; Oxford University Press: New York, NY, USA, 2017; pp. 319–352. [Google Scholar] [CrossRef]
- Klimesch, W. EEG-alpha rhythms and memory processes. Int. J. Psychophysiol. 1997, 26, 319–340. [Google Scholar] [CrossRef]
- Klimesch, W.; Freunberger, R.; Sauseng, P. Oscillatory mechanisms of process binding in memory. Neurosci. Biobehav. Rev. 2010, 34, 1002–1014. [Google Scholar] [CrossRef]
- Müller, V.I.; Langner, R.; Cieslik, E.C.; Rottschy, C.; Eickhoff, S.B. Interindividual differences in cognitive flexibility: Influence of gray matter volume, functional connectivity and trait impulsivity. Brain Struct. Funct. 2015, 220, 2401–2414. [Google Scholar] [CrossRef]
- Mattar, M.G.; Wymbs, N.F.; Bock, A.S.; Aguirre, G.K.; Grafton, S.T.; Bassett, D.S. Predicting future learning from baseline network architecture. NeuroImage 2018, 172, 107–117. [Google Scholar] [CrossRef]
- Ames, A.I. CNS energy metabolism as related to function. Brain Res. Rev. 2000, 34, 42–68. [Google Scholar] [PubMed]
- Lennie, P. The cost of cortical computation. Curr. Biol. 2003, 13, 493–497. [Google Scholar] [CrossRef]
- Raichle, M.E.; Mintun, M.A. Brain work and brain imaging. Annu. Rev. Neurosci. 2006, 29, 449–476. [Google Scholar] [CrossRef]
- Freeman, W.J. Origin, structure, and role of background EEG activity. Part 2. Analytic phase. Clin. Neurophysiol. 2004, 115, 2089–2107. [Google Scholar] [CrossRef]
- Laufs, H.; Krakow, K.; Sterzer, P.; Eger, E.; Beyerle, A.; Salek-Haddadi, A.; Kleinschmidt, A. Electroencephalographic signatures of attentional and cognitive default modes in spontaneous brain activity fluctuations at rest. Proc. Natl. Acad. Sci. USA 2003, 100, 11053–11058. [Google Scholar] [CrossRef] [PubMed]
- Fox, M.D.; Raichle, M.E. Spontaneous fluctuations in brain activity observed with functional magnetic resonance imaging. Nat. Rev. Neurosci. 2007, 8, 700–711. [Google Scholar]
- Landers, D.M.; Boutcher, S.H. Arousal-performance relationships. In Applied Sport Psychology: Personal Growth to Peak Performance, 2nd ed.; Williams, J.M., Ed.; Mayfield Publishing Co.: Mayfield, CA, USA, 1993; pp. 170–184. [Google Scholar]
- Hockey, G.R.J.; Hamilton, P. The cognitive patterning of stress states. In Stress and Fatigue in Human Performance; Hockey, G.R.T., Ed.; John Wiley & Sons: Chichester, UK, 1983; pp. 331–362. [Google Scholar]
- Pariitt, C.G.; Jones, J.G.; Hardy, L. Multidimensional anxiety and performance. In Stress and Performance in Sport; Jones, J.G., Hardy, L., Eds.; John Wiley & Sons: Chichester, UK, 1990; pp. 43–80. [Google Scholar]
- Could, D.; Weinberg, R.S. Sources of worry in junior elite wrestlers. J. Sport Behav. 1985, 8, 115–127. [Google Scholar]
- Fingelkurts, A.A.; Fingelkurts, A.A.; Neves, C.F.H. Neuro-assessment of leadership training. Coaching 2020, 13, 107–145. [Google Scholar] [CrossRef]
- Urry, H.L.; Nitschke, J.B.; Dolski, I.; Jackson, D.C.; Dalton, K.M.; Mueller, C.J.; Rosenkranz, M.A.; Ryff, C.D.; Singer, B.H.; Davidson, R.J. Making a life worth living: Neural correlates of well-being. Psychol. Sci. 2004, 15, 367–372. [Google Scholar] [CrossRef]
- King, M.L. The neural correlates of well-being: A systematic review of the human neuroimaging and neuropsychological literature. Cogn. Affect. Behav. Neurosci. 2019, 19, 779–796. [Google Scholar] [CrossRef]
- Cole, C.; Oetting, E.R.; Hinkle, J. Non-linearity of self-concept discrepancy: The value dimension. Psychol. Rep. 1967, 21, 58–60. [Google Scholar] [CrossRef] [PubMed]
- Sonstroem, R.J.; Bernardo, P. Intraindividual pregame state anxiety and basketball performance: A re-examination of the inverted-U curve. J. Sport Psychol. 1982, 4, 235–245. [Google Scholar] [CrossRef]
- Raglin, J.S.; Turner, P.E. Anxiety and performance in track and field athletes: A comparison of the inverted-U hypothesis with zone of optimal function theory. Personal. Individ. Differ. 1993, 14, 163–171. [Google Scholar] [CrossRef]
- Murphy, P.R.; Vandekerckhove, J.; Nieuwenhuis, S. Pupil-linked arousal determines variability in perceptual decision making. PLoS Comput. Biol. 2014, 10, e1003854. [Google Scholar] [CrossRef] [PubMed]
- McGinley, M.J.; David, S.V.; McCormick, D.A. Cortical membrane potential signature of optimal states for sensory signal detection. Neuron 2015, 87, 179–192. [Google Scholar] [CrossRef]
- McGinley, M.J.; Vinck, M.; Reimer, J.; Batista-Brito, R.; Zagha, E.; Cadwell, C.R.; Tolias, A.S.; Cardin, J.A.; McCormick, D.A. Waking state: Rapid variations modulate neural and behavioral responses. Neuron 2015, 87, 1143–1161. [Google Scholar] [CrossRef]
- Cavanagh, M. Mental-health issues and challenging clients in executive coaching. In Evidence-Based Coaching: Theory, Research and Practice from the Behavioural Sciences; Cavanagh, M., Grant, A.M., Kemp, T., Eds.; Australian Academic Press: Bowen Hills, QLD, Australia, 2005; Volume 1, pp. 21–36. [Google Scholar]
- Sokolov, E.N.; Danilova, N.N.; Khomskaya, E.D. Functional States of the Brain; Moscow University Press: Moscow, Russia, 1975. [Google Scholar]
- Cherniy, T.V. Application of method of EEG integral quantitative analysis for the estimation of zonal distinctions of electroencephalograms, laid in a concept of ‘ideal norm’. In Questions of Experimental and Clinical Medicine; Collection of Articles; 2010; Volume 14, pp. 116–129. [Google Scholar]
- Cona, G.; Koçillari, L.; Palombit, A.; Bertoldo, A.; Maritan, A.; Corbetta, M. Archetypes in human behavior and their brain correlates: An evolutionary trade-off approach. bioRxiv Prepr. 2018; first posted online Ma. 18. [Google Scholar] [CrossRef]
- Liberman, E.A.; Minina, S.V.; Shklovsky-Kordi, N.E. Quantum molecular computer model of the neuron and a pathway to the union of the sciences. BioSystems 1989, 22, 135–154. [Google Scholar] [CrossRef]
- Igamberdiev, A.U.; Shklovskiy-Kordi, N.E. The quantum basis of spatiotemporality in perception and consciousness. Prog. Biophys. Mol. Biol. 2017, 130 Pt A, 15–25. [Google Scholar] [CrossRef]
- Gorelick, P.B.; Furie, K.L.; Iadecola, C.; Smith, E.E.; Waddy, S.P.; Lloyd-Jones, D.M.; Bae, H.J.; Bauman, M.A.; Dichgans, M.; Duncan, P.W.; et al. Defining optimal brain health in adults: A presidential advisory from the American heart association/American stroke association. Stroke 2017, 48, e284–e303. [Google Scholar] [CrossRef]
- Rudrauf, D. Structure-Function relationships behind the phenomenon of cognitive resilience in neurology: Insights for neuroscience and medicine. Adv. Neurosci. 2014, 2014, 462765. [Google Scholar] [CrossRef]
- Thatcher, R.W.; Lubar, J.F. History of the scientific standards of QEEG normative databases. In Introduction to QEEG and Neurofeedback: Advanced Theory and Applications; Budzinsky, T., Budzinski, H., Evans, J., Abarbanel, A., Eds.; Academic Press: San Diego, CA, USA, 2008; pp. 29–62. [Google Scholar]
- Hellhammer, D.; Meinlschmidt, G.; Pruessner, J.C. Conceptual endophenotypes: A strategy to advance the impact of psychoneuroendocrinology in precision medicine. Psychoneuroendocrinology 2018, 89, 147–160. [Google Scholar] [CrossRef]
- McEwen, B.S. Stress, adaptation, and disease: Allostasis and allostatic load. Ann. N. Y. Acad. Sci. 1998, 840, 33–44. [Google Scholar] [CrossRef]
- Sterling, P.; Eyer, J. Allostasis: A new paradigm to explain arousal pathology. In Handbook of Life Stress, Cognition and Health; Fisher, S., Reason, J., Eds.; Wiley: Chichester, UK, 1988; pp. 629–649. [Google Scholar]
- McEwen, B.S. Allostasis and allostatic load: Implications for neuropsychopharmacology. Neuropsychopharmacology 2000, 22, 108–124. [Google Scholar] [CrossRef]
- McEwen, B.S.; Wingfield, J.C. The concept of allostasis in biology and biomedicine. Horm. Behav. 2003, 43, 2–15. [Google Scholar] [CrossRef]
- Anderzhanova, E.; Kirmeier, T.; Wotjak, C.T. Animal models in psychiatric research: The RDoC system as a new framework for endophenotype-oriented translational neuroscience. Neurobiol. Stress 2017, 7, 47–56. [Google Scholar] [CrossRef]
- Kalueff, A.V.; Ren-Patterson, R.F.; LaPorte, J.L.; Murphy, D.L. Domain interplay concept in animal models of neuropsychiatric disorders: A new strategy for high-throughput neurophenotyping research. Behav. Brain Res. 2008, 188, 243–249. [Google Scholar] [CrossRef]
- Kozel, F.A. Identifying phronotypes in psychiatry. Front. Psychiatry 2010, 1, 141, eCollection 2010. [Google Scholar] [CrossRef]
- Gunkelman, J. Transcend the DSM using phenotypes. Biofeedback 2006, 34, 95–98. [Google Scholar]
- Krueger, R.F.; Markon, K.E. Reinterpreting comorbidity: A model-based approach to understanding and classifying psychopathology. Annu. Rev. Clin. Psychol. 2006, 2, 111–133. [Google Scholar] [CrossRef]
- Insel, T.; Cuthbert, B.; Garvey, M.; Heinssen, R.; Pine, D.S.; Quinn, K.; Sanislow, C.; Wang, P. Research domain criteria (RDoC): Toward a new classification framework for research on mental disorders. Am. J. Psychiatry 2010, 167, 748–751. [Google Scholar] [CrossRef]
- Krueger, R.F.; Markon, K.E. A dimensional-spectrum model of psychopathology: Progress and opportunities. Arch. Gen. Psychiatry 2011, 68, 10–11. [Google Scholar] [CrossRef]
- Kapur, S.; Phillips, A.G.; Insel, T.R. Why has it taken so long for biological psychiatry to develop clinical tests and what to do about it? Mol. Psychiatry 2012, 17, 1174–1179. [Google Scholar] [CrossRef]
- Casey, B.J.; Craddock, N.; Cuthbert, B.N.; Hyman, S.E.; Lee, F.S.; Ressler, K.J. DSM-5 and RDoC: Progress in psychiatry research? Nat. Rev. Neurosci. 2013, 14, 810–814. [Google Scholar] [CrossRef]
- Kirmayer, L.J.; Crafa, D. What kind of science for psychiatry? Front. Hum. Neurosci. 2014, 8, 435. [Google Scholar] [CrossRef] [Green Version]




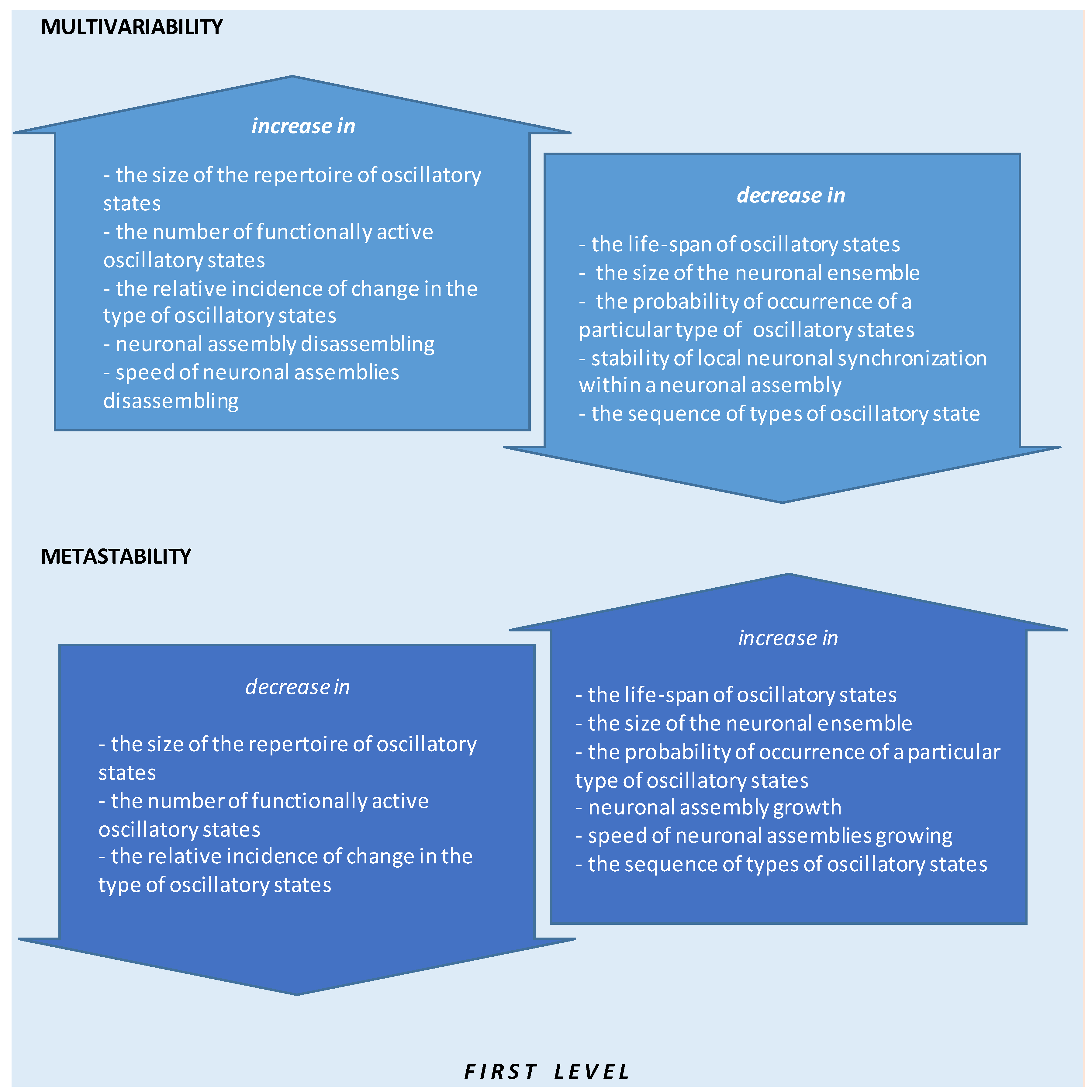{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dysfunction/Disorder | Sensitivity | Specificity | References |
|---|---|---|---|
| Cerebrovascular disease | >80% | [199] | |
| ADHD/ADD vs. normal children | 90% | 94% | [237] |
| ADD vs. DLD | 97% | 84.2% | [252] |
| LD vs. normal children | 72% | 80% | [253] |
| Depression | 72–93% | 75–88% | [254] |
| Panic disorder | 71% | 84% | [255] |
| Dementia | 91.9% | 92.2% | [256] |
| AD | 71–81% | [257,258] | |
| Declining to MCI | 95% | 94.1% | [259] |
| Converting to AD | 96.3% | 94.1% | [259] |
| Alcohol and drug abuse predicting relapse | 61% | 85% | [260] |
| mTBI vs. sTBI | 95.5% | 97.4% | [238,261] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fingelkurts, A.A.; Fingelkurts, A.A. Quantitative Electroencephalogram (qEEG) as a Natural and Non-Invasive Window into Living Brain and Mind in the Functional Continuum of Healthy and Pathological Conditions. Appl. Sci. 2022, 12, 9560. https://0-doi-org.brum.beds.ac.uk/10.3390/app12199560
Fingelkurts AA, Fingelkurts AA. Quantitative Electroencephalogram (qEEG) as a Natural and Non-Invasive Window into Living Brain and Mind in the Functional Continuum of Healthy and Pathological Conditions. Applied Sciences. 2022; 12(19):9560. https://0-doi-org.brum.beds.ac.uk/10.3390/app12199560
Chicago/Turabian StyleFingelkurts, Alexander A., and Andrew A. Fingelkurts. 2022. "Quantitative Electroencephalogram (qEEG) as a Natural and Non-Invasive Window into Living Brain and Mind in the Functional Continuum of Healthy and Pathological Conditions" Applied Sciences 12, no. 19: 9560. https://0-doi-org.brum.beds.ac.uk/10.3390/app12199560