A Pilot Study on Linking Tissue Mechanics with Load-Dependent Collagen Microstructures in Porcine Tricuspid Valve Leaflets

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
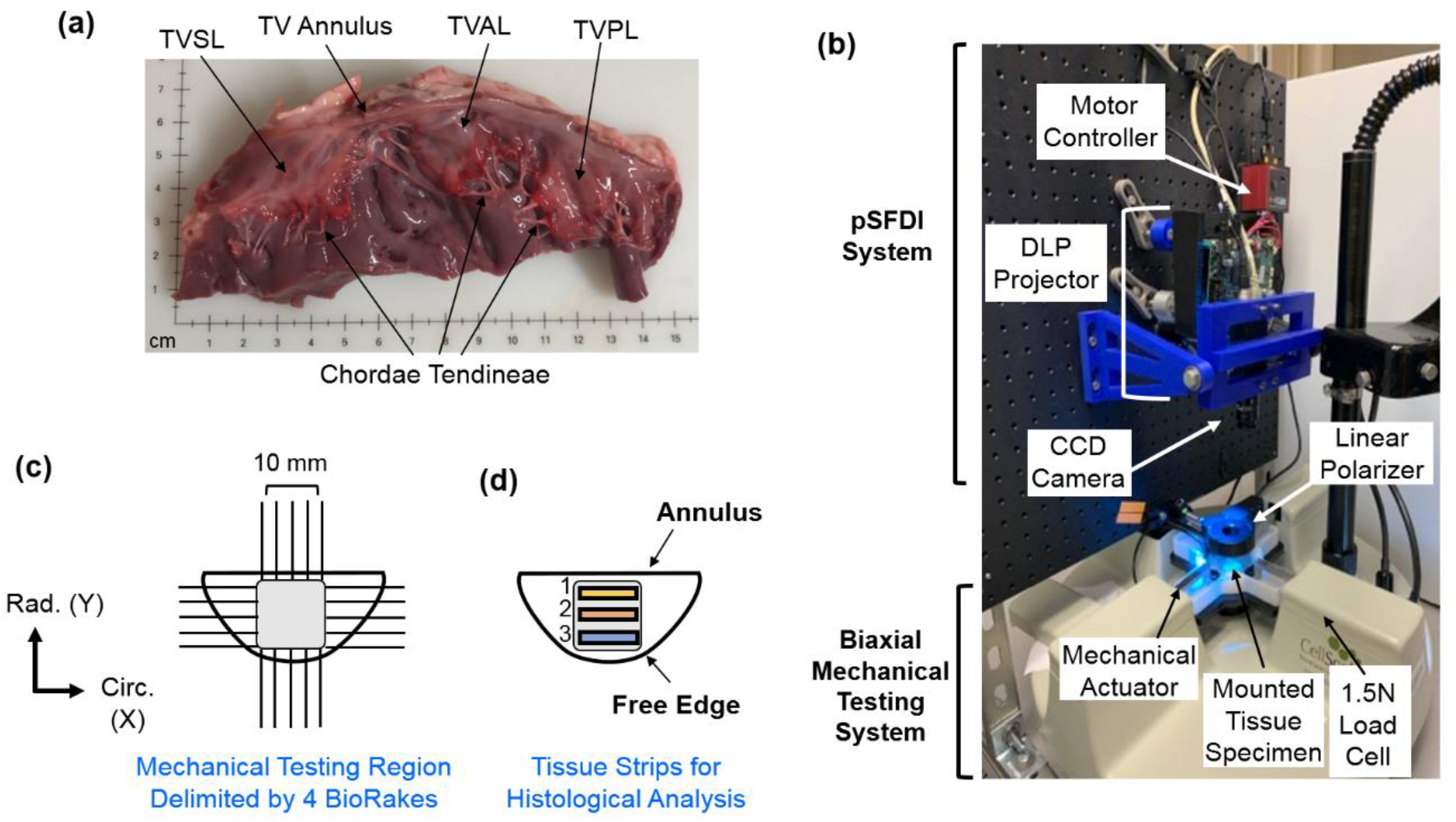
2.1. Porcine Heart Acquisition and Tissue Preparation
2.2. Biaxial Mechanical Testing
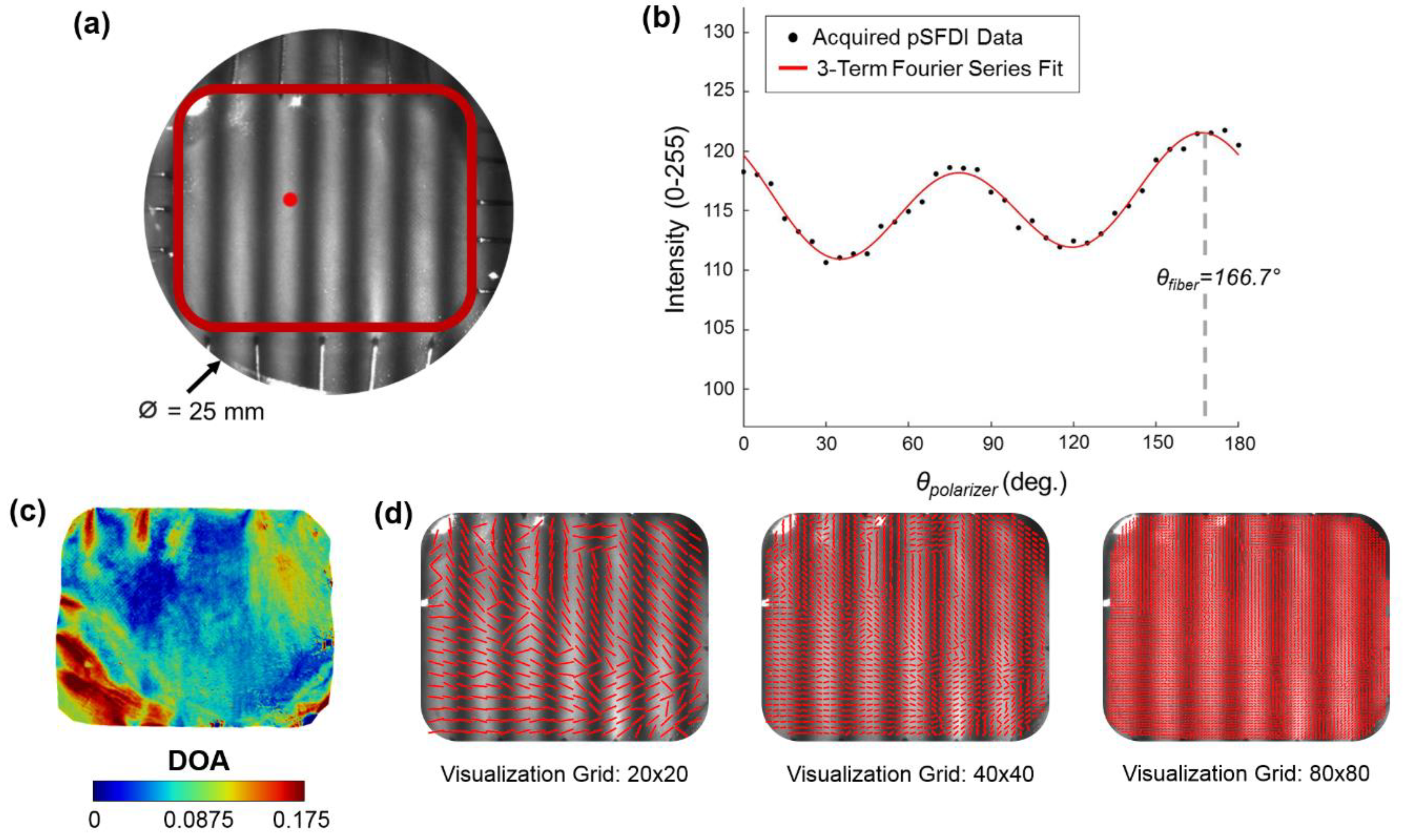
2.3. pSFDI-Based Collagen Microstructure Quantifications
2.4. Histological Analysis
2.5. Statistical Analysis
3. Results
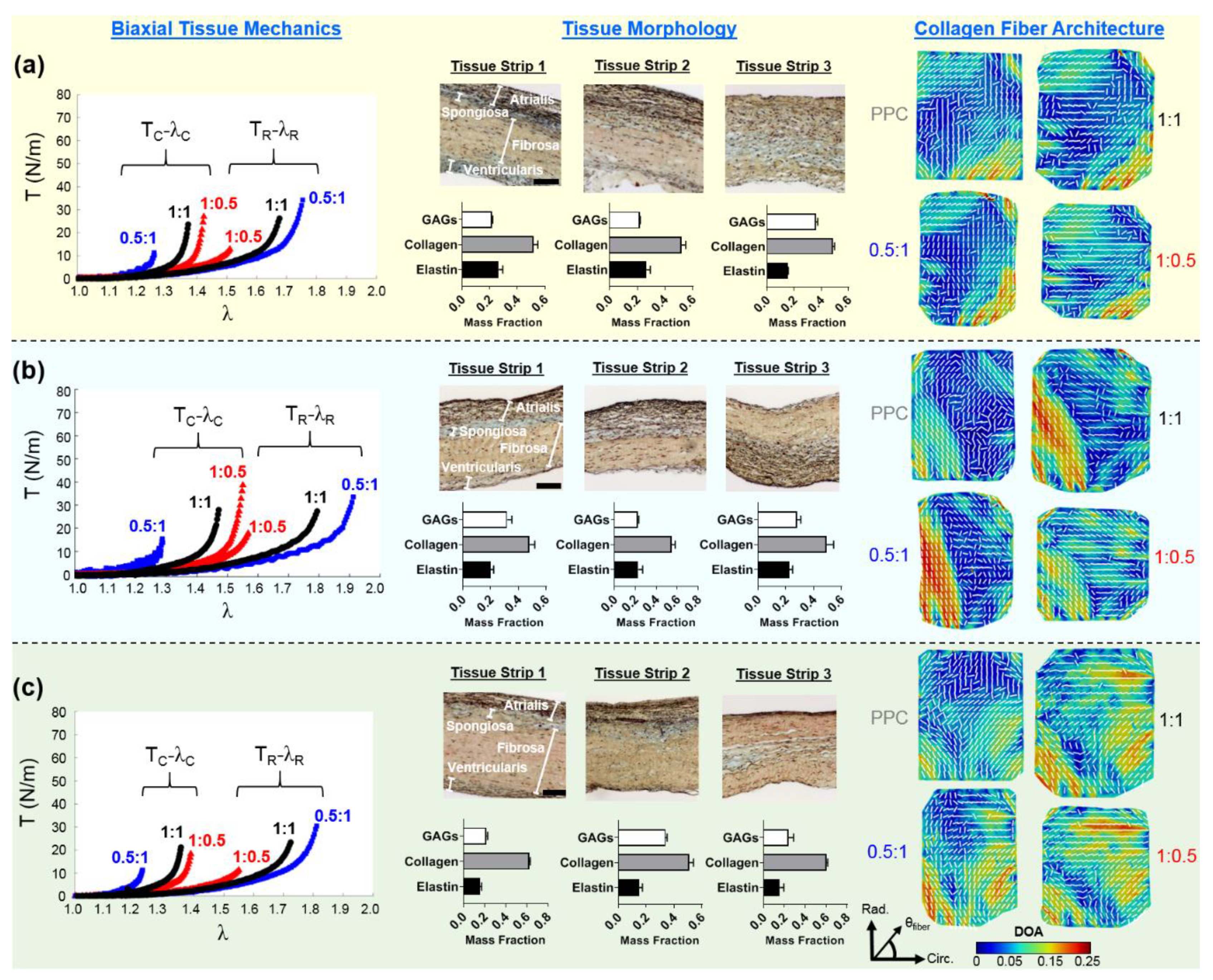
3.1. Biaxial Mechanical Testing Results
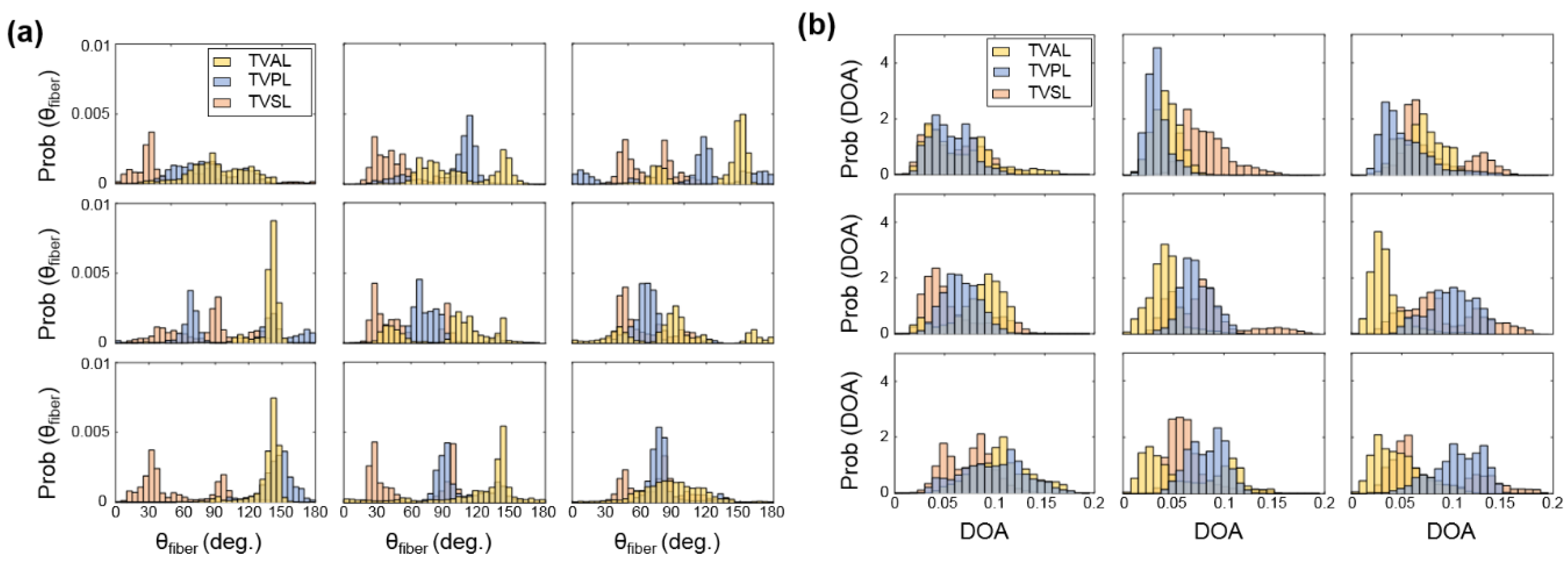
3.2. Histological Results
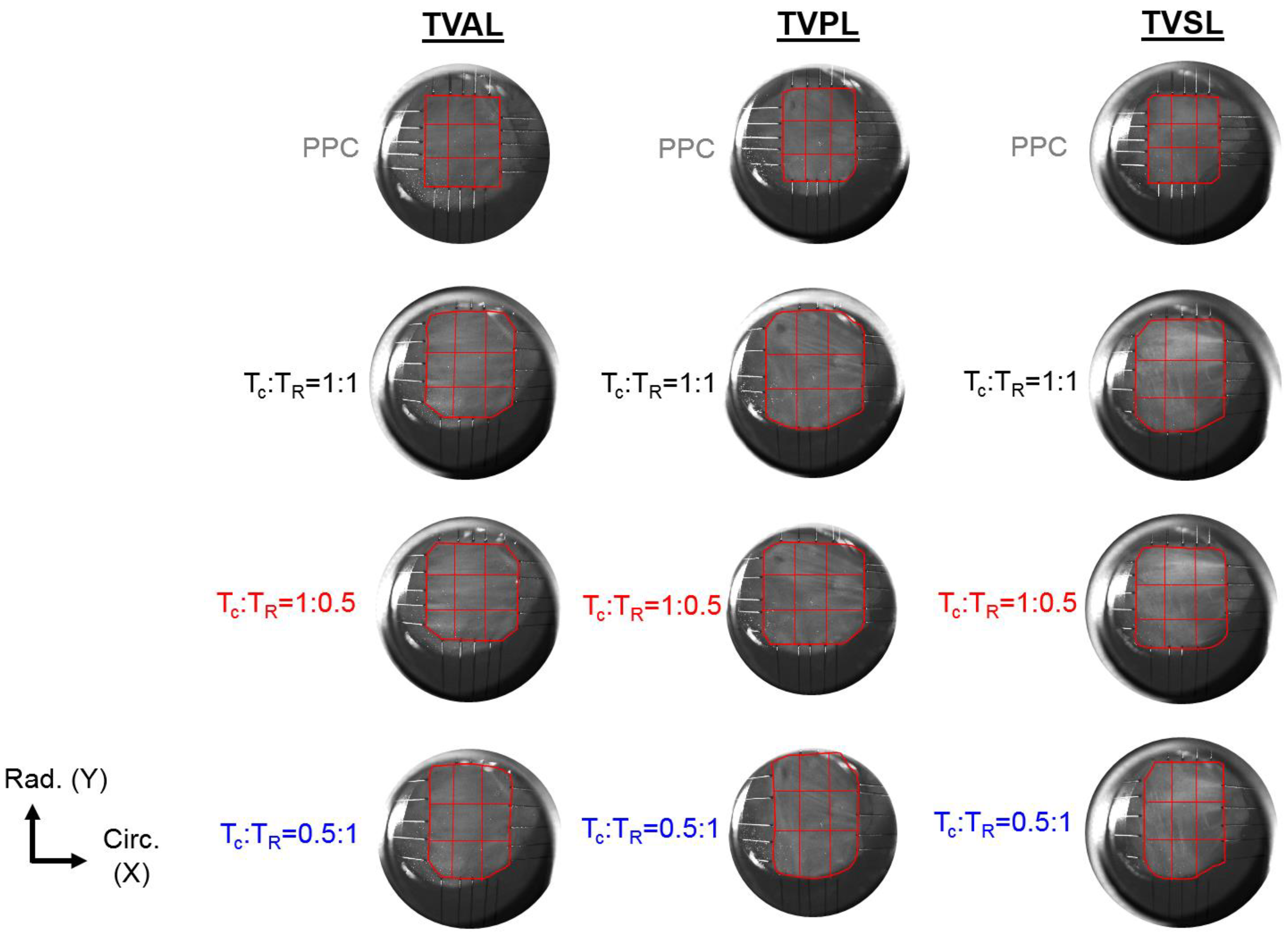
3.3. Load-Dependent Collagen Fiber Architecure
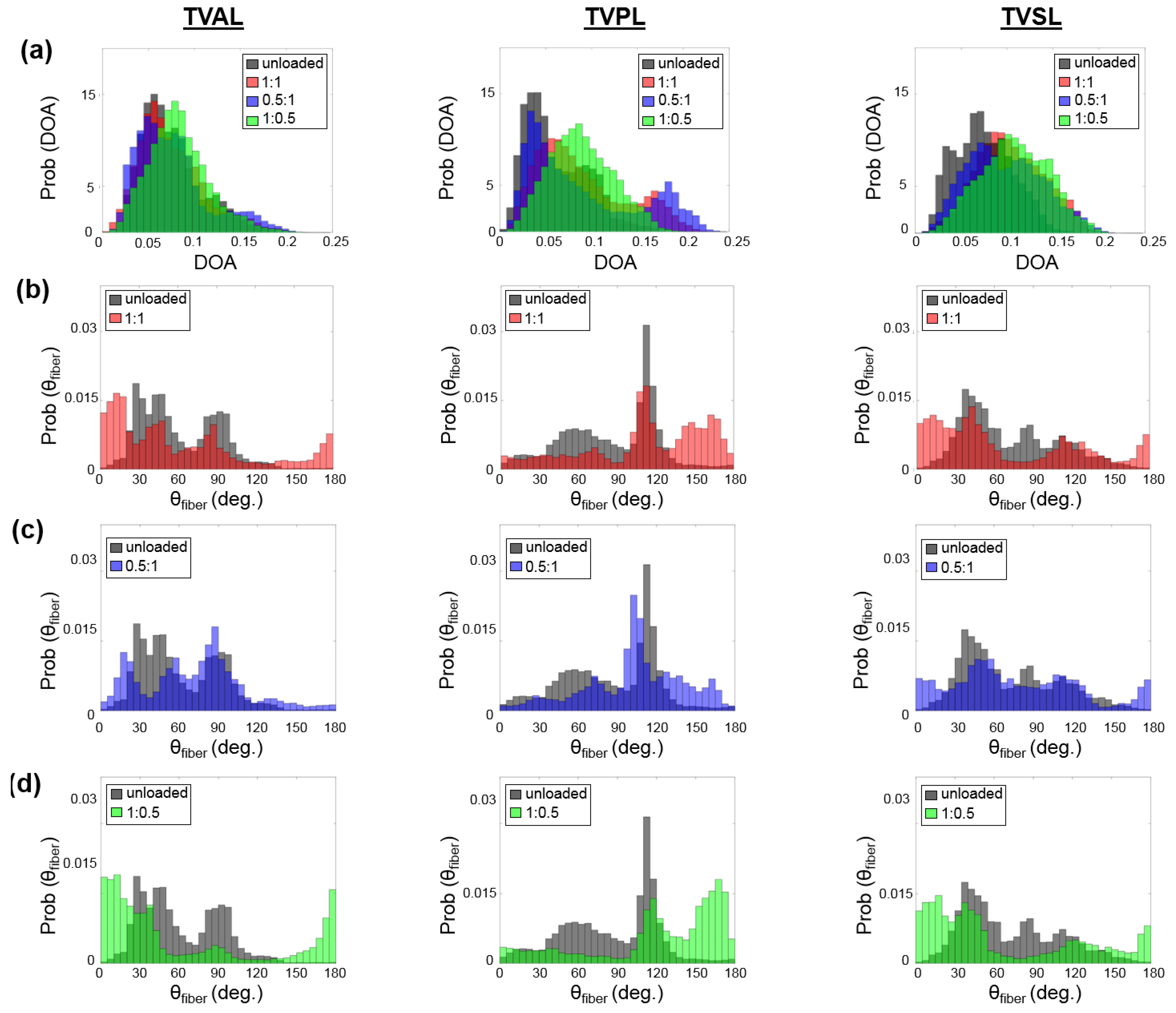
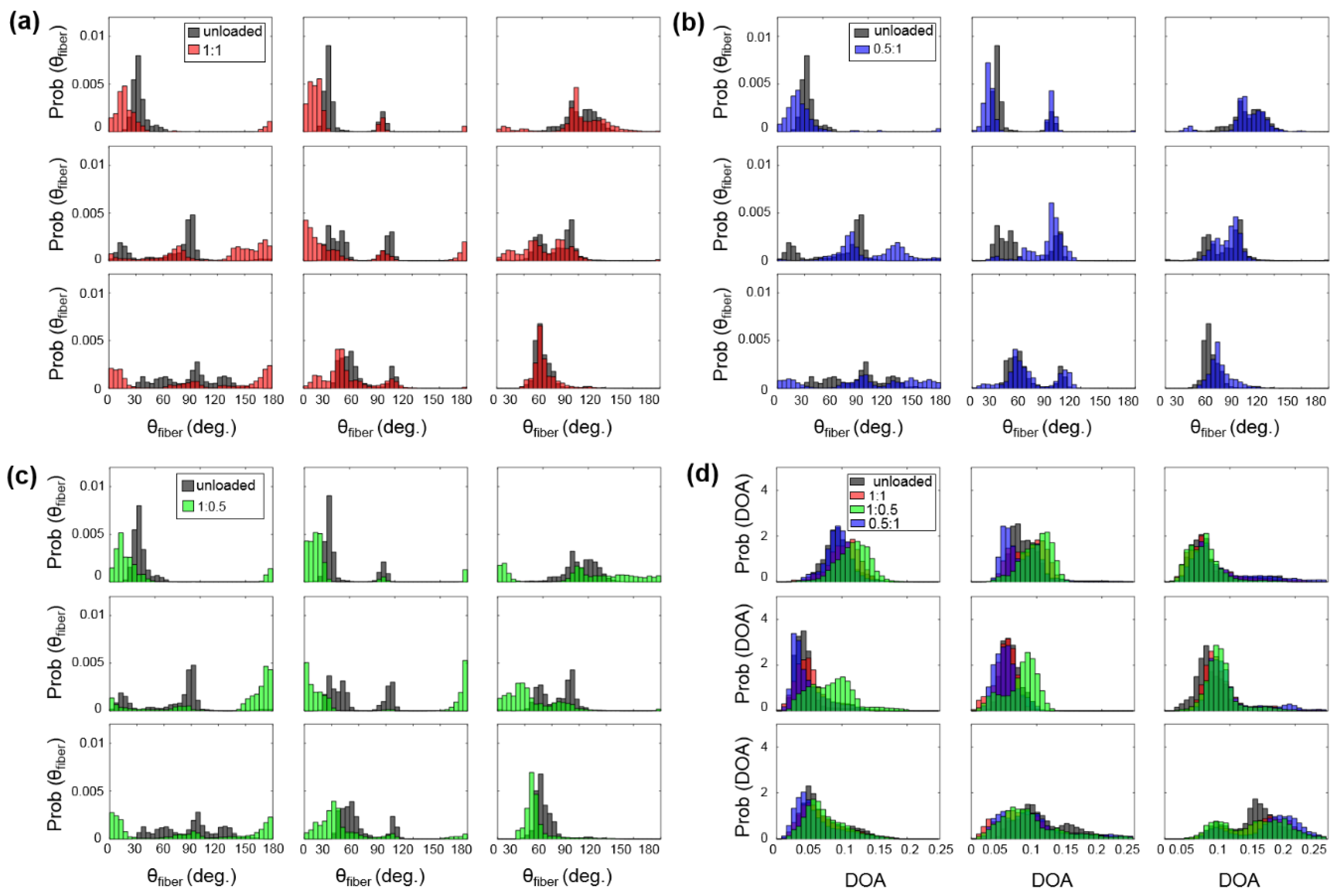
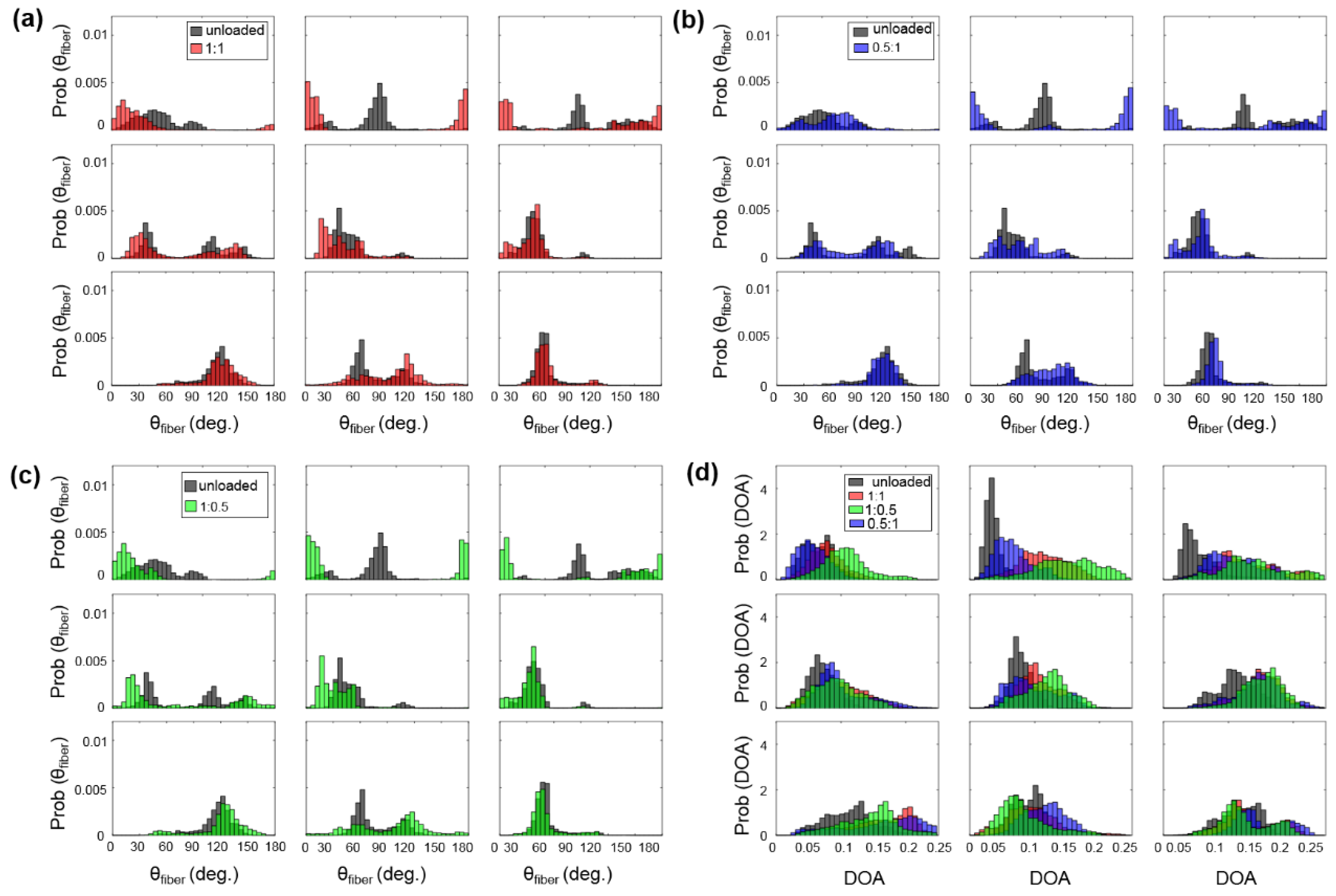
- Firstly, for the non-equibiaxial loading protocols, the collagen fiber orientations displayed a shift towards the direction of the maximum applied loading (Figure 4). Moreover, collagen fiber networks became more aligned, as evidenced by the increasing DOA within the central regions of the tissues (Figure A2, Figure A3 and Figure A4).
- Secondly, the largest changes in the mean collagen fiber orientation were observed in the TVPL under equibiaxial and radially-dominant loading conditions (Table 5), with differences of 37.5% ( 1:1), 36.0% ( 2:2), and 42.2% ( 0.5:1), compared to the post-preconditioning (PPC) state, also referred to as the “unloaded” state in the remaining discussion.
- Thirdly, the largest percent change in the quantified DOA, as compared to the PPC state, occurred under the radially-dominant loading ( 0.5:1), with a 10.8% increase for the TVAL, a 39.1% increase for the TVPL, and a 47.4% increase for the TVSL, respectively (Table 5). Under circumferentially-dominant loading ( 1:0.5), a minimal change of 1.4% in DOA was seen for the TVAL, with a 27.3% and a 32.4% increase for the TVPL and TVSL, respectively (Table 5).
- Fourthly, across all the loading protocols, the TVSL displayed the largest increase in the DOA (47.3%) under the radially-dominant loading, whereas the smallest change was found for the TVAL (1.4%), under equibiaxial and circumferentially-dominant loading (Table 5).
3.3.1. Changes in the CFA Associated with the Equibiaxial Loading States
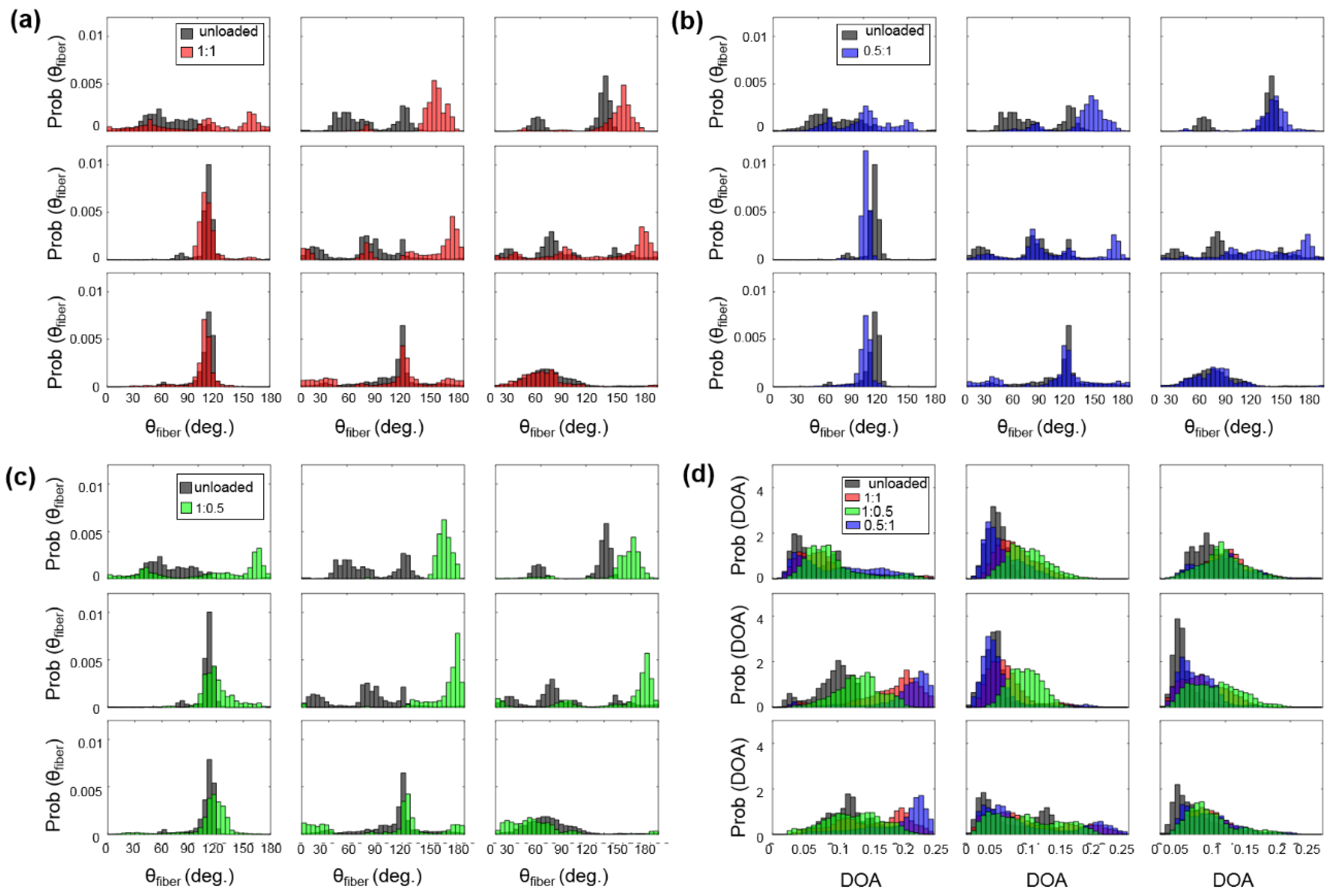
3.3.2. Changes in the CFA Associated with the Circumferentially-Dominant Loading ( 1:0.5)
3.3.3. Changes in the CFA Associated with the Radially-Dominant Loading ( = 0.5:1)
4. Discussion
4.1. Mechanics-Related Observations
4.2. Collagen Fiber Architecture-Related Observations
4.3. Study Limitations and Future Work
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Nomenclature
| Category | Abbreviation | Definition |
| Anatomy | TV | Tricuspid valve |
| TVAL | Tricuspid valve anterior leaflet | |
| TVPL | Tricuspid valve posterior leaflet | |
| TVSL | Tricuspid valve septal leaflet | |
| RV | Right ventricle | |
| FTR | Functional tricuspid regurgitation | |
| ECM | Extracellular matrix | |
| Tissue Layers | A | Atrialis |
| S | Spongiosa | |
| F | Fibrosa | |
| V | Ventricularis | |
| Tissue Layer Constituents | CFA | Collagen fiber architecture |
| GAGs | Glycosaminoglycans | |
| PGs | Proteoglycans | |
| Instrumentation | pSFDI | Polarized spatial frequency domain imaging |
| SALS | Small angle light scattering | |
| SHG | Second harmonic generation | |
| FOV | Field of view | |
| ROI | Region of interest | |
| Collagen Fiber Quantification | Collagen fiber orientation | |
| DOA | Degree of optical anisotropy | |
| Mechanics | C | Tissue’s circumferential direction |
| R | Tissue’s radial direction | |
| T | Membrane tension | |
| PPC | Post-preconditioning | |
| λ | Tissue stretch |
Appendix A. Quantification of Collagen Fiber Orientation and Degree of Optical Anisotropy

Appendix B. Spatial Heterogeneity of CFAs



Appendix C. Quantified Load-Dependent Changes in the CFA for Heart #2 and Heart #3

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DOA (TVAL) | DOA (TVPL) | DOA (TVSL) | |||||||
| Mean | Skewness | SD | Mean | Skewness | SD | Mean | Skewness | SD | |
| PPC | 0.069 | 0.791 | 0.025 | 0.071 | 1.377 | 0.029 | 0.062 | 0.410 | 0.026 |
| 1:1 | −1.6° | 12.4° | 9.5° | −5.3° | −10.0° | 27.0° | 14.3° | −23.1° | 12.2° |
| 1:0.5 | 12.2° | −5.2° | 41.1° | 6.1° | −8.1° | 39.9° | 43.5° | −47.6° | 13.5° |
| 0.5:1 | −3.3° | −18.0° | −0.6° | −7.4° | −12.8° | 6.7° | 15.0° | −16.4° | 19.2° |
| (TVAL) | (TVPL) | (TVSL) | |||||||
| Mean | Skewness | SD | Mean | Skewness | SD | Mean | Skewness | SD | |
| PPC | 93.4° | −0.365 | 30.0° | 105.3° | −0.891 | 26.3° | 106.0° | −0.462 | 31.9° |
| 1:1 | −1.4° | −18.6° | 15.2° | 6.5° | −19.4° | 47.1° | 0.3° | 27.9° | 54.1° |
| 1:0.5 | 5.5° | 12.5° | −2.1° | 14.3° | −16.5° | 2.6° | 11.5° | 54.0° | 3.4° |
| 0.5:1 | −9.9° | −130.9° | 25.6° | −3.5° | −46.5° | 78.4° | −13.8° | −94.0° | 68.9° |
| DOA (TVAL) | DOA (TVPL) | DOA (TVSL) | |||||||
| Mean | Skewness | SD | Mean | Skewness | SD | Mean | Skewness | SD | |
| PPC | 0.072 | 0.861 | 0.026 | 0.065 | 2.774 | 0.032 | 0.082 | 0.515 | 0.028 |
| 1:1 | 0.3° | −5.8° | 20.4° | 25.8° | −82.9° | 6.6° | 4.2° | 5.4° | 12.0° |
| 1:0.5 | 12.9° | −35.8° | 22.3° | 39.5° | −95.8° | 12.2° | 12.0° | 40.2° | 15.9° |
| 0.5:1 | −7.1° | 13.4° | 16.2° | 12.8° | −86.9° | −1.5° | 1.2° | −38.5° | 14.9° |
| (TVAL) | (TVPL) | (TVSL) | |||||||
| Mean | Skewness | SD | Mean | Skewness | SD | Mean | Skewness | SD | |
| PPC | 113.1° | −0.256 | 22.7° | 110.6 | −0.948 | 34.3° | 85.0° | 0.639 | 29.4° |
| 1:1 | −2.1° | 12.9° | 35.5° | −5.9° | −35.4° | −5.4° | −5.9° | 21.0° | 6.2° |
| 1:0.5 | −1.1° | −27.9° | 12.1° | −3.0° | 14.7° | −22.5° | 6.9° | −34.2° | −11.7° |
| 0.5:1 | −6.7° | 59.8° | 60.1° | −13.5° | −70.3° | 32.6° | −17.7° | 98.9° | 18.5° |
References
- Arsalan, M.; Walther, T.; Smith, R.L., II; Grayburn, P.A. Tricuspid regurgitation diagnosis and treatment. Eur. Heart J. 2015, 38, 634–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anyanwu, A.C.; Adams, D.H. Functional tricuspid regurgitation in mitral valve disease: Epidemiology and prognostic implications. Semin. Thorac. Cardiovasc. Surg. 2010, 22, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Di Mauro, M.; Bezante, G.P.; Di Baldassarre, A.; Clemente, D.; Cardinali, A.; Acitelli, A.; Salerni, S.; Penco, M.; Calafiore, A.M.; Gallina, S. Functional tricuspid regurgitation: An underestimated issue. Int. J. Cardiol. 2013, 168, 707–715. [Google Scholar] [CrossRef] [PubMed]
- Braunwald, N.S.; Ross, J., Jr.; Morrow, A.G. Conservative management of tricuspid regurgitation in patients undergoing mitral valve replacement. Circulation 1967, 35, I-63–I-69. [Google Scholar] [CrossRef]
- Dreyfus, G.D.; Corbi, P.J.; Chan, K.M.J.; Bahrami, T. Secondary tricuspid regurgitation or dilatation: Which should be the criteria for surgical repair? Ann. Thorac. Surg. 2005, 79, 127–132. [Google Scholar] [CrossRef]
- Cyron, C.J.; Humphrey, J.D. Growth and remodeling of load-bearing biological soft tissues. Meccanica 2017, 52, 645–664. [Google Scholar] [CrossRef]
- Driessen, N.J.B.; Cox, M.A.J.; Bouten, C.V.C.; Baaijens, F.P.T. Remodelling of the angular collagen fiber distribution in cardiovascular tissues. Biomech. Model. Mechanobiol. 2008, 7, 93–103. [Google Scholar] [CrossRef] [Green Version]
- Song, J.-M.; Jang, M.-K.; Kim, Y.-J.; Kim, D.-H.; Kang, D.-H.; Song, J.-K. Right ventricular remodeling determines tricuspid valve geometry and the severity of functional tricuspid regurgitation: A real-time 3-dimensional echocardiography study. Korean Circ. J. 2010, 40, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Kramer, K.E.; Ross, C.J.; Laurence, D.W.; Babu, A.R.; Wu, Y.; Towner, R.A.; Mir, A.; Burkhart, H.M.; Holzapfel, G.A.; Lee, C.-H. An investigation of layer-specific tissue biomechanics of porcine atrioventricular heart valve leaflets. Acta Biomater. 2019, 96, 368–384. [Google Scholar] [CrossRef]
- Jett, S.V.; Laurence, D.W.; Kunkel, R.P.; Babu, A.R.; Kramer, K.E.; Baumwart, R.; Towner, R.A.; Wu, Y.; Lee, C.-H. An investigation of the anisotropic mechanical properties and anatomical structure of porcine atrioventricular heart valves. J. Mech. Behav. Biomed. Mater. 2018, 87, 155–171. [Google Scholar] [CrossRef]
- Tseng, H.; Grande-Allen, K.J. Elastic fibers in the aortic valve spongiosa: A fresh perspective on its structure and role in overall tissue function. Acta Biomater. 2011, 7, 2101–2108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinton, R.B.; Yutzey, K.E. Heart valve structure and function in development and disease. Annu. Rev. Physiol. 2011, 73, 29–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stella, J.A.; Sacks, M.S. On the biaxial mechanical properties of the layers of the aortic valve leaflet. J. Biomech. Eng. 2007, 129, 757–766. [Google Scholar] [CrossRef] [PubMed]
- Khoiy, K.A.; Amini, R. On the biaxial mechanical response of porcine tricuspid valve leaflets. J. Biomech. Eng. 2016, 138, 104504. [Google Scholar] [CrossRef]
- Mathur, M.; Jazwiec, T.; Meador, W.D.; Malinowski, M.; Goehler, M.; Ferguson, H.; Timek, T.A.; Rausch, M.K. Tricuspid valve leaflet strains in the beating ovine heart. Biomech. Model. Mechanobiol. 2019, 18, 1351–1361. [Google Scholar] [CrossRef]
- Meador, W.D.; Mathur, M.; Rausch, M.K. Tricuspid valve biomechanics: A brief review. In Advances in Heart Valve Biomechanics; Springer: Berlin/Heidelberg, Germany, 2018; pp. 105–114. [Google Scholar]
- Pham, T.; Sulejmani, F.; Shin, E.; Wang, D.; Sun, W. Quantification and comparison of the mechanical properties of four human cardiac valves. Acta Biomater. 2017, 54, 345–355. [Google Scholar] [CrossRef]
- Lim, K.O. Mechanical properties and ultrastructure of normal human tricuspid valve chordae tendineae. Jpn. J. Physiol. 1980, 30, 455–464. [Google Scholar] [CrossRef] [Green Version]
- Pokutta-Paskaleva, A.; Sulejmani, F.; DelRocini, M.; Sun, W. Comparative mechanical, morphological, and microstructural characterization of porcine mitral and tricuspid leaflets and chordae tendineae. Acta Biomater. 2019, 85, 241–252. [Google Scholar] [CrossRef]
- Ross, C.J.; Laurence, D.W.; Hsu, M.-C.; Baumwart, R.; Zhao, Y.D.; Mir, A.; Burkhart, H.M.; Holzapfel, G.A.; Wu, Y.; Lee, C.-H. Mechanics of Porcine Heart Valves’ Strut Chordae Tendineae Investigated as a Leaflet–Chordae–Papillary Muscle Entity. Ann. Biomed. Eng. 2020. [Google Scholar] [CrossRef]
- Basu, A.; He, Z. Annulus tension on the tricuspid valve: An in-vitro study. Cardiovasc. Eng. Technol. 2016, 7, 270–279. [Google Scholar] [CrossRef]
- Makaryus, A.N.; Ismail, H.; Makaryus, J.N.; Fan, D. Geometric comparison of the mitral and tricuspid valve annulus: Insights from three dimensional transesophageal echocardiography. World J. Cardiol. 2017, 9, 757. [Google Scholar] [CrossRef] [PubMed]
- Malinowski, M.; Jazwiec, T.; Goehler, M.; Quay, N.; Bush, J.; Jovinge, S.; Rausch, M.K.; Timek, T.A. Sonomicrometry-derived 3-dimensional geometry of the human tricuspid annulus. J. Thorac. Cardiovasc. Surg. 2018, 157, 1452–1461. [Google Scholar] [CrossRef] [PubMed]
- Rausch, M.K.; Malinowski, M.; Wilton, P.; Khaghani, A.; Timek, T.A. Engineering analysis of tricuspid annular dynamics in the beating ovine heart. Ann. Biomed. Eng. 2018, 46, 443–451. [Google Scholar] [CrossRef]
- Rausch, M.K.; Malinowski, M.; Meador, W.D.; Wilton, P.; Khaghani, A.; Timek, T.A. The effect of acute pulmonary hypertension on tricuspid annular height, strain, and curvature in sheep. Cardiovasc. Eng. Technol. 2018, 9, 365–376. [Google Scholar] [CrossRef]
- Ring, L.; Rana, B.S.; Kydd, A.; Boyd, J.; Parker, K.; Rusk, R.A. Dynamics of the tricuspid valve annulus in normal and dilated right hearts: A three-dimensional transoesophageal echocardiography study. Eur. Heart J. Cardiovasc. Imaging 2012, 13, 756–762. [Google Scholar] [CrossRef] [PubMed]
- Dreyfus, G.D.; Martin, R.P.; Chan, K.J.; Dulguerov, F.; Alexandrescu, C. Functional tricuspid regurgitation: A need to revise our understanding. J. Am. Coll. Cardiol. 2015, 65, 2331–2336. [Google Scholar] [CrossRef] [Green Version]
- Hahn, R.T. Imaging of the Tricuspid Valve: Transoesophageal Echocardiography. In Practical Manual of Tricuspid Valve Diseases; Springer: Berlin/Heidelberg, Germany, 2018; pp. 117–126. [Google Scholar]
- Kim, J.B.; Jung, S.-H.; Choo, S.J.; Chung, C.H.; Lee, J.W. Clinical and echocardiographic outcomes after surgery for severe isolated tricuspid regurgitation. J. Thorac. Cardiovasc. Surg. 2013, 146, 278–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishi, H.; Toda, K.; Miyagawa, S.; Yoshikawa, Y.; Fukushima, S.; Kawamura, M.; Yoshioka, D.; Saito, T.; Ueno, T.; Kuratani, T. Tricuspid annular dynamics before and after tricuspid annuloplasty–Three-dimensional transesophageal echocardiography. Circ. J. 2015, 79, 873–879. [Google Scholar] [CrossRef] [Green Version]
- Utsunomiya, H.; Itabashi, Y.; Mihara, H.; Kobayashi, S.; De Robertis, M.A.; Trento, A.; Shiota, T. Usefulness of 3D echocardiographic parameters of tricuspid valve morphology to predict residual tricuspid regurgitation after tricuspid annuloplasty. Eur. Heart J. Cardiovasc. Imaging 2017, 18, 809–817. [Google Scholar] [CrossRef]
- Driessen, N.J.B.; Bouten, C.V.C.; Baaijens, F.P.T. Improved prediction of the collagen fiber architecture in the aortic heart valve. J. Biomech. Eng. 2005, 127, 329–336. [Google Scholar] [CrossRef]
- Liao, J.; Yang, L.; Grashow, J.; Sacks, M.S. The relation between collagen fibril kinematics and mechanical properties in the mitral valve anterior leaflet. J. Biomech. Eng. 2007, 129, 78–87. [Google Scholar] [CrossRef]
- Lee, C.-H.; Zhang, W.; Liao, J.; Carruthers, C.A.; Sacks, J.I.; Sacks, M.S. On the presence of affine fibril and fiber kinematics in the mitral valve anterior leaflet. Biophys. J. 2015, 108, 2074–2087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meador, W.D.; Mathur, M.; Sugerman, G.P.; Jazwiec, T.; Malinowski, M.; Bersi, M.R.; Timek, T.A.; Rausch, M.K. A detailed mechanical and microstructural analysis of ovine tricuspid valve leaflets. Acta Biomater. 2020, 102, 100–113. [Google Scholar] [CrossRef] [PubMed]
- Laurence, D.W.; Ross, C.J.; Jett, S.V.; Johns, C.H.; Echols, A.L.; Baumwart, R.; Towner, R.A.; Liao, J.; Bajona, P.; Wu, Y.; et al. An investigation of regional variations in the biaxial mechanical properties and stress relaxation behaviors of porcine atrioventricular heart valve leaflets. J. Biomech. 2019, 83, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Nadiarynkh, O.; Plotnikov, S.; Campagnola, P.J. Second harmonic generation microscopy for quantitative analysis of collagen fibrillar structure. Nat. Protoc. 2012, 7, 654–669. [Google Scholar] [CrossRef] [PubMed]
- Schriefl, A.J.; Wolinski, H.; Regitnig, P.; Kohlwein, S.D.; Holzapfel, G.A. An automated approach for three-dimensional quantification of fibrillar structures in optically cleared soft biological tissues. J. R. Soc. Interface 2013, 10, 20120760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Akins, M.L.; Murari, K.; Xi, J.; Li, M.-J.; Luby-Phelps, K.; Mahendroo, M.; Li, X. A compact fiber-optic SHG scanning endomicroscope and its application to visualize cervical remodeling during pregnancy. Proc. Natl. Acad. Sci. USA 2012, 109, 12878–12883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaul, R.T.; Nolan, D.R.; Lally, C. Collagen fibre characterisation in arterial tissue under load using SALS. J. Mech. Behav. Biomed. Mater. 2017, 75, 359–368. [Google Scholar] [CrossRef]
- Sacks, M.S.; Smith, D.B.; Hiester, E.D. A small angle light scattering device for planar connective tissue microstructural analysis. Ann. Biomed. Eng. 1997, 25, 678–689. [Google Scholar] [CrossRef]
- Goth, W.; Lesicko, J.; Sacks, M.S.; Tunnell, J.W. Optical-based analysis of soft tissue structures. Annu. Rev. Biomed. Eng. 2016, 18, 357–385. [Google Scholar] [CrossRef] [Green Version]
- Plotnikov, S.; Juneja, V.; Isaacson, A.B.; Mohler, W.A.; Campagnola, P.J. Optical clearing for improved contrast in second harmonic generation imaging of skeletal muscle. Biophys. J. 2006, 90, 328–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goth, W.; Potter, S.; Allen, A.C.B.; Zoldan, J.; Sacks, M.S.; Tunnell, J.W. Non-destructive reflectance mapping of collagen fiber alignment in heart valve leaflets. Ann. Biomed. Eng. 2019, 47, 1250–1264. [Google Scholar] [CrossRef] [PubMed]
- Jett, S.V.; Hudson, L.T.; Baumwart, R.; Bohnstedt, B.N.; Mir, A.; Burkhart, H.M.; Holzapfel, G.A.; Wu, Y.; Lee, C.-H. Integration of polarized spatial frequency domain imaging (pSFDI) with a biaxial mechanical testing system for quantification of load-dependent collagen architecture in soft collagenous tissues. Acta Biomater. 2020, 102, 149–168. [Google Scholar] [CrossRef]
- Duginski, G.A.; Ross, C.J.; Laurence, D.W.; Johns, C.H.; Lee, C.-H. An investigation of the effect of freezing storage on the biaxial mechanical properties of excised porcine tricuspid valve anterior leaflets. J. Mech. Behav. Biomed. Mater. 2020, 101, 103438. [Google Scholar] [CrossRef] [PubMed]
- Ross, C.J.; Laurence, D.W.; Wu, Y.; Lee, C.-H. Biaxial mechanical characterizations of atrioventricular heart valves. J. Vis. Exp. 2019, 146, e59170. [Google Scholar] [CrossRef] [PubMed]
- Fung, Y.C. On mathematical models of stress-strain relationship for living soft tissues. Polym. Mech. 1975, 11, 726–740. [Google Scholar] [CrossRef]
- Cuccia, D.J.; Bevilacqua, F.; Durkin, A.J.; Tromberg, B.J. Modulated imaging: Quantitative analysis and tomography of turbid media in the spatial-frequency domain. Opt. Lett. 2005, 30, 1354–1356. [Google Scholar] [CrossRef]
- Cuccia, D.J.; Bevilacqua, F.P.; Durkin, A.J.; Ayers, F.R.; Tromberg, B.J. Quantitation and mapping of tissue optical properties using modulated imaging. J. Biomed. Opt. 2009, 14, 024012. [Google Scholar] [CrossRef]
- Mazhar, A.; Saggese, S.; Pollins, A.C.; Cardwell, N.L.; Nanney, L.B.; Cuccia, D.J. Noncontact imaging of burn depth and extent in a porcine model using spatial frequency domain imaging. J. Biomed. Opt. 2014, 19, 086019. [Google Scholar] [CrossRef]
- Ruifrok, A.C.; Johnston, D.A. Quantification of histochemical staining by color deconvolution. Anal. Quant. Cytol. Histol. 2001, 23, 291–299. [Google Scholar]
- Heyden, S.; Nagler, A.; Bertoglio, C.; Biehler, J.; Gee, M.W.; Wall, W.A.; Ortiz, M. Material modeling of cardiac valve tissue: Experiments, constitutive analysis and numerical investigation. J. Biomech. 2015, 48, 4287–4296. [Google Scholar] [CrossRef] [PubMed]
- Jett, S.V.; Laurence, D.W.; Kunkel, R.P.; Babu, A.R.; Kramer, K.E.; Baumwart, R.; Towner, R.A.; Wu, Y.; Lee, C.-H. Biaxial mechanical data of porcine atrioventricular valve leaflets. Data Brief 2018, 21, 358–363. [Google Scholar] [CrossRef] [PubMed]
- Fung, Y.C. Structure and Stress-Strain Relationship of Soft Tissues. Am. Zool. 1984, 24, 13–22. [Google Scholar] [CrossRef] [Green Version]
- Lanir, Y. A structural theory for the homogeneous biaxial stress-strain relationships in flat collagenous tissues. J. Biomech. 1979, 12, 423–436. [Google Scholar] [CrossRef]
- Lanir, Y. Constitutive equations for fibrous connective tissues. J. Biomech. 1983, 16, 1–12. [Google Scholar] [CrossRef]
- Pant, A.D.; Thomas, V.S.; Black, A.L.; Verba, T.; Lesicko, J.G.; Amini, R. Pressure-induced microstructural changes in porcine tricuspid valve leaflets. Acta Biomater. 2018, 67, 248–258. [Google Scholar] [CrossRef]
- Kunzelman, K.S.; Cochran, R.P. Stress/strain characteristics of porcine mitral valve tissue: Parallel versus perpendicular collagen orientation. J. Card. Surg. 1992, 7, 71–78. [Google Scholar] [CrossRef]
- Stella, J.A.; Liao, J.; Sacks, M.S. Time-dependent biaxial mechanical behavior of the aortic heart valve leaflet. J. Biomech. 2007, 40, 3169–3177. [Google Scholar] [CrossRef] [Green Version]
- Anssari-Benam, A.; Screen, H.R.C.; Bucchi, A. Insights into the micromechanics of stress-relaxation and creep behaviours in the aortic valve. J. Mech. Behav. Biomed. Mater. 2019, 93, 230–245. [Google Scholar] [CrossRef]
- Yang, B.; Lesicko, J.; Sharma, M.; Hill, M.; Sacks, M.S.; Tunnell, J.W. Polarized light spatial frequency domain imaging for non-destructive quantification of soft tissue fibrous structures. Biomed. Opt. Express 2015, 6, 1520–1533. [Google Scholar] [CrossRef] [Green Version]





| Circumferential Stretch | Radial Stretch | Pixels in the Tissue ROI (pSFDI) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| TVAL | TVPL | TVSL | TVAL | TVPL | TVSL | TVAL | TVPL | TVSL | |
| PPC | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 368,508 | 434,703 | 377,398 |
| 1:1 † | 1.35 ± 0.03 | 1.43 ± 0.02 | 1.39 ± 0.01 | 1.59 ± 0.06 | 1.71 ± 0.03 | 1.77 ± 0.04 | 536,726 | 624,611 | 541,161 |
| 0.5:1 | 1.25 ± 0.02 | 1.28 ± 0.01 | 1.24 ± 0.01 | 1.67 ± 0.05 | 1.81 ± 0.05 | 1.87 ± 0.04 | 550,747 | 626,641 | 533,910 |
| 1:0.5 | 1.35 ± 0.03 | 1.49 ± 0.02 | 1.43 ± 0.02 | 1.52 ± 0.01 | 1.55 ± 0.01 | 1.61 ± 0.05 | 505,182 | 567,840 | 545,645 |
| 2:2 | 1.40 | 1.50 | 1.38 | 1.75 | 1.87 | 1.81 | 584,990 | 697,596 | 625,324 |
| Tissue Layer | Thickness (Respective %) | ||
|---|---|---|---|
| TVAL | TVPL | TVSL | |
| Atrialis (A) | 168.8 ± 18.1 µm (27.5%) | 235.1 ± 6.3 µm (33.3%) | 230.8 ± 16.3 µm (29.5%) |
| Spongiosa (S) | 77.6 ± 12.4 µm (12.6%) | 86.2 ± 11.7 µm (12.2%) | 90.8 ± 13.8 µm (11.6%) |
| Fibrosa (F) | 297.6 ± 15.6 µm (48.5%) | 289.1 ± 15.4 µm (50.0%) | 363.3 ± 41.1 µm (46.4%) |
| Ventricularis (V) | 69.7 ± 4.5 µm (11.4%) | 73.8 ± 3.2 µm (10.5%) | 98.2 ± 14.9 µm (12.5%) |
| Total Thickness | 614.0 ± 23.9 µm | 705.4 ± 22.2 µm | 783.1 ± 62.6 µm |
| Within the TVAL | Within the TVPL | Within the TVSL | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Collagen | Elastin | GAGs | Collagen | Elastin | GAGs | Collagen | Elastin | GAGs | |
| Region 1 vs. Region 2 | 0.593 | 0.377 | 0.933 | 0.266 | 0.720 | 0.025 | 0.029 | 0.983 | 0.004 |
| Region 1 vs. Region 3 | 0.189 | 0.283 | 0.002 | 0.937 | 0.767 | 0.449 | 0.889 | 0.999 | 0.809 |
| Region 2 vs. Region 3 | 0.686 | 0.023 | 0.001 | 0.429 | 0.997 | 0.244 | 0.073 | 0.989 | 0.017 |
| Within Region 1 | Within Region 2 | Within Region 3 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Collagen | Elastin | GAGs | Collagen | Elastin | GAGs | Collagen | Elastin | GAGs | |
| TVAL vs. TVPL | 0.166 | 0.924 | 0.029 | 0.769 | 0.604 | 0.964 | 0.963 | 0.169 | 0.054 |
| TVAL vs. TVSL | 0.253 | 0.311 | 0.952 | 0.981 | 0.018 | 0.004 | 0.019 | 0.999 | 0.004 |
| TVPL vs. TVSL | 0.006 | 0.510 | 0.016 | 0.659 | 0.121 | 0.007 | 0.034 | 0.176 | 0.441 |
| DOA (TVAL) | DOA (TVPL) | DOA (TVSL) | |||||||
| Mean | Skewness | SD | Mean | Skewness | SD | Mean | Skewness | SD | |
| PPC | 0.074 | 0.873 | 0.033 | 0.064 | 0.708 | 0.036 | 0.074 | 0.314 | 0.031 |
| 1:1 | 0.075 | 0.879 | 0.035 | 0.089 | 0.654 | 0.048 | 0.104 | 0.155 | 0.036 |
| 1:0.5 | 0.073 | 1.264 | 0.038 | 0.088 | 0.730 | 0.059 | 0.098 | 0.317 | 0.041 |
| 0.5:1 | 0.082 | 0.564 | 0.032 | 0.089 | 0.257 | 0.034 | 0.109 | −0.012 | 0.036 |
| 2:2 | 0.072 | 0.930 | 0.034 | 0.087 | 0.700 | 0.048 | 0.102 | 0.142 | 0.037 |
| (TVAL) | (TVPL) | (TVSL) | |||||||
| Mean | Skewness | SD | Mean | Skewness | SD | Mean | Skewness | SD | |
| PPC | 61.6° | 0.588 | 30.8° | 85.9° | −0.377 | 34.6° | 70.9° | 0.621 | 36.3° |
| 1:1 | 60.6° | 0.906 | 52.3° | 112.4° | −0.698 | 46.3° | 67.2° | 0.655 | 53.5° |
| 1:0.5 | 69.7° | 0.348 | 38.3° | 103.3° | −0.462 | 37.2° | 77.1° | 0.398 | 46.9° |
| 0.5:1 | 61.6° | 0.889 | 63.4° | 122.3° | −1.042 | 49.3° | 63.6° | 0.799 | 56.8° |
| 2:2 | 60.1° | 0.928 | 52.5° | 111.1° | −0.665 | 47.2° | 66.8° | 0.655 | 53.6° |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hudson, L.T.; Jett, S.V.; Kramer, K.E.; Laurence, D.W.; Ross, C.J.; Towner, R.A.; Baumwart, R.; Lim, K.M.; Mir, A.; Burkhart, H.M.; et al. A Pilot Study on Linking Tissue Mechanics with Load-Dependent Collagen Microstructures in Porcine Tricuspid Valve Leaflets. Bioengineering 2020, 7, 60. https://0-doi-org.brum.beds.ac.uk/10.3390/bioengineering7020060
Hudson LT, Jett SV, Kramer KE, Laurence DW, Ross CJ, Towner RA, Baumwart R, Lim KM, Mir A, Burkhart HM, et al. A Pilot Study on Linking Tissue Mechanics with Load-Dependent Collagen Microstructures in Porcine Tricuspid Valve Leaflets. Bioengineering. 2020; 7(2):60. https://0-doi-org.brum.beds.ac.uk/10.3390/bioengineering7020060
Chicago/Turabian StyleHudson, Luke T., Samuel V. Jett, Katherine E. Kramer, Devin W. Laurence, Colton J. Ross, Rheal A. Towner, Ryan Baumwart, Ki Moo Lim, Arshid Mir, Harold M. Burkhart, and et al. 2020. "A Pilot Study on Linking Tissue Mechanics with Load-Dependent Collagen Microstructures in Porcine Tricuspid Valve Leaflets" Bioengineering 7, no. 2: 60. https://0-doi-org.brum.beds.ac.uk/10.3390/bioengineering7020060