
Sulfated Polysaccharides from Chaetoceros muelleri: Macromolecular Characteristics and Bioactive Properties
 , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Microalga Cultivation and Polysaccharide Extraction
2.2. Chemical Analysis
2.3. Attenuated Total Reflectance-FourierTransform Infrared (ATR-FTIR) Spectroscopy
2.4. Microstructure and Elemental Composition
2.5. Macromolecular Characteristics
2.6. Cell Line and Culture Conditions
2.7. Cytotoxicity
2.8. Antioxidant Activity
2.9. Glycemic Index (GI)
2.10. Statistical Analysis
3. Results
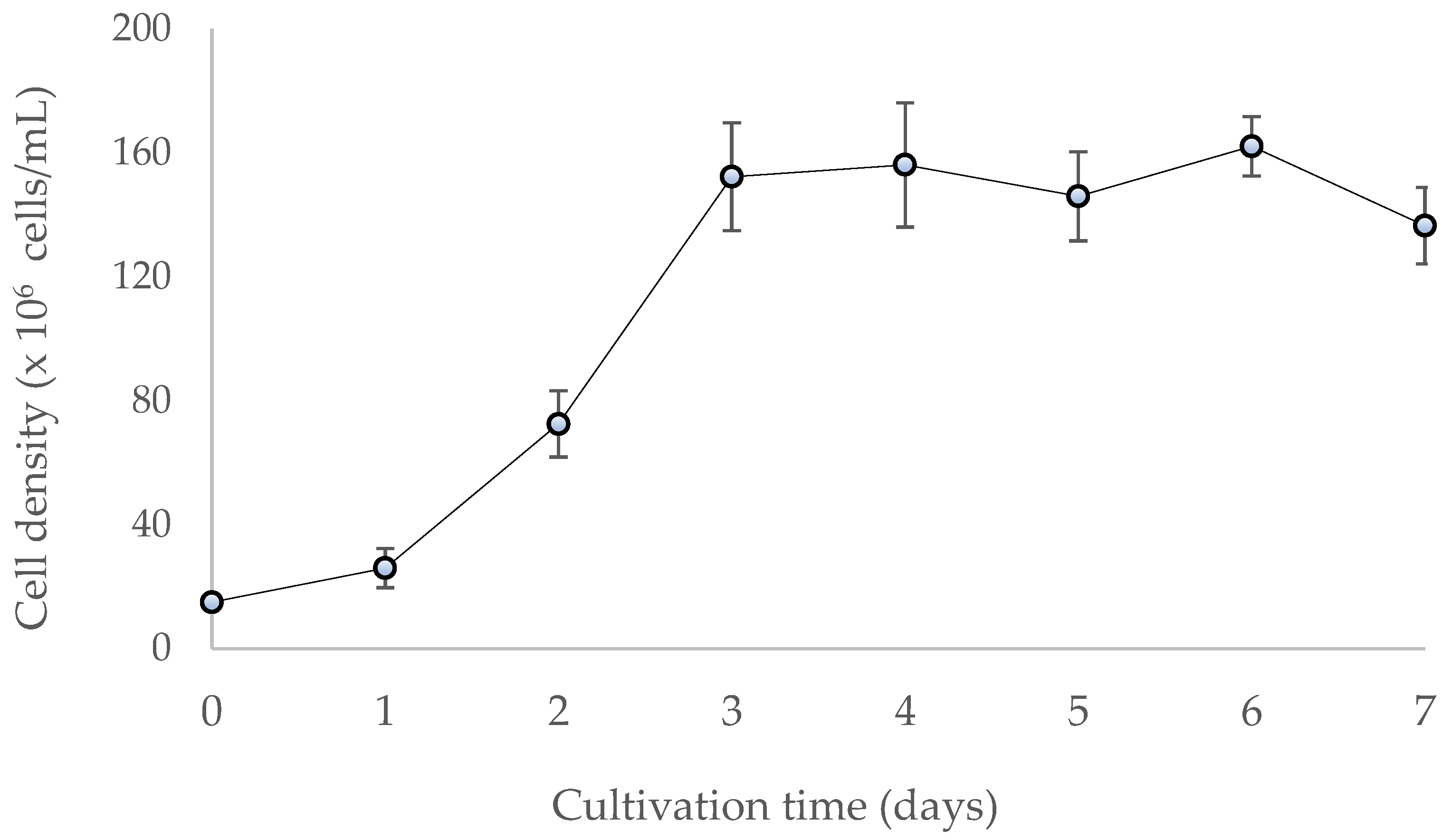
3.1. Cell Culture and Polysaccharide Yield
3.2. Yield and Chemical Analysis of the CMSP
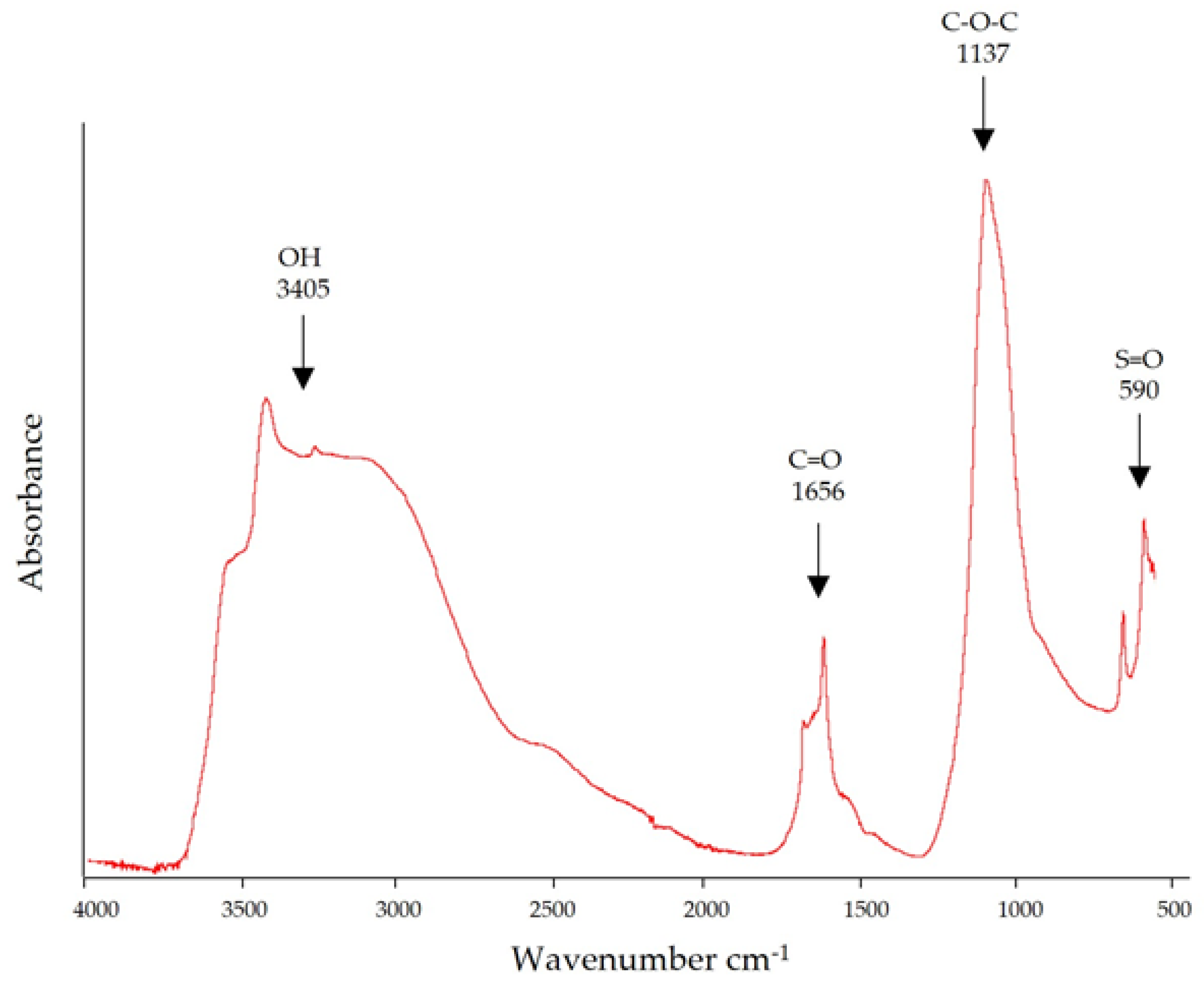
3.3. Fourier Transform Infrared Spectroscopy
3.4. Microstructure and Elemental Composition
3.5. Macromolecular Characteristics
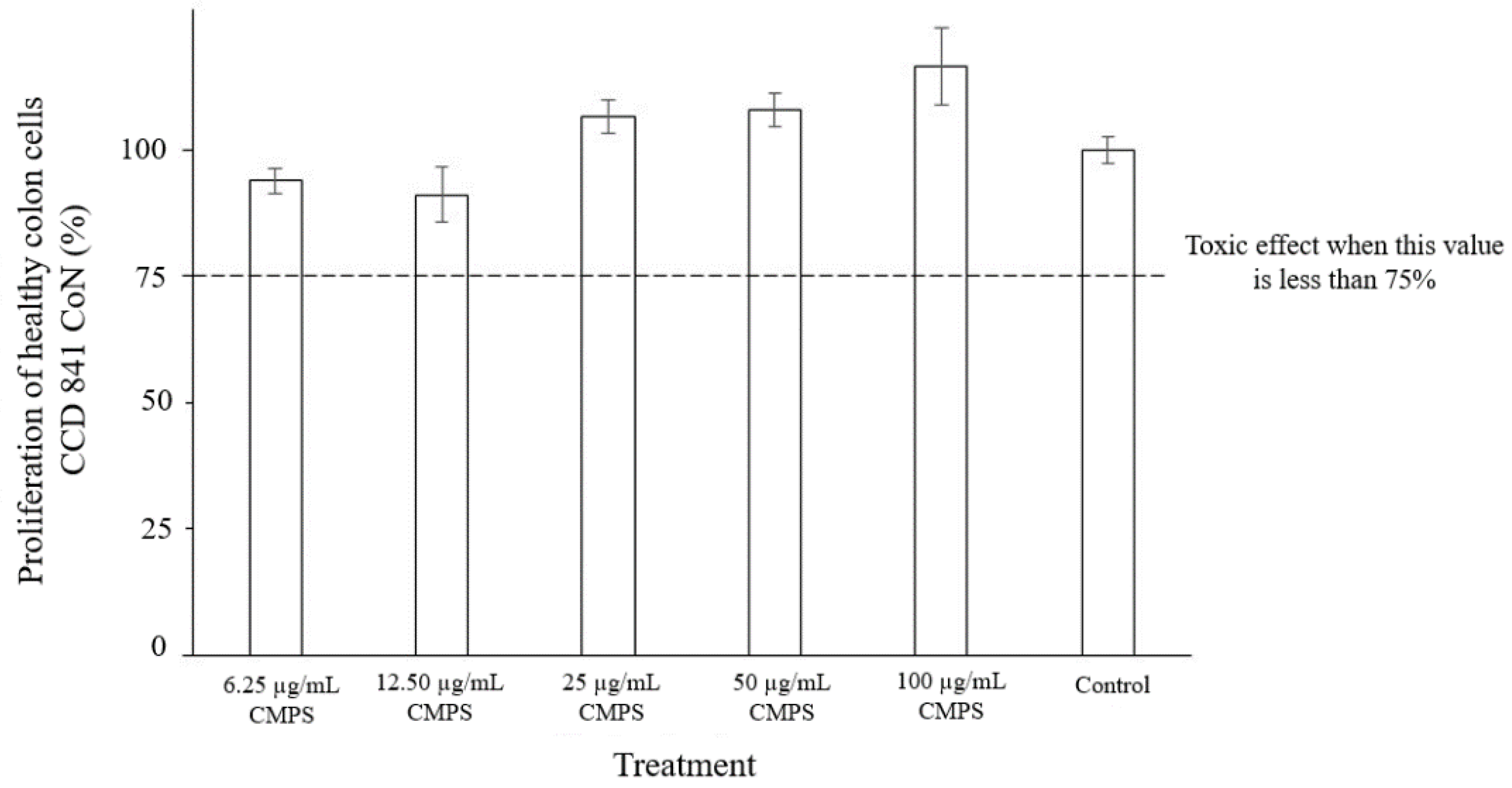
3.6. Cytotoxicity of the CMSP
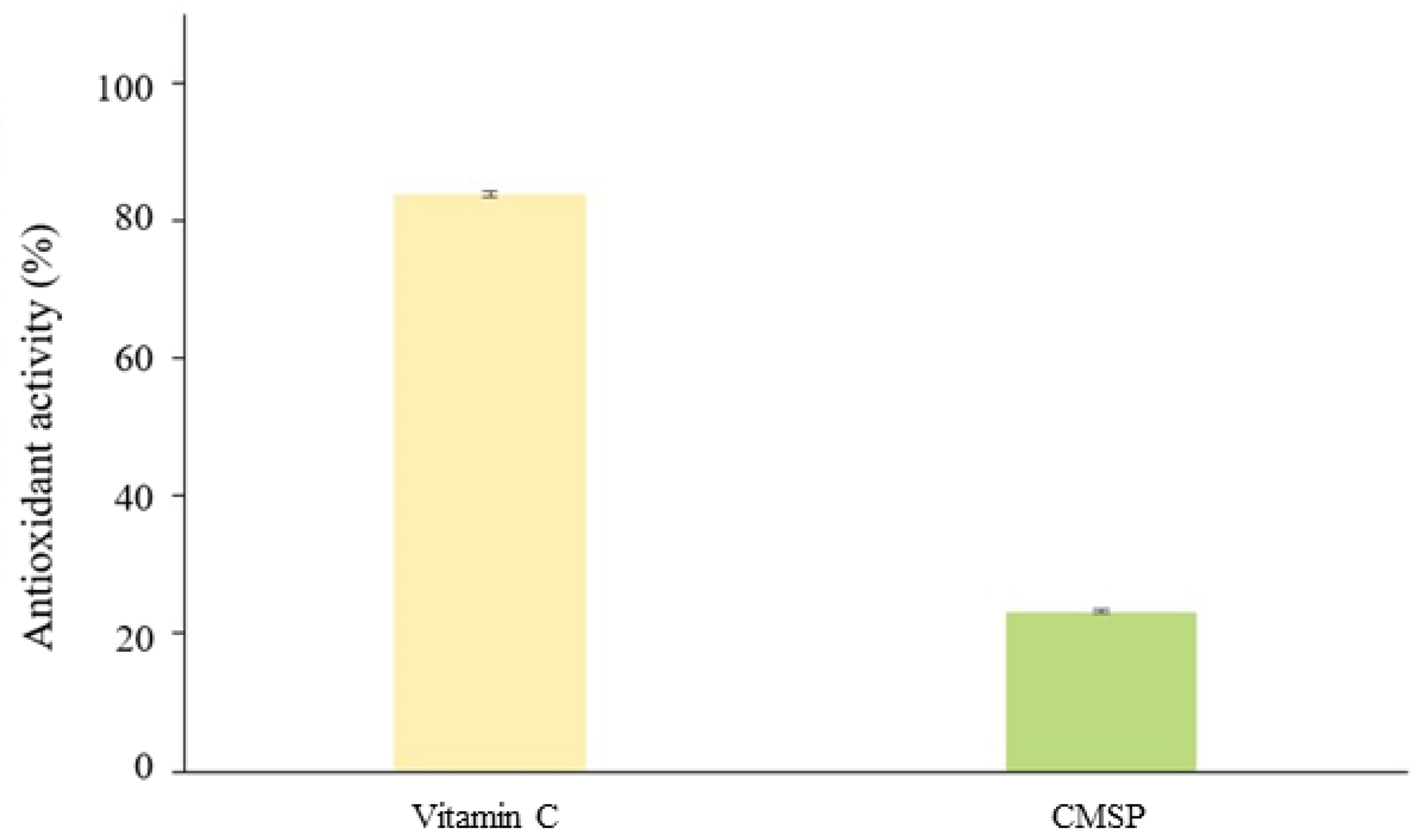
3.7. Antioxidant Activity of the CMSP
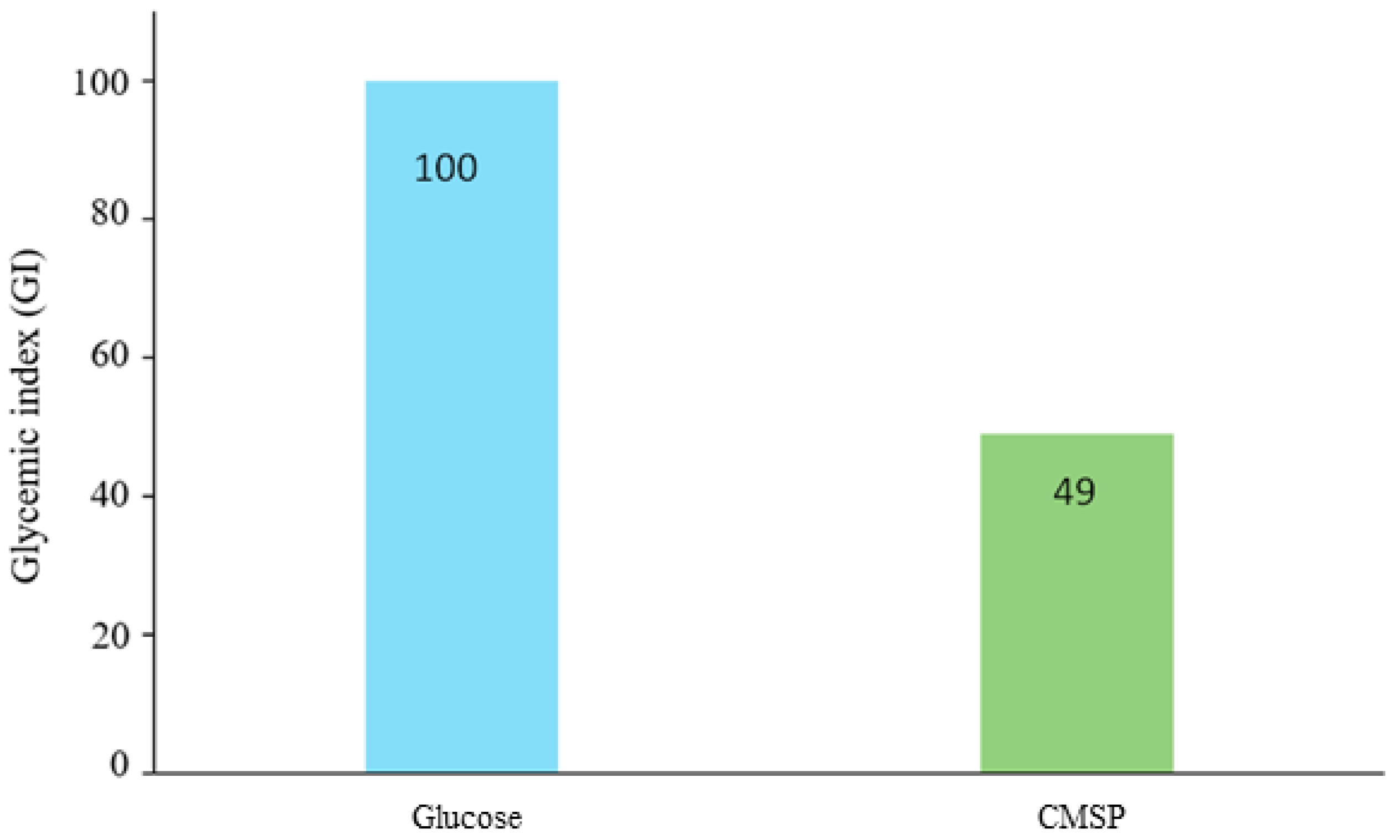
3.8. Glycemic Index (GI)
4. Discussion
4.1. CMSP Yield and Characteristics
4.2. Cytotoxicity, Antioxidant Activity, and Glycemic Index of the CMSP
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brown, E.M.; Allsopp, P.J.; Magee, P.J.; Gill, C.I.; Nitecki, S.; Strain, C.R.; McSorley, E.M. Seaweed and Human Health. Nutr. Rev. 2014, 72, 205–216. [Google Scholar] [CrossRef] [PubMed]
- De Jesus Raposo, M.; de Morais, A.; de Morais, R. Marine Polysaccharides from Algae with Potential Biomedical Applications. Mar. Drugs 2015, 13, 2967–3028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamed, I.; Özogul, F.; Özogul, Y.; Regenstein, J.M. Marine Bioactive Compounds and Their Health Benefits: A Review. Compr. Rev. Food Sci. Food Saf. 2015, 14, 446–465. [Google Scholar] [CrossRef]
- Jiang, J.-L.; Zhang, W.-Z.; Ni, W.-X.; Shao, J.-W. Insight on Structure-Property Relationships of Carrageenan from Marine Red Algal: A Review. Carbohydr. Polym. 2021, 257, 117642. [Google Scholar] [CrossRef]
- Le Costaouëc, T.; Unamunzaga, C.; Mantecon, L.; Helbert, W. New Structural Insights into the Cell-Wall Polysaccharide of the Diatom Phaeodactylum Tricornutum. Algal Res. 2017, 26, 172–179. [Google Scholar] [CrossRef]
- Raposo, M.; De Morais, R.; Bernardo de Morais, A. Bioactivity and Applications of Sulphated Polysaccharides from Marine Microalgae. Mar. Drugs 2013, 11, 233–252. [Google Scholar] [CrossRef] [Green Version]
- Malibari, R.; Sayegh, F.; Elazzazy, A.M.; Baeshen, M.N.; Dourou, M.; Aggelis, G. Reuse of Shrimp Farm Wastewater as Growth Medium for Marine Microalgae Isolated from Red Sea—Jeddah. J. Clean. Prod. 2018, 198, 160–169. [Google Scholar] [CrossRef]
- Rahmadi, A.; Mulyani, Y.; Lewaru, M.W. Effect of Salinity Difference on Lipid Content from Chaetoceros Muelleri on Continuous Reactors. Adv. J. Grad. Res. 2019, 7, 3–10. [Google Scholar] [CrossRef] [Green Version]
- Geresh, S.; Mamontov, A.; Weinstein, J. Sulfation of Extracellular Polysaccharides of Red Microalgae: Preparation, Characterization and Properties. J. Biochem. Biophys. Methods 2002, 50, 179–187. [Google Scholar] [CrossRef]
- Ryther, J.H.; Guillard, R.R.L. Studies of marine planktonic diatoms: III. Some effects of temperature on respiration of five species. Can. J. Microbiol. 1962, 8, 447–453. [Google Scholar] [CrossRef]
- Lai, H.-L.; Yang, L.-C.; Lin, P.-T.; Lai, S.-Y.; Wang, M.-Y. Phagocytosis Activity of Three Sulfated Polysaccharides Purified from a Marine Diatom Cultured in a Semi-Continuous System. Int. J. Biol. Macromol. 2020, 155, 951–960. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Horwitz, W.; Latimer, G.W. Official Methods of Analysis of AOAC International, 18th ed.; AOAC International: Gaithersburg, MD, USA, 2007; ISBN 978-0-935584-78-3. [Google Scholar]
- Fimbres-Olivarria, D.; Carvajal-Millan, E.; Lopez-Elias, J.A.; Martinez-Robinson, K.G.; Miranda-Baeza, A.; Martinez-Cordova, L.R.; Enriquez-Ocaña, F.; Valdez-Holguin, J.E. Chemical Characterization and Antioxidant Activity of Sulfated Polysaccharides from Navicula sp. Food Hydrocoll. 2018, 75, 229–236. [Google Scholar] [CrossRef]
- Rochas, C.; Lahaye, M.; Yaphe, W. Sulfate Content of Carrageenan and Agar Determined by Infrared Spectroscopy. Bot. Mar. 1986, 29, 335–340. [Google Scholar] [CrossRef]
- Mendez-Encinas, M.A.; Valencia-Rivera, D.E.; Carvajal-Millan, E.; Astiazaran-Garcia, H.; Rascón-Chu, A.; Brown-Bojorquez, F. Electrosprayed Highly Cross-Linked Arabinoxylan Particles: Effect of Partly Fermentation on the Inhibition of Caco-2 Cells Proliferation. AIMS Bioeng. 2020, 8, 52–70. [Google Scholar] [CrossRef]
- Chen, L.; Lin, X.; Teng, H. Emulsions Loaded with Dihydromyricetin Enhance Its Transport through Caco-2 Monolayer and Improve Anti-Diabetic Effect in Insulin Resistant HepG2 Cell. J. Funct. Foods 2020, 64, 103672. [Google Scholar] [CrossRef]
- Hernandez, J.; Goycoolea, F.; Quintero, J.; Acosta, A.; Castañeda, M.; Dominguez, Z.; Robles, R.; Vazquez-Moreno, L.; Velazquez, E.; Astiazaran, H.; et al. Sonoran Propolis: Chemical Composition and Antiproliferative Activity on Cancer Cell Lines. Planta Med. 2007, 73, 1469–1474. [Google Scholar] [CrossRef]
- Hou, W.-C.; Hsu, F.-L.; Lee, M.-H. Yam (Dioscorea Batatas) Tuber Mucilage Exhibited Antioxidant Activities in Vitro. Planta Med. 2002, 68, 1072–1076. [Google Scholar] [CrossRef]
- Nieves, M.; López, D.J.; Medina, M.A.; Piña, P.; Leal, S. Producción y calidad de Chaetoceros muelleri a diferentes concentraciones de nutrientes y densidades de inóculo. Rev. Investig. Mar. 2009, 30, 123–133. [Google Scholar]
- Huntsman, S.A.; Sloneker, J.H. An Exocellular Polysaccharide From The Diatom Gomphonema olivaceum. J. Phycol. 1971, 7, 261–264. [Google Scholar] [CrossRef]
- Fimbres-Olivarría, D.; López-Elías, J.; Carvajal-Millán, E.; Márquez-Escalante, J.; Martínez-Córdova, L.; Miranda-Baeza, A.; Enríquez-Ocaña, F.; Valdéz-Holguín, J.; Brown-Bojórquez, F. Navicula Sp. Sulfated Polysaccharide Gels Induced by Fe(III): Rheology and Microstructure. Int. J. Mol. Sci. 2016, 17, 1238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiwari, A.; Melchor-Martínez, E.M.; Saxena, A.; Kapoor, N.; Singh, K.J.; Saldarriaga-Hernández, S.; Parra-Saldívar, R.; Iqbal, H.M.N. Therapeutic Attributes and Applied Aspects of Biological Macromolecules (Polypeptides, Fucoxanthin, Sterols, Fatty Acids, Polysaccharides, and Polyphenols) from Diatoms—A Review. Int. J. Biol. Macromol. 2021, 171, 398–413. [Google Scholar] [CrossRef]
- Chen, S.; Sathuvan, M.; Zhang, X.; Zhang, W.; Tang, S.; Liu, Y.; Cheong, K.-L. Characterization of Polysaccharides from Different Species of Brown Seaweed Using Saccharide Mapping and Chromatographic Analysis. BMC Chem. 2021, 15, 1. [Google Scholar] [CrossRef] [PubMed]
- Shao, P.; Chen, M.; Pei, Y.; Sun, P. In Intro Antioxidant Activities of Different Sulfated Polysaccharides from Chlorophytan Seaweeds Ulva Fasciata. Int. J. Biol. Macromol. 2013, 59, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Saboural, P.; Chaubet, F.; Rouzet, F.; Al-Shoukr, F.; Azzouna, R.; Bouchemal, N.; Picton, L.; Louedec, L.; Maire, M.; Rolland, L.; et al. Purification of a Low Molecular Weight Fucoidan for SPECT Molecular Imaging of Myocardial Infarction. Mar. Drugs 2014, 12, 4851–4867. [Google Scholar] [CrossRef] [Green Version]
- Fernando, I.P.S.; Sanjeewa, K.K.A.; Samarakoon, K.W.; Lee, W.W.; Kim, H.-S.; Kim, E.-A.; Gunasekara, U.K.D.S.S.; Abeytunga, D.T.U.; Nanayakkara, C.; de Silva, E.D.; et al. FTIR Characterization and Antioxidant Activity of Water Soluble Crude Polysaccharides of Sri Lankan Marine Algae. Algae 2017, 32, 75–86. [Google Scholar] [CrossRef] [Green Version]
- Pereira, L.; Gheda, S.F.; Ribeiro-Claro, P.J.A. Analysis by Vibrational Spectroscopy of Seaweed Polysaccharides with Potential Use in Food, Pharmaceutical, and Cosmetic Industries. Int. J. Carbohydr. Chem. 2013, 2013, 537202. [Google Scholar] [CrossRef]
- Pielesz, A. Temperature-Dependent FTIR Spectra of Collagen and Protective Effect of Partially Hydrolysed Fucoidan. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2014, 118, 287–293. [Google Scholar] [CrossRef]
- Alboofetileh, M.; Rezaei, M.; Tabarsa, M. Enzyme-Assisted Extraction of Nizamuddinia Zanardinii for the Recovery of Sulfated Polysaccharides with Anticancer and Immune-Enhancing Activities. J. Appl. Phycol. 2019, 31, 1391–1402. [Google Scholar] [CrossRef]
- Lobus, N.V.; Kulikovskiy, M.S.; Maltsev, Y.I. Multi-Element Composition of Diatom Chaetoceros Spp. from Natural Phytoplankton Assemblages of the Russian Arctic Seas. Biology 2021, 10, 1009. [Google Scholar] [CrossRef] [PubMed]
- Tesson, B.; Genet, M.J.; Fernandez, V.; Degand, S.; Rouxhet, P.G.; Martin-Jézéquel, V. Surface Chemical Composition of Diatoms. ChemBioChem 2009, 10, 1915. [Google Scholar] [CrossRef]
- Ibrahim, M.I.A.; Amer, M.S.; Ibrahim, H.A.H.; Zaghloul, E.H. Considerable Production of Ulvan from Ulva Lactuca with Special Emphasis on Its Antimicrobial and Anti-Fouling Properties. Appl. Biochem. Biotechnol. 2022, 194, 3097–3118. [Google Scholar] [CrossRef] [PubMed]
- del Rio, A.R.; Ramírez-Gilly, M.; Tecante, A. Flow Properties of Lambda Carrageenan in Aqueous Systems. In Biological Activities and Application of Marine Polysaccharides; Shalaby, E.A., Ed.; InTech: London, UK, 2017; ISBN 978-953-51-2859-5. [Google Scholar]
- Danaei, M.; Dehghankhold, M.; Ataei, S.; Hasanzadeh Davarani, F.; Javanmard, R.; Dokhani, A.; Khorasani, S.; Mozafari, M. Impact of Particle Size and Polydispersity Index on the Clinical Applications of Lipidic Nanocarrier Systems. Pharmaceutics 2018, 10, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, M.Q.; Hu, X.; Wang, C.; Ai, L. Polysaccharides: Structure and Solubility. In Solubility of Polysaccharides; Xu, Z., Ed.; InTech: London, UK, 2017; ISBN 978-953-51-3649-1. [Google Scholar]
- Hadj Ammar, H.; Lajili, S.; Ben Said, R.; Le Cerf, D.; Bouraoui, A.; Majdoub, H. Physico-Chemical Characterization and Pharmacological Evaluation of Sulfated Polysaccharides from Three Species of Mediterranean Brown Algae of the Genus Cystoseira. DARU J. Pharm. Sci. 2015, 23, 1. [Google Scholar] [CrossRef] [Green Version]
- Bastías Solís, M.F. Determinación y Cuantificación de La Eficiencia de Asociación de Azitromicina En Nanopartículas Biopoliméricas de Quitosano y Condroitín Sulfato Mediante La Técnica de Diafiltración. Bachelor’s Thesis, Universidad Austral de Chile, Valdivia, Chile, 2017. [Google Scholar]
- Cortés Castell, E. Assessment of cytotoxicity and biosafety of polyphenolic extracts from olive pits. Nutr. Hosp. 2014, 29, 1388–1393. [Google Scholar] [CrossRef] [Green Version]
- Xia, S.; Gao, B.; Li, A.; Xiong, J.; Ao, Z.; Zhang, C. Preliminary Characterization, Antioxidant Properties and Production of Chrysolaminarin from Marine Diatom Odontella Aurita. Mar. Drugs 2014, 12, 4883–4897. [Google Scholar] [CrossRef] [Green Version]
- Chin, Y.X.; Mi, Y.; Cao, W.X.; Lim, P.E.; Xue, C.H.; Tang, Q.J. A Pilot Study on Anti-Obesity Mechanisms of Kappaphycus Alvarezii: The Role of Native κ-Carrageenan and the Leftover Sans-Carrageenan Fraction. Nutrients 2019, 11, 1133. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Xie, Q.; Huang, S.; Shao, P.; You, L.; Pedisić, S. Digestion & Fermentation Characteristics of Sulfated Polysaccharides from Gracilaria Chouae Using Two Extraction Methods in Vitro and in Vivo. Food Res. Int. 2021, 145, 110406. [Google Scholar] [CrossRef]
- Lordan, S.; Ross, R.P.; Stanton, C. Marine Bioactives as Functional Food Ingredients: Potential to Reduce the Incidence of Chronic Diseases. Mar. Drugs 2011, 9, 1056–1100. [Google Scholar] [CrossRef] [Green Version]
- Khan, T.; Date, A.; Chawda, H.; Patel, K. Polysaccharides as Potential Anticancer Agents—A Review of Their Progress. Carbohydr. Polym. 2019, 210, 412–428. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Tao, H.; Xie, J.; Zhang, S.; Xu, X. Degradation and Antioxidant Activity of κ-Carrageenans. J. Appl. Polym. Sci. 2010, 117, 194–199. [Google Scholar] [CrossRef]
- Qi, H.; Zhao, T.; Zhang, Q.; Li, Z.; Zhao, Z.; Xing, R. Antioxidant Activity of Different Molecular Weight Sulfated Polysaccharides from Ulva Pertusa Kjellm (Chlorophyta). J. Appl. Phycol. 2005, 17, 527–534. [Google Scholar] [CrossRef]
- Gropper, S.A.S.; Smith, J.L.; Carr, T.P. Advanced Nutrition and Human Metabolism, 7th ed.; Cengage Learning: Boston, MA, USA, 2018; ISBN 978-1-305-62785-7. [Google Scholar]
- Braithwaite, M.C.; Tyagi, C.; Tomar, L.K.; Kumar, P.; Choonara, Y.E.; Pillay, V. Nutraceutical-Based Therapeutics and Formulation Strategies Augmenting Their Efficiency to Complement Modern Medicine: An Overview. J. Funct. Foods 2014, 6, 82–99. [Google Scholar] [CrossRef]
- Lu, L.W.; Chen, J.-H. Seaweeds as Ingredients to Lower Glycemic Potency of Cereal Foods Synergistically—A Perspective. Foods 2022, 11, 714. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | % (w/w) |
|---|---|
| Total carbohydrates | 78 ± 0.3 |
| Protein | 5 ± 1.0 |
| Ash | 17 ± 0.2 |
| Mw (kDa) | 4.13 |
|---|---|
| Mn (kDa) | 1.94 |
| PDI (Mw/Mn) | 2.12 |
| [η] (mL/g) | 4.68 |
| Rh (nm) | 1.33 |
| Characteristic | Value |
|---|---|
| Sulfation degree (sulfate/carbohydrate ratio) | 0.10 ± 0.01 |
| Z Potential (mV) | +1.91 ± 0.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miranda-Arizmendi, V.; Fimbres-Olivarria, D.; Miranda-Baeza, A.; Martínez-Robinson, K.; Rascón-Chu, A.; De Anda-Flores, Y.; Lizardi-Mendoza, J.; Mendez-Encinas, M.A.; Brown-Bojorquez, F.; Canett-Romero, R.; et al. Sulfated Polysaccharides from Chaetoceros muelleri: Macromolecular Characteristics and Bioactive Properties. Biology 2022, 11, 1476. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11101476
Miranda-Arizmendi V, Fimbres-Olivarria D, Miranda-Baeza A, Martínez-Robinson K, Rascón-Chu A, De Anda-Flores Y, Lizardi-Mendoza J, Mendez-Encinas MA, Brown-Bojorquez F, Canett-Romero R, et al. Sulfated Polysaccharides from Chaetoceros muelleri: Macromolecular Characteristics and Bioactive Properties. Biology. 2022; 11(10):1476. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11101476
Chicago/Turabian StyleMiranda-Arizmendi, Valeria, Diana Fimbres-Olivarria, Anselmo Miranda-Baeza, Karla Martínez-Robinson, Agustín Rascón-Chu, Yubia De Anda-Flores, Jaime Lizardi-Mendoza, Mayra A. Mendez-Encinas, Francisco Brown-Bojorquez, Rafael Canett-Romero, and et al. 2022. "Sulfated Polysaccharides from Chaetoceros muelleri: Macromolecular Characteristics and Bioactive Properties" Biology 11, no. 10: 1476. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11101476