Influence of Nitrogen Source and Growth Phase on Extracellular Biosynthesis of Silver Nanoparticles Using Cultural Filtrates of Scenedesmus obliquus

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microalgal Strain and Culture Conditions
2.2. Microalgal Growth Parameters
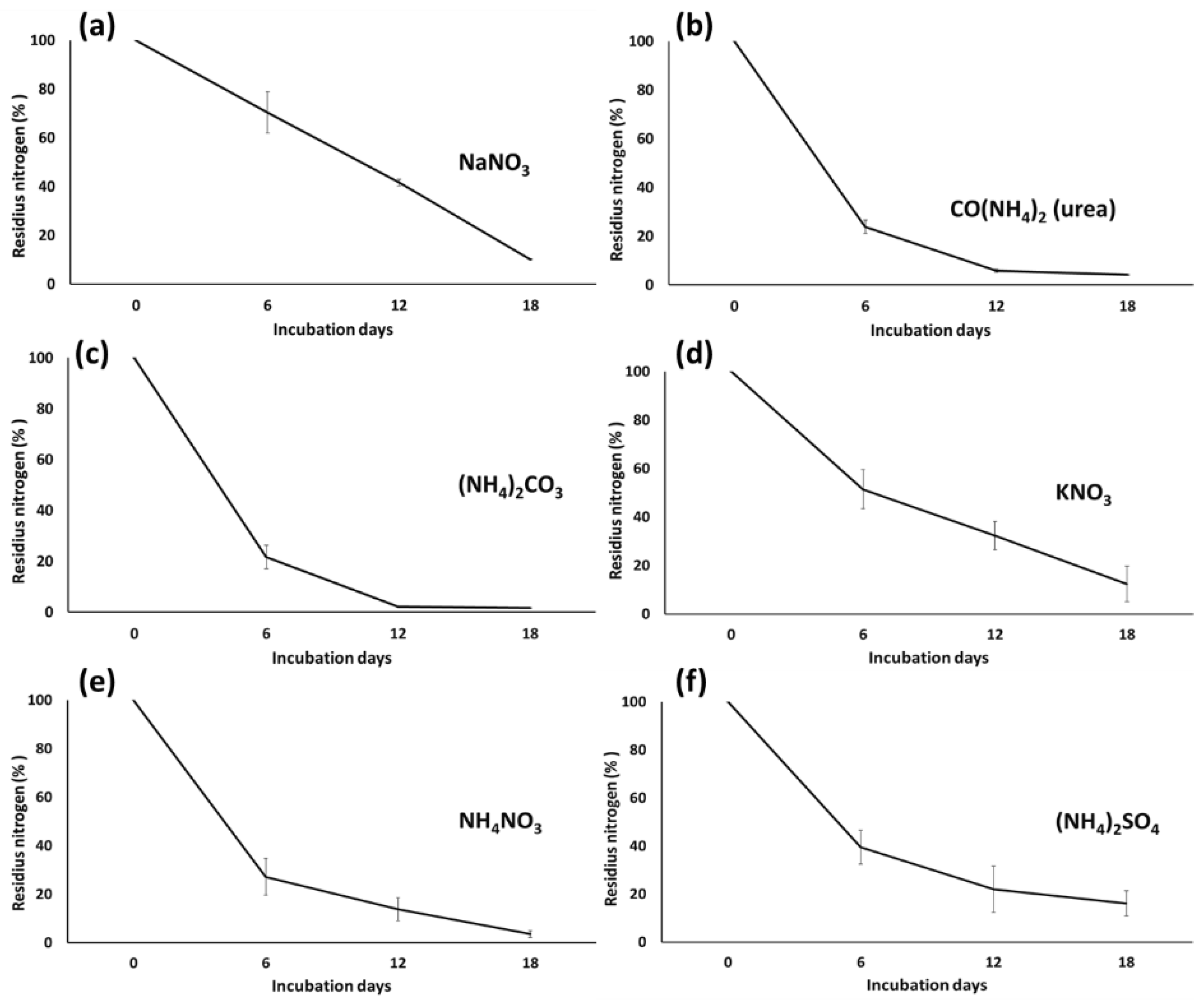
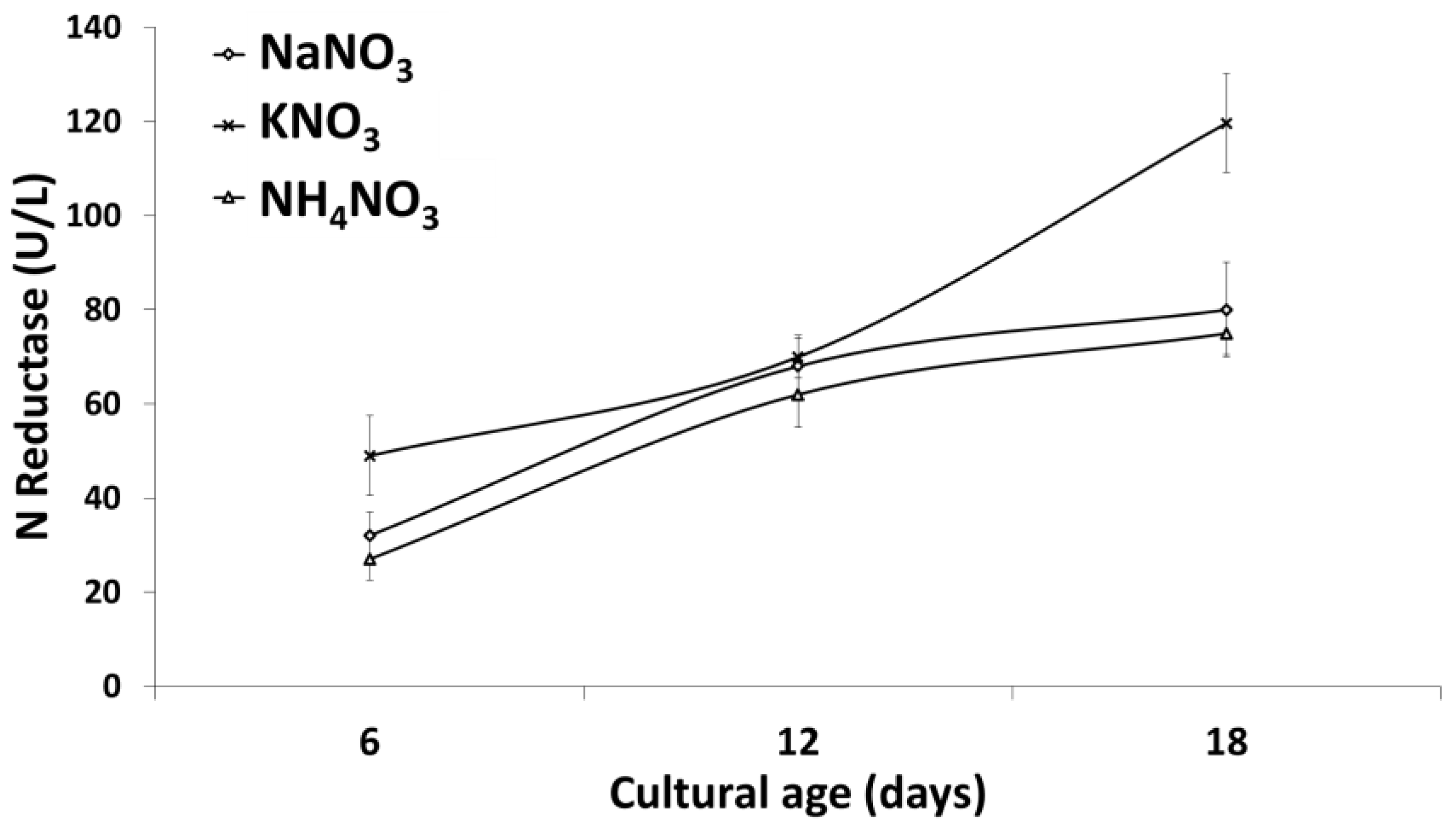
2.3. Nitrate Reductase Activity Assay
2.4. Extracellular Biosynthesis and Characterization of AgNPs
2.5. Antimicrobial Activity of AgNPs
2.6. Statistical Analysis
3. Results and Discussion
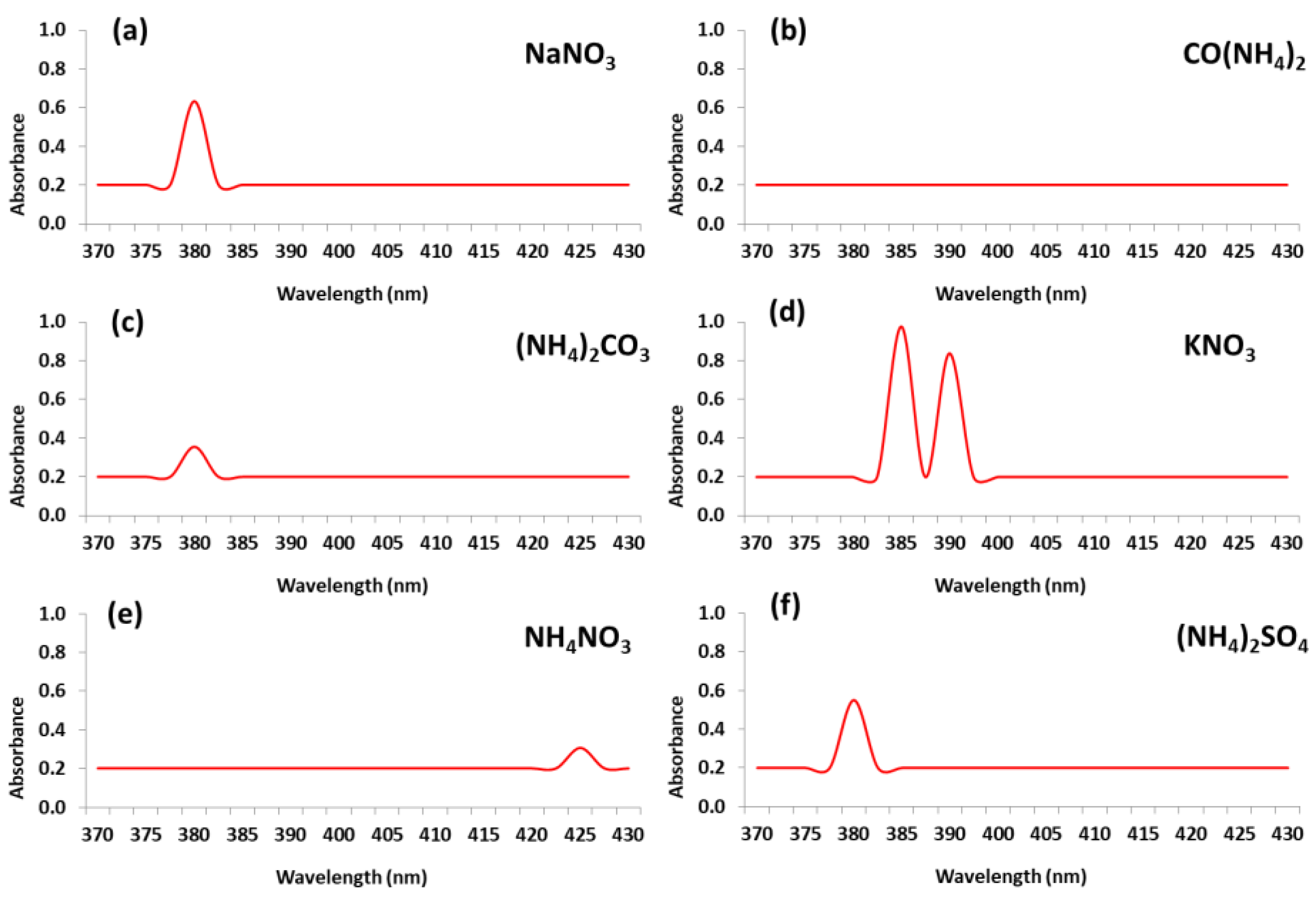
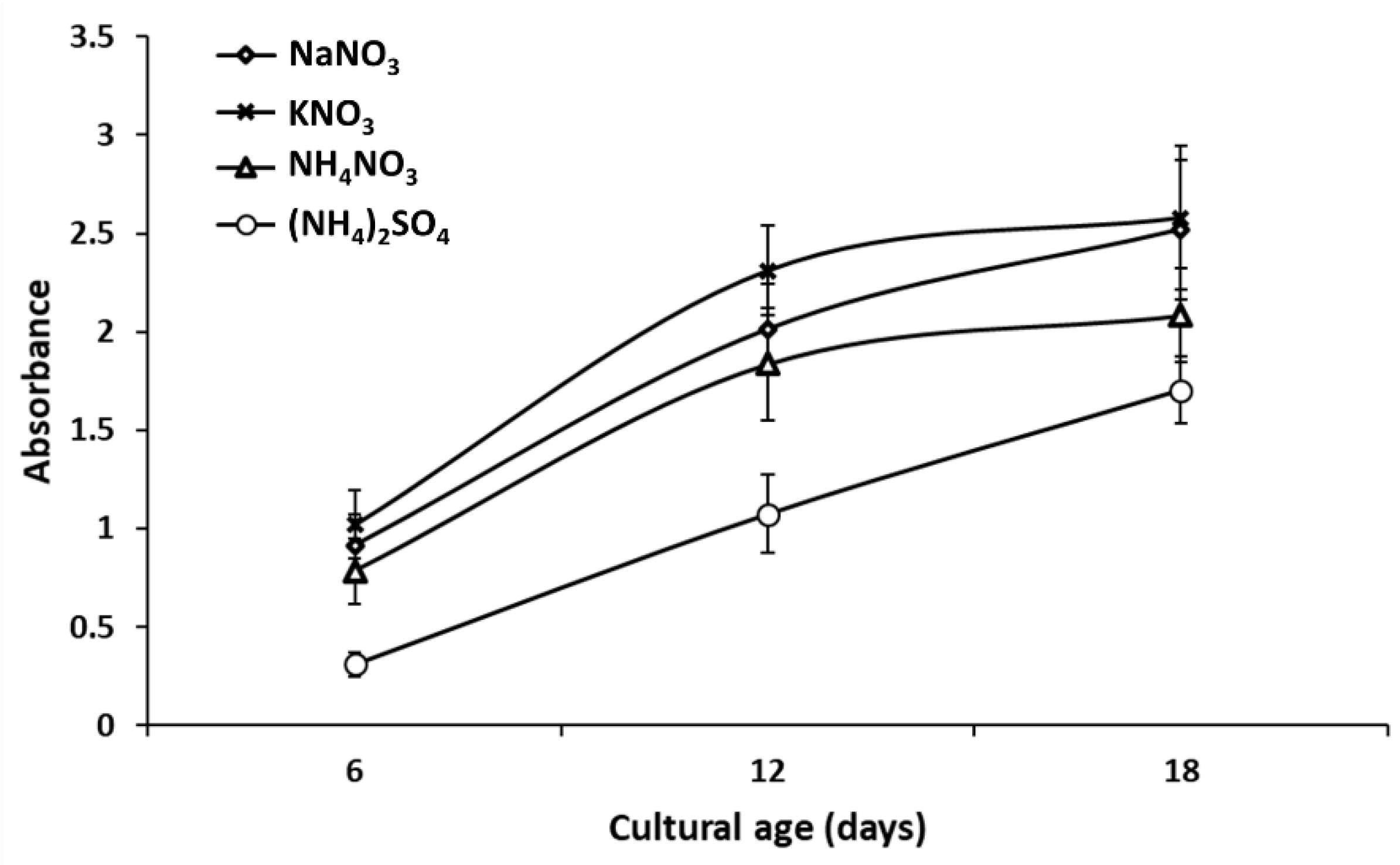
3.1. Visible Observation and UV–Visible Spectroscopy of AgNPs Biosynthesis
3.1.1. Impact of Nitrogen Source
3.1.2. Impact of Microalgal Growth Stage
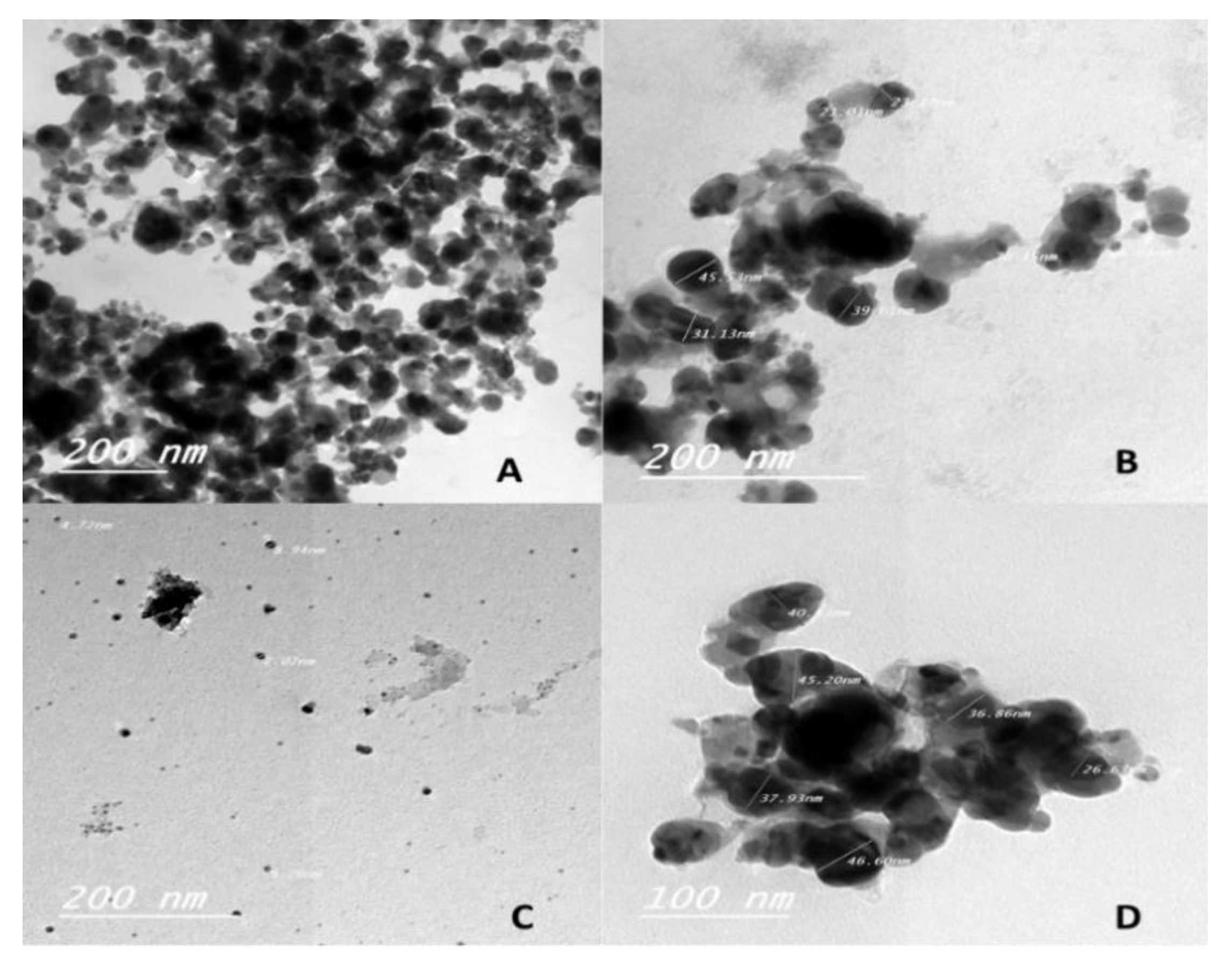
3.2. Characterization of AgNPs by High-Resolution Transmission Electron Microscopy (HRTEM)
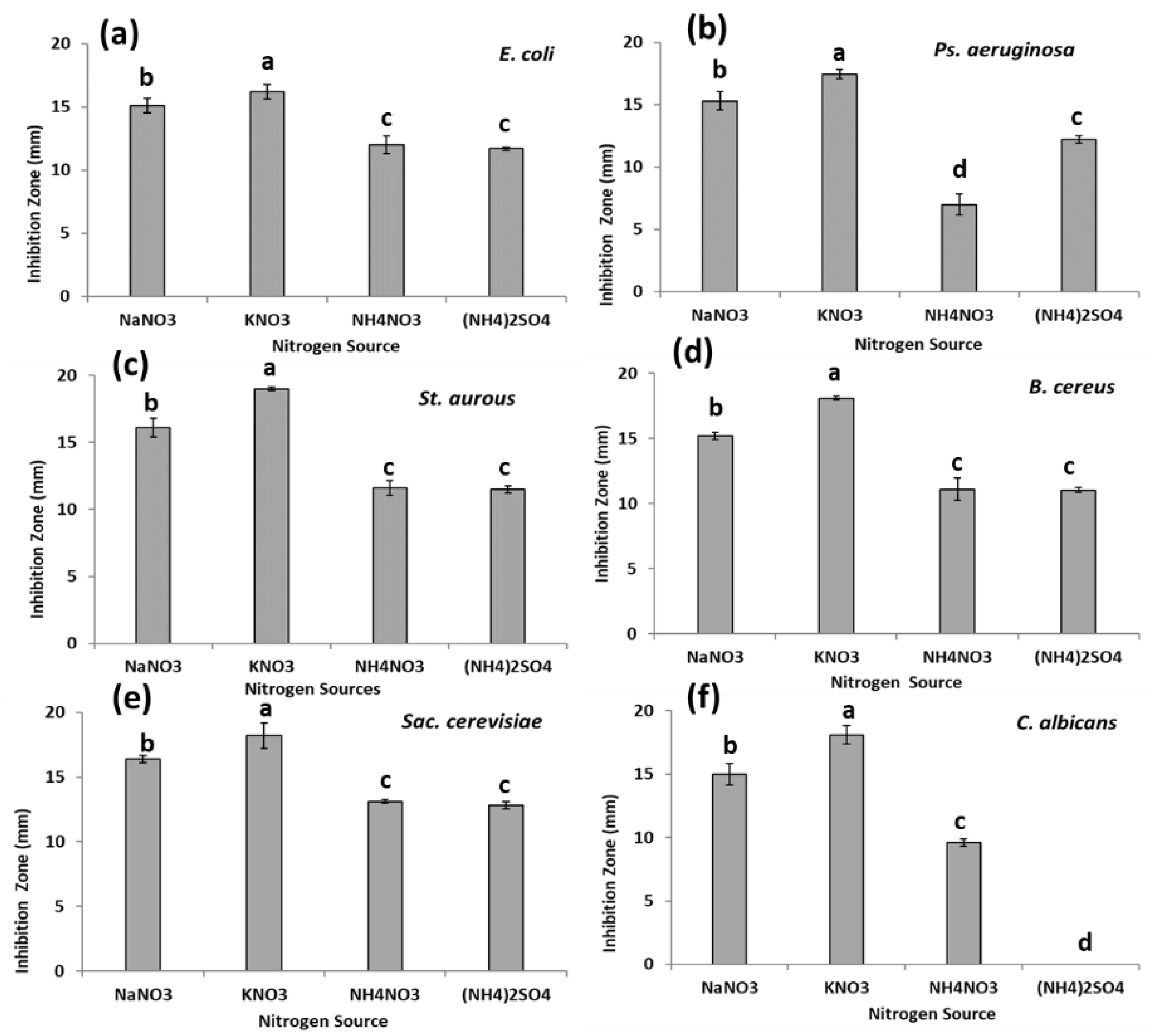
3.3. Antimicrobial Activity of Synthesized AgNPs
4. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Hopkins, M.M.; Kraft, A.; Martin, P.A.; Nightingale, P.; Mahdi, S. Is the biotechnology revolution a myth? In Comprehensive Medicinal Chemistry II; Taylor, J.B., Triggle, D.J., Eds.; Elsevier: London, UK, 2007; Volume 1, pp. 591–613. [Google Scholar]
- Abou-Shanab, R.A.; Matter, I.A.; Kim, S.N.; Oh, Y.K.; Choi, J.; Jeon, B.H. Characterization and identification of lipid-producing microalgae species isolated from a freshwater lake. Biomass Bioenergy 2011, 35, 3079–3085. [Google Scholar] [CrossRef]
- Matter, I.A.; Darwesh, O.M.; Eida, M.F. Harvesting of Scenedesmus obliquus by bioflocculation: Appropriate chitosan concentrations with various pH values at different growth stages. Jordan J. Biol. Sci. 2018, 11, 475–481. [Google Scholar]
- Hasanin, M.S.; Mostafa, A.M.; Mwafy, E.A.; Darwesh, O.M. Eco-friendly cellulose nano fibers via first reported Egyptian Humicola fuscoatra Egyptia X4: Isolation and characterization. Environ. Nanotechnol. Monit. Manag. 2018, 10, 409–418. [Google Scholar] [CrossRef]
- Hasanin, M.S.; Darwesh, O.M.; Matter, I.A.; El-Saied, H. Isolation and characterization of non-cellulolytic Aspergillus flavus EGYPTA5 exhibiting selective ligninolytic potential. Biocatal. Agric. Biotechnol. 2019, 17, 160–167. [Google Scholar] [CrossRef]
- Abou-Shanab, R.A.I.; Khalafallah, M.A.; Emam, N.F.; Aly, M.A.; Abou-Sdera, S.A.; Matter, I.A. Characterisation and identification of carbofuran-utilising bacteria isolated from agricultural soil. Chem. Ecol. 2012, 28, 193–203. [Google Scholar] [CrossRef]
- Saber, M.; Hoballah, E.; Matter, I.; Zaghloul, A. Bioremediation of chlorinated hydrocarbons in a sewaged soil by certain remediative amendments followed by phytoremediation. Res. J. Pharm. Biol. Chem. Sci. 2014, 5, 83–90. [Google Scholar]
- Hoballah, E.; Saber, M.; Matter, I.; Zaghloul, A. Bioremediation of polychlorinated biphenyl (PCBs) in a sewaged soil by certain remediative amendments followed by phytoremediation. Res. J. Pharm. Biol. Chem. Sci. 2014, 5, 91–103. [Google Scholar]
- Barakat, K.M.; Hassan, S.W.M.; Darwesh, O.M. Biosurfactant production by haloalkaliphilic Bacillus strains isolated from Red Sea, Egypt. Egypt. J. Aquat. Res. 2017, 43, 205–211. [Google Scholar] [CrossRef]
- Annamalai, J.; Nallamuthu, T. Green synthesis of silver nanoparticles: Characterization and determination of antibacterial potency. Appl. Nanosci. 2016, 6, 259–265. [Google Scholar] [CrossRef]
- Marrez, D.A.; Abdelhamid, A.E.; Darwesh, O.M. Eco-friendly cellulose acetate green synthesized silver nano-composite as antibacterial packaging system for food safety. Food Packag. Shelf Life 2019, 20, 100302. [Google Scholar] [CrossRef]
- Patel, V.; Berthold, D.; Puranik, P.; Gantar, M. Screening of cyanobacteria and microalgae for their ability to synthesize silver nanoparticles with antibacterial activity. Biotechnol. Rep. 2015, 5, 112–119. [Google Scholar] [CrossRef]
- Darwesh, O.M.; Matter, I.A.; Eida, M.F. Development of peroxidase enzyme immobilized magnetic nanoparticles for bioremediation of textile wastewater dye. J. Environ. Chem. Eng. 2019, 7, 102805. [Google Scholar] [CrossRef]
- Lee, K.; Lee, S.Y.; Praveenkumar, R.; Kim, B.; Seo, J.Y.; Jeon, S.G.; Na, J.G.; Park, J.Y.; Kim, D.M.; Oh, Y.K. Repeated use of stable magnetic flocculant for efficient harvest of oleaginous Chlorella sp. Bioresour. Technol. 2014, 167, 284–290. [Google Scholar] [CrossRef]
- Gu, H.; Ho, P.L.; Tong, E.; Wang, L.; Xu, B. Presenting vancomycin on nanoparticles to enhance antimicrobial activities. Nano Lett. 2003, 3, 1261–1263. [Google Scholar] [CrossRef]
- Schabes-Retchkiman, P.S.; Canizal, G.; Herrera-Becerra, R.; Zorrilla, C.; Liu, H.B.; Ascencio, J.A. Biosynthesis and characterization of Ti/Ni bimetallic nanoparticles. Opt. Mater. 2006, 29, 95–99. [Google Scholar] [CrossRef]
- El-Naggar, N.E.; Abdelwahed, N.A.M.; Darwesh, O.M. Fabrication of biogenic antimicrobial silver nanoparticles by Streptomyces aegyptia NEAE 102 as eco-friendly nanofactory. J. Microbiol. Biotechnol. 2014, 24, 453–464. [Google Scholar] [CrossRef]
- Kim, B.; Praveenkumar, R.; Lee, J.; Nam, B.; Kim, D.M.; Lee, K.; Lee, Y.C.; Oh, Y.K. Magnesium aminoclay enhances lipid production of mixotrophic Chlorella sp. KR-1 while reducing bacterial populations. Bioresour. Technol. 2016, 219, 608–613. [Google Scholar] [CrossRef]
- Jena, J.; Pradhan, N.; Nayak, R.R.; Dash, B.P.; Sukla, L.B.; Panda, P.K.; Mishra, B.K. Microalga Scenedesmus sp.: A potential low-cost green machine for silver nanoparticle synthesis. J. Microbiol. Biotechnol. 2014, 24, 522–533. [Google Scholar] [CrossRef]
- Marambio-Jones, C.; Hoek, E.M. A review of the antibacterial effects of silver nanomaterials and potential implications for human health and the environment. J. Nanopart. Res. 2010, 12, 1531–1551. [Google Scholar] [CrossRef]
- Mahdieh, M.; Zolanvari, A.; Azimee, A.S. Green biosynthesis of silver nanoparticles by Spirulina platensis. Sci. Iran. 2012, 19, 926–929. [Google Scholar] [CrossRef]
- Kabary, H.A.; Attia, M.; Easa, S.M.; Awad, N.M.; Abou-Shanab, R.A.; Eida, M.F. Characterization of magnetotactic bacteria (MTBs) isolated from different habitats in Egypt. Res. J. Pharm. Biol. Chem. Sci. 2016, 7, 244–257. [Google Scholar]
- Kabary, H.; Eida, M.F.; Attia, M.; Awad, N.; Easa, S.M. Magnetotactic characterization and environmental application P. aeruginosa kb1 isolate. Annu. Res. Rev. Biol. 2017, 20, 1–10. [Google Scholar] [CrossRef]
- Elshahawy, I.; Abouelnasr, H.M.; Lashin, S.M.; Darwesh, O.M. First report of Pythium aphanidermatum infecting tomato in Egypt and its control using biogenic silver nanoparticles. J. Plant Prot. Res. 2018, 15, 137–151. [Google Scholar]
- Rahman, A.; Kumar, S.; Bafana, A.; Dahoumane, S.; Jeffryes, C. Biosynthetic conversion of Ag+ to highly sable Ag0 nanoparticles by wild type and cell wall deficient strains of Chlamydomonas reinhardtii. Molecules 2019, 24, 98. [Google Scholar] [CrossRef]
- El-Baz, F.K.; Gad, M.S.; Abdo, S.M.; Abed, K.A.; Matter, I.A. Performance and exhaust emissions of a diesel engine burning algal biodiesel blends. Int. J. Mech. Mech. Eng. 2016, 16, 151–158. [Google Scholar]
- Matter, I.A.; Darwesh, O.M.; Eida, M.F. Harvesting of microalgae Scenedesmus obliquus using chitosan-alginate dual flocculation system. Biosci. Res. 2018, 15, 540–548. [Google Scholar]
- Eida, M.F.; Matter, I.A.; Darwesh, O.M. Cultivation of oleaginous microalgae Scenedesmus obliquus on secondary treated municipal wastewater as growth medium for biodiesel production. Ecol. Eng. 2018, 19, 38–50. [Google Scholar] [CrossRef]
- Perin, G.; Bellan, A.; Bernardi, A.; Bezzo, F.; Morosinotto, T. The potential of quantitative models to improve microalgae photosynthetic efficiency. Physiol. Plant. 2019, in press. [Google Scholar] [CrossRef]
- Amin, N.F.; Khalafallah, M.A.; Ali, M.A.; Abou-Sdera, S.A.; Matter, I.A. Effect of some nitrogen sources on growth and lipid of microalgae Chlorella sp. for biodiesel production. J. Appl. Sci. Res. 2013, 9, 4845–4855. [Google Scholar]
- Sahayaraj, K. Bionanomaterials: Synthesis and applications. In Proceedings of the National Seminar on New Materials Research and Nano Technology; John, N.J., Ed.; Govt. Arts College: Ooty, Tamil Nadu, Indian, 2012; pp. 24–29. [Google Scholar]
- González-Fernández, C.; Ballesteros, M. Microalgae autoflocculation: An alternative to high-energy consuming harvesting methods. J. Appl. Phycol. 2013, 25, 991–999. [Google Scholar] [CrossRef]
- Xiao, R.; Zheng, Y. Overview of microalgal extracellular polymeric substances (EPS) and their applications. Biotechnol. Adv. 2016, 34, 1225–1244. [Google Scholar] [CrossRef]
- Rahman, A.; Kumar, S.; Bafana, A.; Dahoumane, S.A.; Jeffryes, C. individual and combined effects of extracellular polymeric substances and whole cell components of Chlamydomonas reinhardtii on silver nanoparticle synthesis and stability. Molecules 2019, 24, 956. [Google Scholar] [CrossRef]
- El-Sheekh, M.M.; El-Kassas, H.Y. Algal production of nano-silver and gold: Their antimicrobial and cytotoxic activities: A review. J. Genet. Eng. Biotechnol. 2016, 14, 299–310. [Google Scholar] [CrossRef] [Green Version]
- Gong, P.; Li, H.; He, X.; Wang, K.; Hu, J.; Tan, W.; Zhang, S.; Yang, X. Preparation and antibacterial activity of Fe3O4@ Ag nanoparticles. Nanotechnology 2007, 18, 285604. [Google Scholar] [CrossRef]
- Khalil, A.M.; Abdel-Monem, R.A.; Darwesh, O.M.; Hashim, A.I.; Nada, A.A.; Rabie, S.T. Synthesis, characterization, and evaluation of antimicrobial activities of chitosan and carboxymethyl chitosan schiff-base/Silver nanoparticles. J. Chem. 2017, 2017, 1434320. [Google Scholar] [CrossRef]
- Rai, M.; Yadav, A.; Gade, A. Silver nanoparticles as a new generation of antimicrobials. Biotechnol. Adv. 2009, 27, 76–83. [Google Scholar] [CrossRef]
- Pal, S.; Tak, Y.K.; Song, J.M. Does the antibacterial activity of silver nanoparticles depend on the shape of the nanoparticle? A study of the gram-negative bacterium Escherichia coli. Appl. Environ. Microbiol. 2007, 73, 1712–1720. [Google Scholar] [CrossRef]
- Barsanti, L.; Gualtieri, P. Algae: Anatomy, Biochemistry, and Biotechnology, 2nd ed.; CRC Press, Taylor & Francis Group, LLC: Boca Raton, FL, USA, 2014; 361p. [Google Scholar]
- Basova, E.M.; Bulanova, M.A.; Ivanov, V.M. Photometric detection of urea in natural waters. Mosc. Univ. Chem. Bull. 2011, 66, 345–350. [Google Scholar] [CrossRef]
- Redinbaugh, M.G.; Campbell, W.H. Quaternary structure and composition of squash NADH: Nitrate reductase. J. Biol. Chem. 1985, 260, 3380–3385. [Google Scholar]
- Kheiralla, Z.H.; Hewedy, M.A.; Mohammed, H.R.; Darwesh, O.M. Isolation of pigment producing actinomycetes from rhizosphere soil and application it in textiles dyeing. Res. J. Pharm. Biol. Chem. Sci. 2016, 7, 2128–2136. [Google Scholar]
- Darwesh, O.M.; Sultan, Y.Y.; Seif, M.M.; Marrez, D.A. Bio-evaluation of crustacean and fungal nano-chitosan for applying as food ingredient. Toxicol. Rep. 2018, 5, 348–356. [Google Scholar] [CrossRef] [PubMed]
- Sultan, Y.Y.; Ali, M.A.; Darwesh, O.M.; Embaby, M.A.; Marrez, D.A. Influence of nitrogen source in culture media on antimicrobial activity of Microcoleus lacustris and Oscillatoria rubescens. Res. J. Pharm. Biol. Chem. Sci. 2016, 7, 1444–1452. [Google Scholar]
- Kim, G.; Mujtaba, G.; Lee, K. Effects of nitrogen sources on cell growth and biochemical composition of marine chlorophyte Tetraselmis sp. for lipid production. Algae 2016, 31, 257–266. [Google Scholar] [CrossRef]
- Agwa, O.K.; Abu, G.O. Influence of various nitrogen sources on biomass and lipid production by Chlorella vulgaris. Br. Biotechnol. J. 2016, 15, 1–13. [Google Scholar] [CrossRef]
- Ebrahiminezhad, A.; Bagheri, M.; Taghizadeh, S.M.; Berenjian, A.; Ghasemi, Y. Biomimetic synthesis of silver nanoparticles using microalgal secretory carbohydrates as a novel anticancer and antimicrobial. Adv. Nat. Sci. Nanosci. Nanotechnol. 2016, 7, 015018. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, K.; Gupta, R.; Kumar, G.; Kumari, S.; Biswas, S.; Padmanabhan, P. Synthesis of silver nanoparticles by Bacillus clausii and computational profiling of nitrate reductase enzyme involved in production. J. Genet. Eng. Biotechnol. 2018, 16, 527–536. [Google Scholar] [CrossRef]
- Matter, I.A.; Darwesh, O.M.; El-baz, F.K. Using the natural polymer chitosan in harvesting Scenedesmus species under different concentrations and cultural pH values. Int. J. Pharm. Biol. Sci. 2016, 7, 254–260. [Google Scholar]
- Sarayloo, E.; Tardu, M.; Unlu, Y.S.; Simsek, S.; Cevahir, G.; Erkey, C.; Kavakli, I.H. Understanding lipid metabolism in high-lipid-producing Chlorella vulgaris mutants at the genome-wide level. Algal Res. 2017, 28, 244–252. [Google Scholar] [CrossRef]
- Herrero, A.; Muro-Pastor, A.M.; Flores, E. Nitrogen control in cyanobacteria. J. Bacteriol. 2001, 183, 411–425. [Google Scholar] [CrossRef]
- Muro-Pastor, M.I.; Reyes, J.C.; Florencio, F.J. Ammonium assimilation in cyanobacteria. Photosynth. Res. 2005, 83, 135–150. [Google Scholar] [CrossRef]
- Jha, P.; Ali, A.; Raghuram, N. Nitrate-induction of nitrate reductase and its inhibition by nitrite and ammonium ions in Spirulina platensis. Physiol. Mol. Biol. Plants 2007, 13, 163–167. [Google Scholar]
- Khodashenas, B.; Ghorbani, H.R. Optimisation of nitrate reductase enzyme activity to synthesise silver nanoparticles. IET Nanobiotechnol. 2016, 10, 158–161. [Google Scholar] [CrossRef]
- Ramezani, F.; Ramezani, M.; Talebi, S. Mechanistic aspects of biosynthesis of nanoparticles by several microbes. Nanocon 2010, 10, 1–7. [Google Scholar]
- Moteshafi, H.; Mousavi, S.M.; Shojaosadati, S.A. The possible mechanisms involved in nanoparticles biosynthesis. J. Ind. Eng. Chem. 2012, 18, 2046–2050. [Google Scholar] [CrossRef]
- Pradhan, N.; Nayak, R.R.; Pradhan, A.K.; Sukla, L.B.; Mishra, B.K. In situ Synthesis of entrapped silver nanoparticles by a fungus—Penicillium purpurogenum. Nanosci. Nanotechnol. Lett. 2011, 3, 659–665. [Google Scholar] [CrossRef]
- Vahabi, K.; Mansoori, G.A.; Karimi, S. Biosynthesis of silver nanoparticles by fungus Trichoderma reesei (a route for large-scale production of AgNPs). Insci. J. 2011, 1, 65–79. [Google Scholar] [CrossRef]
- Cuellar-Bermudez, S.P.; Aguilar-Hernandez, I.; Cardenas-Chavez, D.L.; Ornelas-Soto, N.; Romero-Ogawa, M.A.; Parra-Saldivar, R. Extraction and purification of high-value metabolites from microalgae: Essential lipids, astaxanthin and phycobiliproteins. Microb. Biotechnol. 2015, 8, 190–209. [Google Scholar] [CrossRef]
- Durán, N.; Marcato, P.D.; Alves, O.L.; De Souza, G.I.; Esposito, E. Mechanistic aspects of biosynthesis of silver nanoparticles by several Fusarium oxysporum strains. J. Nanobiotechnol. 2005, 3, 1–7. [Google Scholar] [CrossRef]
- Younes, A.M.; Mohamed, L.A.; Eida, M.F.; Gaafar, A.Y. Characterization and pathogen challenge of Pseudomonas species from Oreochromis niloticus in Egypt. Res. J. Pharm. Biol. Chem. Sci. 2015, 6, 312–317. [Google Scholar]
- Fernández, M.; Hudson, J.A.; Korpela, R.; de los Reyes-Gavilán, C.G. Impact on human health of microorganisms present in fermented dairy products: An overview. BioMed Res. Int. 2015, 2015, 412714. [Google Scholar] [CrossRef]
- Morones, J.R.; Elechiguerra, J.L.; Camacho, A.; Holt, K.; Kouri, J.B.; Ramírez, J.T.; Yacaman, M.J. The bactericidal effect of silver nanoparticles. Nanotechnology 2005, 16, 2346. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nitrogen Source | OD (Abs 680 nm) | Dry Biomass (g/L) |
|---|---|---|
| NaNO3 | 2.33 ± 0.2 b | 0.63 ± 0.013 c |
| CO(NH4)2 (urea) | 3.20 ± 0.3 a | 1.09 ± 0.046 a |
| (NH4)2CO3 | 2.66 ± 0.2 b | 0.76 ± 0.046 b |
| KNO3 | 1.92 ± 0.2 c | 0.48 ± 0.008 e |
| NH4NO3 | 2.72 ± 0.3 b | 0.56 ± 0.004 d |
| (NH4)2SO4 | 0.84 ± 0.1 d | 0.26 ± 0.001 f |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Darwesh, O.M.; Matter, I.A.; Eida, M.F.; Moawad, H.; Oh, Y.-K. Influence of Nitrogen Source and Growth Phase on Extracellular Biosynthesis of Silver Nanoparticles Using Cultural Filtrates of Scenedesmus obliquus. Appl. Sci. 2019, 9, 1465. https://0-doi-org.brum.beds.ac.uk/10.3390/app9071465
Darwesh OM, Matter IA, Eida MF, Moawad H, Oh Y-K. Influence of Nitrogen Source and Growth Phase on Extracellular Biosynthesis of Silver Nanoparticles Using Cultural Filtrates of Scenedesmus obliquus. Applied Sciences. 2019; 9(7):1465. https://0-doi-org.brum.beds.ac.uk/10.3390/app9071465
Chicago/Turabian StyleDarwesh, Osama M., Ibrahim A. Matter, Mohamed F. Eida, Hassan Moawad, and You-Kwan Oh. 2019. "Influence of Nitrogen Source and Growth Phase on Extracellular Biosynthesis of Silver Nanoparticles Using Cultural Filtrates of Scenedesmus obliquus" Applied Sciences 9, no. 7: 1465. https://0-doi-org.brum.beds.ac.uk/10.3390/app9071465