Reactive Oxygen Species, Antioxidant Responses and Implications from a Microbial Modulation Perspective



1
International Faculty of Applied Technology, Yibin University, Yibin 644000, China
2
Department of Life Sciences, Institute for Plant Biology, Technical University of Braunschweig, 38106 Braunschweig, Germany
*
Author to whom correspondence should be addressed.
Biology 2022, 11(2), 155; https://0-doi-org.brum.beds.ac.uk/10.3390/biology11020155
Submission received: 15 December 2021
/
Revised: 14 January 2022
/
Accepted: 17 January 2022
/
Published: 18 January 2022
(This article belongs to the Section Plant Science)
Abstract
:Simple Summary
Environmental conditions are subject to unprecedented changes due to recent progressive anthropogenic activities on our planet. Plants, as the frontline of food security, are susceptible to these changes, resulting in the generation of unavoidable byproducts of metabolism (ROS), which eventually affect their productivity. The response of plants to these unfavorable conditions is highly intricate and depends on several factors, among them are the species/genotype tolerance level, intensity, and duration of stress factors. Defensive mechanisms in plant systems, by nature, are concerned primarily with generating enzymatic and non-enzymatic antioxidants. In addition to this, plant-microbe interactions have been found to improve immune systems in plants suffering from drought and salinity stress.
Abstract
Plants are exposed to various environmental stresses in their lifespan that threaten their survival. Reactive oxygen species (ROS), the byproducts of aerobic metabolism, are essential signalling molecules in regulating multiple plant developmental processes as well as in reinforcing plant tolerance to biotic and abiotic stimuli. However, intensified environmental challenges such as salinity, drought, UV irradiation, and heavy metals usually interfere with natural ROS metabolism and homeostasis, thus aggravating ROS generation excessively and ultimately resulting in oxidative stress. Cellular damage is confined to the degradation of biomolecular structures, including carbohydrates, proteins, lipids, pigments, and DNA. The nature of the double-edged function of ROS as a secondary messenger or harmful oxidant has been attributed to the degree of existing balance between cellular ROS production and ROS removal machinery. The activities of enzyme-based antioxidants, catalase (CAT, EC 1.11.1.6), monodehydroascorbate reductase (MDHAR, E.C.1.6.5.4), dehydroascorbate reductase (DHAR, EC 1.8.5.1), superoxide dismutase (SOD, EC 1.15.1.1), ascorbate peroxidase (APX, EC 1.11.1.11), glutathione reductase (GR, EC 1.6.4.2), and guaiacol peroxidase (GPX, EC 1.11.1.7); and non-enzyme based antioxidant molecules, ascorbate (AA), glutathione (GSH), carotenoids, α-tocopherol, prolines, flavonoids, and phenolics, are indeed parts of the defensive strategies developed by plants to scavenge excess ROS and to maintain cellular redox homeostasis during oxidative stress. This review briefly summarises current knowledge on enzymatic and non-enzymatic antioxidant machinery in plants. Moreover, additional information about the beneficial impact of the microbiome on countering abiotic/biotic stresses in association with roots and plant tissues has also been provided.
1. Introduction
Constant changes in environmental conditions exacerbates unfavorable, stressful conditions such as salinity, drought, extreme temperatures, waterlogging, or heavy metal stress, severely affecting plant growth, development, and yield through inducing changes in plant physiological and biochemical characteristics [1,2].
Plenty of studies have shown that plants are prone to generate both highly reactive oxygen free radicals and slightly reactive non-radicals of oxygen derivatives after subjection to various environmental biotic and/or abiotic stresses [3,4,5]. This generation, which corresponds to nearly 1–2% of the total plant’s oxygen (O2) consumption [6], is highly premised on the presence and functioning of respiratory burst oxidase homologues (RBOHs); and the plant NADPH oxidases [7,8,9]. The oxidative products are collectively referred to as the reactive oxygen species (ROS) and predominantly include superoxide anion radical (O2•−), hydroxyl radical (OH•), perhydroxyl radical (HO2•), singlet oxygen (1O2) and hydrogen peroxide (H2O2) [3,10]. The interconversion rate among unwanted ROS byproducts is extremely high, making them functionally variable and potent oxidants considering their disparity in stability, reactivity, and ability to transport through/across biological membranes [11]. Both ROS types (radical/non-radical) are naturally formed at basal levels in the course of many aerobic metabolic processes such as chloroplast and mitochondrial electron transport chains [12,13], during photorespiration in peroxisomes [14], or in apoplastic spaces [15,16].
Generally, ROS basic levels generated under optimal environmental conditions cannot cause cellular damage due to the expression of stress-responsive genes [17]. This level of ROS generation, based on some evidence, has been suggested to be connected to their limited natural involvement in developmental processes [18] or the regulation of morphogenetic processes associated with phytohormones such as cytokinins and auxins [19]. On the other hand, stressful environmental conditions excessively accelerate cellular ROS concentrations [20] at levels exceeding the antioxidant scavenging capacities [11,17] employed by plants to neutralise excess ROS production [21]. This feature potentially leads to oxidative stress along with damage to membrane lipids, proteins, and nucleic acids and eventually results in cell death [4,22,23]. Abiotic stress-induced programmed cell death (PCD) is a genetically controlled process in which the intensity of detrimental factors is harnessed through increased ROS generation [24]. Environmental stress factors involved in exerting disturbance on the delicate balance between ROS production and ROS removal pathways include but not limited to drought, salinity, high irradiance, extreme temperatures, heavy metals, pollution, and pathogen infection [22,25,26].
Plant adaptation to the oxidative stress caused by elevated ROS concentrations is often consistent with many key factors, among which are stress duration and severity, instantaneous cellular energy status, plant growth stages, ROS cellular level, and antioxidant capacity [27]. Generally, ROS-scavenging systems in plants are comprised of ascorbate (AA), glutathione (GSH), carotenoids, α-tocopherol, prolines, flavonoids, and phenolic compounds as non-enzymatic antioxidants, monodehydroascorbate reductase (MDHAR), dehydroascorbate reductase (DHAR), superoxide dismutase (SOD), catalase (CAT), ascorbate peroxidase (APX), glutathione reductase (GR), and guaiacol peroxidase (GPX) as well as low molecular mass (LMS) antioxidants and/or enzymatic antioxidants [20,21,27,28,29]. It is well reported that increased activity of antioxidant enzymes or non-enzymatic antioxidants in response to unprecedented environmental stresses helps to ameliorate the degree of damage caused by oxidative stress [5,26]. For instance, in green bean (Phaseolus vulgaris L.) plants affected by salinity stress, a subtle increase in the activity of CAT and APX antioxidants was observed in salt-tolerant GS57 and salt-sensitive 4F-89 genotypes, which was consistent with increased fresh and dry weight in these plants [30]. Similarly, an overall increase was reported in antioxidant enzymes (i.e., APX, SOD, GPX, CAT and GR) of some traditional rice plants suffering from drought stress [31]. The upregulation of AA, GSH, phenolics, phytochelatins (as GSH oligomers), and sugars as non-enzyme-based antioxidants was induced in Antarctica in Colobanthus quitensis (Kunth) Bartl by copper stress treatment [32].
Plant growth-promoting rhizobacteria (PGPRs) are known to be beneficial microorganisms and play a contributing role in reinforcing plant response to stressful conditions [33]. Typically, after colonising the rhizosphere or endo-rhizosphere of plants, the PGPRs adopt several direct and/or indirect mechanisms to promote plant growth at the expense of tackling abiotic stresses [33,34,35,36]. Some of these mechanisms include the induction of osmolyte accumulation [37], the activation of the antioxidant defence system [38], the up/downregulation of stress-responsive genes [39,40], and alteration in root morphology in acquisition of stress tolerance [41,42]. PGPR-induced drought or salinity tolerance in biological systems mainly occurs though induction of physical and chemical alterations in plants, which are collectively referred to as PGPR-induced systemic tolerance (IST) [43,44]. For instance, a consortium of three PGPR strains, viz. Bacillus subtilis SM21, Bacillus cereus AR156, and Serratia sp. XY21, have been shown to enhance the drought tolerance of cucumber (Cucumis sativus L.) plants by increasing the activity of SOD and mitigating the expression of genes encoding the cytosolic APX in cucumber leaf tissues [45].
This review aimed to further extend our understanding of antioxidants and their function in protecting plants against oxidative stress, as well as of plant–microbe interactions that confer abiotic stress tolerance in planta. Therefore, we provided a relatively comprehensive overview of major enzymatic and non-enzymatic antioxidants, pointed out the importance of PGPRs in alleviating plant stress, and summarised some current knowledge on them.
2. Enzymatic Defensive Mechanisms
Preventive mechanisms against ROS overaccumulation are the antioxidant capacity by which enzymatic activities confer plant tolerance to adverse environmental conditions [11,46]. Many genetic studies emphasised the significance of expressing these antioxidant enzymes in increasing plant survival rate [47,48]. There are several main ROS-eliminating enzymatic systems in the site of production (subcellular compartments) in plants, namely, SOD, CAT, APX, GPX, GR, MDHAR, and DHAR [11,26].
2.1. Superoxide Dismutase
This omnipresent metalloenzyme, which can be found in almost all aerobic organisms including plants, in all intracellular organelles and apoplastic spaces [49], is actively present at the forefront of defence against oxidative damage from ROS [4,50]. The primary function of superoxide dismutase (SOD, EC 1.15.1.1) is to dismutate superoxide radicals (O•−2) into H2O2 and O2, such that it eventually wards off the formation of OH• by the metal-catalysed Haber–Weiss reaction [26,49,51]. In other words, the catalysing activity of this enzyme regulates the amount of O2 and H2O2, which are the Haber–Weiss reaction substrates, and reduces the risk of producing highly active OH− radicals. Based on the metal ion it binds, three main isozymes/isoforms (SODs) of this enzyme have been introduced thus far in the Arabidopsis thaliana genome: Cu/Zn-SOD gene localised to the chloroplast thylakoids (CSD2), peroxisomes (CSD3), and the cytosol (CSD1), Fe-SOD gene localised to the chloroplast stroma (FSD1), cytosol (FSD1), thylakoids (FSD2, FSD3), and the nucleus (FSD1), and Mn-SOD (MSD1) gene localised to the mitochondrial matrix [11,46,49,52,53,54]. These isoforms have been identified as the main ROS scavengers under drought stress conditions [55]. Additionally, nutrient deficiency (K, P, Mg, Ca, S or N) increased SOD functioning and introduced new isoforms in maize plants [56]. The existence of coherent coordination between peroxidase (an H2O2 scavenger) and enhancement of the activity of chloroplastidial Fe-SOD usually leads to plant protection when CO2 assimilation is intensely reduced; or otherwise results in elevated cytotoxicity in the Haber–Weiss reaction [57,58]. Such coordination during exposure to oxidative stress has already been observed in leaves of tea [59], tobacco [60], onion [61] and cotton [62] plants. In addition to the patented purification and characterisation techniques (e.g., anion exchange chromatography and ammonium sulfate precipitation) [63], another possible approach to increasing SOD levels in leaves of stress-tolerant plant species would be exposing them to environmental stresses such as low temperature, water deficit, or salinity [64,65].
Regardless of SOD types, they are initially encrypted in the nucleus and then transferred to various organs [46,66]. In chloroplast, the SOD enzyme is attached/bonded to the thylakoid membrane as a solution in the stroma [49]. The former one dismantles the superoxide radical immediately at the site of production, and the stroma-soluble form converts the superoxide radicals released in the stroma into H2O2 [4]. Most of the research conducted on SOD was centred on how environmental stresses or genotypes might affect the enzymatic antioxidants alone [57,67] or combined [68,69,70] with other plant items. In a study by Boguszewska et al. [71], it was found that abiotic stress conditions can upregulate the formation of SODs in potato plants. Heavy metals such as zinc oxide nanoparticles (ZnO-NP) escalate NADP-oxidase activity and thus increase the production of superoxide radicals [58]. Apart from their beneficial antioxidative function in plants suffering from salt [72], oxidative [52], or photooxidative [73] stresses, there are indications showing that SODs have an effect on root development [53], germination [46], flowering [74], and not least, chloroplast development [52].
2.2. Catalase
As a Fe-containing homotetrameric enzyme, catalase (CAT, EC 1.11.1.6) is responsible for the catalysation of H2O2 overproduced during light respiration or photorespiration in peroxisomes, H2O2 produced during β-oxidation of fatty acids in glyoxysomes, or H2O2 produced by SOD [4,75]. In all these cases, CAT mediates the dismutation of oxygenated water (H2O2) into water and oxygen in an energy-efficient way [26]. Peroxisomes are the core sites of CAT activity in response to H2O2 production as a result of oxidative stress, purine catabolism, photorespiration, and not least the β-oxidation of fatty acids [10,76]. Nevertheless, a more recent report highlights their presence in the cytosol, chloroplast, mitochondria, and other subcellular compartments [77]. Despite having a high specificity for H2O2, CAT showed a low affinity for organic peroxides [77]. One of the unique characteristics of this enzyme is its high turnover rate and low demand for reducing equivalents [26]. This enzyme is encoded by different CAT genes in several plant species [11]. For example, these include seven CAT genes reported in cotton (Gossypium hirsutum L.) [78], four in cucumber (Cucumis sativus L.) [79], and in rice (Oryza sativa L.) [80], three in (Arabidopsis thaliana L.) Heynh. [81], pumpkin (Cucurbita Linn.) [82], maize (Zea mays L.) [83], Tex-Mex tobacco (Nicotiana plumbaginifolia Viviani) [84], two in common barley (Hordeum vulgare L.) [85], and in tomato (Lycopersicon esculentum Mill.) [86], and one in sweet potato (Ipomoea batatas L.) Poir. [87], and in castor bean (Ricinus communis L. cv. Hale) [88], among others (Table 1).
In Arabidopsis, the first two (CAT1 and CAT2) genes are predominantly present in glyoxysomes, peroxisomes and cytosol, and the latter (CAT3) is present in mitochondria and cytosol, with no presence reported in chloroplasts [4,77]. Recent reports indicate that these genes are primarily expressed in seeds (CAT1, CAT2, CAT3), pollens (CAT1), roots (CAT2), photosynthetic tissues (CAT2), and vascular tissues (CAT3) [26,121,122,123,124]. The involvement of CATs in the growth, development, or senescence of organs such as roots [122], shoots, flower parts (ovule, pollen), seeds [121,123], and leaves [77,125] has been explicitly documented.
Because of low affinity of CATs to H2O2, only high concentrations of H2O2 have the possibility of being eliminated by CATs [126]. In particular, CAT1 is responsible for scavenging H2O2 under abiotic stress conditions, and CAT2 and CAT3 for removing H2O2 to maintain ROS homeostasis both in light and darkness [78]. Stimulation of enzymatic antioxidants such as CATs has been reported in maize plants exposed to arsenate and arsenite [127]. Leaf senescence significantly impacts the concentration reduction of CAT2 rather than CAT1 in tobacco [110]. As for the model plant, Arabidopsis, all three CATs are engaged with photooxidation responses [11,124,128], CAT1 is involved in the plant’s response to drought and salt stress [129], CAT2 is implicated in toxic metal [130], heat [131], cold and salt [132] stress responses and CAT3 is incorporated in drought stress responses [133].
2.3. Ascorbate Peroxidase
The ascorbate peroxidase (APX, EC 1.1.11.1) enzyme functions in the ascorbate-glutathione cycle and regularly exists in chloroplasts (stroma and thylakoid), mitochondria, peroxisomes, cytosols, vacuoles, and the apoplast [4,11]. Five main isoforms of the APX family in rice plants are named after their constituent amino acids and their subcellular localisation [134]. Recent studies on the Arabidopsis genome revealed the presence of nearly nine putative APX genes, localised in the cytosol (APX1, APX2, APX6), peroxisomes (APX3 and APX5), chloroplast stroma (sAPX), chloroplast thylakoid (tAPX) [135,136], and mitochondria (sAPX) [137], with two of them (APC4 and APX7) being annotated as genes lacking H2O2 detoxifying activity and pseudogene, respectively [138]. The critical importance of cytosolic APXs is not so much linked to their conference of plant tolerance to cold [139], salinity (APX2), high light (APX2), heat and drought (APX1) [140,141] stresses as to their depletion (e.g., APX1) which tends to inactivate chloroplastic H2O2 detoxification [142,143].
More importantly, APXs deal with H2O2 signalling during developmental stages [144,145] through scavenging H2O2 toxicity in the chloroplast and cytosol [26,146]. APX enzymes possess a strong affinity for H2O2, indicating that the ascorbate-glutathione (AA-GSH) cycle plays a vital role in controlling ROS levels in cellular organelles [138]. The way this enzyme decomposes H2O2 into water and oxygen is to use ascorbate (ascorbic acid-AA) as a reducing factor [11]. In this reaction, ascorbate is converted to monodehydroascorbate (MDHA) by APX activity, and this compound is also converted to dehydroascorbate (DHA) through a non-enzymatic pathway. Additionally, the ascorbate is a prerequisite substance to maintain the production cycle. In the AA-GSH cycle, monodehydroascorbate reductase (MDHAR) converts the MDHA to AA utilising NADPH. Also, the conversion reaction of DHA to AA is catalysed by dehydroascorbate reductase (DHAR) with redox glutathione (GSH) oxidation in the process (Figure 1) [147].
Similar to the enzyme CAT, APX also presents either in the thylakoid membrane or as a solution in the stroma. The form attached to the thylakoid membrane immediately detoxifies the hydrogen peroxide at the production site, and the stroma-soluble type decomposes the hydrogen peroxide released into the stroma. In comparison to CAT, APX has a stronger affinity for H2O2; hence it is a better scavenger at times of stress [26]. The noticeable concentration of SOD, CAT and APX in the heavy metal (HM)-resistant plant Pteris vittata in proportion to the HM-sensitive plant Pteris ensiformis suggests that the so-called enzymes play a central role in the detoxification of zinc oxide nanoparticles [148].
2.4. Guaiacol Peroxidase (GPX)
With its 40–50 kDa monomers, this heme-containing enzyme, GPX (EC 1.11.1.7), is capable of eliminating excess H2O2, whether by normal or stress-driven metabolism [26]. GPX isoenzymes are localised in vacuoles, cell walls and the cytosol (Figure 1) [149]. The two main functions of GPX are lignin biosynthesis and biotic stress defence by virtue of indole acetic acid (IAA) degradation and H2O2 utilisation in the process [150]. GPX prefers aromatic compounds (e.g., guaiacol and pyrogallol) over non-aromatic ones due to their higher electron-donating ability [151]. As a consequence of its fairly high activity in intra- and extracellular spaces or cell walls, it has been postulated to be a critical enzyme in H2O2 removal [152]. Because this enzyme undertakes both peroxidative and hydroxylic reactions, it comes as no surprise that it can participate in different plant processes such as incorporation in biosynthetic processes, auxin metabolism, cell wall elongation and lignification, as well as pathogenic resistance [153]. A number of environment-born stress factors including herbicide [154], potentially toxic metals [155,156], polycyclic aromatic hydrocarbon (PAH) [157], and ozone (O3) [158] are involved in the induction of GPX activation.
2.5. Glutathione Reductase (GR)
This enzyme (GR, EC 1.6.4.2), also known as flavoprotein oxidoreductase, is the last in the ascorbate-glutathione cycle that is found predominantly in chloroplasts (GR2) and partially in the cytosol (GR1), mitochondria (GR2) [11,26,159] and peroxisomes (GR1) [11]. The proper enzymatic activity of GR mainly springs from its possession of triple domains, including NADPH-binding domain, FAD-binding domain, and a dimerisation domain [160]. The above-mentioned isozymes of GR are involved in tolerance to high light (GR1, GR2) [161,162], salt stress (GR1) [163,164], chilling stress (GR2) [165,166], methyl viologen (MV2+)-induced oxidative stress (GR2) [167], and toxic metals (GR1) [168,169]. Moreover, GR2 has been proven to interfere with developmental processes involving embryo development [170], seed germination [171], root growth, and apical meristems maintenance [172]. Thus, it is follows from the above that abiotic [173,174] and possibly biotic [175] stress factors induce the activity of GR in plants.
GR mediated plant tolerance to stress conditions often refers to its role in ROS detoxification through GSH (reduced glutathione) regeneration [176]. GR has been proven to mediate the conversion of oxidised glutathione (glutathione disulfide-GSSG) to GSH, using the electron donor (reductant) NADPH. Moreover, most of the synthesised GSH is involved in the regeneration of ascorbate (AA) from DHA under DHAR mediation (Figure 1) [26], and the activation of several CO2-stabilizing chloroplast enzymes [4]. Also, GR has the potential to maintain a high GSH/GSSH cellular ratio by catalysing the formation of glutathione disulfide [26], signifying that the pool of GSH consumed by the DHAR reaction is replenished by GR [11]. Increasing GR activity increases the NADP+/NADPH ratio, thus elevating the amount of available NADP+ as the last electron receptor in photosynthetic light reactions, and eventually reducing the likelihood of electrons being transferred to O2 for superoxide radical production [177,178,179].
2.6. Monodehydroascorbate Reductase (MDHAR)
AA regeneration from transient MDHA is catalysed by the FAD (flavin adenine dinucleotide)-dependent MDHAR (E.C.1.6.5.4) enzyme using electrons donated by NADPH [26,180,181]. Notably, MDHAR and APX enzymes are co-localised in the mitochondria and peroxisomes, where their reducing and oxidising activity creates a balance between AA and MDHA pool sizes [10,182]. According to Obara et al. [183], six functional proteins have been encoded from only five Arabidopsis genes. MDHAR isozymes are laid out in different subcellular compartments, including mitochondria (MDHAR5), peroxisomes (MDHAR1) and its membrane glyoxysomes (MDHAR4), chloroplast (MDHAR6), and cytosol (MDHAR2 and MDHAR3) [11,184]. Other than their antioxidant potential, not much information is available on the specific function of MDHAR genes. As a case in point, Eastmond [185] suggested the possible interference of MDHAR4 in different growth stages of Arabidopsis, from germination to post-germination, through senescence. Based on studies conducted so far on rice, tobacco and Arabidopsis, it was revealed that overexpression of MDHAR genes resulted in an increased tolerance to salt, ozone, and osmotic stress as well as higher germination rate and grain weight [186,187].
2.7. Dehydroascorbate Reductase (DHAR)
What is certain in the reduction process of DHA to AA is the DHAR utilisation and transformation of the reduced GSH as an electron donor, which eventually maintains the redox state of plant cells [186,188]. This introduces DHAR as the second catalytic agent along with the so-called MDHAR in regulating the regeneration of the cellular AA pool both in symplast and apoplast [189]. The regulation of AA homeostasis by DHAR during the plant’s developmental processes is no more likely than GSH homeostasis regulation by DHAR [190]. DHAR is usually found in seeds, roots, and shoots with (green) or without (etiolated) chlorophyll content [26]. To date, three types of DHAR proteins have been identified as soluble monomeric enzymes in the chloroplast (DHAR3) and cytosol (DHAR1, DHAR2) [11,188]. A plethora of studies have defined various functions for DHARs at their site of localisation, where, for example, DHAR1 is involved in plant responses to high light, high temperature, and MV2+-induced oxidative stresses [167,191,192], DHAR2 in plant protection against Polyethylene glycol (PEG), salt, drought, and ozone [186,193], or DHAR3 in high light response conference [194].
3. Non-Enzymatic Defensive Mechanisms
Non-enzymatic antioxidants are low molecular weight molecules with specific structures, chemical properties, and locations. Possessing a pivotal role in eliminating free radicals by virtue of donating electrons or hydrogen, these compounds are divided into two groups [4,26]: (1) Fat-soluble membrane antioxidants such as α-tocopherol, carotenoids and xanthophyll; (2) Water-soluble antioxidants such as glutathione, ascorbate and phenolic compounds [195].
3.1. Ascorbic Acid
As one of the most abundant antioxidants in plant systems, ascorbic acid ((AA),vitamin C) has a critical function in plant growth and development [196]. The metabolisation of ROS species by the redox buffer AA could have been explained by safeguarding cells against free oxygen radicals generated under different environmental stress factors [197]. Usually, continuous oxidation of AA is observed under the disguise of such scavenging [196]. Different pathways are in place to ensure the recycling of AA in spite of its slow and time-consuming biosynthesis [196]. Two key enzymes are involved in maintaining the AA homeostasis in living organisms under solicited and unsolicited exogenous stimuli; MDHAR and DHAR [198]. The oxidised (MDHA, DHA), or reduced (MDHAR, DHAR) forms of AA are tapped in AA regeneration [199]. As primary products of AA oxidation, the MDHA molecules can bind each other to form AA or DHA [196]. There has always been the possibility of generating an irreversible form of 2,3-diketogulonic acid in the course of spontaneous DHA hydrolysis [198]. Notably, NAD(P)H figures as an MDHAR electron donor, reducing MDHA to AA in an enzymatic reaction [200].
DHA reduction to AA is mediated by the coupled activity of DHAR and GSH (hydrogen donor) oxidation (Figure 1). There are positive reports on the impact of DHAR overexpression on improved grain and biomass yield in several plants [193,201,202,203,204]. For instance, Kim et al. [204] reported that transgenic japonica rice genotypes with high DHAR expression had increased concentrations of AA and higher crop productivity than wild-type rice. Based on these results, one may speculate on the existence of a direct relationship between AA pools and the grain yield and decreased ROS in plants lacking sufficient environmental adaptation. DHAR down-regulating tobacco plants have shown lower ascorbate levels, signifying the importance of DHAR proteins in AA regeneration [205]. An opposite trend in DHAR expression, however, led to higher AA (oxidised) and GSH (reduced) concentrations [198]. Despite the above statements, a recent finding by Rahantaniaina et al. [188] demonstrated that there is not much difference between an Arabidopsis (Arabidopsis thaliana) mutant without all three DHAR proteins and a wild-type plant with respect to AA regeneration levels, as well as of growth and development. In addition to the analysis of DHAR mutants that show DHARs are required to tie the H2O2 metabolism with GSH oxidation, this finding has brought uncertainty in the efficiency of DAHRs in AA recycling [197].
Physiological studies demonstrate that DAHRs function properly in the course of ascorbate oxidation and increase in ascorbate pool size in a state of high-light stress conditions [206,207]. A study on the Arabidopsis Ddhar mutant identified that the DHAR proteins contributed to increased growth and ascorbate homeostasis under low-light stress [197]. Under high-light stress, both the Ddhar mutant and wild-type plants elevate their ascorbate concentration, with a minor reduction seen in the ascorbate accumulation of Ddhar compared to the wild type that was ascribed to an ascorbate degradation product, the threonate [207]. Accordingly, the authors assumed that the ascorbate pool size determined the activity range of DAHRs and that the necessity of DAHRs activity (to regenerate ascorbate) was a function of higher ascorbate levels. In other words, the smaller the Arabidopsis pool of ascorbate, the lower the DAHRs regeneration activity to sustain ascorbate recycling. It appears that the non-enzymatic reduction of AA by GSH can act as a backup to maintain DAHR activity [197]. In agreement with earlier results [208], a recent report shows that only 30% of the average GSH level in a wild type plant is sufficient to sustain AA recycling under high-light stress conditions [197]. However, an appreciable reduction in AA accumulation as a result of prolonged high-light stress and/or pharmacologically induced GSH deficiency was noticed in the Ddhar pad2-1 quadruple mutants. This was attributed to lower photochemical activity, bleaching, and increased accumulation of threonate [197]. This study put forward the hypothesis that glutathione compensates for the loss of DAHR function under high-light conditions [207]. In the case of Arabidopsis, the activity of DAHR proteins is necessary on the condition that the accumulation level of AA is high (high light) or GSH is less available [207]. Unlike the earlier reports on the indispensable role of DAHRs on ascorbate recycling in tobacco [189,205], recent studies in Arabidopsis underscore the neutral function of DAHRs in ascorbate recycling or maintaining its redox state without affecting ascorbate levels [188].
Even though AA is naturally synthesised in the inner membrane of mitochondria, it may also be found in the cytosol, cell walls, chloroplasts, vacuoles, and apoplasts [209]. AA performs different roles in cells, such as controlling cell cycles, growth and development, and not least having effects on cell wall elongation and the redox level adjustment [196,199]. Ascorbate is a precursor to oxalate and tartrate as well as a cofactor of enzymes involved in the synthesis of glycoproteins rich in hydroxyproline (Hyp), ethylene, gibberellin, and anthocyanin [196]. In addition to being known as a substrate of many peroxidases, AA has been shown to be one of the main components of the ascorbate-glutathione and/or water–water cycle that effectively eliminates ROS [4]. Also, the possibility of AA oxidation in direct reaction with active oxygen species, such as superoxide, singlet oxygen, or hydroxyl radicals, or AA utilisation as a reducing agent in α-tocopherol regeneration to protect membranes from oxidative stress was reported earlier by Parida and Das [147]. Another point is that AA in conjunction with α-tocopherol scavenges lipid peroxyl radicals and prevents the spread of lipid peroxidation in membranes [210,211]. As an example, in chloroplasts, AA functions as a cofactor for the enzyme violaxanthin de-epoxidase (VDE) and participates in the distribution of excessive excitation energy [4].
3.2. Glutathione
Glutathione (GSH) is an intact tripeptide, α-glutamylcysteinyl glycine present in all cell parts, including cytosol, chloroplast, endoplasmic reticulum vacuole and mitochondria, with the highest amount reported in the chloroplast [26,212]. The GSH pool is an essential component of the cellular redox system, which effectively controls the amount of H2O2 through the ascorbate–glutathione cycle and the glutathione cycle [4]. Furthermore, the regeneration process of ascorbate during the ascorbate–glutathione cycle is exclusively premiered on the functioning of the GSH [147]. The antioxidant GSH can act as an eliminator of ROS species. In addition to interfering with the antioxidant defence system, glutathione may also be involved in regulating other processes such as cell entry into the G1 phase and cell differentiation and death. It is believed that this compound is one of the primary sources of non-protein thiols in most plant cells. Given the high reactivity domain of thiol (Sulfhydryl -SH) groups of the low molecular weight compound, GSH, these substances are tapped into many chemical reactions [26]. The nucleophilic nature of the thiol group is critical in the formation of bonds with metals and electrophilic materials. For instance, GSH has been found to positively impact phytochelatin formation via phytochelatin synthase [213]. This reactivity, combined with high stability and solubility of glutathione in water, makes it an ideal substance for protecting plants against environmental stimuli such as heavy metal stress [26,214]. Another beneficial aspect of this molecule is the high reduction potential of the glutathione molecule thanks to its central (C-terminal) cysteine residues [4].
3.3. Proline
The amino acid proline has also been regarded as a non-enzymatic antioxidant in plant systems that can easily counteract the harmful effect of ROS [26,215,216]. Proline synthesis from substrate glutamate (glutamic acid) is a consecutive reaction catalysed by two enzymes, ð1-pyrroline 5-carboxylate (P5C) synthase (P5C-S) and P5C reductase (P5C-R) [215]. The involvement of proline in scavenging ROS damage through direct reaction with ROS has been widely investigated [217,218,219]. For example, some studies revealed that proline osmolytes containing polypeptides could react with H2O2 and OH• to generate stable free radicals by adducting to prolines and hydroxyproline derivatives such as as 4-hydroxyproline and 3-hydroxyproline [217,220]. Further observations by Kaul et al. [218] indicated that proline indirectly scavenges the cellular H2O2 or O2•−. In Brassica juncea plants, proline involvement could dramatically suppress the production of 1O2 in the thylakoids [221]. The 1O2 quenching feature of prolines has been suggested to help stabilise proteins, DNA, and membranes [222]. One of the attractive functions of prolines is to restore the cellular redox balance disrupted by ROS during heavy metal stress [219]. Sharma et al. [223] reported that proline has the potential to prevent zinc (Zn) and cadmium (Cd) from reducing the activity of the cellular enzymes by forming complexes with these metals. Similar protection was earlier observed in copper (Cu)-tolerant Armeria maritime (Mill.) Willd [224]. Multiple studies have identified the progressive impact of proline on antioxidant activities of defensive enzymes such as CAT-in oxidative stress [225,226], SOD- in Cd stress [227], or enzymes related to GSH- in salt stress [228], and AA-GSH cycle- in Cd stress [229].
3.4. α-Tocopherols
α-tocopherol with its three methyl substituents is considered to be one of the most reactive antioxidants among the four well-known lipoperophilic isomers of tocopherol (α, β, γ, and δ) [26]. Regardless of present tocopherol groups, they are responsible for scavenging lipid peroxy radicals, oxygen free radicals and singlet oxygen [230,231,232]. Tocopherols are often localised in the green tissues of plants where photosynthetic organelles and pigments are found [152]. After tocopherol synthesis is completed in the inner envelope of chloroplasts, it has been proposed that they transfer and accumulate in all chloroplast membranes [233,234]. Given that tocopherols are generally incorporated in the protection of lipids, and proteins and pigments are incorporated in the photosynthetic apparatus against oxidative stress, loss of them would be expected to adversely affect growth and photosynthesis in plant systems [235]. There are indications showing that tocopherols’ presence in chloroplasts reinforced plant tolerance to salinity, chilling and water deficit stresses [236,237,238,239].
A combination of five catalysing enzymes (i.e., 4-hydroxyphenylpyruvate dioxygenase, 2-methyl-6-phytylbenzoquinol methyl-transferase, homogentisate phytyl transferases, γ-tocopherol methyl-transferase, tocopherol cyclase (VTE1), and two precursors (homogentisic acid and phytyl diphosphate) compounds are required for tocopherol biosynthesis [240]. Notably, the enzyme γ-tocopherol methyl-transferase (γ-TMT) has been found to catalyse the biosynthesis of α-tocopherol from γ-tocopherol [26,241]. The penultimate step in tocopherol biosynthesis in leaves is dependent on the availability of the VTE1 enzyme [235]. This suggests that VTE1 deficiency would result in a significant reduction in tocopherol synthesis [242].
The way tocopherols are utilised to indirectly protect the structure and function of photosystem II (PSII) is to establish a chemical reaction with O2 and physically quench its excess energy, therefore protecting lipids and other components in the double-membrane of chloroplasts [243]. The prevention of membrane lipid autoxidation through α-tocopherols interaction with lipid radicals (i.e., RO•, ROO•, and RO*) has identified them as protectors of biological membranes [241,244]. On the other hand, the free radical trapper α-tocopherol represses the chain propagation step in the lipid peroxidation (LPO) cycle [26,152,245]. Cellular regeneration of oxidised tocopherols (TOH•) is often driven by coenzyme Q [246] or AA and GSH [247,248]. More specifically, the benzoquinone ring (after full substitution) or the phytyl chain (after full reduction) in tocopherols may act as an efficient antioxidant for 1O2 neutralisation [249,250,251]. In addition to the inhibition of non-enzyme based LPO under stress conditions [241], α-tocopherols are capable of protecting seed storage lipids, activating plant defence responses, functioning in membrane stability, or involving in seedling germination, transcript regulation and intracellular signalling [252,253,254].
3.5. Carotenoids
Carotenoids are tetraterpene antioxidants found in the plastids of photosynthetic and non-photosynthetic tissues of plants [255,256] and are synthesised by geranyl pyrophosphate (GPP) in the course of isoprenoids [26]. Carotenoids containing pure hydrocarbons (carotene) and those with one or more oxygen atoms (xanthophyll) are two main types of carotenoids in plant tissues [257]. Apart from their presence in plants, carotenoids have also been found in algae and photosynthetic microorganisms [152,256]. Their light-harvesting behaviour in chloroplasts encompasses not only light absorption by antenna molecules (450–570 nm) and transfer of energy to chlorophyll (Chl) molecules but also photosynthetic machinery protection [258,259,260]. Carotenoids exert their antioxidative functioning in photosynthetic apparatus through scavenging singlet oxygen activity, reacting with LPOs to halt the chain reaction of ROS production, quenching triplet (3Chl*), and exciting (Chl*) Chl molecules to prevent the formation of singlet oxygen, and dissipating excess excitation energy in the xanthophyll cycle [26,261,262,263]. Depending on plants’ resistance threshold to stressful conditions, the amounts of carotenoids are affected [264,265]. Sugarcane plants with high carotenoids contents exhibited a better adaptation to saline conditions [266].
3.6. Phenolic Compounds
Flavonoids, anthocyanins, tannins, hydroxycinnamic acid esters, and lignins are phenolic compounds that belong to the secondary metabolites arising from the phenylpropanoid pathway in plant tissues [267]. The enzyme phenylalanine ammonia-lyase (PAL) is the tropical initiator of phenylpropanoid, which converts L-phenylalanine to trans-cinnamic acid by deamination. This pathway is necessary for the biosynthesis of secondary metabolites in living cells [268]. Phenolic compounds are naturally synthesised in the cell under optimal conditions, but when there is biotic or abiotic stress, the concentration of these products are significantly affected [152]. Also, any alteration in the activity of biosynthesising or degrading enzymes may influence the amount of these compounds in plant cells.
Flavonoids are a large group of secondary metabolites that are widely distributed among plants and have multiple roles, including contributing to colouring of flowers, seeds and pollen grains, helping in pollination, germination and pollen tube growth, and auxin transport [26], as well as inducing protection against photosynthetic damages caused by excess excitation energy [269]. The latter function is presumably associated with its ROS scavenging capacity [152]. Thanks to the presence of flavonoids, living cells are capable of alleviating the damage of 1O2 on the outer envelope of the chloroplastic membrane [269,270]. The main enzymes involved in the biosynthesis of flavonoids are PAL and chalcone synthase (CHS). Until now, about 12 groups of flavonoids have been identified, the three most important of which are flavonoids, flavonols and anthocyanins [271]. Aside from the roles mentioned above, phenolic compounds also have an antioxidant property in the cell.
Considering the fundamental role of phenolic compounds in reducing or inhibiting lipid auto-oxidation, eliminating oxygen free radicals, quenching singlet oxygen or decomposing peroxides, they have also been known as essential antioxidants responsible for protection against proliferation and advancement of the oxidation chain and defence against reactive oxygen species [271]. The antioxidant properties of phenolic compounds are tied to their chemical structure that may act as an electron or H+ donor. Polyphenols have been shown to chelate intermediate metals such as iron, thus preventing the Fenton and/or Haber–Weiss reaction [272]. Numerous studies in plants demonstrated the impact of potentially toxic heavy metals on the amounts of phenolic compounds, glutathione, phytochelatins, ascorbate, carotenoids, anthocyanins and the activity of PAL enzyme. For instance, the accumulation of these compounds in the presence of zinc oxide nanoparticles was reported [273].
4. Antioxidant Machinery in Plant Systems and Microbial Mediation in Promoting Plant Tolerance
ROS signals are initially perceived and transduced in plants before being translated into sufficient responses [11]. The oxidising nature of ROS aggregates determines the modification/modulation level of potential signalling targets such as transcription factors, kinases, and stress-induced proteins. Notably, this modulation is subject to ROS capability in affecting the protein redox status through oxidation of thiol groups and methionine residues [274]. Thio- and gluta-redoxins are two examples of proteins with the capacity to regulate cellular redox states via their interactive activation/deactivation or reversible oxidation/reduction [275]. For instance, a redox-sensing mechanism was introduced for apoplastic H2O2 perception and transduction by Wu et al. [276]. Protein oxidation or the attachment of carbonyl groups (ketones and aldehydes) to the protein side chains of threonine, lysine, proline, or arginine is known as ROS-mediated carbonylation [277]. This process might lead to protein instability and susceptibility to proteolysis [278,279]. ROS-driven redox perturbations have been found to be transduced by metabolic signals to switch on rapid adaptive mechanisms by mitochondrial/chloroplastic retrograde signalling [280]. Also, ROS can mediate the plastids to the nucleus retrograde-signalling pathway [11]. As such, the nucleus can host the H2O2 generated in plastids at the expense of activating the defence gene expression [281]. There are few reports showing that ROS can interplay with other secondary messengers, such as reactive nitrogen species (RNS) and Ca2+ [11,282,283]. Thanks to their high oxidative potential, ROS interact with nitric oxide (NO) messengers, leading to the generation of (non-)radical RNS products including nitrous acid (HNO2), nitroxyl anion (NO−), nitrosonium cation (NO+), peroxynitrite (ONOO−), nitrate (NO•3), nitric oxide (NO•), and nitric dioxide (NO•2) [284,285,286,287]. These NOx species by nature are involved in plant development, metabolic processes, stress-dependent responses and stomatal closure [288]. Delledonne et al. [289] were the first to report the existence of an interplay between NO and H2O2 during plant hypertension responses. Usually, the crosstalk of ROS and RNS accompanies, by direct or indirect modulation, antioxidant enzymes [285] and may have deleterious or beneficial effects on plant cells, and is highly dependent on the concentration and specific subcellular microcompartment/organelle type [286,290,291].
Increased ROS generation as a result of different environmental stresses has always been naturally responded to in biological systems in multiple ways under the disguise of antioxidative defence mechanisms [11,292]. In addition to this, mitigating the damaging consequences of adverse environmental conditions via the generation of ROS-response antioxidants can also occur by exogenous factors such as plant growth-promoting rhizobacteria (PGPR) [293,294], and plant growth-promoting fungi (PGPF) [295]. It has been suggested that growth-promoting bacteria have the potential to confer enhanced tolerance to abiotic stresses through the induction of physical and chemical alterations in planta, a mechanism that offers protection and is referred to as PGPR-induced systemic tolerance (IST) [296,297,298,299,300,301,302,303]. However, in the case of biotic stress, the so-called eliciting function of PGPRs is known as induced systemic resistance (ISR) [304,305].
The PGPRs function in inducing plant tolerance against abiotic stresses, such as salt, drought, and extreme temperatures, is of key importance in alleviating the adverse effect of climate change on sustainable crop production [299]. As outlined earlier in this review, Wang et al. [45] have shown that the inoculation of Cucumis sativus (cucumber) with a consortium of three PGPR strains could induce systemic tolerance in drought-imposed plants by maintaining root vigour, photosynthetic performance, and increased generation and activity of SOD, CAT and prolines in the leaves. In Lycopersicon esculentum (tomato) plants treated by a Bacillus cereus AR156 supernatant, an appreciable enhancement to drought stress was reported by Wang et al. [306]. The induced tolerance was associated with the increased chlorophyll a and b contents, as well as the enhanced activities of CAT, SOD, and peroxidase (POX). Enhanced salinity tolerance in Panicum turgidum Forssk plants has been attributed to the Arbuscular mycorrhizal fungi contribution by modifying photosynthetic and antioxidant pathways [307]. Similarly, PGPR alleviated drought stress in potato (Solanum tuberosum L.) plants treated with Bacillus subtilis HAS31 by maintaining higher photosynthetic processes, total soluble sugars, proteins and prolines with elevated activity of POX, CAT, and SOD [281]. In another study by Banik et al. [308], it was revealed that PGPR treatment significantly augmented drought tolerance in Agrostis palustris by lowering MDA accumulation and developing osmotic adjustments associated with higher synthesis and accumulation of compatible solutes, including soluble sugars, free amino acids, proteins, and not least the non-enzymatic prolines. Plant inoculation with PGPRs adds up to the available quantity of proline in plant systems under stress conditions [309]. As such, a sizable quantity of prolines was increased in Zea mays L. after inoculation with P. fluorescens under drought stress [310]. A noticeable enhancement in drought tolerance of Lavandula dentate plants treated with PGPR B. thuringiensis was recorded under drought conditions, which was attributed to the increased shoot proline contents [311]. Also, there was an excess secretion of proline in the root of tomato (Lycopersicon esculentum Mill cv. Anakha) plants after exposition to Bacillus polymyxa [312]. Similar to plants, PGPRs secrete osmolytes (e.g., proline) at the time of water scarcity, which cumulatively helps in stimulating plant growth [313].
The application of Trichoderma afroharzianum strain T22 has been shown to enhance tomato (Solanum lycopersicum L.) seed germination under biotic stress conditions by alleviating oxidative damage in salt-stressed seedlings [314]. Similarly, Zhang et al. [282] demonstrated that the PGPF Trichoderma longibrachiatum strain T6 induced the tolerance of wheat seedlings to salt stress through upregulation of SOD, CAT and POX genes, improving the antioxidant defence machinery. Compared to uninoculated rice genotypes Swarna and Swarna sub-1 under severe drought conditions, the PGPFs (Fusarium pallidoroseum strain-10 and Trichoderma harzianum strain-35) inoculated rice genotypes showed higher activity of SOD, POX, and CAT [315]. Table 2 briefly presents the antioxidative responses in selected plant species after exposure to various environmental stresses.
5. Conclusions and Future Prospects
ROS are natural byproducts of many metabolic pathways or their respective electron transport activities in different cellular compartments. Thanks to cellular auto adjustment mechanisms, there is always a homeostatic balance between ROS production and removal machinery which make ROS less harmful to plants in normal environments. However, prolonged environmental stresses such as salinity, chilling, drought, water deficit, and UV radiation can severely exacerbate the production level of ROS by disrupting the natural cellular homeostasis, changing the ROS role as a signalling molecule to a damaging oxidant capable of harming lipids, proteins and DNA. Plants have developed different enzymatic and non-enzymatic antioxidative pathways to alleviate the adverse effects of oxidative damages. The term Induced Systemic Tolerance (IST) has been coined in this review to capture the concept of microbial modulation of enhanced plant tolerance against abiotic stresses through inducing physical and chemical alterations. The beneficial microbiome related to roots and plant tissues suppresses plant stress by a variety of processes. As an example among them, PGPRs are capable of enhancing plant micronutrient uptake, regulating phytohormones homeostasis, and stimulating the plant immune system against biotic and abiotic stresses.
Insofar as the results of ROS experiments under a variety of imposed stresses are concerned, it is not entirely clear how plants deal with stress-induced ROS homeostasis disruption and the consequent cellular degradation. There are still many unsolved questions regarding the simultaneous antioxidative activities of enzymatic antioxidants and their non-enzymatic counterparts. These uncertainties can be considered as hints for further studies in ROS formation and ROS removal machinery. In light of ROS studies, the use of state of art analytical and/or imaging techniques might provide a broader insight into understanding the complex antioxidant networks involved in plant responses to elevated ROS levels. Further, the combined application of advanced functional genomics, proteomics and metabolomics might be helpful in ROS network elaboration. Transgenic techniques can also be used for producing plants with high tolerance to multiple stresses.
Author Contributions
Gathering the required information and drafting the manuscript: P.Z. and E.S.; Revising and editing the manuscript: E.S. and P.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research did not receive any funding from authors or their corresponding Universities.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No associated data marked.
Conflicts of Interest
The authors declared that there is no conflict of interest with reference to this review.
References
- Demirel, U.; Morris, W.L.; Ducreux, L.J.M.; Yavuz, C.; Asim, A.; Tindas, I.; Campbell, R.; Morris, J.A.; Verrall, S.R.; Hedley, P.; et al. Physiological, Biochemical, and Transcriptional Responses to Single and Combined Abiotic Stress in Stress-Tolerant and Stress-Sensitive Potato Genotypes. Front. Plant Sci. 2020, 11, 169. [Google Scholar] [CrossRef] [PubMed]
- Rahman, K.; Rahman, M.; Ahmed, N.; Alam, M.; Rahman, A.; Islam, M.; Hasanuzzaman, M. Morphophysiological changes and reactive oxygen species metabolism in Corchorus olitorius L. under different abiotic stresses. Open Agric. 2021, 6, 549–562. [Google Scholar] [CrossRef]
- Ahsan, H.; Ali, A.; Ali, R. Oxygen free radicals and systemic autoimmunity. Clin. Exp. Immunol. 2003, 131, 398–404. [Google Scholar] [CrossRef] [PubMed]
- Jaleel, C.A.; Riadh, K.; Gopi, R.; Manivannan, P.; Inès, J.; Al-Juburi, H.J.; Chang-Xing, Z.; Hong-Bo, S.; Panneerselvam, R. Antioxidant defense responses: Physiological plasticity in higher plants under abiotic constraints. Acta Physiol. Plant. 2009, 31, 427–436. [Google Scholar] [CrossRef]
- Sewelam, N.; Kazan, K.; Schenk, P.M. Global Plant Stress Signaling: Reactive Oxygen Species at the Cross-Road. Front. Plant Sci. 2016, 7, 187. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharjee, S. Reactive oxygen species and oxidative burst: Roles in stress, senescence and signal. Curr. Sci. 2005, 89, 1113–1121. Available online: https://0-www-jstor-org.brum.beds.ac.uk/stable/24110963 (accessed on 10 November 2021).
- Marino, D.; Andrio, E.; Danchin, E.G.J.; Oger, E.; Gucciardo, S.; Lambert, A.; Puppo, A.; Pauly, N. A Medicago truncatula NADPH oxidase is involved in symbiotic nodule functioning. New Phytol. 2010, 189, 580–592. [Google Scholar] [CrossRef] [Green Version]
- Kadota, Y.; Shirasu, K.; Zipfel, C. Regulation of the NADPH Oxidase RBOHD During Plant Immunity. Plant Cell Physiol. 2015, 56, 1472–1480. [Google Scholar] [CrossRef] [Green Version]
- Stankovic-Valentin, N.; Melchior, F. Control of SUMO and Ubiquitin by ROS: Signaling and disease implications. Mol. Asp. Med. 2018, 63, 3–17. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Dvořák, P.; Krasylenko, Y.; Zeiner, A.; Šamaj, J.; Takáč, T. Signaling Toward Reactive Oxygen Species-Scavenging Enzymes in Plants. Front. Plant Sci. 2021, 11, 618835. [Google Scholar] [CrossRef] [PubMed]
- Gleason, C.; Huang, S.; Thatcher, L.; Foley, R.C.; Anderson, C.R.; Carroll, A.J.; Millar, A.H.; Singh, K.B. Mitochondrial complex II has a key role in mitochondrial-derived reactive oxygen species influence on plant stress gene regulation and defense. Proc. Natl. Acad. Sci. USA 2011, 108, 10768–10773. [Google Scholar] [CrossRef] [Green Version]
- Foyer, C.H. Reactive oxygen species, oxidative signaling and the regulation of photosynthesis. Environ. Exp. Bot. 2018, 154, 134–142. [Google Scholar] [CrossRef]
- Del Río, L.A.; Lopez-Huertas, E. ROS Generation in Peroxisomes and its Role in Cell Signaling. Plant Cell Physiol. 2016, 57, 1364–1376. [Google Scholar] [CrossRef]
- Bindschedler, L.V.; Dewdney, J.; Blee, K.A.; Stone, J.M.; Asai, T.; Plotnikov, J.; Denoux, C.; Hayes, T.; Gerrish, C.; Davies, D.R.; et al. Peroxidase-dependent apoplastic oxidative burst in Arabidopsis required for pathogen resistance. Plant J. 2006, 47, 851–863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roychoudhury, A.; Basu, S. Ascorbate-Glutathione and plant tolerance to various abiotic stresses. In Oxidative Stress in Plants: Causes, Consequences and Tolerance; Anjum, N.A., Umar, S., Ahmad, A., Eds.; IK International Publishers: New Delhi, India, 2012; pp. 177–258. [Google Scholar]
- Foyer, C.H.; Noctor, G. Redox homeostasis and antioxidant signaling: A metabolic interface between stress perception and physiological responses. Plant Cell 2005, 17, 1866–1875. [Google Scholar] [CrossRef] [Green Version]
- Mhamdi, A.; Van Breusegem, F. Reactive oxygen species in plant development. Development 2018, 145, dev164376. [Google Scholar] [CrossRef] [Green Version]
- Xia, X.J.; Zhou, Y.H.; Shi, K.; Zhou, J.; Foyer, C.H.; Yu, J.Q. Interplay between reactive oxygen species and hormones in the control of plant development and stress tolerance. J. Exp. Bot. 2015, 66, 2839–2856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Nath, M.; Bhatt, D.; Prasad, R.; Gill, S.S.; Anjum, N.A.; Tuteja, N. Reactive Oxygen Species Generation-Scavenging and Signaling during Plant-Arbuscular Mycorrhizal and Piriformospora indica Interaction under Stress Condition. Front. Plant Sci. 2016, 7, 1574. [Google Scholar] [CrossRef] [Green Version]
- Baxter, A.; Mittler, R.; Suzuki, N. ROS as key players in plant stress signalling. J. Exp. Bot. 2013, 65, 1229–1240. [Google Scholar] [CrossRef]
- Huang, H.; Ullah, F.; Zhou, D.X.; Yi, M.; Zhao, Y. Mechanisms of ROS regulation of plant development and stress responses. Front. Plant Sci. 2019, 10, 800. [Google Scholar] [CrossRef]
- Petrov, V.; Hille, J.; Mueller-Roeber, B.; Gechev, T.S. ROS-mediated abiotic stress-induced programmed cell death in plants. Front. Plant Sci. 2015, 6, 69. [Google Scholar] [CrossRef] [Green Version]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Van Breusegem, F. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef] [Green Version]
- Miller, G.; Suzuki, N.; Ciftci-Yilmaz, S.; Mittler, R. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef]
- Rasool, S.; Ahmad, A.; Siddiqi, T.O.; Ahmad, P. Changes in growth, lipid peroxidation and some key antioxidant enzymes in chickpea genotypes under salt stress. Acta Physiol. Plant. 2012, 35, 1039–1050. [Google Scholar] [CrossRef]
- Talbi, S.; Romero-Puertas, M.C.; Hernández, A.; Terrón, L.; Ferchichi, A.; Sandalio, L.M. Drought tolerance in a Saharian plant Oudneya africana: Role of antioxidant defences. Environ. Exp. Bot. 2015, 111, 114–126. [Google Scholar] [CrossRef]
- Yasar, F.; Ellialtioglu, S.; Yildiz, K. Effect of salt stress on antioxidant defense systems, lipid peroxidation, and chlorophyll content in green bean. Russ. J. Plant Physiol. 2008, 55, 782–786. [Google Scholar] [CrossRef]
- Nahar, S.; Vemireddy, L.R.; Sahoo, L.; Tanti, B. Antioxidant Protection Mechanisms Reveal Significant Response in Drought-Induced Oxidative Stress in Some Traditional Rice of Assam, India. Rice Sci. 2018, 25, 185–196. [Google Scholar] [CrossRef]
- Contreras, R.A.; Pizarro, M.; Köhler, H.; Sáez, C.A.; Zúñiga, G.E. Copper stress induces antioxidant responses and accumulation of sugars and phytochelatins in Antarctic Colobanthus quitensis (Kunth) Bartl. Biol. Res. 2018, 51, 48. [Google Scholar] [CrossRef]
- Yogendra, S.G.; Singh, U.S.; Sharma, A.K. Bacterial mediated amelioration of drought stress in drought tolerant and susceptible cultivars of rice (Oryza sativa L.). Afr. J. Biotechnol. 2015, 14, 764–773. [Google Scholar] [CrossRef] [Green Version]
- Grover, M.; Ali, S.Z.; Sandhya, V.; Rasul, A.; Venkateswarlu, B. Role of microorganisms in adaptation of agriculture crops to abiotic stresses. World J. Microbiol. Biotechnol. 2010, 27, 1231–1240. [Google Scholar] [CrossRef]
- Rolli, E.; Marasco, R.; Vigani, G.; Ettoumi, B.; Mapelli, F.; DeAngelis, M.L.; Gandolfi, C.; Casati, E.; Previtali, F.; Gerbino, R.; et al. Improved plant resistance to drought is promoted by the root-associated microbiome as a water stress-dependent trait. Environ. Microbiol. 2014, 17, 316–331. [Google Scholar] [CrossRef]
- Berg, G.; Zachow, C.; Müller, H.; Philipps, J.; Tilcher, R. Next-Generation Bio-Products Sowing the Seeds of Success for Sustainable Agriculture. Agronomy 2013, 3, 648–656. [Google Scholar] [CrossRef]
- Sandhya, V.; Ali, S.Z.; Grover, M.; Reddy, G.; Venkateswarlu, B. Effect of plant growth promoting Pseudomonas spp. on compatible solutes, antioxidant status and plant growth of maize under drought stress. Plant Growth Regul. 2010, 62, 21–30. [Google Scholar] [CrossRef]
- Jha, Y.; Subramanian, R.B. PGPR regulate caspase-like activity, programmed cell death, and antioxidant enzyme activity in paddy under salinity. Physiol. Mol. Biol. Plants 2014, 20, 201–207. [Google Scholar] [CrossRef]
- Suárez, R.; Wong, A.; Ramírez, M.; Barraza, A.; Orozco, M.D.C.; Cevallos, M.A.; Lara, M.; Hernández, G.; Iturriaga, G. Improvement of Drought Tolerance and Grain Yield in Common Bean by Overexpressing Trehalose-6-Phosphate Synthase in Rhizobia. Mol. Plant-Microbe Interact. 2008, 21, 958–966. [Google Scholar] [CrossRef] [Green Version]
- Kasim, W.A.; Osman, M.E.; Omar, M.N.; El-Daim, I.A.A.; Bejai, S.; Meijer, J. Control of Drought Stress in Wheat Using Plant-Growth-Promoting Bacteria. J. Plant Growth Regul. 2012, 32, 122–130. [Google Scholar] [CrossRef]
- Pereyra, M.A.; Zalazar, C.; Barassi, C. Root phospholipids in Azospirillum-inoculated wheat seedlings exposed to water stress. Plant Physiol. Biochem. 2006, 44, 873–879. [Google Scholar] [CrossRef] [PubMed]
- Dimkpa, C.; Weinand, T.; Asch, F. Plant-rhizobacteria interactions alleviate abiotic stress conditions. Plant Cell Environ. 2009, 32, 1682–1694. [Google Scholar] [CrossRef]
- Hartmann, A.; Klink, S.; Rothballer, M. Plant Growth Promotion and Induction of Systemic Tolerance to Drought and Salt Stress of Plants by Quorum Sensing Auto-Inducers of the N-acyl-homoserine Lactone Type: Recent Developments. Front. Plant Sci. 2021, 12, 683546. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Dias, M.C.; Freitas, H. Drought and Salinity Stress Responses and Microbe-Induced Tolerance in Plants. Front. Plant Sci. 2020, 11, 591911. [Google Scholar] [CrossRef]
- Wang, C.-J.; Yang, W.; Wang, C.; Gu, C.; Niu, D.-D.; Liu, H.-X.; Wang, Y.-P.; Guo, J.-H. Induction of Drought Tolerance in Cucumber Plants by a Consortium of Three Plant Growth-Promoting Rhizobacterium Strains. PLoS ONE 2012, 7, e52565. [Google Scholar] [CrossRef] [Green Version]
- Dvořák, P.; Krasylenko, Y.; Ovečka, M.; Basheer, J.; Zapletalová, V.; Šamaj, J.; Takáč, T. In vivo light-sheet microscopy resolves localisation patterns of FSD1, a superoxide dismutase with function in root development and osmoprotection. Plant Cell Environ. 2020, 44, 68–87. [Google Scholar] [CrossRef] [PubMed]
- Noctor, G.; Reichheld, J.-P.; Foyer, C.H. ROS-related redox regulation and signaling in plants. Semin. Cell Dev. Biol. 2018, 80, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Foyer, C.H.; Noctor, G. Redox Homeostasis and Signaling in a Higher-CO2World. Annu. Rev. Plant Biol. 2020, 71, 157–182. [Google Scholar] [CrossRef]
- Pilon, M.; Ravet, K.; Tapken, W. The biogenesis and physiological function of chloroplast superoxide dismutases. Biochim. Biophys. Acta Bioenerg. 2011, 1807, 989–998. [Google Scholar] [CrossRef] [Green Version]
- Kliebenstein, D.J.; Monde, R.-A.; Last, R.L. Superoxide Dismutase in Arabidopsis: An Eclectic Enzyme Family with Disparate Regulation and Protein Localization. Plant Physiol. 1998, 118, 637–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alscher, R.G.; Erturk, N.; Heath, L.S. Role of superoxide dismutases (SODs) in controlling oxidative stress in plants. J. Exp. Bot. 2002, 53, 1331–1341. [Google Scholar] [CrossRef]
- Myouga, F.; Hosoda, C.; Umezawa, T.; Iizumi, H.; Kuromori, T.; Motohashi, R.; Shono, Y.; Nagata, N.; Ikeuchi, M.; Shinozaki, K. A Heterocomplex of Iron Superoxide Dismutases Defends Chloroplast Nucleoids against Oxidative Stress and Is Essential for Chloroplast Development in Arabidopsis. Plant Cell 2008, 20, 3148–3162. [Google Scholar] [CrossRef] [Green Version]
- Morgan, M.J.; Lehmann, M.; Schwarzländer, M.; Baxter, C.J.; Sienkiewicz-Porzucek, A.; Williams, T.C.; Schauer, N.; Fernie, A.R.; Fricker, M.D.; Ratcliffe, R.G.; et al. Decrease in Manganese Superoxide Dismutase Leads to Reduced Root Growth and Affects Tricarboxylic Acid Cycle Flux and Mitochondrial Redox Homeostasis. Plant Physiol. 2008, 147, 101–114. [Google Scholar] [CrossRef] [Green Version]
- Jamdhade, A.R.; Sunkar, R.; Hivrale, V.K. Zymographic Method for Distinguishing Different Classes of Superoxide Dismutases in Plants. Methods Mol. Biol. 2017, 1631, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Sales, C.R.G.; Ribeiro, R.V.; Silveira, J.A.G.; Machado, E.C.; Martins, M.O.; Lagôa, A.M.M.A. Superoxide dismutase and ascorbate peroxidase improve the recovery of photosynthesis in sugarcane plants subjected to water deficit and low substrate temperature. Plant Physiol. Biochem. 2013, 73, 326–336. [Google Scholar] [CrossRef] [PubMed]
- Tewari, R.K.; Kumar, P.; Tewari, N.; Srivastava, S.; Sharma, P.N. Macronutrient deficiencies and differential antioxidant responses—Influence on the activity and expression of superoxide dismutase in maize. Plant Sci. 2004, 166, 687–694. [Google Scholar] [CrossRef]
- Yordanova, R.Y. Antioxidative enzymes in barley plants subjected to soil flooding. Environ. Exp. Bot. 2004, 51, 93–101. [Google Scholar] [CrossRef]
- Stephenie, S.; Chang, Y.P.; Gnanasekaran, A.; Esa, N.M.; Gnanaraj, C. An insight on superoxide dismutase (SOD) from plants for mammalian health enhancement. J. Funct. Foods 2020, 68, 103917. [Google Scholar] [CrossRef]
- Li, J.; Arkorful, E.; Cheng, S.; Zhou, Q.; Li, H.; Chen, X.; Sun, K.; Li, X. Alleviation of cold damage by exogenous application of melatonin in vegetatively propagated tea plant (Camellia sinensis L. O. Kuntze). Sci. Hortic. 2018, 238, 356–362. [Google Scholar] [CrossRef]
- Gupta, A.S.; Webb, R.P.; Holaday, A.S.; Allen, R.D. Overexpression of Superoxide Dismutase Protects Plants from Oxidative Stress (Induction of Ascorbate Peroxidase in Superoxide Dismutase-Overexpressing Plants). Plant Physiol. 1993, 103, 1067–1073. [Google Scholar] [CrossRef] [Green Version]
- Rady, M.O.A.; Semida, W.M.; El-Mageed, T.A.A.; Hemida, K.A.; Rady, M.M. Up-regulation of antioxidative defense systems by glycine betaine foliar application in onion plants confer tolerance to salinity stress. Sci. Hortic. 2018, 240, 614–622. [Google Scholar] [CrossRef]
- Yi, X.-P.; Zhang, Y.-L.; Yao, H.-S.; Luo, H.-H.; Gou, L.; Chow, W.S.; Zhang, W.-F. Rapid recovery of photosynthetic rate following soil water deficit and re-watering in cotton plants (Gossypium herbaceum L.) is related to the stability of the photosystems. J. Plant Physiol. 2016, 194, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Lin, R.; Zhang, Z.; Zhou, M. Purification and characterisation of superoxide dismutase from tartary buckwheat leaves. Fagopyrum 1993, 13, 31–34. [Google Scholar]
- Szőllősi, R. Superoxide dismutase (SOD) and abiotic stress tolerance in plants: An overview. In Oxidative Damage to Plants; Ahmad, A., Ed.; Academic Press: New York, NY, USA, 2014; pp. 89–129. [Google Scholar] [CrossRef]
- Río, L.A.D.; Corpas, F.J.; López-Huertas, E.; Palma, J.M. Plant superoxide dismutases: Function under abiotic stress conditions. In Antioxidants and Antioxidant Enzymes in Higher Plants, 1st ed.; Gupta, D., Palma, J., Corpas, F., Eds.; Springer: Cham, Switzerland, 2018; pp. 1–26. [Google Scholar] [CrossRef]
- Bowler, C.; Van Camp, W.; Van Montagu, M.; Inzé, D.; Asada, P.K. Superoxide Dismutase in Plants. Crit. Rev. Plant Sci. 1994, 13, 199–218. [Google Scholar] [CrossRef]
- Lima, C.S.; Ferreira-Silva, S.L.; Carvalho, F.E.L.; Neto, M.C.L.; Aragão, R.M.; Silva, E.N.; Sousa, R.M.J.; Silveira, J.A.G. Antioxidant protection and PSII regulation mitigate photo-oxidative stress induced by drought followed by high light in cashew plants. Environ. Exp. Bot. 2018, 149, 59–69. [Google Scholar] [CrossRef] [Green Version]
- Lin, K.-H.; Kuo, W.-S.; Chiang, C.-M.; Hsiung, T.-C.; Chiang, M.-C.; Lo, H.-F. Study of sponge gourd ascorbate peroxidase and winter squash superoxide dismutase under respective flooding and chilling stresses. Sci. Hortic. 2013, 162, 333–340. [Google Scholar] [CrossRef] [Green Version]
- Ju, Y.-L.; Yue, X.-F.; Zhao, X.-F.; Zhao, H.; Fang, Y.-L. Physiological, micro-morphological and metabolomic analysis of grapevine (Vitis vinifera L.) leaf of plants under water stress. Plant Physiol. Biochem. 2018, 130, 501–510. [Google Scholar] [CrossRef]
- Dias, M.C.; Mariz-Ponte, N.; Santos, C. Lead induces oxidative stress in Pisum sativum plants and changes the levels of phytohormones with antioxidant role. Plant Physiol. Biochem. 2019, 137, 121–129. [Google Scholar] [CrossRef]
- Boguszewska, D.; Grudkowska, M.; Zagdanska, B. Drought-Responsive Antioxidant Enzymes in Potato (Solanum tuberosum L.). Potato Res. 2010, 53, 373–382. [Google Scholar] [CrossRef]
- Shafi, A.; Chauhan, R.; Gill, T.; Swarnkar, M.K.; Sreenivasulu, Y.; Kumar, S.; Kumar, N.; Shankar, R.; Ahuja, P.S.; Singh, A.K. Expression of SOD and APX genes positively regulates secondary cell wall biosynthesis and promotes plant growth and yield in Arabidopsis under salt stress. Plant Mol. Biol. 2015, 87, 615–631. [Google Scholar] [CrossRef]
- Xing, Y.; Cao, Q.; Zhang, Q.; Qin, L.; Jia, W.; Zhang, J. MKK5 Regulates High Light-Induced Gene Expression of Cu/Zn Superoxide Dismutase 1 and 2 in Arabidopsis. Plant Cell Physiol. 2013, 54, 1217–1227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizhsky, L.; Liang, H.; Mittler, R. The Water-Water Cycle Is Essential for Chloroplast Protection in the Absence of Stress. J. Biol. Chem. 2003, 278, 38921–38925. [Google Scholar] [CrossRef] [Green Version]
- Tuzet, A.; Rahantaniaina, M.-S.; Noctor, G. Analyzing the Function of Catalase and the Ascorbate–Glutathione Pathway in H2O2 Processing: Insights from an Experimentally Constrained Kinetic Model. Antioxid. Redox Signal. 2019, 30, 1238–1268. [Google Scholar] [CrossRef]
- Frugoli, J.A.; Zhong, H.H.; Nuccio, M.L.; McCourt, P.; McPeek, M.A.; Thomas, T.L.; McClung, C.R. Catalase is encoded by a multigene family in Arabidopsis thaliana (L.) Heynh. Plant Physiol. 1996, 112, 327–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scandalios, J.G. Oxidative stress: Molecular perception and transduction of signals triggering antioxidant gene defenses. Braz. J. Med. Biol. Res. 2005, 38, 995–1014. [Google Scholar] [CrossRef]
- Wang, W.; Cheng, Y.; Chen, D.; Liu, D.; Hu, M.; Dong, J.; Zhang, X.; Song, L.; Shen, F. The Catalase Gene Family in Cotton: Genome-Wide Characterization and Bioinformatics Analysis. Cells 2019, 8, 86. [Google Scholar] [CrossRef] [Green Version]
- Hu, L.; Yang, Y.; Jiang, L.; Liu, S. The catalase gene family in cucumber: Genome-wide identification and organization. Genet. Mol. Biol. 2016, 39, 408–415. [Google Scholar] [CrossRef] [Green Version]
- Joo, J.; Lee, Y.H.; Song, S.I. Rice CatA, CatB, and CatC are involved in environmental stress response, root growth, and photorespiration, respectively. J. Plant Biol. 2014, 57, 375–382. [Google Scholar] [CrossRef]
- Du, Y.-Y.; Wang, P.; Chen, J.; Song, C.-P. Comprehensive Functional Analysis of the Catalase Gene Family in Arabidopsis thaliana. J. Integr. Plant Biol. 2008, 50, 1318–1326. [Google Scholar] [CrossRef]
- Esaka, M.; Yamada, N.; Kitabayashi, M.; Setoguchi, Y.; Tsugeki, R.; Kondo, M.; Nishimura, M. cDNA cloning and differential gene expression of three catalases in pumpkin. Plant Mol. Biol. 1997, 33, 141–155. [Google Scholar] [CrossRef] [PubMed]
- Guan, L.; Scandalios, J.G. Developmentally related responses of maize catalase genes to salicylic acid. Proc. Natl. Acad. Sci. USA 1995, 92, 5930–5934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willekens, H.; Villarroel, R.; Van Montagu, M.; Inzé, D.; Van Camp, W. Molecular identification of catalases from Nicotiana plumbaginifolia (L.). FEBS Lett. 1994, 352, 79–83. [Google Scholar] [CrossRef] [Green Version]
- Skadsen, R.W.; Schulze-Lefert, P.; Herbst, J.M. Molecular cloning, characterization and expression analysis of two catalase isozyme genes in barley. Plant Mol. Biol. 1995, 29, 1005–1014. [Google Scholar] [CrossRef]
- Drory, A.; Woodson, W.R. Molecular cloning and nucleotide sequence of a cDNA encoding catalase from tomato. Plant Physiol. 1992, 100, 1605–1606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.-J.; Wu, S.-D.; Huang, G.-J.; Shen, C.-Y.; Afiyanti, M.; Li, W.-J.; Lin, Y.-H. Expression of a cloned sweet potato catalase SPCAT1 alleviates ethephon-mediated leaf senescence and H2O2 elevation. J. Plant Physiol. 2012, 169, 86–97. [Google Scholar] [CrossRef] [PubMed]
- González, E. The C-terminal domain of plant catalases Implications for a glyoxysomal targeting sequence. JBIC J. Biol. Inorg. Chem. 1991, 199, 211–215. [Google Scholar] [CrossRef]
- Raza, A.; Su, W.; Gao, A.; Mehmood, S.; Hussain, M.; Nie, W.; Lv, Y.; Zou, X.; Zhang, X. Catalase (CAT) Gene Family in Rapeseed (Brassica napus L.): Genome-Wide Analysis, Identification, and Expression Pattern in Response to Multiple Hormones and Abiotic Stress Conditions. Int. J. Mol. Sci. 2021, 22, 4281. [Google Scholar] [CrossRef] [PubMed]
- Eising, R.; Trelease, R.N.; Ni, W. Biogenesis of catalase in glyoxysomes and leaf-type peroxisomes of sunflower cotyledons. Arch. Biochem. Biophys. 1990, 278, 258–264. [Google Scholar] [CrossRef]
- Kleff, S.; Trelease, R.N.; Eising, R. Nucleotide and deduced amino acid sequence of a putative higher molecular weight precursor for catalase in sunflower cotyledons. Biochim. Biophys. Acta-Bioenerg. 1994, 1224, 463–466. [Google Scholar] [CrossRef]
- Azpilicueta, C.E.; Pena, L.B.; Tomaro, M.L.; Gallego, S.M. Modifications in catalase activity and expression in developing sunflower seedlings under cadmium stress. Redox Rep. 2008, 13, 40–46. [Google Scholar] [CrossRef]
- Ni, W.; Trelease, R.N. Post-Transcriptional Regulation of Catalase Isozyme Expression in Cotton Seeds. Plant Cell 1991, 3, 737–744. [Google Scholar] [CrossRef] [Green Version]
- Isin, S.H.; Allen, R.D. Isolation and characterization of a pea catalase cDNA. Plant Mol. Biol. 1991, 17, 1263–1265. [Google Scholar] [CrossRef]
- Corpas, F.J.; Palma, J.M.; Sandalio, L.M.; Lopez-Huertas, E.; Romero-Puertas, M.C.; Barroso, J.B.; Del Río, L.A. Purification of Catalase from Pea Leaf Peroxisomes: Identification of Five Different Isoforms. Free. Radic. Res. 1999, 31, 235–241. [Google Scholar] [CrossRef]
- Morita, S.; Tasaka, M.; Fujisawa, H.; Ushimaru, T.; Tsuji, H. A cDNA Clone Encoding a Rice Catalase Isozyme. Plant Physiol. 1994, 105, 1015–1016. [Google Scholar] [CrossRef]
- Alam, N.B.; Ghosh, A. Comprehensive analysis and transcript profiling of Arabidopsis thaliana and Oryza sativa catalase gene family suggests their specific roles in development and stress responses. Plant Physiol. Biochem. 2018, 123, 54–64. [Google Scholar] [CrossRef]
- Chevalier, C.; Yamaguchi, J.; McCourt, P. Nucleotide Sequence of a cDNA for Catalase from Arabidopsis thaliana. Plant Physiol. 1992, 99, 1726–1728. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Cui, L.; Xie, Z.; Zhang, Z.; Liu, E.; Peng, X. Two NCA1 isoforms interact with catalase in a mutually exclusive manner to redundantly regulate its activity in rice. BMC Plant Biol. 2019, 19, 105. [Google Scholar] [CrossRef] [PubMed]
- Contento, A.L.; Bassham, D.C. Increase in catalase-3 activity as a response to use of alternative catabolic substrates during sucrose starvation. Plant Physiol. Biochem. 2010, 48, 232–238. [Google Scholar] [CrossRef]
- Mhamdi, A.; Queval, G.; Chaouch, S.; Vanderauwera, S.; Van Breusegem, F.; Noctor, G. Catalase function in plants: A focus on Arabidopsis mutants as stress-mimic models. J. Exp. Bot. 2010, 61, 4197–4220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isin, S.H. Regulation of Catalase Gene Expression in Soybean. Ph.D. Thesis, Texas Tech. University, Lubbock, TX, USA, 1992. [Google Scholar]
- Redinbaugh, M.G.; Wadsworth, G.J.; Scandalios, J.G. Characterization of catalase transcripts and their differential expression in maize. Biochim. Biophys. Acta-Gene Struct. Expr. 1988, 951, 104–116. [Google Scholar] [CrossRef]
- Wadsworth, G.J.; Scandalios, J.G. Differential expression of the maize catalase genes during kernel development: The role of steady-state mRNA levels. Dev. Genet. 1989, 10, 304–310. [Google Scholar] [CrossRef]
- Guan, L.; Scandalios, J.G. Characterization of the catalase antioxidant defense gene Cat1 of maize, and its developmentally regulated expression in transgenic tobacco. Plant J. 1993, 3, 527–536. [Google Scholar] [CrossRef] [PubMed]
- Guan, L.; Polidoros, A.N.; Scandalios, J.G. Isolation, characterization and expression of the maize Cat2 catalase gene. Plant Mol. Biol. 1996, 30, 913–924. [Google Scholar] [CrossRef]
- Abler, M.L.; Scandalios, J.G. The CAT-2 null phenotype in maize is likely due to a DNA insertion into the Cat2 gene. Theor. Appl. Genet. 1991, 81, 635–640. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; An, C.S. Differential expression of three catalase genes in hot pepper (Capsicum annuum L.). Mol. Cells 2005, 20, 247–255. [Google Scholar]
- Schultes, N.P.; Zelitch, I.; Mcgonigle, B.; Nelson, T. The Primary Leaf Catalase Gene from Nicotiana tabacum and Nicotiana sylvestris. Plant Physiol. 1994, 106, 399–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niewiadomska, E.; Polzien, L.; Desel, C.; Rozpądek, P.; Miszalski, Z.; Krupinska, K. Spatial patterns of senescence and development-dependent distribution of reactive oxygen species in tobacco (Nicotiana tabacum) leaves. J. Plant Physiol. 2009, 166, 1057–1068. [Google Scholar] [CrossRef]
- Suzuki, M.; Ario, T.; Hattori, T.; Nakamura, K.; Asahi, T. Isolation and characterization of two tightly linked catalase genes from castor bean that are differentially regulated. Plant Mol. Biol. 1994, 25, 507–516. [Google Scholar] [CrossRef]
- Luna, C.M.; Pastori, G.M.; Driscoll, S.; Groten, K.; Bernard, S.; Foyer, C.H. Drought controls on H2O2 accumulation, catalase (CAT) activity and CAT gene expression in wheat. J. Exp. Bot. 2005, 56, 417–423. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Shah, D.M. Isolation and characterization of a potato catalase cDNA. Plant Physiol. 1995, 108, 1748–1749. [Google Scholar]
- Almeida, J.M.; Fidalgo, F.; Confraria, A.; Santos, A.; Pires, H.; Santos, I. Effect of hydrogen peroxide on catalase gene expression, isoform activities and levels in leaves of potato sprayed with homobrassinolide and ultrastructural changes in mesophyll cells. Funct. Plant Biol. 2005, 32, 707–720. [Google Scholar] [CrossRef]
- Kerdnaimongkol, K.; Woodson, W.R. Inhibition of Catalase by Antisense RNA Increases Susceptibility to Oxidative Stress and Chilling Injury in Transgenic Tomato Plants. J. Am. Soc. Hortic. Sci. 1999, 124, 330–336. [Google Scholar] [CrossRef] [Green Version]
- Meena, M.; Zehra, A.; Dubey, M.K.; Aamir, M.; Gupta, V.K.; Upadhyay, R.S. Comparative Evaluation of Biochemical Changes in Tomato (Lycopersicon esculentum Mill.) Infected by Alternaria alternata and Its Toxic Metabolites (TeA, AOH, and AME). Front. Plant Sci. 2016, 7, 1408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Havir, E.A.; McHale, N.A. Regulation of Catalase Activity in Leaves of Nicotiana sylvestris by High CO2. Plant Physiol. 1989, 89, 952–957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, H.; Imaseki, H. cDNA for Catalase from Etiolated Mung Bean (Vigna radiata) Hypocotyls. Plant Physiol. 1993, 102, 691–692. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, M.; Grief, J.; Feierabend, J. Mode of translational activation of the catalase (cat1) mRNA of rye leaves (Secale cereale L.) and its control through blue light and reactive oxygen. Planta 2006, 223, 835–846. [Google Scholar] [CrossRef]
- Sakajo, S.; Nakamura, K.; Asahi, T. Molecular cloning and nucleotide sequence of full-length cDNA for sweet potato catalase mRNA. JBIC J. Biol. Inorg. Chem. 1987, 165, 437–442. [Google Scholar] [CrossRef]
- Su, T.; Wang, P.; Li, H.; Zhao, Y.; Lu, Y.; Dai, P.; Ren, T.; Wang, X.; Li, X.; Shao, Q.; et al. The Arabidopsiscatalase triple mutant reveals important roles of catalases and peroxisome-derived signaling in plant development. J. Integr. Plant Biol. 2018, 60, 591–607. [Google Scholar] [CrossRef]
- Yang, Z.; Mhamdi, A.; Noctor, G. Analysis of catalase mutants underscores the essential role of CATALASE2 for plant growth and day length-dependent oxidative signalling. Plant Cell Environ. 2018, 42, 688–700. [Google Scholar] [CrossRef] [PubMed]
- Palma, J.M.; Mateos, R.M.; Lopez-Jaramillo, F.J.; Ruiz, M.R.; González-Gordo, S.; Lechuga-Sancho, A.; Corpas, F.J. Plant catalases as NO and H2S targets. Redox Biol. 2020, 34, 101525. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Li, C.; Ren, H.; Zhao, T.; Li, Q.; Wang, S.; Zhang, Y.; Xiao, F.; Wang, X. BAK1 Mediates Light Intensity to Phosphorylate and Activate Catalases to Regulate Plant Growth and Development. Int. J. Mol. Sci. 2020, 21, 1437. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Ji, T.-T.; Li, T.-T.; Tian, Y.-Y.; Wang, L.-F.; Liu, W.-C. Jasmonic acid promotes leaf senescence through MYC2-mediated repression of CATALASE2 expression in Arabidopsis. Plant Sci. 2020, 299, 110604. [Google Scholar] [CrossRef]
- Wang, L.; Chen, W.J.; Wang, Q.; Eneji, A.E.; Li, Z.H.; Duan, L.S. Coronatine Enhances Chilling Tolerance in Cucumber (Cucumis sativus L.) Seedlings by Improving the Antioxidative Defence System. J. Agron. Crop. Sci. 2009, 195, 377–383. [Google Scholar] [CrossRef]
- Mylona, P.; Polidoros, A.; Scandalios, J.G. Modulation of antioxidant responses by arsenic in maize. Free Radic. Biol. Med. 1998, 25, 576–585. [Google Scholar] [CrossRef]
- Vandenabeele, S.; Vanderauwera, S.; Vuylsteke, M.; Rombauts, S.; Langebartels, C.; Seidlitz, H.K.; Zabeau, M.; Van Montagu, M.; Inzé, D.; Van Breusegem, F. Catalase deficiency drastically affects gene expression induced by high light in Arabidopsis thaliana. Plant J. 2004, 39, 45–58. [Google Scholar] [CrossRef]
- Xing, Y.; Jia, W.; Zhang, J. AtMEK1 mediates stress-induced gene expression of CAT1 catalase by triggering H2O2 production in Arabidopsis. J. Exp. Bot. 2007, 58, 2969–2981. [Google Scholar] [CrossRef]
- Corpas, F.J.; Barroso, J.B. Lead-induced stress, which triggers the production of nitric oxide (NO) and superoxide anion (O2−) in Arabidopsis peroxisomes, affects catalase activity. Nitric Oxide 2017, 68, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Ono, M.; Isono, K.; Sakata, Y.; Taji, T. CATALASE2 plays a crucial role in long-term heat tolerance of Arabidopsis thaliana. Biochem. Biophys. Res. Commun. 2020, 534, 747–751. [Google Scholar] [CrossRef] [PubMed]
- Bueso, E.; Alejandro, S.; Carbonell, P.; Perez-Amador, M.A.; Fayos, J.; Bellés, J.M.; Rodriguez, P.L.; Serrano, R.; Carbonell-Bejerano, P. The lithium tolerance of the Arabidopsiscat2mutant reveals a cross-talk between oxidative stress and ethylene. Plant J. 2007, 52, 1052–1065. [Google Scholar] [CrossRef]
- Zou, J.-J.; Li, X.-D.; Ratnasekera, D.; Wang, C.; Liu, W.-X.; Song, L.-F.; Zhang, W.-Z.; Wu, W.-H. Arabidopsis calcium-dependent protein kinase8 and CATALASE3 Function in Abscisic Acid-Mediated Signaling and H2O2 Homeostasis in Stomatal Guard Cells under Drought Stress. Plant Cell 2015, 27, 1445–1460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, P.; Dubey, R. Ascorbate peroxidase from rice seedlings: Properties of enzyme isoforms, effects of stresses and protective roles of osmolytes. Plant Sci. 2004, 167, 541–550. [Google Scholar] [CrossRef]
- Maruta, T.; Inoue, T.; Noshi, M.; Tamoi, M.; Yabuta, Y.; Yoshimura, K.; Ishikawa, T.; Shigeoka, S. Cytosolic ascorbate peroxidase 1 protects organelles against oxidative stress by wounding-and jasmonate-induced H2O2 in Arabidopsis plants. Biochim. Biophys. Acta-Gen. Subj. 2012, 1820, 1901–1907. [Google Scholar] [CrossRef]
- Maruta, T.; Sawa, Y.; Shigeoka, S.; Ishikawa, T. Diversity and Evolution of Ascorbate Peroxidase Functions in Chloroplasts: More Than Just a Classical Antioxidant Enzyme? Plant Cell Physiol. 2016, 57, pcv203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chew, O.; Whelan, J.; Millar, A.H. Molecular Definition of the Ascorbate-Glutathione Cycle in Arabidopsis Mitochondria Reveals Dual Targeting of Antioxidant Defenses in Plants. J. Biol. Chem. 2003, 278, 46869–46877. [Google Scholar] [CrossRef] [Green Version]
- Granlund, I.; Storm, P.; Schubert, M.; García-Cerdán, J.G.; Funk, C.; Schröder, W.P. The TL29 Protein is Lumen Located, Associated with PSII and Not an Ascorbate Peroxidase. Plant Cell Physiol. 2009, 50, 1898–1910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Buer, J.; Cvetkovic, J.; Baier, M. Cold regulation of plastid ascorbate peroxidases serves as a priming hub controlling ROS signaling in Arabidopsis thaliana. BMC Plant Biol. 2016, 16, 163. [Google Scholar] [CrossRef] [Green Version]
- Rossel, J.B.; Walter, P.B.; Hendrickson, L.; Chow, W.S.; Poole, A.; Mullineaux, P.M.; Pogson, B.J. A mutation affecting ascorbate peroxidase 2 gene expression reveals a link between responses to high light and drought tolerance. Plant Cell Environ. 2005, 29, 269–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, N.; Miller, G.; Sejima, H.; Harper, J.; Mittler, R. Enhanced seed production under prolonged heat stress conditions inArabidopsis thalianaplants deficient in cytosolic ascorbate peroxidase 2. J. Exp. Bot. 2012, 64, 253–263. [Google Scholar] [CrossRef] [Green Version]
- Pnueli, L.; Liang, H.; Rozenberg, M.; Mittler, R. Growth suppression, altered stomatal responses, and augmented induction of heat shock proteins in cytosolic ascorbate peroxidase (Apx1)-deficient Arabidopsis plants. Plant J. 2003, 34, 187–203. [Google Scholar] [CrossRef] [Green Version]
- Davletova, S.; Rizhsky, L.; Liang, H.; Shengqiang, Z.; Oliver, D.J.; Coutu, J.; Shulaev, V.; Schlauch, K.; Mittler, R. Cytosolic Ascorbate Peroxidase 1 Is a Central Component of the Reactive Oxygen Gene Network of Arabidopsis. Plant Cell 2005, 17, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Pandey, S.; Fartyal, D.; Agarwal, A.; Shukla, T.; James, D.; Kaul, T.; Negi, Y.K.; Arora, S.; Reddy, M.K. Abiotic Stress Tolerance in Plants: Myriad Roles of Ascorbate Peroxidase. Front. Plant Sci. 2017, 8, 581. [Google Scholar] [CrossRef] [Green Version]
- Chin, D.-C.; Kumar, R.S.; Suen, C.-S.; Chien, C.-Y.; Hwang, M.-J.; Hsu, C.-H.; Xuhan, X.; Lai, Z.X.; Yeh, K.-W. Plant Cytosolic Ascorbate Peroxidase with Dual Catalytic Activity Modulates Abiotic Stress Tolerances. iScience 2019, 16, 31–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Letnik, I.; Hacham, Y.; Dobrev, P.; Ben-Daniel, B.-H.; Vanková, R.; Amir, R.; Miller, G. ASCORBATE PEROXIDASE6 Protects Arabidopsis Desiccating and Germinating Seeds from Stress and Mediates Cross Talk between Reactive Oxygen Species, Abscisic Acid, and Auxin. Plant Physiol. 2014, 166, 370–383. [Google Scholar] [CrossRef] [Green Version]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, M.; Ma, L.Q.; Singh, N.; Singh, S. Antioxidant responses of hyper-accumulator and sensitive fern species to arsenic. J. Exp. Bot. 2005, 56, 1335–1342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asada, K. Ascorbate peroxidase—A hydrogen peroxide-scavenging enzyme in plants. Physiol. Plant. 1992, 85, 235–241. [Google Scholar] [CrossRef]
- Caverzan, A.; Casassola, A.; Brammer, S.P. Reactive oxygen species and antioxidant enzymes involved in plant tolerance to stress. In Abiotic and Biotic Stress in Plants-Recent Advances and Future Perspectives; Shanker, A.K., Shanker, C., Eds.; IntechOpen: London, UK, 2016. [Google Scholar] [CrossRef] [Green Version]
- Asada, K. The water-water cycle in chloroplasts: Scavenging of Active Oxygens and Dissipation of Excess Photons. Annu. Rev. Plant Biol. 1999, 50, 601–639. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive Oxygen Species, Oxidative Damage, and Antioxidative Defense Mechanism in Plants under Stressful Conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef] [Green Version]
- Passardi, F.; Longet, D.; Penel, C.; Dunand, C. The class III peroxidase multigenic family in rice and its evolution in land plants. Phytochemistry 2004, 65, 1879–1893. [Google Scholar] [CrossRef]
- Ivanov, S.; Shopova, E.; Kerchev, P.; Sergiev, I.; Miteva, L.; Polizoev, D.; Alexieva, V. Long-term impact of sublethal atrazine perturbs the redox homeostasis in pea (Pisum sativum L.) plants. Protoplasma 2012, 250, 95–102. [Google Scholar] [CrossRef]
- Mishra, S.; Jha, A.B.; Dubey, R.S. Arsenite treatment induces oxidative stress, upregulates antioxidant system, and causes phytochelatin synthesis in rice seedlings. Protoplasma 2010, 248, 565–577. [Google Scholar] [CrossRef]
- Srivastava, S.; Dubey, R.S. Manganese-excess induces oxidative stress, lowers the pool of antioxidants and elevates activities of key antioxidative enzymes in rice seedlings. Plant Growth Regul. 2010, 64, 1–16. [Google Scholar] [CrossRef]
- Song, H.; Wang, Y.-S.; Sun, C.-C.; Wang, Y.-T.; Peng, Y.-L.; Cheng, H. Effects of pyrene on antioxidant systems and lipid peroxidation level in mangrove plants, Bruguiera gymnorrhiza. Ecotoxicology 2012, 21, 1625–1632. [Google Scholar] [CrossRef]
- Li, C.; Wang, T.; Li, Y.; Zheng, Y.-H.; Jiang, G.-M. Flixweed Is More Competitive than Winter Wheat under Ozone Pollution: Evidences from Membrane Lipid Peroxidation, Antioxidant Enzymes and Biomass. PLoS ONE 2013, 8, e60109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kataya, A.R.A.; Reumann, S. Arabidopsis glutathione reductase 1 is dually targeted to peroxisomes and the cytosol. Plant Signal. Behav. 2010, 5, 171–175. [Google Scholar] [CrossRef] [Green Version]
- Berkholz, D.S.; Faber, H.R.; Savvides, S.N.; Karplus, P.A. Catalytic Cycle of Human Glutathione Reductase Near 1 Å Resolution. J. Mol. Biol. 2008, 382, 371–384. [Google Scholar] [CrossRef] [Green Version]
- Karpinski, S.; Escobar, C.; Karpinska, B.; Creissen, G.; Mullineaux, P.M. Photosynthetic electron transport regulates the expression of cytosolic ascorbate peroxidase genes in Arabidopsis during excess light stress. Plant Cell 1997, 9, 627–640. [Google Scholar] [CrossRef] [Green Version]
- Müller-Schüssele, S.J.; Wang, R.; Gütle, D.D.; Romer, J.; Rodriguez-Franco, M.; Scholz, M.; Buchert, F.; Lüth, V.M.; Kopriva, S.; Dörmann, P.; et al. Chloroplasts require glutathione reductase to balance reactive oxygen species and maintain efficient photosynthesis. Plant J. 2020, 103, 1140–1154. [Google Scholar] [CrossRef]
- Csiszár, J.; Brunner, S.; Horváth, E.; Bela, K.; Ködmön, P.; Riyazuddin, R.; Gallé, Á.; Hurton, Á.; Papdi, C.; Szabados, L.; et al. Exogenously applied salicylic acid maintains redox homeostasis in salt-stressed Arabidopsis gr1 mutants expressing cytosolic roGFP1. Plant Growth Regul. 2018, 86, 181–194. [Google Scholar] [CrossRef]
- Wang, Q.; Pu, Y.; Yang, D.; Yin, X.; He, Z.; Yang, Y.; Yang, Y. Molecular cloning and characterization of the glutathione reductase gene from Stipa purpurea. Biochem. Biophys. Res. Commun. 2018, 495, 1851–1857. [Google Scholar] [CrossRef]
- Kornyeyev, D.; Logan, B.A.; Payton, P.R.; Allen, R.D.; Holaday, A.S. Elevated chloroplastic glutathione reductase activities decrease chilling-induced photoinhibition by increasing rates of photochemistry, but not thermal energy dissipation, in transgenic cotton. Funct. Plant Biol. 2003, 30, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Logan, B.A.; Monteiro, G.; Kornyeyev, D.; Payton, P.; Allen, R.D.; Holaday, A.S. Transgenic overproduction of glutathione reductase does not protect cotton, Gossypium hirsutum (Malvaceae), from photoinhibition during growth under chilling conditions. Am. J. Bot. 2003, 90, 1400–1403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Ding, H.; Chen, Q.; Ouyang, L.; Li, S.; Zhang, J. Enhanced Tolerance to Methyl Viologen-Mediated Oxidative Stress via AtGR2 Expression From Chloroplast Genome. Front. Plant Sci. 2019, 10, 1178. [Google Scholar] [CrossRef]
- Guo, B.; Liu, C.; Li, H.; Yi, K.; Ding, N.; Li, N.; Lin, Y.; Fu, Q. Endogenous salicylic acid is required for promoting cadmium tolerance of Arabidopsis by modulating glutathione metabolisms. J. Hazard. Mater. 2016, 316, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Mano, J.; Tanaka, K.; Wang, S.; Zhang, M.; Deng, X.; Zhang, S. High level of reduced glutathione contributes to detoxification of lipid peroxide-derived reactive carbonyl species in transgenic Arabidopsis overexpressing glutathione reductase under aluminum stress. Physiol. Plant. 2017, 161, 211–223. [Google Scholar] [CrossRef] [PubMed]
- Marty, L.; Bausewein, D.; Müller, C.; Bangash, S.A.K.; Moseler, A.; Schwarzländer, M.; Müller-Schüssele, S.J.; Zechmann, B.; Riondet, C.; Balk, J.; et al. Arabidopsis glutathione reductase 2 is indispensable in plastids, while mitochondrial glutathione is safeguarded by additional reduction and transport systems. New Phytol. 2019, 224, 1569–1584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sumugat, M.R.; Donahue, J.L.; Cortes, D.F.; Stromberg, V.K.; Grene, R.; Shulaev, V.; Welbaum, G.E. Seed Development and Germination in anArabidopsis thalianaLine Antisense to Glutathione Reductase 2. J. New Seeds 2010, 11, 104–126. [Google Scholar] [CrossRef]
- Yu, X.; Pasternak, T.; Eiblmeier, M.; Ditengou, F.; Kochersperger, P.; Sun, J.; Wang, H.; Rennenberg, H.; Teale, W.; Paponov, I.; et al. Plastid-Localized Glutathione Reductase2-Regulated Glutathione Redox Status Is Essential for Arabidopsis Root Apical Meristem Maintenance. Plant Cell 2013, 25, 4451–4468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, M. Water stress-induced abscisic acid accumulation triggers the increased generation of reactive oxygen species and up-regulates the activities of antioxidant enzymes in maize leaves. J. Exp. Bot. 2002, 53, 2401–2410. [Google Scholar] [CrossRef] [Green Version]
- Romero-Puertas, M.C.; Corpas, F.J.; Sandalio, L.M.; Leterrier, M.; Rodriguez-Serrano, M.; Rio, L.A.; Palma, J.M. Glutathione reductase from pea leaves: Response to abiotic stress and characterization of the peroxisomal isozyme. New Phytol. 2006, 170, 43–52. [Google Scholar] [CrossRef]
- Kuźniak, E.; Skłodowska, M. Ascorbate, glutathione and related enzymes in chloroplasts of tomato leaves infected by Botrytis cinerea. Plant Sci. 2001, 160, 723–731. [Google Scholar] [CrossRef]
- Gill, S.S.; Anjum, N.A.; Hasanuzzaman, M.; Gill, R.; Trivedi, D.K.; Ahmad, I.; Pereira, E.; Tuteja, N. Glutathione and glutathione reductase: A boon in disguise for plant abiotic stress defense operations. Plant Physiol. Biochem. 2013, 70, 204–212. [Google Scholar] [CrossRef]
- Sudhakar, C.; Lakshmi, A.; Giridarakumar, S. Changes in the antioxidant enzyme efficacy in two high yielding genotypes of mulberry (Morus alba L.) under NaCl salinity. Plant Sci. 2001, 161, 613–619. [Google Scholar] [CrossRef]
- Bandeoğlu, E.; Eyidoğan, F.; Yücel, M.; Öktem, H.A. Antioxidant responses of shoots and roots of lentil to NaCl-salinity stress. Plant Growth Regul. 2004, 42, 69–77. [Google Scholar] [CrossRef]
- Wang, L.-J.; Li, S.-H. Salicylic acid-induced heat or cold tolerance in relation to Ca2+ homeostasis and antioxidant systems in young grape plants. Plant Sci. 2006, 170, 685–694. [Google Scholar] [CrossRef]
- Hossain, M.A.; Asada, K. Monodehydroascorbate reductase from cucumber is a flavin adenine dinucleotide enzyme. J. Biol. Chem. 1985, 260, 12920–12926. [Google Scholar] [CrossRef]
- Suman, S.; Bagal, D.; Jain, D.; Singh, R.; Singh, I.K.; Singh, A. Biotic stresses on plants: Reactive oxygen species generation and antioxidant mechanism, Chapter 14. In Frontiers in Plant-Soil Interactions; Aftab, T., Hakeem, K.R., Eds.; Academic Press: New York, NY, USA, 2021; pp. 381–411. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Anee, T.I.; Parvin, K.; Nahar, K.; Mahmud, J.A.; Fujita, M. Regulation of Ascorbate-Glutathione Pathway in Mitigating Oxidative Damage in Plants under Abiotic Stress. Antioxidants 2019, 8, 384. [Google Scholar] [CrossRef] [Green Version]
- Obara, K.; Sumi, K.; Fukuda, H. The use of multiple transcription starts causes the dual targeting of Arabidopsis putative monodehydroascorbate reductase to both mitochondria and chloroplasts. Plant Cell Physiol. 2002, 43, 697–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, N.; Hu, J. Defining the Plant Peroxisomal Proteome: From Arabidopsis to Rice. Front. Plant Sci. 2011, 2, 103. [Google Scholar] [CrossRef] [Green Version]
- Eastmond, P.J. Monodehyroascorbate reductase4 is Required for Seed Storage Oil Hydrolysis and Postgerminative Growth in Arabidopsis. Plant Cell 2007, 19, 1376–1387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eltayeb, A.E.; Kawano, N.; Badawi, G.H.; Kaminaka, H.; Sanekata, T.; Shibahara, T.; Inanaga, S.; Tanaka, K. Overexpression of monodehydroascorbate reductase in transgenic tobacco confers enhanced tolerance to ozone, salt and polyethylene glycol stresses. Planta 2007, 225, 1255–1264. [Google Scholar] [CrossRef]
- Sultana, S.; Khew, C.Y.; Morshed, M.; Namasivayam, P.; Napis, S.; Ho, C.-L. Overexpression of monodehydroascorbate reductase from a mangrove plant (AeMDHAR) confers salt tolerance on rice. J. Plant Physiol. 2011, 169, 311–318. [Google Scholar] [CrossRef]
- Rahantaniaina, M.-S.; Li, S.; Chatel-Innocenti, G.; Tuzet, A.; Issakidis-Bourguet, E.; Mhamdi, A.; Noctor, G. Cytosolic and Chloroplastic DHARs Cooperate in Oxidative Stress-Driven Activation of the Salicylic Acid Pathway. Plant Physiol. 2017, 174, 956–971. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Gallie, D.R. Dehydroascorbate Reductase Affects Leaf Growth, Development, and Function. Plant Physiol. 2006, 142, 775–787. [Google Scholar] [CrossRef] [Green Version]
- Ding, H.; Wang, B.; Han, Y.; Li, S. The pivotal function of dehydroascorbate reductase in glutathione homeostasis in plants. J. Exp. Bot. 2020, 71, 3405–3416. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Xiao, Y.; Chen, W.; Tang, K.; Zhang, L. Increased Vitamin C Content Accompanied by an Enhanced Recycling Pathway Confers Oxidative Stress Tolerance in Arabidopsis. J. Integr. Plant Biol. 2010, 52, 400–409. [Google Scholar] [CrossRef]
- Noshi, M.; Yamada, H.; Hatanaka, R.; Tanabe, N.; Tamoi, M.; Shigeoka, S. Arabidopsis dehydroascorbate reductase 1 and 2 modulate redox states of ascorbate-glutathione cycle in the cytosol in response to photooxidative stress. Biosci. Biotechnol. Biochem. 2017, 81, 523–533. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, S.; Tamaoki, M.; Shikano, T.; Nakajima, N.; Ogawa, D.; Ioki, M.; Aono, M.; Kubo, A.; Kamada, H.; Inoue, Y.; et al. Cytosolic Dehydroascorbate Reductase is Important for Ozone Tolerance in Arabidopsisthaliana. Plant Cell Physiol. 2006, 47, 304–308. [Google Scholar] [CrossRef]
- Noshi, M.; Hatanaka, R.; Tanabe, N.; Terai, Y.; Maruta, T.; Shigeoka, S. Redox regulation of ascorbate and glutathione by a chloroplastic dehydroascorbate reductase is required for high-light stress tolerance in Arabidopsis. Biosci. Biotechnol. Biochem. 2016, 80, 870–877. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Viveros, G.; Ferrera-Cerrato, R.; Alarcón, A. Short-Term Effects of Arsenate-Induced Toxicity on Growth, Chlorophyll and Carotenoid Contents, and Total Content of Phenolic Compounds of Azolla filiculoides. Water Air Soil Pollut. 2010, 217, 455–462. [Google Scholar] [CrossRef]
- Do, H.; Kim, I.-S.; Jeon, B.W.; Lee, C.W.; Park, A.K.; Wi, A.R.; Shin, S.C.; Park, H.; Kim, Y.-S.; Yoon, H.-S.; et al. Structural understanding of the recycling of oxidized ascorbate by dehydroascorbate reductase (OsDHAR) from Oryza sativa L. japonica. Sci. Rep. 2016, 6, 19498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dell’Aglio, E.; Mahmdi, A. What are the Roles for Dehydroascorbate Reductases and Glutathione in Sustaining Ascorbate Accumulation? Available online: https://plantae.org/what-are-the-roles-for-dehydroascorbate-reductases-and-glutathione-in-sustaining-ascorbate-accumulation (accessed on 10 October 2021).
- Chen, Z.; Young, T.E.; Ling, J.; Chang, S.-C.; Gallie, D.R. Increasing vitamin C content of plants through enhanced ascorbate recycling. Proc. Natl. Acad. Sci. USA 2003, 100, 3525–3530. [Google Scholar] [CrossRef] [Green Version]
- Smirnoff, N. Ascorbic acid metabolism and functions: A comparison of plants and mammals. Free Radic. Biol. Med. 2018, 122, 116–129. [Google Scholar] [CrossRef]
- Potters, G.; De Gara, L.; Asard, H.; Horemans, N. Ascorbate and glutathione: Guardians of the cell cycle, partners in crime? Plant Physiol. Biochem. 2002, 40, 537–548. [Google Scholar] [CrossRef]
- Ushimaru, T.; Nakagawa, T.; Fujioka, Y.; Daicho, K.; Naito, M.; Yamauchi, Y.; Nonaka, H.; Amako, K.; Yamawaki, K.; Murata, N. Transgenic Arabidopsis plants expressing the rice dehydroascorbate reductase gene are resistant to salt stress. J. Plant Physiol. 2006, 163, 1179–1184. [Google Scholar] [CrossRef]
- Yin, L.; Wang, S.; Eltayeb, A.E.; Uddin, I.; Yamamoto, Y.; Tsuji, W.; Takeuchi, Y.; Tanaka, K. Overexpression of dehydroascorbate reductase, but not monodehydroascorbate reductase, confers tolerance to aluminum stress in transgenic tobacco. Planta 2009, 231, 609–621. [Google Scholar] [CrossRef] [PubMed]
- Eltelib, H.A.; Badejo, A.A.; Fujikawa, Y.; Esaka, M. Gene expression of monodehydroascorbate reductase and dehydroascorbate reductase during fruit ripening and in response to environmental stresses in acerola (Malpighia glabra). J. Plant Physiol. 2011, 168, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-S.; Kim, I.-S.; Bae, M.-J.; Choe, Y.-H.; Kim, Y.-H.; Park, H.-M.; Kang, H.-G.; Yoon, H.-S. Homologous expression of cytosolic dehydroascorbate reductase increases grain yield and biomass under paddy field conditions in transgenic rice (Oryza sativa L. japonica). Planta 2013, 237, 1613–1625. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Gallie, D.R. Dehydroascorbate Reductase Affects Non-photochemical Quenching and Photosynthetic Performance. J. Biol. Chem. 2008, 283, 21347–21361. [Google Scholar] [CrossRef] [Green Version]
- Dowdle, J.; Ishikawa, T.; Gatzek, S.; Rolinski, S.; Smirnoff, N. Two genes in Arabidopsis thaliana encoding GDP-l-galactose phosphorylase are required for ascorbate biosynthesis and seedling viability. Plant J. 2007, 52, 673–689. [Google Scholar] [CrossRef]
- Terai, Y.; Ueno, H.; Ogawa, T.; Sawa, Y.; Miyagi, A.; Kawai-Yamada, M.; Ishikawa, T.; Maruta, T. Dehydroascorbate Reductases and Glutathione Set a Threshold for High-Light–Induced Ascorbate Accumulation. Plant Physiol. 2020, 183, 112–122. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Chaouch, S.; Mhamdi, A.; Queval, G.; Zechmann, B.; Noctor, G. Functional Analysis of Arabidopsis Mutants Points to Novel Roles for Glutathione in Coupling H2O2 to Activation of Salicylic Acid Accumulation and Signaling. Antioxid. Redox Signal. 2013, 18, 2106–2121. [Google Scholar] [CrossRef] [Green Version]
- Niak, E.B.K. Ascorbate and ascorbate-dependent enzymes in detached tomato leaves under conditions modulating the ascorbate pool. Acta Physiol. Plant. 2004, 26, 1–6. [Google Scholar] [CrossRef]
- Liebler, D.C.; Kling, D.S.; Reed, D.J. Antioxidant protection of phospholipid bilayers by alpha-tocopherol. Control of α-tocopherol status and lipid peroxidation by ascorbic acid and glutathione. J. Biol. Chem. 1986, 261, 12114–12119. [Google Scholar] [CrossRef]
- Szarka, A.; Tomasskovics, B.; Bánhegyi, G. The ascorbate-glutathione-α-tocopherol triad in abiotic stress response. Int. J. Mol. Sci. 2012, 13, 4458–4483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mullineaux, P.M.; Rausch, T. Glutathione, photosynthesis and the redox regulation of stress-responsive gene expression. Photosynth. Res. 2005, 86, 459–474. [Google Scholar] [CrossRef] [PubMed]
- Roychoudhury, A.; Pradhan, S.; Chaudhuri, B.; Das, K. Phytoremediation of toxic metals and the involvement of Brassica species. In Phytotechnologies: Remediation of Environmental Contaminants; Anjum, N.A., Pereira, M.E., Ahmad, I., Duarte, A.C., Umar, S., Khan, N.A., Eds.; CRC Press: Boca Raton, FL, USA, 2012; pp. 219–251. [Google Scholar]
- RoyChoudhury, A.; Das, K.; Ghosh, S.; Mukherjee, R.N.; Banerjee, R. Transgenic plants: Benefits and controversies. J. Bot. Soc. Bengal 2012, 66, 29–35. [Google Scholar]
- Liang, X.; Zhang, L.; Natarajan, S.K.; Becker, D.F. Proline Mechanisms of Stress Survival. Antioxid. Redox Signal. 2013, 19, 998–1011. [Google Scholar] [CrossRef] [Green Version]
- Szabados, L.; Savouré, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Trelstad, R.L.; Lawley, K.R.; Holmes, L.B. Nonenzymatic hydroxylations of proline and lysine by reduced oxygen derivatives. Nature 1981, 289, 310–312. [Google Scholar] [CrossRef]
- Kaul, S.; Sharma, S.S.; Mehta, I.K. Free radical scavenging potential of L-proline: Evidence from in vitro assays. Amino Acids 2008, 34, 315–320. [Google Scholar] [CrossRef]
- Sharma, S.S.; Dietz, K.-J. The relationship between metal toxicity and cellular redox imbalance. Trends Plant Sci. 2009, 14, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Floyd, R.A.; Nagy, I.Z. Formation of long-lived hydroxyl free radical adducts of proline and hydroxyproline in a fenton reaction. Biochim. Biophys. Acta-Protein Struct. Mol. Enzym. 1984, 790, 94–97. [Google Scholar] [CrossRef]
- Saradhi, P.P.; Mohanty, P. Involvement of proline in protecting thylakoid membranes against free radical-induced photodamage. J. Photochem. Photobiol. B Biol. 1997, 38, 253–257. [Google Scholar] [CrossRef]
- Matysik, J.; Alia Bhalu, B.; Mohanty, P. Molecular mechanisms of quenching of reactive oxygen species by proline under stress in plants. Curr. Sci. 2002, 82, 525–532. [Google Scholar]
- Sharma, S.S.; Schat, H.; Vooijs, R. In vitro alleviation of heavy metal-induced enzyme inhibition by proline. Phytochemistry 1998, 49, 1531–1535. [Google Scholar] [CrossRef]
- Farago, M.; Mullen, W. Plants which accumulate metals. Part IV. A possible copper-proline complex from the roots of armeria maritima. Inorg. Chim. Acta 1979, 32, L93–L94. [Google Scholar] [CrossRef]
- Chen, C.; Dickman, M.B. From The Cover: Proline suppresses apoptosis in the fungal pathogen Colletotrichum trifolii. Proc. Natl. Acad. Sci. USA 2005, 102, 3459–3464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoque, A.; Okuma, E.; Banu, M.N.A.; Nakamura, Y.; Shimoishi, Y.; Murata, Y. Exogenous proline mitigates the detrimental effects of salt stress more than exogenous betaine by increasing antioxidant enzyme activities. J. Plant Physiol. 2007, 164, 553–561. [Google Scholar] [CrossRef]
- Xu, J.; Yin, H.; Li, X. Protective effects of proline against cadmium toxicity in micropropagated hyperaccumulator, Solanum nigrum L. Plant Cell Rep. 2008, 28, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Hoque, A.; Banu, M.N.A.; Nakamura, Y.; Shimoishi, Y.; Murata, Y. Proline and glycinebetaine enhance antioxidant defense and methylglyoxal detoxification systems and reduce NaCl-induced damage in cultured tobacco cells. J. Plant Physiol. 2008, 165, 813–824. [Google Scholar] [CrossRef]
- Islam, M.M.; Hoque, A.; Okuma, E.; Jannat, R.; Banu, M.N.A.; Jahan, S.; Nakamura, Y.; Murata, Y. Proline and Glycinebetaine Confer Cadmium Tolerance on Tobacco Bright Yellow-2 Cells by Increasing Ascorbate-Glutathione Cycle Enzyme Activities. Biosci. Biotechnol. Biochem. 2009, 73, 2320–2323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diplock, T.; Machlin, L.J.; Packer, L.; Pryor, W.A. Vitamin E: Biochemistry and Health Implications; Annals of the New York Academy of Sciences: New York, NY, USA, 1989; pp. 372–378. [Google Scholar]
- Kamal-Eldin, A.; Appelqvist, L.-Å. The chemistry and antioxidant properties of tocopherols and tocotrienols. Lipids 1996, 31, 671–701. [Google Scholar] [CrossRef]
- Kiffin, R.; Bandyopadhyay, U.; Cuervo, A.M. Oxidative Stress and Autophagy. Antioxid. Redox Signal. 2006, 8, 152–162. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K.; Prenzel, U.; Douce, R.; Joyard, J. Localization of prenylquinones in the envelope of spinach chloroplasts. Biochim. Biophys. Acta-Biomembr. 1981, 641, 99–105. [Google Scholar] [CrossRef]
- Soll, J.; Schultz, G.; Joyard, J.; Douce, R.; Block, M.A. Localization and synthesis of prenylquinones in isolated outer and inner envelope membranes from spinach chloroplasts. Arch. Biochem. Biophys. 1985, 238, 290–299. [Google Scholar] [CrossRef] [Green Version]
- Kanwischer, M.; Porfirova, S.; Bergmuüller, E.; Doörmann, P. Alterations in Tocopherol Cyclase Activity in Transgenic and Mutant Plants of Arabidopsis Affect Tocopherol Content, Tocopherol Composition, and Oxidative Stress. Plant Physiol. 2005, 137, 713–723. [Google Scholar] [CrossRef] [Green Version]
- Shinozaki, K. A novel cis-acting element in an Arabidopsis gene is involved in responsiveness to drought, low-temperature, or high-salt stress. Plant Cell 1994, 6, 251–264. [Google Scholar] [CrossRef] [Green Version]
- Munné-Bosch, S.; Schwarz, K.; Alegre, L. Enhanced Formation of α-Tocopherol and Highly Oxidized Abietane Diterpenes in Water-Stressed Rosemary Plants. Plant Physiol. 1999, 121, 1047–1052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.; Liu, X.; Li, X.; Chen, S.; Jin, Z.; Liu, G. Overexpression of VTE1 from Arabidopsis resulting in high vitamin E accumu-lation and salt stress tolerance increase in tobacco plant. Chin. J. Appl. Environ. Biol. 2006, 12, 468–471. (In Chinese) [Google Scholar]
- Bafeel, S.O.; Ibrahim, M.M. Antioxidants and accumulation of α-tocopherol induce chilling tolerance in Medicago sativa. Int. J. Agric. Biol. 2008, 10, 593–598. [Google Scholar]
- Li, Y.; Zhou, Y.; Wang, Z.; Sun, X.; Tang, K. Engineering tocopherol biosynthetic pathway in Arabidopsis leaves and its effect on antioxidant metabolism. Plant Sci. 2010, 178, 312–320. [Google Scholar] [CrossRef]
- Fritsche, S.; Wang, X.; Jung, C. Recent Advances in our Understanding of Tocopherol Biosynthesis in Plants: An Overview of Key Genes, Functions, and Breeding of Vitamin E Improved Crops. Antioxidants 2017, 6, 99. [Google Scholar] [CrossRef] [Green Version]
- DellaPenna, D.; Last, R. Progress in the dissection and manipulation of plant vitamin E biosynthesis. Physiol. Plant. 2006, 126, 356–368. [Google Scholar] [CrossRef]
- Ivanov, B.N.; Khorobrykh, S. Participation of Photosynthetic Electron Transport in Production and Scavenging of Reactive Oxygen Species. Antioxid. Redox Signal. 2003, 5, 43–53. [Google Scholar] [CrossRef]
- Matringe, M.; Ksas, B.; Rey, P.; Havaux, M. Tocotrienols, the Unsaturated Forms of Vitamin E, Can Function as Antioxidants and Lipid Protectors in Tobacco Leaves. Plant Physiol. 2008, 147, 764–778. [Google Scholar] [CrossRef] [Green Version]
- Mène-Saffrané, L.; DellaPenna, D. Biosynthesis, regulation and functions of tocochromanols in plants. Plant Physiol. Biochem. 2009, 48, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Kagan, V.E.; Fabisiak, J.P.; Quinn, P.J. Coenzyme Q and vitamin E need each other as antioxidants. Protoplasma 2000, 214, 11–18. [Google Scholar] [CrossRef]
- Fryer, M.J. The antioxidant effects of thylakoid Vitamin E (alpha-tocopherol). Plant Cell Environ. 1992, 15, 381–392. [Google Scholar] [CrossRef]
- Igamberdiev, A.U.; Seregelyes, C.; Manac, N.; Hill, R.D. NADH-dependent metabolism of nitric oxide in alfalfa root cultures expressing barley hemoglobin. Planta 2004, 219, 95–102. [Google Scholar] [CrossRef]
- Fukuzawa, K.; Tokumura, A.; Ouchi, S.; Tsukatani, H. Antioxidant activities of tocopherols on Fe2+-ascorbate-induced lipid peroxidation in lecithin liposomes. Lipids 1982, 17, 511–513. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Reactive Species and Antioxidants. Redox Biology Is a Fundamental Theme of Aerobic Life. Plant Physiol. 2006, 141, 312–322. [Google Scholar] [CrossRef] [Green Version]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine, 5th ed.; Oxford University Press: Oxford, UK, 2015; pp. 381–382. [Google Scholar]
- Maeda, H.; Song, W.; Sage, T.L.; DellaPenna, D. Tocopherols Play a Crucial Role in Low-Temperature Adaptation and Phloem Loading in Arabidopsis. Plant Cell 2006, 18, 2710–2732. [Google Scholar] [CrossRef] [Green Version]
- Azzi, A. Molecular mechanism of α-tocopherol action. Free Radic. Biol. Med. 2007, 43, 16–21. [Google Scholar] [CrossRef]
- Hyun, T.K.; Kumar, K.; Rao, K.P.; Sinha, A.K.; Roitsch, T. Role of α-tocopherol in cellular signaling: α-tocopherol inhibits stress-induced mitogen-activated protein kinase activation. Plant Biotechnol. Rep. 2010, 5, 19–25. [Google Scholar] [CrossRef]
- Young, A.J. The photoprotective role of carotenoids in higher plants. Physiol. Plant. 1991, 83, 702–708. [Google Scholar] [CrossRef]
- Havaux, M. Carotenoid oxidation products as stress signals in plants. Plant J. 2013, 79, 597–606. [Google Scholar] [CrossRef]
- Lohr, M. Carotenoids. In The Chlamydomonas Sourcebook, 2nd ed.; Harris, E.H., Stern, D.B., Witman, G.B., Eds.; Academic Press: Cambridge, MA, USA, 2009; pp. 799–817. [Google Scholar] [CrossRef]
- Siefermann-Harms, D. The light-harvesting and protective functions of carotenoids in photosynthetic membranes. Physiol. Plant. 1987, 69, 561–568. [Google Scholar] [CrossRef]
- Cogdell, R.J.; Gardiner, A.T. Functions of carotenoids in photosynthesis. Meth. Enzymol. 1993, 214, 185–193. [Google Scholar] [CrossRef]
- Mallik, N.; Rai, L.C. Physiological responses of non-vascular plants to heavy metals, Chapter 5. In Metals Physiology and Bio-chemistry of Metal Toxicity and Tolerance in Plants; Prasad, M.N., Strzatka, K., Eds.; Springer: Dordrecht, The Netherlands, 2002; pp. 111–147. [Google Scholar]
- Johnson, M.; Havaux, M.; Triantaphylides, C.; Ksas, B.; Pascal, A.; Robert, B.; Davison, P.A.; Ruban, A.V.; Horton, P. Elevated Zeaxanthin Bound to Oligomeric LHCII Enhances the Resistance of Arabidopsis to Photooxidative Stress by a Lipid-protective, Antioxidant Mechanism. J. Biol. Chem. 2007, 282, 22605–22618. [Google Scholar] [CrossRef] [Green Version]
- Triantaphylidès, C.; Havaux, M. Singlet oxygen in plants: Production, detoxification and signaling. Trends Plant Sci. 2009, 14, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Jahns, P.; Holzwarth, A.R. The role of the xanthophyll cycle and of lutein in photoprotection of photosystem II. Biochim. Biophys. Acta 2012, 1817, 182–193. [Google Scholar] [CrossRef] [Green Version]
- DellaPenna, D.; Pogson, B.J. Vitamin synthesis in plants: Tocopherols and Carotenoids. Annu. Rev. Plant Biol. 2006, 57, 711–738. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Vallabhaneni, R.; Yu, J.; Rocheford, T.; Wurtzel, E.T. The Maize Phytoene Synthase Gene Family: Overlapping Roles for Carotenogenesis in Endosperm, Photomorphogenesis, and Thermal Stress Tolerance. Plant Physiol. 2008, 147, 1334–1346. [Google Scholar] [CrossRef] [Green Version]
- Gomathi, R.; Rakkiyapan, P. Comparative lipid peroxidation, leaf membrane thermostability, and antioxidant system in four sugarcane genotypes differing in salt tolerance. Int. J. Plant Physiol. Biochem. 2011, 3, 67–74. [Google Scholar] [CrossRef]
- Grace, S.C.; Logan, B. Energy dissipation and radical scavenging by the plant phenylpropanoid pathway. Philos. Trans. R. Soc. B Biol. Sci. 2000, 355, 1499–1510. [Google Scholar] [CrossRef] [Green Version]
- Michalak, A. Phenolic compounds and their antioxidant activity in plants growing under heavy metal stress. Pol. J. Environ. Stud. 2006, 15, 523–530. [Google Scholar]
- Fini, A.; Brunetti, C.; Di Ferdinando, M.; Ferrini, F.; Tattini, M. Stress-induced flavonoid biosynthesis and the antioxidant machinery of plants. Plant Signal. Behav. 2011, 6, 709–711. [Google Scholar] [CrossRef]
- Agati, G.; Azzarello, E.; Pollastri, S.; Tattini, M. Flavonoids as antioxidants in plants: Location and functional significance. Plant Sci. 2012, 196, 67–76. [Google Scholar] [CrossRef]
- Sakihama, Y.; Cohen, M.F.; Grace, S.C.; Yamasaki, H. Plant phenolic antioxidant and prooxidant activities: Phenolics-induced oxidative damage mediated by metals in plants. Toxicology 2002, 177, 67–80. [Google Scholar] [CrossRef]
- Rice-Evans, C.; Miller, N.; Paganga, G. Antioxidant properties of phenolic compounds. Trends Plant Sci. 1997, 2, 152–159. [Google Scholar] [CrossRef]
- Finnegan, P.M.; Chen, W. Arsenic Toxicity: The Effects on Plant Metabolism. Front. Physiol. 2012, 3, 182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waszczak, C.; Akter, S.; Jacques, S.; Huang, J.; Messens, J.; Van Breusegem, F. Oxidative post-translational modifications of cysteine residues in plant signal transduction. J. Exp. Bot. 2015, 66, 2923–2934. [Google Scholar] [CrossRef] [Green Version]
- Waszczak, C.; Carmody, M.; Kangasjärvi, J. Reactive Oxygen Species in Plant Signaling. Annu. Rev. Plant Biol. 2018, 69, 209–236. [Google Scholar] [CrossRef] [Green Version]
- Wu, F.; Chi, Y.; Jiang, Z.; Xu, Y.; Xie, L.; Huang, F.; Wan, D.; Ni, J.; Yuan, F.; Wu, X.; et al. Hydrogen peroxide sensor HPCA1 is an LRR receptor kinase in Arabidopsis. Nature 2020, 578, 577–581. [Google Scholar] [CrossRef] [PubMed]
- Yalcinkaya, T.; Uzilday, B.; Ozgur, R.; Turkan, I.; Mano, J. Lipid peroxidation-derived reactive carbonyl species (RCS): Their interaction with ROS and cellular redox during environmental stresses. Environ. Exp. Bot. 2019, 165, 139–149. [Google Scholar] [CrossRef]
- Suzuki, Y.J.; Carini, M.; Butterfield, D.A. Protein carbonylation. Antioxid. Redox Signal. 2010, 12, 323–325. [Google Scholar] [CrossRef] [PubMed]
- Ciacka, K.; Tymiński, M.; Gniazdowska, A.; Krasuska, U. Carbonylation of proteins—An element of plant ageing. Planta 2020, 252, 12. [Google Scholar] [CrossRef]
- Cui, F.; Brosche, M.; Shapiguzov, A.; He, X.-Q.; Vainonen, J.P.; Leppälä, J.; Trotta, A.; Kangasjärvi, S.; Salojärvi, J.; Kangasjärvi, J.; et al. Interaction of methyl viologen-induced chloroplast and mitochondrial signalling in Arabidopsis. Free. Radic. Biol. Med. 2019, 134, 555–566. [Google Scholar] [CrossRef]
- Exposito-Rodriguez, M.; Laissue, P.P.; Yvon-Durocher, G.; Smirnoff, N.; Mullineaux, P.M. Photosynthesis-dependent H2O2 transfer from chloroplasts to nuclei provides a high-light signalling mechanism. Nat. Commun. 2017, 8, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trebak, M.; Ginnan, R.; Singer, H.A.; Jourd’Heuil, D. Interplay Between Calcium and Reactive Oxygen/Nitrogen Species: An Essential Paradigm for Vascular Smooth Muscle Signaling. Antioxid. Redox Signal. 2010, 12, 657–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hempel, N.; Trebak, M. Crosstalk between calcium and reactive oxygen species signaling in cancer. Cell Calcium 2017, 63, 70–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, L.; Liao, W. Hydrogen Peroxide Signaling in Plant Development and Abiotic Responses: Crosstalk with Nitric Oxide and Calcium. Front. Plant Sci. 2016, 7, 230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farnese, F.D.S.; Menezes-Silva, P.E.; Gusman, G.S.; Oliveira, J. When Bad Guys Become Good Ones: The Key Role of Reactive Oxygen Species and Nitric Oxide in the Plant Responses to Abiotic Stress. Front. Plant Sci. 2016, 7, 471. [Google Scholar] [CrossRef] [Green Version]
- Lindermayr, C.; Durner, J. Interplay of Reactive Oxygen Species and Nitric Oxide: Nitric Oxide Coordinates Reactive Oxygen Species Homeostasis. Plant Physiol. 2015, 167, 1209–1210. [Google Scholar] [CrossRef] [Green Version]
- Del Río, L.A. ROS and RNS in plant physiology: An overview. J. Exp. Bot. 2015, 66, 2827–2837. [Google Scholar] [CrossRef] [Green Version]
- Piterková, J.; Luhová, L.; Navrátilová, B.; Sedlářová, M.; Petřivalský, M. Early and long-term responses of cucumber cells to high cadmium concentration are modulated by nitric oxide and reactive oxygen species. Acta Physiol. Plant. 2015, 37, 19. [Google Scholar] [CrossRef]
- Delledonne, M.; Zeier, J.; Marocco, A.; Lamb, C. Signal interactions between nitric oxide and reactive oxygen intermediates in the plant hypersensitive disease resistance response. Proc. Natl. Acad. Sci. USA 2001, 98, 13454–13459. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Loake, G.J.; Chu, C. Cross-talk of nitric oxide and reactive oxygen species in plant programed cell death. Front. Plant Sci. 2013, 4, 314. [Google Scholar] [CrossRef] [Green Version]
- Kohli, S.K.; Khanna, K.; Bhardwaj, R.; Allah, E.F.A.; Ahmad, P.; Corpas, F.J. Assessment of Subcellular ROS and NO Metabolism in Higher Plants: Multifunctional Signaling Molecules. Antioxidants 2019, 8, 641. [Google Scholar] [CrossRef] [Green Version]
- Kankofer, M. Antioxidative Defence Mechanisms Against Reactive Oxygen Species in Bovine Retained and Not-Retained Placenta: Activity of Glutathione Peroxidase, Glutathione Transferase, Catalase and Superoxide Dismutase. Placenta 2001, 22, 466–472. [Google Scholar] [CrossRef] [PubMed]
- Vardharajula, S.; Ali, S.Z.; Grover, M.; Reddy, G.; Bandi, V. Drought-tolerant plant growth promotingBacillusspp.: Effect on growth, osmolytes, and antioxidant status of maize under drought stress. J. Plant Interact. 2011, 6, 1–14. [Google Scholar] [CrossRef]
- Batool, T.; Ali, S.; Seleiman, M.F.; Naveed, N.H.; Ali, A.; Ahmed, K.; Abid, M.; Rizwan, M.; Shahid, M.R.; Alotaibi, M.; et al. Plant growth promoting rhizobacteria alleviates drought stress in potato in response to suppressive oxidative stress and antioxidant enzymes activities. Sci. Rep. 2020, 10, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Gan, Y.; Xu, B. Application of Plant-Growth-Promoting Fungi Trichoderma longibrachiatum T6 Enhances Tolerance of Wheat to Salt Stress through Improvement of Antioxidative Defense System and Gene Expression. Front. Plant Sci. 2016, 7, 1405. [Google Scholar] [CrossRef] [Green Version]
- Borriss, R.; Wu, H.; Gao, X. Secondary metabolites of the plant growth promoting model rhizobacterium Bacillus velezensis FZB42 are involved in direct suppression of plant pathogens and in stimulation of plant-induced systemic resistance. In Metabolites of Plant Growth Promoting Rhizomicroorganisms: Discovery and Applications, 1st ed.; Singh, H.B., Keswani, C., Reddy, M.S., Sansinenea, E., García-Estrada, C., Eds.; Springer: Singapore, 2019; pp. 147–168. [Google Scholar]
- Shrivastava, P.; Kumar, R. Soil salinity: A serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi J. Biol. Sci. 2014, 22, 123–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno-Galván, A.; Romero-Perdomo, F.A.; Estrada-Bonilla, G.; Meneses, C.H.S.G.; Bonilla, R.R. Dry-Caribbean Bacillus spp. Strains Ameliorate Drought Stress in Maize by a Strain-Specific Antioxidant Response Modulation. Microorganisms 2020, 8, 826. [Google Scholar] [CrossRef] [PubMed]
- Carlson, R.; Tugizimana, F.; Steenkamp, P.A.; Dubery, I.A.; Hassen, A.I.; Labuschagne, N. Rhizobacteria-induced systemic tolerance against drought stress in Sorghum bicolor (L.) Moench. Microbiol. Res. 2019, 232, 126388. [Google Scholar] [CrossRef] [PubMed]
- Lucas, J.A.; García-Cristobal, J.; Bonilla, A.; Ramos, B.; Gutierrez-Mañero, J. Beneficial rhizobacteria from rice rhizosphere confers high protection against biotic and abiotic stress inducing systemic resistance in rice seedlings. Plant Physiol. Biochem. 2014, 82, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Naseem, H.; Bano, A. Role of plant growth-promoting rhizobacteria and their exopolysaccharide in drought tolerance of maize. J. Plant Interact. 2014, 9, 689–701. [Google Scholar] [CrossRef] [Green Version]
- Ngumbi, E.; Kloepper, J. Bacterial-mediated drought tolerance: Current and future prospects. Appl. Soil Ecol. 2016, 105, 109–125. [Google Scholar] [CrossRef]
- Yang, J.; Kloepper, J.W.; Ryu, C.-M. Rhizosphere bacteria help plants tolerate abiotic stress. Trends Plant Sci. 2009, 14, 1–4. [Google Scholar] [CrossRef]
- Harish, S.; Kavino, M.; Kumar, N.; Saravanakumar, D.; Soorianathasundaram, K.; Samiyappan, R. Biohardening with Plant Growth Promoting Rhizosphere and Endophytic bacteria induces systemic resistance against Banana bunchy top virus. Appl. Soil Ecol. 2008, 39, 187–200. [Google Scholar] [CrossRef]
- Walters, D.R.; Fountaine, J.M. Practical application of induced resistance to plant diseases: An appraisal of effectiveness under field conditions. J. Agric. Sci. 2009, 147, 523–535. [Google Scholar] [CrossRef]
- Wang, C.; Guo, Y.; Wang, C.; Liu, H.; Niu, D.; Wang, Y.; Guo, J. Enhancement of tomato (Lycopersicon esculentum) tolerance to drought stress by plant-growth-promoting rhizobacterium (PGPR) Bacillus cereus AR156. J. Agric. Biotechnol. 2012, 20, 1097–1105. (In Chinese) [Google Scholar]
- Gallé, A.; Haldimann, P.; Feller, U. Photosynthetic performance and water relations in young pubescent oak (Quercus pubescens) trees during drought stress and recovery. New Phytol. 2007, 174, 799–810. [Google Scholar] [CrossRef]
- Banik, P.; Zeng, W.; Tai, H.; Bizimungu, B.; Tanino, K. Effects of drought acclimation on drought stress resistance in potato (Solanum tuberosum L.) genotypes. Environ. Exp. Bot. 2016, 126, 76–89. [Google Scholar] [CrossRef]
- Vurukonda, S.S.K.P.; Vardharajula, S.; Shrivastava, M.; SkZ, A. Enhancement of drought stress tolerance in crops by plant growth promoting rhizobacteria. Microbiol. Res. 2016, 184, 13–24. [Google Scholar] [CrossRef]
- Ansary, M.H.; Rahmani, H.A.; Ardakani, M.R.; Paknejad, F.; Habibi, D.; Mafakheri, S. Effect of Pseudomonas fluorescens on proline and phytohormonal status of maize (Zea mays L.) under water deficit stress. Ann. Biol. Res. 2012, 3, 1054–1062. [Google Scholar]
- Armada, E.; Roldan, A.; Azcon, R. Differential Activity of Autochthonous Bacteria in Controlling Drought Stress in Native Lavandula and Salvia Plants Species Under Drought Conditions in Natural Arid Soil. Microb. Ecol. 2013, 67, 410–420. [Google Scholar] [CrossRef]
- Shintu, P.V.; Jayaram, K.M. Phosphate solubilising bacteria (Bacillus polymyxa)—An effective approach to mitigate drought in tomato (Lycopersicon esculentum Mill). Trop. Plant Res. 2015, 2, 17–22. [Google Scholar]
- Paul, M.J.; Primavesi, L.F.; Jhurreea, D.; Zhang, Y. Trehalose Metabolism and Signaling. Annu. Rev. Plant Biol. 2008, 59, 417–441. [Google Scholar] [CrossRef] [Green Version]
- Mastouri, F.; Björkman, T.; Harman, G.E. Seed Treatment with Trichoderma harzianum Alleviates Biotic, Abiotic, and Physiological Stresses in Germinating Seeds and Seedlings. Phytopathology 2010, 100, 1213–1221. [Google Scholar] [CrossRef] [Green Version]
- Yogendra, S.G.; Singh, U.S.; Sharma, A.K. Enhance activity of stress related enzymes in rice (Oryza sativa L.) induced by plant growth promoting fungi under drought stress. Afr. J. Agric. Res. 2014, 9, 1430–1434. [Google Scholar] [CrossRef] [Green Version]
- Zlatev, Z.S.; Lidon, F.C.; Ramalho, J.; Yordanov, I.T. Comparison of resistance to drought of three bean cultivars. Biol. Plant. 2006, 50, 389–394. [Google Scholar] [CrossRef]
- Chang-Quan, W.; Rui-Chang, L. Enhancement of superoxide dismutase activity in the leaves of white clover (Trifolium repens L.) in response to polyethylene glycol-induced water stress. Acta Physiol. Plant. 2008, 30, 841–847. [Google Scholar] [CrossRef]
- Mafakheri, A.; Siosemardeh, A.; Bahramnejad, B.; Struik, P.C.; Sohrabi, Y. Effect of drought stress on yield, proline and chlo-rophyll contents in three chickpea cultivars. Aust. J. Crop Sci. 2010, 4, 580–585. [Google Scholar]
- Kukreja, S.; Nandwal, A.S.; Kumar, N.; Sharma, S.K.; Sharma, S.K.; Unvi, V.; Sharma, P.K. Plant water status, H2O2 scavenging enzymes, ethylene evolution and membrane integrity of Cicer arietinum roots as affected by salinity. Biol. Plant. 2005, 49, 305–308. [Google Scholar] [CrossRef]
- Eyidogan, F.; Oz, M.T. Effect of salinity on antioxidant responses of chickpea seedlings. Acta Physiol. Plant. 2007, 29, 485–493. [Google Scholar] [CrossRef]
- Gapińska, M.; Skłodowska, M.; Gabara, B. Effect of short- and long-term salinity on the activities of antioxidative enzymes and lipid peroxidation in tomato roots. Acta Physiol. Plant. 2007, 30, 11–18. [Google Scholar] [CrossRef]
- Takshak, S.; Agrawal, S.B. Effect of ultraviolet-B radiation on biomass production, lipid peroxidation, reactive oxygen species, and antioxidants in withania somnifera. Biol. Plant. 2014, 58, 328–334. [Google Scholar] [CrossRef]
- Zhang, F.; Zhang, H.; Xia, Y.; Wang, G.; Xu, L.; Shen, Z. Exogenous application of salicylic acid alleviates cadmium toxicity and reduces hydrogen peroxide accumulation in root apoplasts of Phaseolus aureus and Vicia sativa. Plant Cell Rep. 2011, 30, 1475–1483. [Google Scholar] [CrossRef] [PubMed]
- Sandalio, L.; Dalurzo, H.; Gómez, M.; Romero-Puertas, M.; del Río, L. Cadmium-induced changes in the growth and oxidative metabolism of pea plants. J. Exp. Bot. 2001, 52, 2115–2126. [Google Scholar] [CrossRef]
- Li, F.-T.; Qi, J.-M.; Zhang, G.-Y.; Lin, L.-H.; Fang, P.-P.; Tao, A.-F.; Xu, J.-T. Effect of Cadmium Stress on the Growth, Antioxidative Enzymes and Lipid Peroxidation in Two Kenaf (Hibiscus cannabinus L.) Plant Seedlings. J. Integr. Agric. 2013, 12, 610–620. [Google Scholar] [CrossRef]
- Gao, S.; Ouyang, C.; Wang, S.; Xu, Y.; Tang, L.; Chen, F. Effects of salt stress on growth, antioxidant enzyme and phenylalanine ammonia-lyase activities in Jatropha curcas L. seedlings. Plant Soil Environ. 2008, 54, 374–381. [Google Scholar] [CrossRef] [Green Version]
- Badawi, G.H.; Kawano, N.; Yamauchi, Y.; Shimada, E.; Sasaki, R.; Kubo, A.; Tanaka, K. Over-expression of ascorbate peroxidase in tobacco chloroplasts enhances the tolerance to salt stress and water deficit. Physiol. Plant. 2004, 121, 231–238. [Google Scholar] [CrossRef]
- Kim, S.Y.; Lim, J.-H.; Park, M.R.; Kim, Y.J.; Park, T.I.; Seo, Y.W.; Choi, K.G.; Yun, S.J. Enhanced antioxidant enzymes are associated with reduced hydrogen peroxide in barley roots under saline stress. J. Biochem. Mol. Biol. 2005, 38, 218–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Ying, Y.; Chen, J.; Wang, X. Transgenic Arabidopsis overexpressing Mn-SOD enhanced salt-tolerance. Plant Sci. 2004, 167, 671–677. [Google Scholar] [CrossRef]
- Rao, M.V.; Paliyath, G.; Ormrod, D.P. Ultraviolet-B- and Ozone-Induced Biochemical Changes in Antioxidant Enzymes of Arabidopsis thaliana. Plant Physiol. 1996, 110, 125–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malar, S.; Shivendra Vikram, S.; Favas, P.J.; Perumal, V. Lead heavy metal toxicity induced changes on growth and antioxidative enzymes level in water hyacinths [Eichhornia crassipes (Mart.)]. Bot. Stud. 2016, 55, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roychoudhury, A.; Ghosh, S. Physiological and biochemical responses of mungbean (Vigna radiata L. Wilczek) to varying concentrations of cadmium chloride or sodium chloride. Unique J. Pharm. Biol. Sci. 2013, 1, 11–21. [Google Scholar]
- Kandziora-Ciupa, M.; Ciepał, R.; Nadgórska-Socha, A.; Barczyk, G. A comparative study of heavy metal accumulation and antioxidant responses in Vaccinium myrtillus L. leaves in polluted and non-polluted areas. Environ. Sci. Pollut. Res. 2013, 20, 4920–4932. [Google Scholar] [CrossRef] [Green Version]
- Aghaei, K.; Ehsanpour, A.A.; Komatsu, S. Potato Responds to Salt Stress by Increased Activity of Antioxidant Enzymes. J. Integr. Plant Biol. 2009, 51, 1095–1103. [Google Scholar] [CrossRef]
- Sharma, P.; Dubey, R.S. Drought Induces Oxidative Stress and Enhances the Activities of Antioxidant Enzymes in Growing Rice Seedlings. Plant Growth Regul. 2005, 46, 209–221. [Google Scholar] [CrossRef]
- Basu, S.; Roychoudhury, A.; Saha, P.P.; Sengupta, D.N. Differential antioxidative responses of indica rice cultivars to drought stress. Plant Growth Regul. 2009, 60, 51–59. [Google Scholar] [CrossRef]
- Roychoudhury, A.; Basu, S.; Sengupta, D.N. Antioxidants and stress-related metabolites in the seedlings of two indica rice varieties exposed to cadmium chloride toxicity. Acta Physiol. Plant. 2011, 34, 835–847. [Google Scholar] [CrossRef]
- Roychoudhury, A.; Basu, S.; Sarkar, S.N.; Sengupta, D.N. Comparative physiological and molecular responses of a common aromatic indica rice cultivar to high salinity with non-aromatic indica rice cultivars. Plant Cell Rep. 2008, 27, 1395–1410. [Google Scholar] [CrossRef]
- Vaidyanathan, H.; Sivakumar, P.; Chakrabarty, R.; Thomas, G. Scavenging of reactive oxygen species in NaCl-stressed rice (Oryza sativa L.)—differential response in salt-tolerant and sensitive varieties. Plant Sci. 2003, 165, 1411–1418. [Google Scholar] [CrossRef]
- Mishra, P.; Bhoomika, K.; Dubey, R.S. Differential responses of antioxidative defense system to prolonged salinity stress in salt-tolerant and salt-sensitive Indica rice (Oryza sativa L.) seedlings. Protoplasma 2011, 250, 3–19. [Google Scholar] [CrossRef]
- Çelik, Ö.; Çakır, B.C.; Atak, Ç. Identification of the antioxidant defense genes which may provide enhanced salt tolerance in Oryza sativa L. Physiol. Mol. Biol. Plants 2018, 25, 85–99. [Google Scholar] [CrossRef]
- Chutipaijit, S.; Cha-Um, S.; Sompornpailin, K. Differential accumulations of proline and flavonoids in indica rice varieties against salinity. Pak. J. Bot. 2009, 41, 2497–2506. [Google Scholar]
- Xu, S.; Li, J.; Zhang, X.; Wei, H.; Cui, L. Effects of heat acclimation pretreatment on changes of membrane lipid peroxidation, antioxidant metabolites, and ultrastructure of chloroplasts in two cool-season turfgrass species under heat stress. Environ. Exp. Bot. 2006, 56, 274–285. [Google Scholar] [CrossRef]
- Chakraborty, U.; Pradhan, D. High temperature-induced oxidative stress in Lens culinaris, role of antioxidants and amelioration of stress by chemical pre-treatments. J. Plant Interact. 2011, 6, 43–52. [Google Scholar] [CrossRef]
- Sairam, R.; Deshmukh, P.; Saxena, D. Role of Antioxidant Systems in Wheat Genotypes Tolerance to Water Stress. Biol. Plant. 1998, 41, 387–394. [Google Scholar] [CrossRef]
- Qayyum, A.; Al Ayoubi, S.; Sher, A.; Bibi, Y.; Ahmad, S.; Shen, Z.; Jenks, M.A. Improvement in drought tolerance in bread wheat is related to an improvement in osmolyte production, antioxidant enzyme activities, and gaseous exchange. Saudi J. Biol. Sci. 2021, 28, 5238–5249. [Google Scholar] [CrossRef]
- Khanna-Chopra, R.; Chauhan, S. Wheat cultivars differing in heat tolerance show a differential response to oxidative stress during monocarpic senescence under high temperature stress. Protoplasma 2015, 252, 1241–1251. [Google Scholar] [CrossRef]
- Almeselmani, M.; Deshmukh, P.S.; Sairam, R.K. High temperature stress tolerance in wheat genotypes: Role of antioxidant defence enzymes. Acta Agron. Hung. 2009, 57, 1–14. [Google Scholar] [CrossRef]
- Kaur, H.; Sirhindi, G.; Bhardwaj, R.; Alyemeni, M.N.; Siddique, K.; Ahmad, P. 28-homobrassinolide regulates antioxidant enzyme activities and gene expression in response to salt- and temperature-induced oxidative stress in Brassica juncea. Sci. Rep. 2018, 8, 8735. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Gupta, D.; Nayyar, H. Comparative response of maize and rice genotypes to heat stress: Status of oxidative stress and antioxidants. Acta Physiol. Plant. 2011, 34, 75–86. [Google Scholar] [CrossRef]
- Shabankareh, H.G.; Khorasaninejad, S.; Soltanloo, H.; Shariati, V. Physiological response and secondary metabolites of three lavender genotypes under water deficit. Sci. Rep. 2021, 11, 1–22. [Google Scholar] [CrossRef]
- Munné-Bosch, S.; Alegre, L. Changes in carotenoids, tocopherols and diterpenes during drought and recovery, and the biological significance of chlorophyll loss in Rosmarinus officinalis plants. Planta 2000, 210, 925–931. [Google Scholar] [CrossRef]
- Kim, T.Y.; Ku, H.; Lee, S.-Y. Crop Enhancement of Cucumber Plants under Heat Stress by Shungite Carbon. Int. J. Mol. Sci. 2020, 21, 4858. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Sullivan, J.; Garrett, W.M.; Caperna, T.J.; Natarajan, S. Impact of solar Ultraviolet-B on the proteome in soybean lines differing in flavonoid contents. Phytochemistry 2008, 69, 38–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Schematic model showing the reactive oxygen species (ROS) generation in plants as well as the ascorbate-glutathione (ascorbic acid/AA-GSH) and the guaiacol peroxidase (GPX) cycles. AA is oxidised by ROS and converted into monodehydroascorbate (MDHA). A set of three enzymes, including FAD-dependent monodehydroascorbate reductase (MDHAR), GSH-dependent dehydroascorbate reductase (DHAR) and glutathione reductase (GR), catalyse the recycling of ascorbate. Superoxide dismutase (SOD) converts O2●− into H2O2. Ascorbate peroxidase (APX) (1), catalase (CAT) (2), and GPX (3) act as the main H2O2 detoxifying enzymes. AA and GSH are antioxidants. Abbreviations: ascorbate peroxidase (APX), oxidised glutathione (GSH), reduced glutathione (GSSG).
Figure 1.
Schematic model showing the reactive oxygen species (ROS) generation in plants as well as the ascorbate-glutathione (ascorbic acid/AA-GSH) and the guaiacol peroxidase (GPX) cycles. AA is oxidised by ROS and converted into monodehydroascorbate (MDHA). A set of three enzymes, including FAD-dependent monodehydroascorbate reductase (MDHAR), GSH-dependent dehydroascorbate reductase (DHAR) and glutathione reductase (GR), catalyse the recycling of ascorbate. Superoxide dismutase (SOD) converts O2●− into H2O2. Ascorbate peroxidase (APX) (1), catalase (CAT) (2), and GPX (3) act as the main H2O2 detoxifying enzymes. AA and GSH are antioxidants. Abbreviations: ascorbate peroxidase (APX), oxidised glutathione (GSH), reduced glutathione (GSSG).


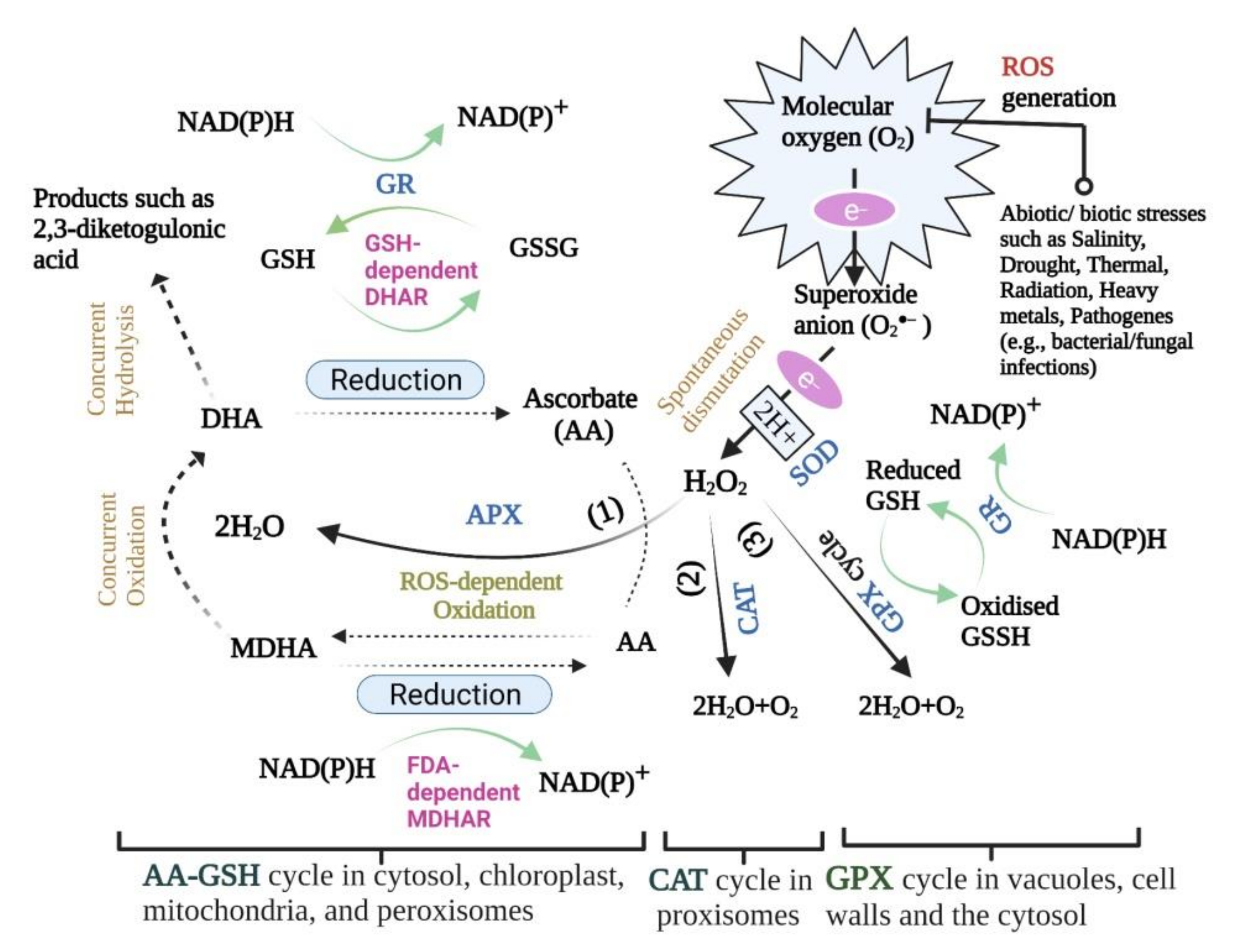{kind=link}
Table 1.
CAT cDNA isolated and characterised from different plant species.
| Plant Species | Common Name | Genomic DNA or Protein | Subcellular Localisation and Site of Detection in Plant | References |
|---|---|---|---|---|
| Brassica napus | Rapeseed | CAT1–CAT14 | Peroxisomes (CAT2–CAT3, CAT6–CAT8, CAT11–CAT14), cytoskeleton (CAT1), cytoplasm (CAT4), mitochondrion (CAT9), and chloroplast (CAT5, CAT10) in root, leaf, stem, and silique samples | [89] |
| Helianthus annus | Sunflower | CAT1–CAT8 | Peroxisomes in cotyledons and roots | [90,91,92] |
| Gossypium hirsutum | Cotton | CAT1–CAT7 | Peroxisomes in leaves | [78,93] |
| Pisum sativum | Pea | CAT1–CAT5 | Peroxisomes in leaves and the whole fruit | [94,95] |
| Cucumis sativus | Cucumber | CAT1–CAT4 | Peroxisomes in roots, stem, leaves, flowers, and fruits | [79] |
| Oryza sativa L. | Rice | OsCATA–OsCATD | Cytosol (OsCATA–OsCATC), peroxisomes (OsCATB, OsCATC), and plasma membrane (OsCATD) in immature seeds and rice seedlings | [80,96,97] |
| Nicotiana plumbaginofolia | Tex-Mex tobacco | CAT1–CAT3 | Peroxisomes in leaves | [84] |
| Arabidopsis thaliana | Arabidopsis | CAT1–CAT3 | Peroxisomes in bolts, leaves, siliques (CAT1, CAT2), in pollen and seeds (CAT1), stems and roots (CAT3) | [81,98,99,100,101] |
| Cucurpita pepo | Pumpkin | CAT1–CAT3 | Glyoxysomes in seeds and early seedlings (CAT1), mature leaves, stem (CAT2), green cotyledons and green hypocotyls (CAT2, CAT3), and roots (CAT3) | [82] |
| Glycine max | Soybean | CAT1–CAT3 | Glyoxysomes in developing kernels (CAT1), green leaves (CAT2, CAT3), roots (CAT3), and epicotyl of the developing seedlings (CAT3) | [102] |
| Zea mays | Maize | CAT1–CAT3 | In scutella, milky endosperm of immature kernels, leaves and epicotyls (CAT1); Post-germinative scutella, with lower levels in leaves, epicotyls and growing kernels (CAT2); Epicotyls and, to a lesser extent, in leaves and scutella (CAT3) | [83,103,104,105,106,107] |
| Capsicum annuum | Hot pepper | CAT1–CAT3 | Peroxisomes in leaf and stem | [108] |
| Nicotiana tabacum | Tobacco | CAT1–CAT3 | Peroxisomes in leaves | [109,110] |
| Ricinus cammunis | Castor bean | CAT1, CAT2 | Glyoxysomes, peroxisomes in hypocotyls and roots (CAT2), in endosperms and cotyledons (CAT1) | [88,111] |
| Hurdeom vulgare | Barley | CAT1, CAT2 | Peroxisomes in whole endosperms, in isolated aleurones and in developing seeds (CAT1), in etiolated seedling shoots and leaf blades (CAT2) | [85] |
| Triticum aestivum | Wheat | CAT1, CAT2 | Peroxisomes in leaves | [112] |
| Solanum tuberosum | Potato | CAT1, CAT2 | Peroxisomes in leaves | [113,114] |
| Lycopersicum sculentum | Tomato | CAT1, CAT2 | Peroxisomes in leaves | [86,115,116] |
| Nicotiana sylvestris | Woodland tobacco | CAT1, CAT4 | Peroxisomes in leaves | [117] |
| Vigna radiata | Mung bean | CAT1 | Glyoxysomes, leaf peroxisomes, and nonspecialised microbodies (etiolated or nongreen tissues) | [118] |
| Secale cereale | Rye | CAT1 | Peroxisomes in leaves | [119] |
| Ipomoea batatas | Sweet potato | CAT1 | Peroxisomes in leaves | [87,120] |
Table 2.
Enzymatic/non-enzymatic antioxidants reported in selected plant species under stress conditions.
Table 2.
Enzymatic/non-enzymatic antioxidants reported in selected plant species under stress conditions.
| Plant Species | Stress Type | Antioxidant Reported + (Increase)/−(Decrease) | Reference |
|---|---|---|---|
| Phaseolus vulgaris L. | Drought | +SOD, +APX in leaves | [316] |
| Trifolium repens L. | Drought | +SOD in leaves | [317] |
| Cicer arietinum L. cv. ILC482 | Drought | +Proline in leaves | [318] |
| Cicer arietinum L. cv. CSG-8962 | Salinity | +SOD, +CAT, + Peroxidase (POX), +APX, +GR, and –AA in roots | [319] |
| Cicer arietinum L. cv. Gökçe | Salinity | +SOD in roots and leaves, +APX and +GR in leaves, and +CAT in roots, –CAT in leaves | [320] |
| Lycopersicon esculentum Mill. cv. Perkoz | Salinity | +SOD, +Glutathione S-transferase (GST), and +Glutathione peroxidase (GSH-PX) in roots | [321] |
| Withania somnifera L. | UV-B radiation | +CAT, +GR, +POX, +Polyphenol oxidase (PPO), and +SOD in roots > leaves | [322] |
| Vicia sativa L. | Cd stress | +SOD, +APX, and +CAT in roots | [323] |
| Pisum sativum L. | Cd stress | –SOD, –CAT, –GPX, and +Lipid peroxidation (LPO) in leaves | [324] |
| Hibiscus cannabinus L. cvs. Fuhong 991 and ZM 412 | Cd stress | +GR in leaves (Fuhong 991), +SOD, +CAT, and +POX in roots (Fuhong 991 and ZM 412), +Lipid peroxidation (LPO) in roots (ZM 412) | [325] |
| Jatropha curcas L. | Salinity | +SOD, +CAT, +POX in the cotyledons, hypocotyls and radicles | [326] |
| Nicotiana tabacum L. line Chl-APX5 | Salinity and drought | +APX in chloroplast | [327] |
| Nicotiana tabacum L. | Salinity | +SOD, +CAT, +APX, +POX, and +GR in roots | [328] |
| Nicotiana tabacum L. | Salt, O3 and polyethylene glycol (PEG) stresses | +MDHAR in leaves | [186] |
| Arabidopsis thaliana L. | Salinity | +DHAR in leaves | [201] |
| Arabidopsis thaliana L. | Salinity | +SOD, +CAT, and +POX in leaves | [329] |
| Arabidopsis thaliana L. | UV-B/Ozone (O3) radiation | +SOD, +APX, +GPX, +POX (UV-B and O3), and +GR (O3) in leaves | [330] |
| Eichhornia crassipes (water hyacinth) | Pb stress | +APX, +POX, +SOD, +CAT in leaf and root tissues | [331] |
| Vigna radiata L. Wilczek | CdCl2 stress | +MDHA +POX, +CAT, and +Proline in roots and leaves | [332] |
| Vaccinium myrtillus L. | Heavy metal stress (Cd, Pb, and Zn) | +GSH, +Non-protein thiols, +Proline, and +GPX in leaves | [333] |
| Solanum tuberosum L. | Salt stress | +APX, +CAT and +GR in roots and shoots | [334] |
| Oryza sativa L. | Drought | +SOD, +APX, +MDHAR, +DHAR, +GR, and –CAT in roots and leaves | [335] |
| Oryza sativa L. cvs. IR-29 and Pusa Basmati | Drought | +Flavonoids, +Phenolics, –CAT, –SOD, and +GPX in leaves | [336] |
| Oryza sativa L. vars. IR-29 and Nonabokra | CdCl2/NaCl stress | + Anthocyanin, + Carotenoids, +GPX, +APX, +Proline, +Polyamines (spermidine and spermine) (IR-29/Nonabokra), –Cysteine and –CAT (IR-29), +Cysteine and +CAT (Nonabokra) in leaves | [337,338] |
| Oryza sativa L. cv. Pokkali | Salt stress | +CAT, +AA, and +GSH in leaves | [339] |
| Oryza sativa L. cvs. CSR-27 and Osmancık-97 | Salt stress | +SOD, +APX, +GPX, +CAT, +MDHAR, +DHAR, +GR, and +Proline in roots and leaves | [340,341] |
| Oryza sativa L. vars. Jasmine (KDML105) and Sangyod (SY) | Salt stress | +Flavonoids, and +Proline in shoots | [342] |
| Turfgrass species | Heat stress | +GSH, and +AA in leaves | [343] |
| Lens culinaris Medik. | Heat stress | +CAT, +APX, +SOD, and +LPO in leaves | [344] |
| Triticum aestivum L. cv. C306 | Drought | +SOD, +APX, +CAT, and +AA in leaves | [345] |
| Triticum aestivum L. cv. Chakwal-50 | Drought | +Proline,+CAT, +SOD, +POX, and +MDHAR in leaves | [346] |
| Triticum aestivum L. cv. Hindi62 | Heat stress | +GSH, +SOD, +CAT, +APX, +GR, and +MDHAR in leaves | [347] |
| Triticum aestivum L.cv. C 306. | Heat stress | +SOD, +APX, +CAT, +GR and +POX in leaves | [348] |
| Brassica juncea L. | High temperature and salt stress | +SOD, +CAT, +APX, +GR, +DHAR, and +MDHAR in seedlings | [349] |
| Zea mays L. | Heat stress | +CAT, +APX, +GR, +AA, +GSH, and +Proline in leaves | [350] |
| Lavandula angustifolia cv. Hidcote, Lavandula angustifolia cv. Munstead, and Lavandula stricta | Drought | +Proline, +CAT, +APX, +POX, +SOD, +MDHA, +Flavonoids, and +Phenols in leaves | [351] |
| Rosmarinus officinalis L. | Drought | +Carotenoids, and +α-tocopherol in leaves | [352] |
| Cucumis sativus L. | Heat stress | +SOD, +CAT, and +POX in leaves | [353] |
| Glycine max L. cv. Clark | UV-B radiation | –APX, –CAT, +SOD, +DHA, and +Flavonoid in leaves | [354] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zandi, P.; Schnug, E. Reactive Oxygen Species, Antioxidant Responses and Implications from a Microbial Modulation Perspective. Biology 2022, 11, 155. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11020155
AMA Style
Zandi P, Schnug E. Reactive Oxygen Species, Antioxidant Responses and Implications from a Microbial Modulation Perspective. Biology. 2022; 11(2):155. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11020155
Chicago/Turabian StyleZandi, Peiman, and Ewald Schnug. 2022. "Reactive Oxygen Species, Antioxidant Responses and Implications from a Microbial Modulation Perspective" Biology 11, no. 2: 155. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11020155
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.