Bacterial and Biodeterioration Analysis of the Waterlogged Wooden Lacquer Plates from the Nanhai No. 1 Shipwreck

 , , ,
, , ,
Abstract
:Featured Application
Abstract
1. Introduction
2. Materials and Methods
2.1. Water Sample Collection
2.2. High-Throughput Sequencing
2.3. Isolation of Dominant Bacteria
2.4. DNA Extraction and Sequencing of the 16S rRNA Gene
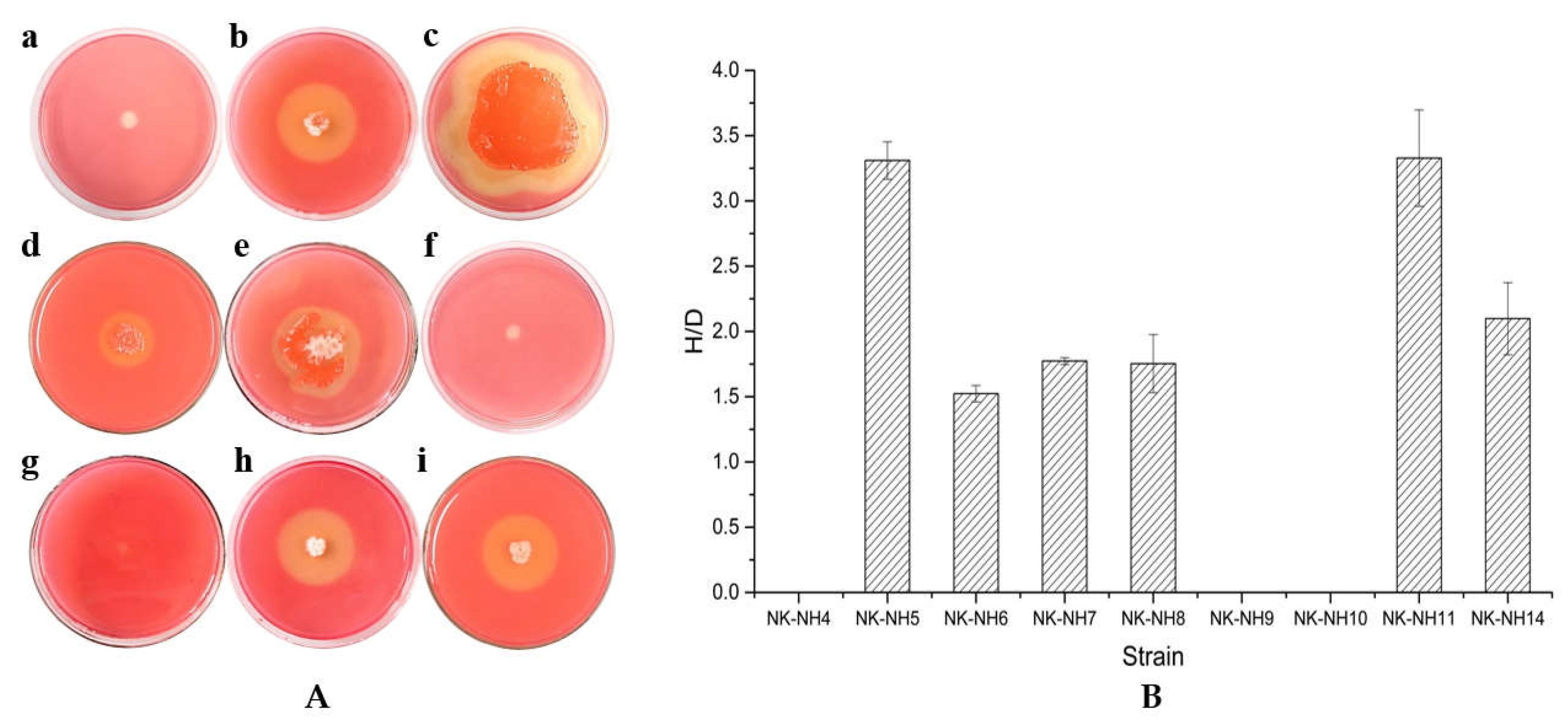
2.5. Determination of Ligninolytic Enzymatic Activity
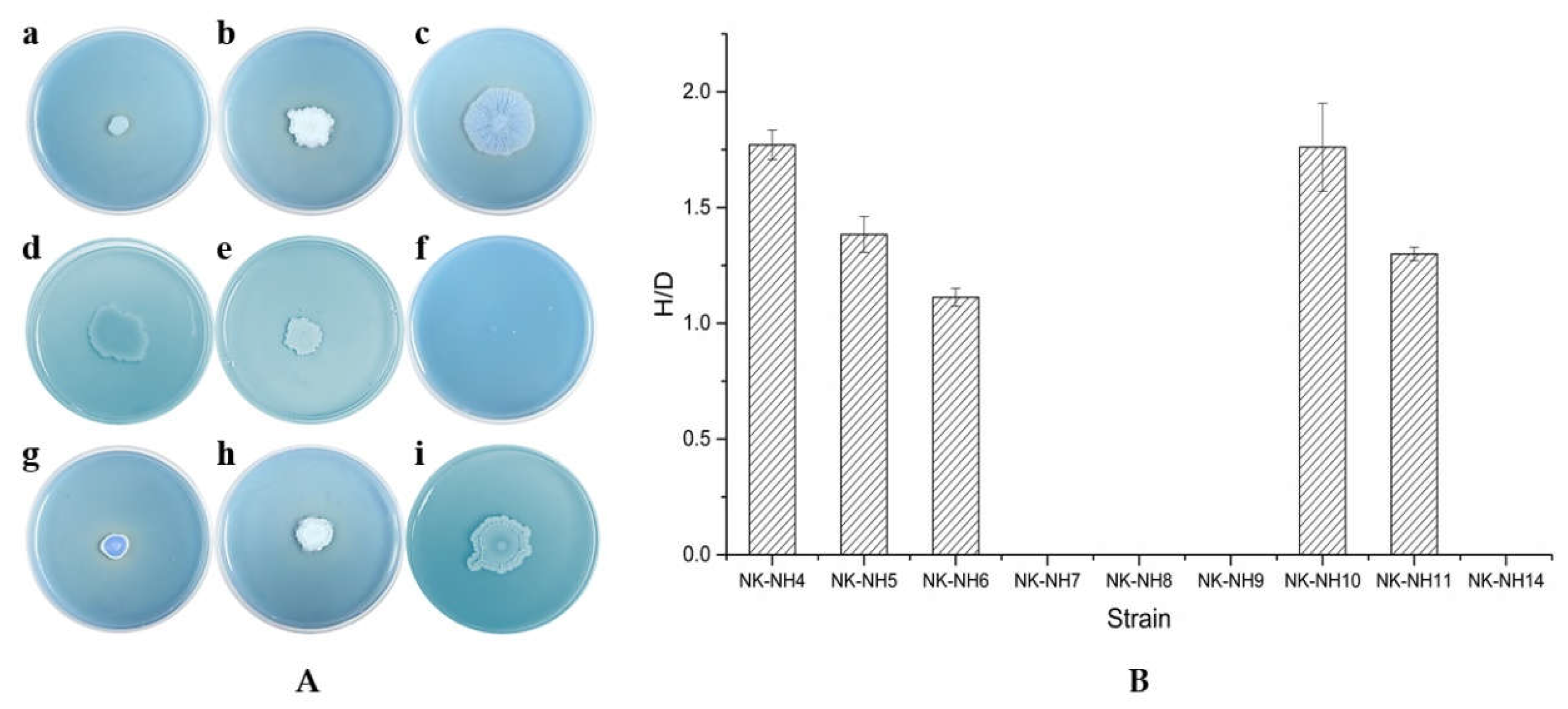
2.6. Determination of Cellulolytic Enzymatic Activity
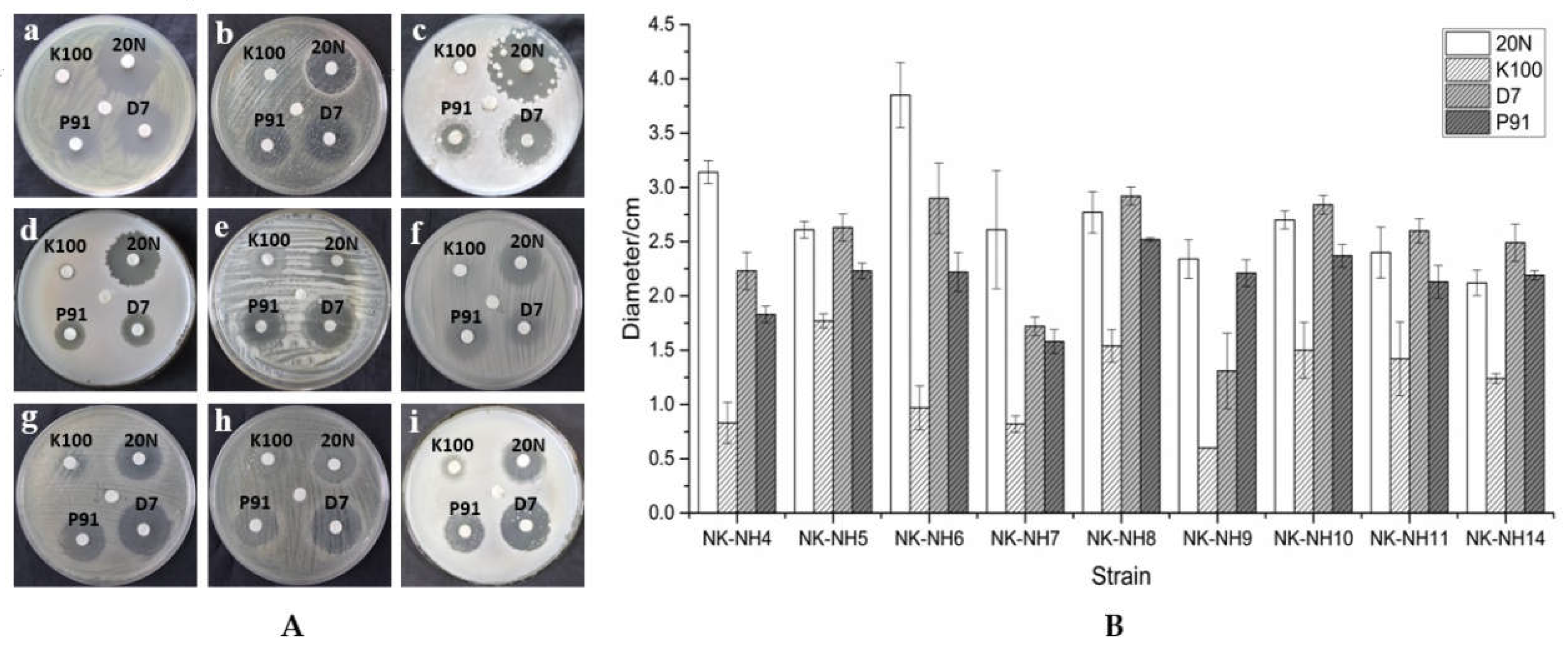
2.7. Determination of Biocide Effectiveness by Inhibition Assay
3. Results
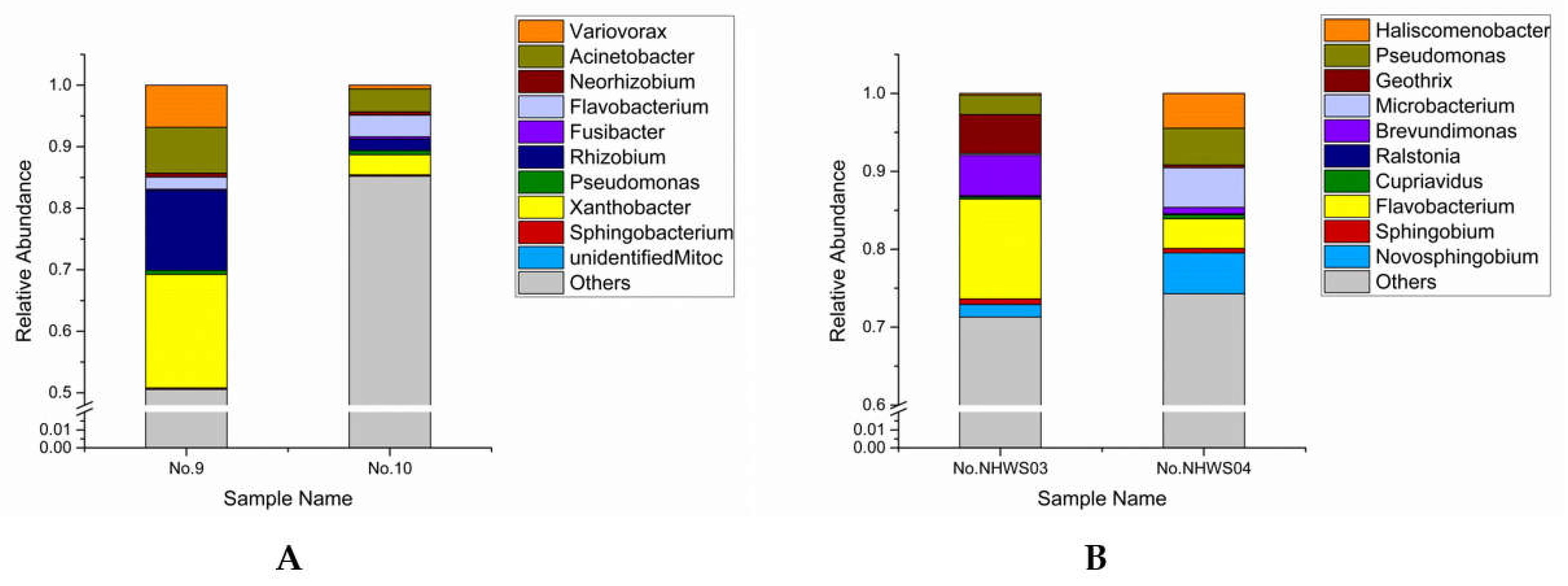
3.1. Bacterial Community Analyses
3.2. Isolation of Dominant Bacteria
3.3. Degradation of Cellulose and Lignin by Isolated Bacterial Strains
3.4. Biocide Susceptibility
4. Discussion
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Wang, Y.L.; Xiao, D.S. The excavation of the “Nanhai No. 1” shipwreck of the Song Dynasty in 2014. Archaeology 2016, 12, 56–83. [Google Scholar]
- Liu, Z.; Fu, T.; Hu, C.; Shen, D.; Macchioni, N.; Sozzi, L.; Chen, Y.; Liu, J.; Tian, X.; Ge, Q. Microbial community analysis and biodeterioration of waterlogged archaeological wood from the Nanhai No. 1 shipwreck during storage. Sci. Rep. 2018, 8, 7170. [Google Scholar] [CrossRef] [PubMed]
- Mcqueen, C.M.A.; Tamburini, D.; Łucejko, J.J.; Braovac, S.; Gambineri, F.; Modugno, F.; Colombini, M.P.; Kutzke, H. New insights into the degradation processes and influence of the conservation treatment in alum-treated wood from the Oseberg collection. Microchem. J. 2017, 132, 119–129. [Google Scholar] [CrossRef]
- Huang, J.H.; Yang, L.; Wang, L.Q.; Yan, S.M.; Wang, W.F. The conservation of a polychrome lacquer plate in archaeology site. Wenbo 2009, 6, 292–297. [Google Scholar]
- Yizheng Museum. The auspicial lacquer plate excavated from the Yizheng, Early Han cemetery in Yangzhou, Jiangsu. Cult. Southeast China 2007, 6, 19–22. [Google Scholar]
- Mo, Z. The preservation and the repairing of the lacquer plate from Zhangzheng Tomb of the Early Han Dynasty. Rep. Relics China 2018, 9, 1–6. [Google Scholar]
- Tamburini, D.; Łucejko, J.J.; Modugno, F.; Colombini, M.P. Combined pyrolysis-based techniques to evaluate the state of preservation of archaeological wood in the presence of consolidating agents. J. Anal. Appl. Pyrolysis 2016, 112, 429–441. [Google Scholar] [CrossRef]
- Chen, Y.H. The Separation, Identification and Degradation of Lignin of the Bamboo Slips of the Three Kingdoms Period. Master’s Thesis, Central South University, Changsha, China, 2009. [Google Scholar]
- Gao, M.G.; Zhang, Q.F.; Jin, T.; Luo, P.; Li, Q.; Xu, R.L. Observation and damage assessment of microbial diseases in some wooden cultural relics from the ancient marine shipwreck, Ningbo Xiaobaijiao No. 1. Sci. Conserv. Archaeol. 2017, 29, 93–102. [Google Scholar]
- Chen, J.C.; Huang, X.; Chen, X.L.; Chen, Z.X. Corrosion type and conservation of archaeological waterlogged wood. Mater. Rev. 2015, 29, 96–101. [Google Scholar]
- Tamburini, D.; Łucejko, J.J.; Zborowska, M.; Modugno, F.; Prądzyński, W.; Colombini, M.P. Archaeological wood degradation at the site of Biskupin (Poland): Wet chemical analysis and evaluation of specific Py-GC/MS profiles. J. Anal. Applied Pyrolysis 2015, 115, 7–15. [Google Scholar] [CrossRef]
- Sun, F. Diversity of bacteria isolated from the South China Sea sediments. Acta Microbiol. Sin. 2008, 48, 1578–1587. [Google Scholar]
- Abdellaoui, R.; Gouja, H.; Sayah, A.; Neffati, M. An efficient DNA extraction method for desert Calligonum species. Biochem. Genet. 2011, 49, 695. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.F.; Yu, Z.L. A strain of lignin-degrading bacteria’s screening and optimization of enzyme-producing condition. J. Agric. Sci. Technol. 2014, 16, 143–148. [Google Scholar]
- Ahmad, B.; Nigar, S.; Shah, S.S.A.; Bashir, S.; Ali, J.; Yousaf, S.; Bangash, J.A. Isolation and identification of cellulose degrading bacteria from municipal waste and their screening for potential antimicrobial activity. World Appl. Sci. J. 2013, 27, 1420–1426. [Google Scholar]
- Ji, H.W.; Zhang, L.; Liu, J.; Wang, F. Progress on analytical method for isothiazolinone derivatives used as industrial biocides. Chem. Reagents 2016, 38, 523–527. [Google Scholar]
- Liu, L. MicrobiaI corrosion and anti-corrosion measures for waterlogged wood objects. China Cult. Heritage Sci. Res. 2014, 4, 83–85. [Google Scholar]
- Powell, K.L.; Pedley, S.; Daniel, G.; Corfield, M. Ultrastructural observations of microbial succession and decay of wood buried at a Bronze Age archaeological site. Int. Biodeterior. Biodegradation 2001, 47, 165–173. [Google Scholar] [CrossRef]
- Cui, Z.; Lai, Q.; Dong, C.; Shao, Z. Biodiversity of polycyclic aromatic hydrocarbon-degrading bacteria from deep sea sediments of the Middle Atlantic Ridge. Environ. Microbiol. 2010, 10, 2138–2149. [Google Scholar] [CrossRef]
- Li, Y.H. Screening and identification of cellulose degradation bacteria. Mod. Agric. Sci. Technol. 2012, 2, 34–35. [Google Scholar]
- Gontang, E.A.; William, F.; Jensen, P.R. Phylogenetic diversity of gram-positive bacteria cultured from marine sediments. Appl. Environ. Microbiol. 2007, 73, 3272–3282. [Google Scholar] [CrossRef]
- Bao, Y.C.; Jin, L.M.; Chen, J.L.; Wang, G.T.; Wang, L.L.; Quan, C.S. Optimization of fermentation conditions of Marine Bacteria Ochrobactrum sp. LJY313. Food Ind. 2017, 11, 47–49. [Google Scholar]
- Bernardet, J.F.; Segers, P.; Vancanneyt, M.F.; Kersters, K.; Vandamme, P. Cutting a Gordian knot: Emended classification and description of the genus Flavobacterium, emended description of the family Flavobacteriaceae, and proposal of Flaviobacterium hydatis nom. nov. (basonym, Cytophaga aquatalis Strohl and Tait 1978). Int. J. Syst. Bacteriol. 1996, 46, 128–148. [Google Scholar] [CrossRef]
- Doughari, H.J.; Ndakidemi, P.A.; Human, I.S.; Benade, S. The ecology, biology and pathogenesis of Acinetobacter spp.: An overview. Microbes Environ. 2011, 26, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Garrity, G.M.; Brenner, D.J.; Krieg, N.R.; Staley, J.T. The proteobacteria, part C: The alpha-, beta-, delta-, and Epsilonproteobacteria. In Bergey’s Manual of Systematic Bacteriology; Bergey, D.H., Garrity, G.M., Brenner, D.J., Eds.; Springer: New York, NY, USA, 2005; Volume 2. [Google Scholar]
- Zheng, Y.K.; Liu, K.; Xiong, Z.J.; Miao, C.P.; Chen, Y.W.; Li-Hua, X.U.; Zhao, L.X. Effect of large-scale planting water hyacinth on cultivable bacterial community structure in the eutrophic lake. Microbiol. China 2015, 42, 42–53. [Google Scholar]
- Wu, G.; Xi, Y.; Zhao, Y.J. The latest development of research on Algae-Lysing bacteria. Res. Environ. Sci. 2002, 15, 43–46. [Google Scholar]
- Dong, L.; Yi, L.; Zheng, T. Advance in the research of marine algicidal functional bacteria and their algicidal mechanism. Adv. Earth Sci. 2013, 28, 243–252. [Google Scholar]
- Blanchette, R.A. A review of microbial deterioration found in archaeological wood from different environments. Int. Biodeterior. Biodegrad. 2000, 46, 189–204. [Google Scholar] [CrossRef]
- Huang, X.; Yang, T.; Wu, H.; Zhang, X.Y.; Li, J.; Zhou, X.; Xiao, L.; Zhao, K. Bacterial community structure in waterlogged bamboo slips excavated from the Laoguanshan Han Dynasty Tomb of Chengdu. J. Appl. Environ. Biol. 2018, 3, 1–9. [Google Scholar]
- Diaz-Herraiz, M.; Jurado, V.; Cuezva, S.; Laiz, L.; Pallecchi, P.; Tiano, P.; Sanchez-Moral, S.; Saiz-Jimenez, C. The actinobacterial colonization of Etruscan paintings. Sci. Rep. 2013, 3, 1440. [Google Scholar] [CrossRef]
- Björdal, C.G.; Nilsson, T.; Daniel, G. Microbial decay of waterlogged archaeological wood found in Sweden Applicable to archaeology and conservation. Int. Biodeterior. Biodegrad. 1999, 43, 63–73. [Google Scholar] [CrossRef]
- Fan, Q.Q.; Liu, Y.L.; Luo, Y.T.; Fu, R. Selection of cellulose degradation microbial floras in environment and analysis of degradation capability. Sichuan Chem. Ind. 2009, 12, 48–51. [Google Scholar]
- Song, W.Z. Biodegradation of Lignin by Composite Flora. Master’s Thesis, South China University of Technology, Guangzhou, China, 2014. [Google Scholar]
- Zhang, J.Y.; Gong, L.P.; Luo, Y.X.; Xu, W.Y.; Ling, J. Biodegradation of lignin in wheat straw by Alkaliphilic bacteria with compounded carbons. Environ. Sci. 2002, 23, 70–73. [Google Scholar] [CrossRef]
- Kumar, L.; Rathore, V.S.; Srivastava, H.S. (14)C-[lignin]-lignocellulose biodegradation by bacteria isolated from polluted soil. Indian J. Exp. Biol. 2001, 39, 584–589. [Google Scholar] [PubMed]
- Chen, J.; Chai, D.; Zhou, J.; Huang, X.; Chen, S. Shape recovery of collapsed archaeological wood ware with active alkali-urea treatment. J. Archaeol. Sci. 2009, 36, 434–440. [Google Scholar]
- Helms, A.C.; Martiny, A.C.; Hofman-Bang, J.; Ahring, B.K.; Kilstrup, M. Identification of bacterial cultures from archaeological wood using molecular biological techniques. Int. Biodeterior. Biodegrad. 2004, 53, 79–88. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, R.; Mcnamara, C.J. Cultural heritage microbiology. Tijdschr. Filos. 2010, 35, 917–920. [Google Scholar]
- Martin-Sanchez, P.M.; Nováková, A.; Bastian, F.; Alabouvette, C.; Saiz-Jimenez, C. Use of biocides for the control of fungal outbreaks in subterranean environments: The case of the Lascaux Cave in France. Environ. Sci. Technol. 2012, 46, 3762. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molecular Identification of Strains Isolated from the Water Samples | |||||
|---|---|---|---|---|---|
| Bacteria | Accession Number | Closet Relative Strain | Accession Number | Similarity (%) | Source |
| NK-NH4 | MH997669 | Microbacterium sp. | DQ658916.1 | 99% | water samples |
| NK-NH5 | MH997670 | Bacillus tequilensis | MG519475.1 | 93% | water samples |
| NK-NH6 | MH997671 | Bacillus subtilis | KY510927.1 | 99% | water samples |
| NK-NH7 | MH997672 | Pseudomonas sp. | MF436694.1 | 99% | water samples |
| NK-NH8 | MH997673 | Bacillus sp. | MG827113.1 | 99% | water samples |
| NK-NH9 | MH997674 | Ochrobactrum sp. | MF442305.1 | 99% | water samples |
| NK-NH10 | MH997675 | Bacillus megaterium | MG818943.1 | 99% | water samples |
| NK-NH11 | MH997676 | Bacillus velezensis | MH014956.1 | 99% | water samples |
| NK-NH14 | MH997679 | Stenotrophomonas sp. | MH144251.1 | 99% | water samples |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yin, L.; Jia, Y.; Wang, M.; Yu, H.; Jing, Y.; Hu, C.; Zhang, F.; Sun, M.; Liu, Z.; Chen, Y.; et al. Bacterial and Biodeterioration Analysis of the Waterlogged Wooden Lacquer Plates from the Nanhai No. 1 Shipwreck. Appl. Sci. 2019, 9, 653. https://0-doi-org.brum.beds.ac.uk/10.3390/app9040653
Yin L, Jia Y, Wang M, Yu H, Jing Y, Hu C, Zhang F, Sun M, Liu Z, Chen Y, et al. Bacterial and Biodeterioration Analysis of the Waterlogged Wooden Lacquer Plates from the Nanhai No. 1 Shipwreck. Applied Sciences. 2019; 9(4):653. https://0-doi-org.brum.beds.ac.uk/10.3390/app9040653
Chicago/Turabian StyleYin, Liuyu, Yin Jia, Mei Wang, Huanhuan Yu, Ying Jing, Cuiting Hu, Fengyu Zhang, Mingliang Sun, Zijun Liu, Yue Chen, and et al. 2019. "Bacterial and Biodeterioration Analysis of the Waterlogged Wooden Lacquer Plates from the Nanhai No. 1 Shipwreck" Applied Sciences 9, no. 4: 653. https://0-doi-org.brum.beds.ac.uk/10.3390/app9040653