
Ethylene, ACC, and the Plant Growth-Promoting Enzyme ACC Deaminase



1
Dipartimento di Scienze e Innovazione Tecnologica, Università del Piemonte Orientale, Viale T. Michel 11, 15121 Alessandria, Italy
2
Department of Biology, University of Waterloo, Waterloo, ON N2L 3G1, Canada
*
Author to whom correspondence should be addressed.
Biology 2023, 12(8), 1043; https://0-doi-org.brum.beds.ac.uk/10.3390/biology12081043
Submission received: 29 June 2023
/
Revised: 20 July 2023
/
Accepted: 24 July 2023
/
Published: 25 July 2023
(This article belongs to the Collection Plant Growth-Promoting Bacteria: Mechanisms and Applications)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
The molecule 1-aminocyclopropane-1-carboxylate is the immediate precursor of the plant hormone ethylene in most seed plant species. Both 1-aminocyclopropane-1-carboxylate and ethylene can affect plant growth and development in a variety of ways. In addition, the bacterial enzyme 1-aminocyclopropane-1-carboxylate deaminase can cleave 1-aminocyclopropane-1-carboxylate and prevent both 1-aminocyclopropane-1-carboxylate and ethylene from affecting plant gene expression and subsequent behavior. In this review, the roles of 1-aminocyclopropane-1-carboxylate, ethylene and 1-aminocyclopropane-1-carboxylate deaminase in the development of plants and their regulation are explored. Since D-amino acids stimulate ethylene synthesis in plants, a section of the review is dedicated to this topic. It is suggested that 1-aminocyclopropane-1-carboxylate was synthesized in plants prior to the plant synthesis of ethylene, giving 1-aminocyclopropane-1-carboxylate some control of plant gene expression in response to environmental signals.
Abstract
Here, a brief summary of the biosynthesis of 1-aminocyclopropane-1-carboxylate (ACC) and ethylene in plants, as well as overviews of how ACC and ethylene act as signaling molecules in plants, is presented. Next, how the bacterial enzyme ACC deaminase cleaves plant-produced ACC and thereby decreases or prevents the ethylene or ACC modulation of plant gene expression is considered. A detailed model of ACC deaminase functioning, including the role of indoleacetic acid (IAA), is presented. Given that ACC is a signaling molecule under some circumstances, this suggests that ACC, which appears to have evolved prior to ethylene, may have been a major signaling molecule in primitive plants prior to the evolution of ethylene and ethylene signaling. Due to their involvement in stimulating ethylene production, the role of D-amino acids in plants is then considered. The enzyme D-cysteine desulfhydrase, which is structurally very similar to ACC deaminase, is briefly discussed and the possibility that ACC deaminase arose as a variant of D-cysteine desulfhydrase is suggested.
1. Introduction
Plant growth and development are regulated by several different phytohormones including cytokinins, gibberellins, auxins, salicylic acid, jasmonates, brassinosteroids, abscisic acid, strigolactones, and ethylene [1,2,3,4,5,6]. A key phytohormone, and of particular interest to this manuscript, is ethylene (C2H4), a low molecular weight (28.05 g/mol) gaseous hydrocarbon that is produced in all higher plants and modulates a wide range of plant physiological and biochemical activities [7,8,9]. Thus, for example, “ethylene is involved in seed germination, tissue differentiation, formation of root and shoot primordia, root branching and elongation, lateral bud development, flowering initiation, anthocyanin synthesis, flower opening and senescence, fruit ripening and degreening, production of volatile organic compounds, …aroma formation in fruits, storage product hydrolysis, leaf senescence, leaf and fruit abscission, Rhizobia nodule formation, mycorrhizae-plant interaction, and (importantly) the response of plants to various biotic and abiotic stress” [7]. The impact of ethylene on a particular plant trait may be either stimulatory or inhibitory, and is a consequence of the genus and species of the plant, the age of the plant, and the soil conditions, including the presence of soil microbes, the weather, and the amount of ethylene that is produced. Moreover, depending upon these conditions, a very wide range of ethylene concentrations (~4000-fold) may exhibit biological activity [7,10]. In addition, in some plants under specific conditions, the presence of increasing ethylene may impact the synthesis of other phytohormones including abscisic acid, gibberellin, cytokinin, and auxin [11,12]. Thus, some of the biological activities largely attributed to ethylene may be the consequence of ethylene affecting the concentration of, or acting in concert with, other phytohormones.
In this review, the role of the phytohormone ethylene and 1-aminocyclopropane-1-carboxylate (ACC), ethylene’s immediate precursor, to act as signaling molecules in plants is reviewed and discussed. In addition, the evidence for the ability of the widespread soil microbial enzyme ACC deaminase [13] to modulate ethylene [14,15,16] and ACC [17] signaling in primitive as well as modern plants is discussed. The similarity between ACC deaminase and the enzyme D-cysteine desulfhydratase [13,18] is also discussed and the possibility that ACC deaminase originated as a variant form of D-cysteine desulfhydratase is considered. Finally, the possible role of D-cysteine desulfhydratase and D-amino acids in general in primitive plants is addressed.
2. Ethylene and ACC Biosynthesis in Plants
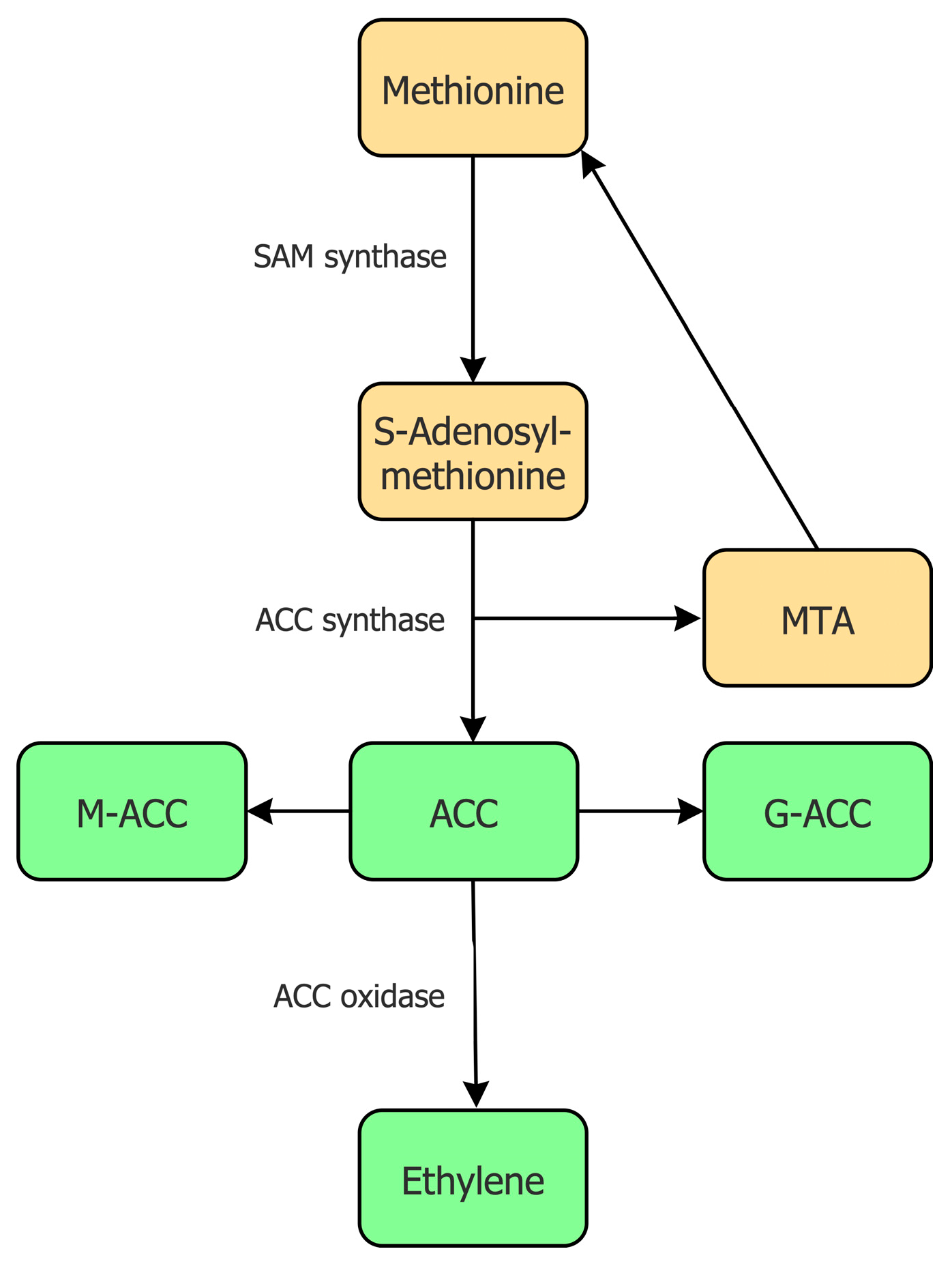
The biosynthesis of ethylene has been studied in detail in numerous higher plants and it appears that all of these plants utilize essentially the same mechanism to synthesize ethylene [19,20]. In addition to the plant biosynthesis of ethylene, some microorganisms can also synthesize ethylene, albeit using an entirely different biosynthetic pathway [21,22]. In plants, the biosynthesis of ethylene begins with the conversion of the relatively rare, but nonetheless very important, amino acid L-methionine into the compound S-adenosylmethionine (SAM) by the enzyme SAM synthase, which is encoded by a small multi-gene family (Figure 1).
Being at the junction of several biosynthetic pathways [23], SAM is moderately abundant within plant tissues. For the synthesis of ethylene, the compound SAM is converted into 1-aminocyclopropane-1-carboxylate (ACC) and 5′-methylthioadenosine (MTA) by the enzyme ACC synthase [24,25]. The MTA that is formed as a byproduct of this reaction is recycled to form the amino acid L-methionine. This allows the amount of L-methionine in a plant cell to remain relatively constant even during fairly high rates of ethylene production. Some scientists believe that the synthesis of ACC from SAM is the committal, or rate-limiting, step in the biosynthesis of ethylene [26,27]. There are several, nearly identical, ACC synthase enzymes present in a plant cell as a consequence of the fact that the genes that encode this enzyme are part of a multi-gene family. Moreover, considerable evidence suggests that the transcription of different genes that encode this enzyme are regulated under a range of different environmental or plant physiological conditions (e.g., [25]). When high levels of ACC synthesis are not required, the amount of ACC synthase in plant cells remains relatively low. The subsequent conversion of ACC to ethylene is catalyzed by the enzyme ACC oxidase [28,29,30], which is also generally present constitutively in most plant tissues at very low levels. Similar to ACC synthase genes, ACC oxidase genes are part of a multi-gene family with different isoforms of this enzyme being actively transcribed under different environmental or plant physiological conditions [31]. Thus, in many physiological conditions, both ACC synthase and ACC oxidase may be considered to be inducible enzymes. The fact that both enzymes are inducible is somewhat unusual in that in most metabolic pathways that have been studied only a single step is typically thought of as rate limiting. However, inducible ACC synthesis followed by inducible ethylene synthesis is consistent with the idea, posited below, that ACC synthesis and ethylene synthesis evolved separately from one another.
Plant cells often make more ACC than they require at any particular time. This enables them to rapidly respond to changing environmental conditions and to quickly synthesize ethylene from this storehouse of ACC. However, to remove some of the excessive ACC when it is not immediately needed, plant cells are able to convert ACC into inactive conjugated forms of this compound. For example, the conjugation of ACC with malonate or glutathione occurs as a consequence of the action of either the enzyme ACC N-malonyl transferase [32,33] or the enzyme gamma-glutamyl transpeptidase [34]. These reactions result in the production of either 1-(malonyl)-ACC (M-ACC) or 1-(glutamyl)-ACC (G-ACC), respectively (Figure 1). Kinetic studies have determined that the tightness of the enzyme ACC N-malonyl transferase binding to its substrate ACC is much lower than the tightness of the enzyme ACC oxidase for the same substrate. Thus, if these two enzymes are present in cells in similar amounts, the ACC will bind preferentially to ACC oxidase, and it will subsequently be converted into ethylene [19]. However, as indicated earlier, there is often an excess of ACC in the cell that needs to be removed, and it would be detrimental to the plant to convert all the available ACC into ethylene [35]. In this regard, the conjugation reactions remove ACC only when it is present in relatively high levels, i.e., there is more ACC than is required for the necessary ethylene synthesis.
3. Ethylene as a Signaling Molecule
Ethylene is one of the simplest signaling molecules with hormone-like behavior that is synthesized by plants. When it plays the role of a hormone, it regulates plant development (seed germination, cell elongation, fruit ripening, seed dispersal) and plant responses to environmental stresses such as soil pollution by metals, high salinity levels, low water availability, and sub-optimal temperatures, as well as pathogen (fungi, insects, nematodes) attack. The long history of research on plant ethylene, well described in the review paper written by Bakshi et al. [36], started in 1885 with the first observation by George Fahnestock reporting that illuminating gas used for lighting in both homes and streets negatively affected plant health and growth in a greenhouse in Philadelphia [37]. About fifteen years later, Dimitry Neljubow, a plant physiologist at the Botanical Institute of St. Petersburg University in Russia, identified ethylene as the active molecule in illuminating gas that affects plants [38]. Since then, a quite large number of papers have focused on the roles of ethylene in plants and on its biosynthetic pathways (see Section 2 of this review), and only more recently has attention been given to the perception of this gas through ethylene-binding sites and to the genes and proteins differentially expressed as a consequence of ethylene synthesis. All the major molecular elements involved in ethylene signaling pathways have been identified and described through a combination of molecular biology, cell biology, biochemistry, and genetic tools [10].
Based on the results of several studies, performed mainly on Arabidopsis thaliana, it has been hypothesized that the plant gene(s) dedicated to ethylene perception are derived from a cyanobacterium that transferred this coding DNA to the chloroplast genome [39,40,41]. According to one pioneering study, it has been estimated that about 4000 ethylene-binding sites are distributed through each tobacco leaf cell [42], mainly located in the endoplasmic reticulum membrane [43,44]. All of the ethylene receptors described in the literature show a hydrophobic N-terminal domain comprising the ethylene-binding domain [41,45,46], followed by a cytosolic domain, which contains ubiquitous sequences occurring in a plethora of other signaling molecules expressed by members of all the three kingdoms of life, playing a pivotal role in protein–protein interactions between the receptors [47,48,49,50]. At the base of the ethylene receptor is a protein homodimer that binds noncovalently with other homodimers leading to the formation of higher order homomeric and heteromeric complexes [49].
The binding of ethylene with its receptor is supported by a copper-based cofactor, which is required for ethylene-receptor functions, and is provided by the RAN1 (Responsive to Antagonist1) copper transporter [41]. According to the studies performed on A. thaliana, there are five ethylene receptors in this model plant that can be categorized into two clades, the first one containing ETR1 and ESR1 (Ethylene Response 1 and Ethylene Response Sensor 1, respectively) and the second represented by ETR2, ERS2, and EIN4 (Ethylene Response 2, Ethylene Response Sensor 2, and Ethylene Insensitive 4, respectively) [51]. A structural model of this ethylene receptor has been proposed by Schott-Verdugo et al. [52] and, very recently, the structure of the ethylene-binding domain of ETR1 has been elucidated by Azhar et al. [53]. Based on the fact that the functioning of the ethylene receptor is dependent on copper availability, it has been hypothesized that the whole mechanism of ethylene perception depends on an ancient copper transport mechanism that protects plant cells from the toxicity induced by high concentrations of this metal [54]. However, since the publication of this paper, there have been no further findings in support of this hypothesis.
The gene CTR1 (CONSTITUTIVE TRIPLE RESPONSE 1), coding for a serine/threonine protein kinase, behaves as a negative regulator of ethylene responses, where ethylene response in plants is suppressed by its protein kinase activity [55]. The N-terminal regulatory domain is closely connected to ETR1. Although this association is required in order to ensure the kinase activity of CTR1, the mechanism by which CTR1 is activated by the ethylene receptors is still unknown [56,57,58,59].
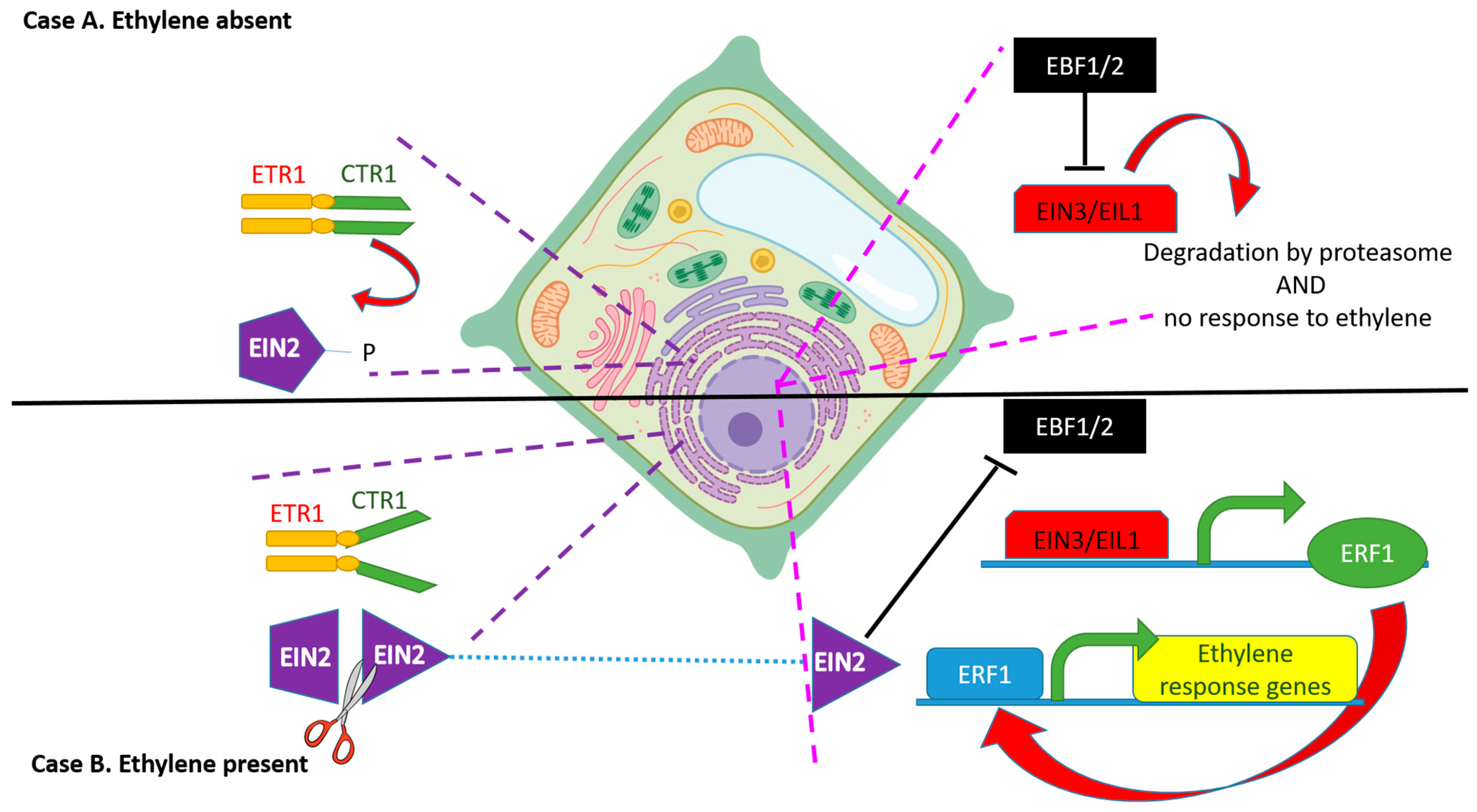
The protein EIN2 (ETHYLENE-INSENSITIVE 2), located in the endoplasmic reticulum membrane, shows 12 trans membrane domains at its N terminus and a plant specific domain that is involved in the activation of the ethylene downstream response at the C terminus [60,61]. It has been shown that A. thaliana has a single-copy EIN2 gene whose sequence is conserved from the charophyte green algae to land plants [62]. Its role is to transfer the ethylene plant response to EIN3 (ETHYLENE-INSENSITIVE 3), which is in the nucleus; in fact, when ethylene is perceived by the plant, EIN2 is cleaved by an unknown protease at the C terminal portion and the remaining sequence can move to the nucleus [63,64].
Additionally, EIN2-C can bind to the EBF1/EBF2 RNAs and be sequestered in cytoplasmic granules composed of translationally repressed mRNAs and proteins related to mRNA decay, called processing bodies (P-bodies) [16,64]. Finally, EIN2 stabilizes the two transcriptional factors EIN3 and EIL1 (ETHYLENE-INSENSITIVE 3-like 1 protein, homologous to EIN3), proteins which are key transcriptional factors involved in the modulation of the ethylene response genes such as ETHYLENE RESPONSE FACTORS (ERFs) [65,66]. The identification of this pathway allowed scientists to describe how an ethylene signal goes from the site of perception at the ER membrane and then to the nucleus. The pathway of ethylene signaling is depicted in Figure 2.
If ethylene is present (case B), it binds to the receptor and inactivates CTR1. This inactivation promotes the cleavage of the C terminus of EIN 2 protein.
The EIN2 C-terminal domain (EIN2-C), released upon cleavage, inhibits the translation of EBF1/EBF2 thus allowing accumulation of the EIN3- and EIN3-LIKE1 (EIL1)-transcription factors that activate the transcription of ERF1 (ETHYLENE RESPONSE FACTOR 1) and of many other genes involved in ethylene response. Altogether, the transcription of these genes then activates the plant ethylene response. Furthermore, EIN2-C can bind to the EBF1/EBF2 RNAs and become sequestered in processing bodies (P-bodies) in the cytoplasm.
The response of plants to ethylene is the same regardless of whether the ethylene is exogenously provided or endogenously synthesized.
4. ACC as a Signaling Molecule
Several studies have provided evidence that, in addition to acting as a precursor for the synthesis of ethylene, ACC itself can act as a hormonal signal [7,17,27,66,67,68,69,70,71]. The first indication of this possibility, using chemical inhibitors of ethylene biosynthesis or ethylene perception, was that ACC appeared to be a signaling molecule for root to shoot communication under conditions where ethylene perception was blocked [66]. In other experiments, ACC was shown to play a role in stomal development in A. thaliana [72]. In addition, other studies suggested that ACC plays a direct role in plant defenses against the fungal phytopathogen Verticillium dahliae [73]. For example, during periods of flooding, ACC, which is primarily synthesized in plant roots, is transported through the xylem to the shoots where, as a consequence of the availability of oxygen, the ACC is converted to ethylene [74,75]. It is possible that the precise role of ACC as a signaling molecule might be better defined, at least in some cases, by repeating some of the experiments that have previously been performed using chemical ethylene (synthesis or perception) inhibitors or ethylene biosynthesis mutants and instead include the presence of the enzyme ACC deaminase, which is an ACC rather than an ethylene inhibitor (see Section 5). Interestingly, it has been demonstrated that ACC behaves as a signal molecule involved in the recruitment of specific bacteria able to cleave ACC into ammonia and α-ketobutyrate, so that it shapes the rhizosphere microbiome. In turn, these bacteria reduce the stress levels in plants and this new physiological condition can subsequently modulate the composition of the plant-associated bacterial communities [76,77].
Finally, assuming that ACC is in fact a plant-signaling molecule, at least under certain circumstances, this is consistent with the possibility that ACC may have been a major signaling molecule in primitive plants prior to the development of ethylene and ethylene signaling, and prior to ACC becoming a precursor for the synthesis of ethylene in seed plants [78].
5. ACC Deaminase
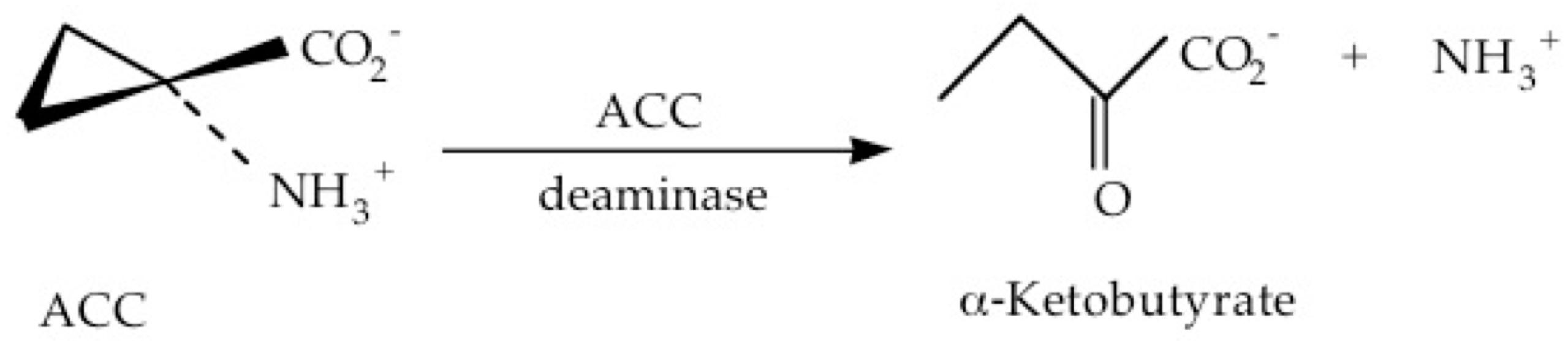
Plant growth-promoting bacteria (PGPB) that contain the enzyme ACC deaminase provide a significant advantage to plants growing in nature, in that this enzyme cleaves ACC into ammonia and α-ketobutyrate (Figure 3) thereby preventing the ACC from being converted to ethylene.
This is especially important during stressful conditions when plants often synthesize stress ethylene, which is usually inhibitory to plant growth [79,80]. For example, when scientists isolated PGPB from the rhizosphere of wild barley plants growing on two opposite slopes of a canyon in northern Israel, separated by ~250 m, they observed that the south- and north-facing slopes contained very different PGPB [81]. The south-facing slope was quite arid with a large amount of sunlight and had only very sparse plant growth. On the north-facing slope, the plant growth was quite lush. Both the south- and north-facing slopes contained similar plants (i.e., wild barley) and similar genera of bacteria within the plant rhizospheres. On closer examination, rhizosphere bacteria from the stressed south-facing slope all contained ACC deaminase, an activity that was present to a much lesser extent in the bacteria from the north-facing slope. Thus, the highly stressful conditions (drought and excess sunlight) on the south-facing slope selected for rhizospheric PGPB, containing ACC deaminase, which better enabled both the PGPB and the plants to survive under these harsh conditions. Importantly, ACC deaminase activity is not limited to the PGPB that occupy the rhizospheres of obviously stressed plants. Rather, it has been found in α, β, and γ Proteobacteria, Actinobacteria, Firmicutes, Bacteroidetes, Archaea, various fungi, and yeast [13,82].
Notwithstanding the abovementioned experiment, most studies of the efficacy of ACC deaminase in thwarting abiotic and biotic stresses have been performed in laboratory experiments, including both growth chamber and greenhouse experiments [83]. Based on many hundreds of reports of ACC deaminase containing PGPB protecting plants against a wide range of both abiotic and biotic stresses [84,85,86,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101,102,103,104,105,106,107,108,109,110,111,112], it is clear that this enzyme is a key component of the ability of plants to survive and thrive in a multitude of different stressful environments.
Where it has been studied, ACC deaminase has been found to be a cytoplasmically localized enzyme that is not secreted to the external medium [113]. Rather, the substrate, ACC, must be taken up by the bacterium that contains this enzyme. The active form of ACC deaminase is multimeric (probably tetrameric) with one mole of the co-factor pyridoxal phosphate per mole of the monomeric enzyme subunit [114]. ACC deaminase enzymes from different microbe’s function optimally between pH 8.0 and 9.0 and typically have a subunit molecular mass of 33–42 kDa [115]. Analyses of ACC deaminase enzyme structures suggest that this enzyme is relatively thermodynamically stable with a Tm = ~60 °C [116].
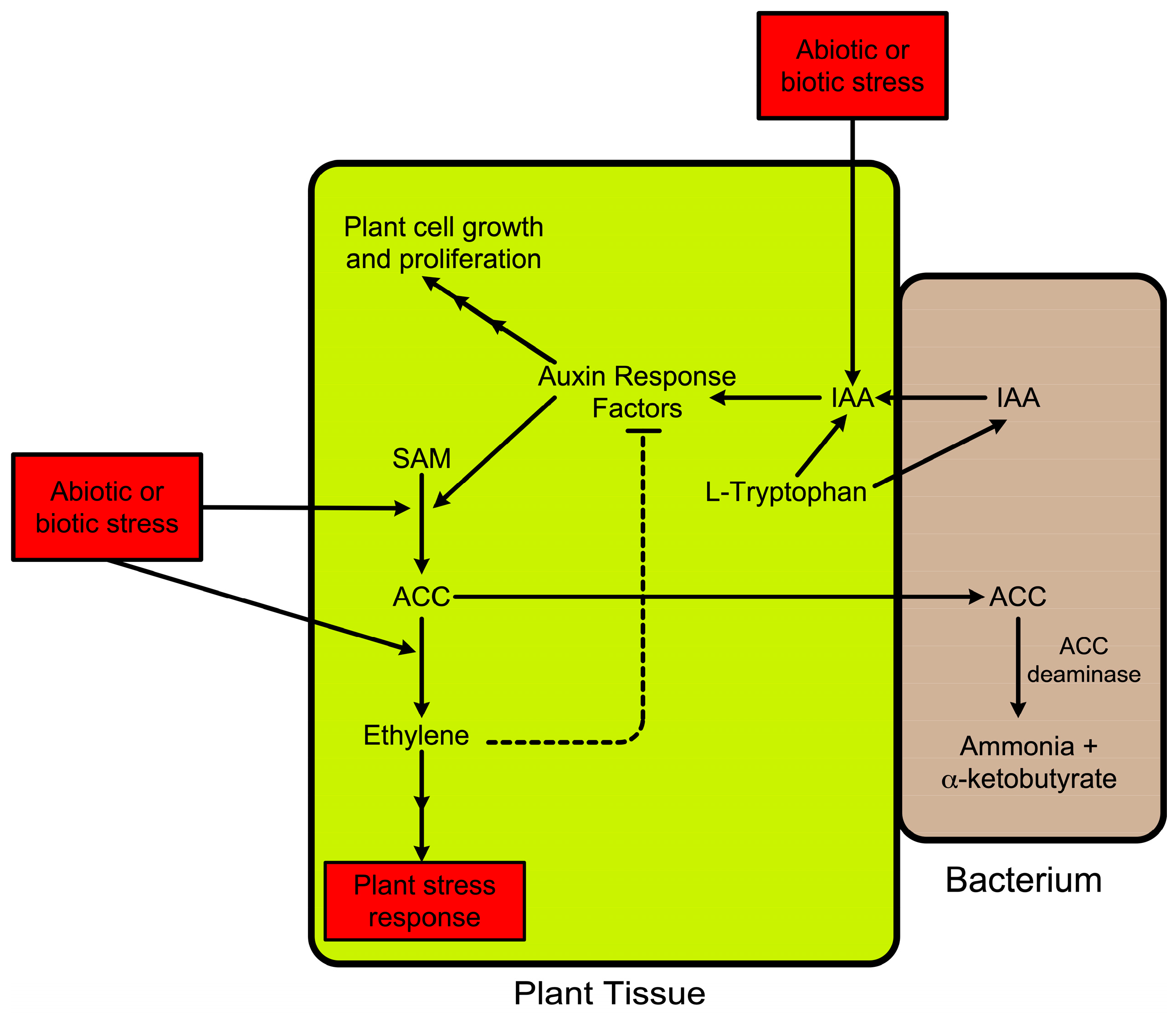
A model that was developed to understand the promotion of plant growth and development by ACC deaminase from PGPB [15,117] includes several different steps (Figure 4).
In this model, following the binding of a bacterium to a plant tissue (usually plant roots or plant seeds) or, in the case of endophytic bacteria, the uptake of the bacteria into plant tissues, the bacteria take up the amino acid L-tryptophan from the plant and use it to synthesize the phytohormone indoleacetic acid [118,119]. Some of the newly synthesized IAA is secreted by the bacteria (which are typically either bound to the surface of plant roots or localized within the root endosphere), taken up by plant tissue, and added to the endogenous IAA pool in the plant. IAA, through a series of metabolic steps (including auxin response factors and auxin transport proteins) [120], can either promote plant-cell growth and proliferation or activate the transcription of ACC synthase [121]. Increased ACC synthase transcription will result in an increase in ACC and ethylene synthesis with the increased amount of ethylene feed-back inhibiting auxin response factors [120] thereby limiting the additional functioning of IAA. Increasing IAA also results in plant-cell wall loosening [122] therefore increasing the amount of exudation of small molecules, including ACC, from the plant [84]. The exuded ACC is taken up by the plant-associated bacteria and cleaved into ammonia and alpha-ketobutyrate, both of which can be metabolized by the bacteria. This results in a decreased level of ethylene inside of the plant cells and a decreased amount of ethylene-caused feed-back inhibition of auxin response factors. Thus, in the presence of ACC deaminase, more IAA is directed toward plant-cell growth and proliferation. The net result of this scheme is that a PGPB that contains both ACC deaminase and the ability to synthesize IAA can (i) lower the ethylene inhibition of plant growth and (ii) simultaneously increase the IAA metabolic flux in the plant, both of which promote plant growth. It should be noted that while not all rhizospheric and endophytic PGPB produce IAA, multiple studies have suggested that around 85–90% of these bacteria have the ability to synthesize IAA. Finally, since both abiotic and biotic stresses generally increase plant ethylene levels, this model explains how ACC deaminase can decrease some of the deleterious effects of these stresses on plants.
6. D-Amino Acids in Plants
The L-form of amino acids, called proteinogenic amino acids, is by far the more prevalent one. There is a consensus on the fact that the prevalence of the L isoforms of amino acids dates back to ancient times. In fact, it has been reported that L-enantiomers of amino acids are dominant in the composition of a carbonaceous chondrite meteorite (Murchison meteorite) that fell in Australia in 1969 [123,124,125]. Since the formation of carbonaceous chondrites dates to ~4.5 billion years ago, it has been hypothesized that the amino acids’ molecular asymmetry appeared before the emergence of life on Earth [123]. This trait was probably randomly selected during the molecular evolution of life. Although amino acid homochirality is a quite rigorous condition, proteins and peptides containing D-amino acids are synthesized mainly after the incorporation of D-amino acids by a non-ribosomal peptide synthetase (NRPS) or through post translational modifications of peptide precursors based on L-amino acids [126]. While the first strategy is followed only by prokaryotes, the second one is exploited by both prokaryotes and eukaryotes. When the first occurrence of D-amino acids was observed in organisms, it was thought that they were peculiar but without any specific role. More recently, it has been demonstrated that D-amino acids have several structural and physiological functions. For example, the D-amino acids, D-alanine and D-glutamate, together with other D-amino acids such as D-aspartate and D-serine, are fundamental building blocks in bacterial peptidoglycan synthesis [127]. These molecules are generally considered to be detrimental for plant growth; however, plants are able to synthesize D-amino acids thus suggesting a functional role for these molecules in plants [128]. In fact, it has been demonstrated that D-serine behaves as a signal molecule modulating pollen growth in the pistil of A. thaliana [129].
Plants are constantly in contact with D-amino acids, which are very concentrated in the soil (in the order of milligrams per kg of soil) surrounding the root system [130]. Once they are transported inside plants, D-amino acids may be used as a nitrogen source modulating chloroplast division and the production of ethylene and affecting plant development according to the D-amino acid and the plant species involved [131,132,133]. Moreover, several genes involved in D-amino acid synthesis have been detected in plant genomes [134].
Regarding D-amino acid transport inside plants, workers have identified a lysine histidine transporter 1 (LTH1) as being responsible for the uptake of different amino acids at the root level [131,135]. Besides LTH1, two other families of transporters have been found to be able to take up D-amino acids. They are (1) the Amino acid Permease 1 (AAP1), which is mainly responsible for the uptake of D-methionine and D-phenylalanine [136], and (2) the proline transporter family ProT [137]. The common trait among these uptake systems is that they are not specific for D-amino acids since they are primarily responsible for the transport of L-amino acids.
On the other hand, D-amino acids are passively exuded through diffusion in A. thaliana [138]. This is interesting, since the uptake of D-amino acids occurs through an active ATP-consuming mechanism while their release by roots is mediated using passive transport [138]. Whether D-amino acids, together with all the other molecules contained in rhizo-deposits, are involved in the stimulation, inhibition, or establishment of certain bacterial communities remains to be elucidated.
As previously stated, according to the plant species and the D-amino acid considered, the effect of these molecules on plant development varies. The exogenous treatment of pepper plants with a mixture of D-Leucine, D-valine, and D-cysteine led to growth promotion possibly related to the utilization of these amino acids as a nitrogen source [130]. On the other hand, negative effects on plant growth were induced in A. thaliana by treatments with D-alanine at concentrations higher than 0.5 mM [131].
In some plant species, exogenous treatment with D-methionine enhanced the ethylene level. Interestingly, in A. thaliana, D-methionine is one of the preferred substrates of the enzyme AtDAT1 (a transaminase specific for D-amino acids). In the presence of D-methionine, the seeds of A. thaliana that lack the gene encoding AtDAT1 or are unable to synthesize this enzyme show a decreased level of germination due to increased ethylene levels. In fact, mutants lacking this gene are characterized by a high malonylation of D-methionine, combined with a reduction in malonyl-ACC content, which is the major product of ACC degradation [139]. Consequently, D-methionine regulates ethylene synthesis with high concentrations of D-methionine leading to this D-amino acid to outcompete ACC for the enzyme N-malonyl-transferase. As a result, the high amount of residual ACC is oxidized to ethylene by the enzyme ACC oxidase [30].
A functional role has also been recognized for D-cysteine, behaving as a precursor of hydrogen sulfide that is involved in plant stress responses [140]. Finally, a study performed on the moss Physcomitrella patens (now Physcomitrium patens) revealed the presence of the dipeptide D-Ala-D-Ala, as it typically occurs in the bacterial peptidoglycan, in the plastid envelope. Mutants of P. patens lacking the ability to synthesize the D-Ala-D-Ala dimer are hampered in plastid division [141]. Since the mutants of A. thaliana that lack the orthologous genes remained unaffected in plastid division, it has been hypothesized that the ability to synthesize peptidoglycan and integrate it into plastidic envelopes is a trait that has been lost during the evolution of land plants after the development of lycophytes [142].
However, it is also true that genes for peptidoglycan synthesis have been found in Picea abies and Pinus taeda and at least four genes (if not the complete set) involved in peptidoglycan formation have been detected in several angiosperms [143]. Thus, the possible presence of peptidoglycan in higher plants is still sparking scientific debate.
7. D-Cysteine Desulfhydrase
D-cysteine desulfhydrase is a pyridoxal-5′-phosphate-dependent enzyme that breaks down D-cysteine into pyruvate, hydrogen sulfide, and ammonia [144]. This enzyme can also break down β-chloro-D-alanine [145]. This enzyme has been detected in both plants [18,146,147,148,149] and bacteria [144,150]. Moreover, the amino acid sequence of D-cysteine desulfhydrase is highly homologous to several ACC deaminases [13,18,150] and the ACC molecule is able to bind to the active site of crystallized D-cysteine desulfhydrase from Salmonella typhimurium [145] even though D-cysteine desulfhydrase does not have any ACC deaminase activity. In E. coli and yeast, D-cysteine is toxic to the microbes, possibly because of its inhibition of the activity of the enzyme threonine deaminase [150]. In addition to plants and microbes, D-cysteine desulfhydrase activity has been purified and characterized from the green alga Chlorella fusca [147]. Interestingly, the properties of all the investigated D-cysteine desulfhydrases are quite similar.
In a key experiment, tomato (Solanum lycopersicum) cDNA that was predicted to encode a putative ACC deaminase was isolated and expressed in E. coli [18]. However, a detailed kinetic assessment of this expressed cDNA revealed that it encoded D-cysteine desulfhydrase and not ACC deaminase. The subsequent site-directed mutagenesis of this cDNA, altering the codons for two amino acid residues that were within the predicted active site of the enzyme D-cysteine desulfhydrase, changed the activity of the enzyme from D-cysteine desulfhydrase to ACC deaminase. Moreover, the site-directed mutagenesis of the codons for two amino acid residues within the same position of the active site of ACC deaminase from the plant growth-promoting bacterium Pseudomonas sp. UW4 [151] changed its activity from ACC deaminase to D-cysteine desulfhydrase. Following a detailed phylogenetic study [13] of ACC deaminase genes, it was observed that ACC deaminase genes and D-cysteine desulfhydrase genes clustered separately, albeit nearby, to one another. Together, these data are consistent with, but do not prove, the possibility that ACC deaminase evolved from the enzyme D-cysteine desulfhydrase or a “similar pyridoxal phosphate dependent deaminase related to tryptophan synthase beta subunit and sharing a common origin” [13].
8. Plant Evolution
The founding event in the origin of plants and other photosynthetic eukaryotes was the acquisition of the chloroplast, the cellular organelle responsible for photosynthesis, and it is estimated to date to ~1.6 billion years ago. Chloroplasts originate from a cyanobacterium that was incorporated into a eukaryotic cell, most likely a freshwater protozoon that fed on bacteria, by phagocytosis and then transformed into an organelle to be inherited from one generation to the next. Molecular data support a monophyletic origin of chloroplasts in all plants [152,153,154].
The transformation of a phagocyted cyanobacterium was associated with a few changes in both organisms; the protozoon acquired a cell wall, lost the ability to feed by phagocytosis, and became autotrophic for carbon while the cyanobacterium became an organelle surrounded by two membranes. Both organisms underwent a rearrangement of their genomes, since a number of genes were transferred from the prokaryote to the nucleus of the protozoon. Following these events, the cyanobacterium became unable to live on its own, integrated into the eukaryotic cell, and the two eventually formed a permanent partnership. Most plant biologists agree on the above description concerning the origin of chloroplasts and plastid phylogenomic analysis has been used to explore the whole phylogeny of green plants [155,156]. However, it must be acknowledged that alternative hypotheses are still attempting to challenge the above-mentioned view [157].
A recent model for eukaryote classification is based on the organization into eight supergroups (plus some other smaller groups), which are rather different from the conventional eukaryotic Kingdoms: Amorphea (Amebozoa, Fungi, Animalia), Archaeplastida (Glaucophyta, Rhodophyta, and Chloroplastida), TSAR (Stramenopila, Alveolata, and Rhizaria), Haptista, Cryptista, CRuMs, Hemimastigophora, and Excavata [158]. Photosynthetic organisms are present in several supergroups, but here the focus is on Archaeplastida, since the photosynthetic members of other eukaryotic supergroups most likely originated through secondary symbiosis involving unicellular organisms belonging to the clade Archaeplastida.
Archeaeplastida comprise three main lines: Glaucophyta (a group of unicellular algae), Rhodophyta (red algae), and Chloroplastida [159]. In Glaucophyta and Rhodophyta, chloroplasts still morphologically resemble cyanobacteria, and their photosynthetic pigment composition is characterized by the presence of both chlorophyll a and phycobiliproteins, like cyanobacteria. Chloroplastida are green plants comprising “green algae” and land plants, with phycobiliproteins replaced with chlorophyll b, a change that allows for the reorganization and packing of the thylakoid membranes, the system found in chloroplasts and cyanobacteria [160,161,162]. In addition, green plants accumulate starch (the main carbon reserve material in Plantae) in chloroplasts, while Glaucophyta and Rhodophyta accumulate starch in the cytoplasm.
Green plants developed along two main lines, one was caused by the Chlorophyta, the other one by the Streptophyta. Chlorophyta represent the vast majority of green algae in the seas and freshwaters, while Streptophyta include a small group of freshwater algae, Charophytina, and all the land plants, Embryophytina. Streptophyta represent the most interesting group for the purposes of this review since, in this clade, ethylene production via the action of the enzymes ACS and ACO has been described in seed plants.
Indeed, the ethylene reception system seems to be much older than that concerning the phytohormone biosynthesis since a gene, known as Etr1 or PIXA or UirS, coding for an ethylene receptor protein, was found in the cyanobacterium Synechocystis; the receptor comprises three domains: an ethylene binding one, a phytochrome-like one able to respond to photons, and a histidine kinase one, and it is involved in photaxis [163,164,165,166]. A recent study concerning the investigation of transcriptomic changes occurring in Synechocystis in response to ethylene during phototaxis revealed that the application of ethylene modulated over 500 gene transcripts [167]. Plants probably inherited this gene, through transfer from the plastid, about 1–1.5 billion years ago [42].
Data concerning chlorophyte green algae indicate that the ethylene receptor gene was lost when this group of organisms and the charophyte green algae diverged, over one billion years ago [168].
Ju et al. [62] investigated the origin of ethylene as a plant hormone by mining the transcriptome datasets of five species of charophytes, representative of the main lineages of these green algae, and performing a comparison with data from representatives of five lineages of embryophytes: mosses, lycophytes, gymnosperms, monocot angiosperms, and eudicot angiosperms. In the charophyte sequences (with some differences in the five considered species), homologues of all the central genes in the ethylene biosynthesis and the signaling pathway of plants were identified; Spyrogyra pratensis and Choleochaete orbicularis presented the most complete sets of ethylene-related homologous genes. Therefore, S. pratensis was tested for ethylene biosynthesis and response. The results showed that S. pratensis cultures produced detectable levels of ethylene and the levels of ethylene increased when exogenous ACC was provided to the medium but to a low extent compared with the amount of applied ACC. The enzyme possibly involved is yet unknown. The treatment of submerged S. pratensis cultures with exogenous ethylene resulted in dose-dependent cell elongation; this response was prevented using a prior treatment with a competitive inhibitor of ethylene binding in land plants. Ethylene-dependent cell elongation is a typical response to submersion in land plants [11]. It is worth mentioning that ACC was also able to induce cell elongation in the submerged cultures of S. pratensis, providing evidence that this alga can respond to endogenous ethylene. Finally, the transformation of the Arabidopsis ETR1 ethylene receptor, SpETR1, with the S. pratensis homologous gene was able to alleviate the short hypocotyl phenotype of the triple ethylene receptor mutant Arabidopsis etr1-7, etr2-3, and ein4-4.
Overall, these results show that ethylene functions as a hormone in some lines of charophytes and their signaling pathways are conserved compared with Arabidopsis. Therefore, these results strongly suggest that ethylene was already a functional hormone in the common ancestor of charophytes and land plants, at least 450 million years ago.
Ethylene synthesis upon ACC treatment was observed in some additional organisms, like cyanobacteria, red algae, and some chlorophytes [169,170,171,172,173], but the enzymes for the synthesis of ethylene are unknown in these organisms.
An increasing body of evidence supports the idea that ACC plays a role as a signaling molecule on its own, beyond being the precursor of ethylene ([78] and Section 4 of this review). Both ACS activity and ACC were detected in liverworts, mosses, and ferns [172,173,174]. It is therefore possible that the system for ethylene perception may have evolved before the ability to synthesize ethylene from ACC, since the former was already present in cyanobacteria [167]. So far, ACO homologs have not been found in the genome sequences of non-seed plants [175], while ACS homologs are conserved in land plants, consistent with the notion that ACC biosynthesis evolved before the ACC-dependent synthesis of ethylene.
9. Concluding Remarks
Ethylene is one of the key phytohormones affecting a wide range of plant gene expression and subsequent behavior. The currently available data suggest that ethylene was already a functional plant hormone ~450 million years ago (around the time that land plants began to emerge). In addition, other data are consistent with the notion that ACC acted as a plant hormonal signal prior to the emergence of plant-produced ethylene as a phytohormone (with ACC currently retaining a vestige of that early hormonal activity). The enzyme, ACC deaminase, synthesized by numerous PGPB (which have positively interacted with plants for many millions of years), cleaves ACC thereby preventing it from being converted to ethylene and from acting as a phytohormone. It appears that ACC deaminase may have evolved from the enzyme D-cysteine desulfhydrase, or another similar pyridoxal phosphate-dependent deaminase related to the tryptophan synthase beta subunit. Taken together, these data help to facilitate an understanding of the central role played by ethylene in plant growth and development. Moreover, these data emphasize the key role of PGPB that produce ACC deaminase in regulating plant ACC and ethylene levels, especially during periods of stress.
Author Contributions
Conceptualization, B.R.G.; writing—original draft preparation, E.G., G.L. and B.R.G.; writing—review and editing, E.G., G.L. and B.R.G.; supervision B.R.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Orozco-Mosqueda, M.C.; Santoyo, X.; Glick, B.R. Recent advances in the bacterial phytohormone modulation of plant growth. Plants 2023, 12, 606. [Google Scholar] [CrossRef]
- Weyers, J.D.B.; Patterson, N.W. Plant hormones and the control of physiological processes. N. Phytol. 2001, 152, 375–407. [Google Scholar] [CrossRef]
- Santaner, A.; Calderon-Villalobos, L.I.A.; Estelle, M. Plant hormones are versatile regulators of plant growth. Nat. Chem. Biol. 2009, 5, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Sabagh, A.E.L.; Islam, M.S.; Hossain, A.; Iqbal, M.A.; Mubeen, M.; Waleed, M.; Reginato, M.; Battaglia, M.; Ahmed, S.; Rehman, A.; et al. Phytohormones as growth regulators during abiotic stress tolerance in plants. Front. Agron. 2022, 4, 765068. [Google Scholar] [CrossRef]
- Zhao, B.; Liu, Q.; Wang, B.; Yuan, F. Roles of phytohormones and their signaling pathways in leaf development and stress responses. J. Agric. Food Chem. 2021, 69, 3566–3584. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Bano, A.; Ali, S.; Babar, M.A. Crosstalk amongst phytohormones from planta and PGPR under biotic and abiotic stress. Plant Grow. Regulat. 2020, 90, 189–203. [Google Scholar] [CrossRef]
- Abeles, F.; Morgan, P.W.; Saltveit, M.E., Jr. Ethylene in Plant Biology, 2nd ed.; Academic Press: San Diego, CA, USA, 1992. [Google Scholar]
- Glick, B.R. Beneficial Plant-Bacterial Interactions, 2nd ed.; Springer: Heidelberg/Berlin, Germany, 2020; 383p. [Google Scholar]
- Mattoo, A.K.; Suttle, J.C. The Plant Hormone Ethylene; CRC Press, Inc.: Boca Raton, FL, USA, 1991; 347p. [Google Scholar]
- Chang, C.; Qamp, A. How do plants respond to ethylene and what is its importance? BMC Biol. 2016, 14, 7. [Google Scholar] [CrossRef] [Green Version]
- Jackson, M.B. Ethylene-promoted elongation: An adaptation to submergence stress. Ann. Bot. 2008, 101, 229–248. [Google Scholar] [CrossRef]
- Iqbal, N.; Khan, N.A.; Ferrante, A.; Tivellini, A.; Francini, A.; Khan, M.I.R. Ethylene role in plant growth, development and senescence: Interaction with other phytohormones. Front. Plant Sci. 2017, 8, 475. [Google Scholar] [CrossRef] [Green Version]
- Nascimento, F.X.; McConkey, B.J.; Glick, B.R. New insights into ACC deaminase phylogeny, evolution, and evolutionary significance. PLoS ONE 2014, 9, e99168. [Google Scholar] [CrossRef] [Green Version]
- Glick, B.R. The enhancement of plant growth by free-living bacteria. Can. J. Microbiol. 1995, 41, 109–117. [Google Scholar] [CrossRef]
- Glick, B.R.; Penrose, D.M.; Li, J. A model for the lowering of plant ethylene concentrations by plant growth promoting bacteria. J. Theor. Biol. 1998, 190, 63–68. [Google Scholar] [CrossRef]
- Binder, B.M. Ethylene signaling in plants. J. Biol. Chem. 2020, 295, 7710–7725. [Google Scholar] [CrossRef] [PubMed]
- Yoon, G.M.; Kieber, J.J. 14-3-3 regulates 1-aminocyclopropane-1-carboxylate synthase protein turnover in Arabidopsis. Plant Cell 2013, 25, 1016–1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todorovic, B.; Glick, B.R. The interconversion of ACC deaminase and D-cysteine desulfhydrase by directed mutagenesis. Planta 2008, 229, 193–205. [Google Scholar] [CrossRef]
- Pattyn, J.; Vaughn-Hirsch, J.; Van de Poel, B. The regulation of ethylene biosynthesis: A complex multilevel control circuitry. N. Phytol. 2021, 229, 770–782. [Google Scholar] [CrossRef] [PubMed]
- Fluhr, R.; Matoo, A.K.; Dilley, D.R. Ethylene—Biosynthesis and perception. Crit. Rev. Plant Sci. 2008, 15, 479–523. [Google Scholar]
- Freebairn, H.T.; Buddenhagen, I.W. Ethylene production by Pseudomonas solanacearum. Nature 1964, 202, 313–314. [Google Scholar] [CrossRef]
- Eckert, C.; Xu, W.; Xiong, W.; Lynch, S.; Ungerer, J.; Tao, L.; Gill, R.; Maness, P.-C.; Yu, J. Ethylene-forming enzyme and bioethylene production. Biotechnol. Biofuels 2014, 7, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, B.; Li, C.; Tarczynski, M.C. High free-methionine and decreased lignin content result from a mutation in the Arabidopsis S-adenosyl-L-methionine synthetase 3 gene. Plant J. 2002, 29, 371–380. [Google Scholar] [CrossRef]
- Zarembinski, T.I.; Theologis, A. Ethylene biosynthesis and action: A case of conservation. Plant Mol. Biol. 1994, 26, 1579–1597. [Google Scholar] [CrossRef]
- Eun, H.-D.; Ali, S.; Jung, H.; Kim, K.; Kim, W.-C. Profiling of ACC synthase gene (ACS11) expression in Arabidopsis induced by abiotic stresses. Appl. Biol. Chem. 2019, 62, 42. [Google Scholar] [CrossRef] [Green Version]
- Harpaz-Saad, S.; Yoon, G.M.; Mattoo, A.; Kieber, J.J. The formation of ACC and competition between polyamides and ethylene for SAM. Ann. Plant Rev. 2012, 44, 53–81. [Google Scholar]
- Polko, J.K.; Kieber, J.J. 1-Aminocyclopropane 1-carboxylic acid and its emerging role as an ethylene-dependent growth regulator. Front. Plant Sci. 2019, 10, 1602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kende, H. Enzymes of ethylene biosynthesis. Plant Physiol. 1989, 91, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, A.J.; Lycett, G.W.; Grierson, D. Antisense gene that inhibits synthesis of the hormone ethylene in transgenic plants. Nature 1990, 346, 284–287. [Google Scholar] [CrossRef]
- Houben, M.; Van de Poel, B. 1-Aminocyclopropane-1-carboxylic acid oxidase (ACO): The enzyme that makes the plant hormone ethylene. Front. Plant Sci. 2019, 10, 695. [Google Scholar] [CrossRef] [Green Version]
- Barry, C.S.; Blume, B.; Bouzayen, M.; Cooper, W.; Hamilton, A.J.; Grierson, D. Differential expression of the 1-aminocyclopropane-1-carboxylate oxidase gene family of tomato. Plant J. 1996, 9, 525–535. [Google Scholar] [CrossRef] [PubMed]
- Peiser, G.; Yang, S.F. Evidence for 1-(malonylamino)cyclopropane-1-carboxylic acid being the major conjugate of aminocyclopropane-1-carboxylic acid in tomato fruit. Plant Physiol. 1998, 116, 1527–1532. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.F. The biosynthesis and metabolism of 1-(malonylamino) cyclopropane-1-carboxylic acid in relation to ethylene production. In Conjugated Plant Hormones: Structure, Metabolism, and Function; Schreiber, K., Schütte, H.R., Sembdner, G., Eds.; VEB Deutscher Verlag der Wissenschaaften: Berlin, Germany, 1987; pp. 92–101. [Google Scholar]
- Martin, M.N.; Cohen, J.D.; Saftner, R.A. A new 1-aminocyclopropane-1-carboxylic acid conjugating activity in tomato fruit. Plant Physiol. 1995, 109, 917–926. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.F.; Hoffman, N.E. Ethylene biosynthesis and its regulation in higher plants. Annu. Rev. Plant Physiol. 1984, 35, 155–189. [Google Scholar] [CrossRef]
- Bakshi, A.; Shemansky, J.M.; Chang, C.; Binder, B.M. History of research on the plant hormone ethylene. J. Plant Growth Regul. 2015, 34, 809–827. [Google Scholar] [CrossRef]
- Fahnestock, G.W. Memoranda of the effects of carbureted hydrogen gas upon a collection of exotic plants. Proc. Acad. Nat. Sci. Phil. 1858, 9–10, 118–134. [Google Scholar]
- Neljubow, D. Uber die horizontale nutation der stengel von Pisum sativum und einiger anderen pflanzen. Beih. Bot. Zentralb. 1901, 10, 128–139. [Google Scholar]
- Bleecker, A.B. Ethylene perception and signaling: An evolutionary perspective. Trends Plant Sci. 1999, 4, 269–274. [Google Scholar] [CrossRef]
- Mount, S.M.; Chang, C. Evidence for a plastid origin of plant ethylene receptor gene. Plant Physiol. 2002, 130, 10–14. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, F.I.; Esch, J.J.; Hall, A.E.; Binder, B.M.; Schaller, G.E.; Bleecker, A.B. A copper cofactor for the ethylene receptor ETR1 from Arabidopsis. Science 1999, 283, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Sisler, E.C. Measurement of ethylene binding in plant tissue. Plant Physiol. 1979, 64, 538–542. [Google Scholar] [CrossRef] [Green Version]
- Evans, D.E.; Bengochea, T.; Cairns, A.J.; Dodds, J.H.; Hall, M.A. Studies on ethylene binding by cell-free preparations from cotyledons of Phaseolus vulgaris L.: Subcellular localization. Plant Cell Environ. 1982, 5, 101–107. [Google Scholar]
- Evans, D.E.; Dodds, J.H.; Lloyd, P.C.; Apgwynn, I.; Hall, M.A. A study of the subcellular localization of an ethylene binding site in developing cotyledons of Phaseolus vulgaris L. by high resolution autoradiography. Planta 1982, 154, 48–52. [Google Scholar] [CrossRef] [PubMed]
- O’Malley, R.C.; Rodriguez, F.I.; Esch, J.J.; Binder, B.M.; O’Donnell, P.; Klee, H.J.; Bleecker, A.B. Ethylene-binding activity, gene expression levels, and receptor system output for ethylene receptor family members from Arabidopsis and tomato. Plant J. 2005, 41, 651–659. [Google Scholar] [CrossRef]
- Hall, A.E.; Findell, J.L.; Schaller, G.E.; Sisler, E.C.; Bleecker, A.B. Ethylene perception by the ERS1 protein in Arabidopsis. Plant Physiol. 2000, 123, 1449–1458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, F.; Liu, Q.; Wen, C.-K. Receptor signal output mediated by the ETR1 N terminus is primarily subfamily I receptor dependent. Plant Physiol. 2006, 142, 492–508. [Google Scholar] [CrossRef] [Green Version]
- Grefen, C.; Städele, K.; Růzicka, K.; Obrdlik, P.; Harter, K.; Horák, J. Subcellular localization and in vivo interactions of the Arabidopsis thaliana ethylene receptor family members. Mol. Plant 2008, 1, 308–320. [Google Scholar] [CrossRef]
- Gao, Z.; Wen, C.-K.; Binder, B.M.; Chen, Y.-F.; Chang, J.; Chiang, Y.-H.; Kerris, R.J.; Chang, C.; Schaller, G.E. Heteromeric interactions among ethylene receptors mediate signaling in Arabidopsis. J. Biol. Chem. 2008, 283, 23801–23810. [Google Scholar] [CrossRef] [Green Version]
- Ho, Y.S.; Burden, L.M.; Hurley, J.H. Structure of the GAF domain, a ubiquitous signaling motif and a new class of cyclic GMP receptor. EMBO J. 2000, 19, 5288–5299. [Google Scholar] [CrossRef] [Green Version]
- Binder, B.M.; Rodriguez, F.I.; Bleecker, A.B. The copper transporter RAN1 is essential for biogenesis of ethylene receptors in Arabidopsis. J. Biol. Chem. 2010, 285, 37263–37270. [Google Scholar] [CrossRef] [Green Version]
- Schott-Verdugo, S.; Müller, L.; Classen, E.; Gohlke, H.; Groth, G. Structural model of the ETR1 Ethylene Receptor Transmembrane Sensor Domain. Sci. Rep. 2019, 9, 8869. [Google Scholar] [CrossRef] [Green Version]
- Azhar, B.J.; Abbas, S.; Aman, S.; Yamburenko, M.V.; Chen, W.; Müller, L.; Uzun, B.; Jewell, D.A.; Dong, J.; Shakeel, S.N.; et al. Basis for high-affinity ethylene binding by the ethylene receptor ETR1 of Arabidopsis. Proc. Natl. Acad. Sci. USA 2023, 120, e2215195120. [Google Scholar] [CrossRef]
- Matoo, K.; Baker, J.E.; Moline, H.E. Induction by Copper Ions of Ethylene Production in Spirodela oligorrhiza: Evidence for a Pathway Independent of 1-Aminocyclopropane-1-carboxylic Acid. J. Plant Physiol. 1983, 123, 193–202. [Google Scholar] [CrossRef]
- Roman, G.; Lubarsky, B.; Kieber, J.J.; Rothenberg, M.; Ecker, J.R. Genetic analysis of ethylene signal transduction in Arabidopsis thaliana: Five novel mutant loci integrated into a stress response pathway. Genetics 1995, 139, 1393–1409. [Google Scholar] [CrossRef] [PubMed]
- Clark, K.L.; Larsen, P.B.; Wang, X.; Chang, C. Association of the Arabidopsis CTR1 Raf-like kinase with the ETR1 and ERS ethylene receptors. Proc. Natl. Acad. Sci. USA 1998, 95, 5401–5406. [Google Scholar] [CrossRef]
- Cancel, J.D.; Larsen, P.B. Loss-of-function mutations in the ethylene receptor ETR1 cause enhanced sensitivity and exaggerated response to ethylene in Arabidopsis. Plant Physiol. 2002, 129, 1557–1567. [Google Scholar] [CrossRef] [Green Version]
- Gao, Z.; Chen, Y.F.; Randlett, M.D.; Zhao, X.C.; Findell, J.L.; Kieber, J.J.; Schaller, G.E. Localization of the Raf-like kinase CTR1 to the endoplasmic reticulum of Arabidopsis through participation in ethylene receptor signaling complexes. J. Biol. Chem. 2003, 278, 34725–34732. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Li, H.; Hutchison, C.E.; Laskey, J.; Kieber, J.J. Biochemical and functional analysis of CTR1, a protein kinase that negatively regulates ethylene signaling in Arabidopsis. Plant J. 2003, 33, 221–233. [Google Scholar] [CrossRef]
- Alonso, J.M.; Hirayama, T.; Roman, G.; Nourizadeh, S.; Ecker, J.R. EIN2, a bifunctional transducer of ethylene and stress responses in Arabidopsis. Science 1999, 284, 2148–2152. [Google Scholar] [CrossRef]
- Bisson, M.M.; Bleckmann, A.; Allekotte, S.; Groth, G. EIN2, the central regulator of ethylene signaling, is localized at the ER membrane where it interacts with the ethylene receptor ETR1. Biochem. J. 2009, 23, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Ju, C.; Van de Poel, B.; Cooper, E.D.; Thierer, J.H.; Gibbons, T.R.; Delwiche, C.F.; Chang, C. Conservation of ethylene as a plant hormone over 450 million years of evolution. Nat. Plants 2015, 1, 14004. [Google Scholar] [CrossRef]
- Qiao, H.; Shen, Z.; Huang, S.S.; Schmitz, R.J.; Urich, M.A.; Briggs, S.P.; Ecker, J.R. Processing and subcellular trafficking of ER-tethered EIN2 control response to ethylene gas. Science 2012, 338, 390–393. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Ma, M.; Feng, Y.; Li, H.; Wang, Y.; Ma, Y.; Li, M.; An, F.; Guo, H. EIN2-directed translational regulation of ethylene signaling in Arabidopsis. Cell 2015, 163, 670–683. [Google Scholar] [CrossRef] [Green Version]
- Dolgikh, V.A.; Pukhovaya, E.M.; Zemlyanskaya, E.V. Shaping ethylene response: The role of EIN3/EIL1 Transcription Factors. Front. Plant Sci. 2019, 10, 1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, S.L.; Rahman, A.; Baskin, T.I.; Kieber, J.J. Two leucine-rich repeat receptor kinases mediate signaling linking cell wall biosynthesis and ACC synthase in Arabidopsis. Plant Cell 2008, 20, 3065–3079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsang, D.L.; Edmond, C.; Harrington, J.L.; Nuhse, T.S. Cell wall integrity controls root elongation via a general 1-aminocyclopropane-1-carboxylic acid-dependent, ethylene-independent pathway. Plant Physiol. 2011, 156, 596–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanderstraeten, L.; Depaepe, T.; Bertrand, S.; Van Der Straeten, D. The ethylene precursor ACC affects early vegetative development independently of ethylene signaling. Front. Plant Sci. 2019, 10, 1591. [Google Scholar] [CrossRef] [Green Version]
- Van de Poel, B.; Van Der Straeten, D. 1-Aminocyclopropane-1-carboxylic acid (ACC) in plants: More than just the precursor of ethylene! Front. Plant Sci. 2014, 5, 640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mou, W.; Kao, Y.-T.; Michard, E.; Simon, A.A.; Li, D.; Wudick, M.M.; Lizzio, M.A.; Feijo, J.A.; Chang, C. Ethylene-independent signalling by the ethylene precursor ACC in Arabidopsis ovular pollen tube attraction. Nat. Common. 2020, 11, 4082. [Google Scholar] [CrossRef] [PubMed]
- Tsuchisaka, A.; Yu, G.; Jin, H.; Alonso, J.M.; Ecker, J.R.; Zhang, X.; Gao, S.; Theologis, A. A combinatorial interplay among the 1-aminocyclopropane-1-carboxylate isoforms regulates ethylene biosynthesis in Arabidopsis thaliana. Genetics 2009, 183, 979–1003. [Google Scholar] [CrossRef] [Green Version]
- Yin, J.; Zhang, X.; Zhang, G.; Wen, Y.; Liang, G.; Chen, X. Aminocyclopropane-1-carboxylic acid is a key regulator of guard mother cell terminal division in Arabidopsis thaliana. J. Exp. Bot. 2018, 70, 897–908. [Google Scholar] [CrossRef] [Green Version]
- Tsolakidou, M.-D.; Pantelides, L.S.; Tzima, A.K.; Kang, S.; Paplomatas, E.J.; Tsaltas, D. Disruption and overexpression of the gene encoding acc (1-aminocyclopropane-1-carboxylic acid) deaminase in soil-borne fungal pathogen Verticillium dahlia revealed the role of ACC as a potential regulator of virulence and plant defense. Mol. Plant Microbe Interact. 2019, 32, 639–653. [Google Scholar] [CrossRef]
- Bradford, K.J.; Yang, S.F. Xylem transport of 1-aminocyclopropane-1-carboxylic acid, an ethylene precursor, in waterlogged tomato plants. Plant Physiol. 1980, 65, 322–326. [Google Scholar] [CrossRef] [Green Version]
- Else, M.A.; Jackson, M.B. Transport of 1-aminocyclopropane-1-carboxylic acid (ACC) in the transpiration stream of tomato (Lycopersicon esculentum) in relation to foliar ethylene production and petiole epinasty. Aust. J. Plant Physiol. 1998, 25, 453–458. [Google Scholar] [CrossRef]
- Chen, Y.; Bonkowski, M.; Shen, Y.; Griffiths, B.S.; Jiang, Y.; Wang, X.; Sun, B. Root ethylene mediates rhizosphere microbial community reconstruction when chemically detecting cyanide produced by neighbouring plants. Microbiome 2020, 8, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nascimento, F.; Rossi, M.; Glick, B. Ethylene and 1-aminocyclopropane-1-carboxylate (ACC) in plant-bacterial interactions. Front. Plant Sci. 2018, 9, 114. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Mou, W.; Van de Poel, B.; Chang, C. Something old, something new: Conservation of the ethylene precursor 1-amino-cyclopropane-1-carboxylic acid as a signaling molecule. Curr. Opin. Plant Biol. 2022, 65, 102116. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.-J.; Chen, H.-W.; Ma, B.; Zhang, W.-K.; Chen, S.Y.; Zhang, J.-S. The role of ethylene in plants under salinity stress. Front. Plant Sci. 2015, 6, 1059. [Google Scholar] [CrossRef] [Green Version]
- Stearns, J.C.; Glick, B.R. Transgenic plants with altered ethylene biosynthesis or perception. Biotechnol. Adv. 2003, 21, 193–210. [Google Scholar] [CrossRef]
- Timmusk, S.; Paalme, V.; Pavlicek, T.; Bergquist, J.; Vangala, A.; Danilas, T.; Nevo, E. Bacterial distribution in the rhizosphere of wild barley under contrasting microclimates. PLoS ONE 2011, 6, 17968. [Google Scholar] [CrossRef]
- Ekimova, G.A.; Fedorov, D.N.; Tani, A.; Doronina, N.V.; Trotsenko, Y.A. Distribution of 1-aminocyclopropane-1-carboxylate deaminase and d-cysteine desulfhydrase genes among type species of the genus Methylobacterium. Antonie Leeuwenhoek 2018, 111, 1723–1734. [Google Scholar] [CrossRef]
- Reed, M.L.E.; Glick, B.R. Applications of Plant Growth-Promoting Bacteria for Plant and Soil Systems; Gupta, V.K., Schmoll, M., Maki, M., Tuohy, M., Mazutti, M.A., Eds.; CRC Press: Boca Raton, FL, USA, 2013. [Google Scholar]
- Grichko, V.P.; Glick, B.R. Amelioration of flooding stress by ACC deaminase-containing plant growth-promoting bacteria. Plant Physiol. Biochem. 2001, 39, 11–17. [Google Scholar] [CrossRef]
- Glick, B.R.; Penrose, D.M. The use of ACC deaminase-containing plant growth-promoting bacteria to protect plants against the deleterious effects of ethylene. In Plant Surface Microbiology; Springer: Berlin/Heidelberg, Germany, 2004; pp. 133–144. [Google Scholar]
- Glick, B.R. Bacterial ACC deaminase and the alleviation of plant stress. Adv. Appl. Microbiol. 2004, 56, 291–312. [Google Scholar]
- Glick, B.R. Modulation of plant ethylene levels by the enzyme ACC deaminase. FEMS Microbiol. Lett. 2005, 251, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Saleh-Lakha, S.; Hontzeas, N.; Glick, B.R. Physiological and genetic effects of bacterial ACC deaminase on plants. In Microbial Biotechnology in Agriculture and Aquaculture; Science Publishers: Enfield, UK, 2006; Volume II, pp. 199–224. [Google Scholar]
- Gerhardt, K.; Greenberg, B.M.; Glick, B.R. The role of ACC deaminase in facilitating the phytoremediation of organics, metals and salt. Curr. Trends Microbiol. 2007, 2, 61–73. [Google Scholar]
- Cheng, Z.; Park, E.; Glick, B.R. 1-Aminocyclopropane-1-carboxylate (ACC) deaminase from Pseudomonas putida UW4 facilitates the growth of canola in the presence of salt. Can. J. Microbiol. 2007, 53, 912–918. [Google Scholar] [CrossRef]
- Hao, Y.; Charles, T.C.; Glick, B.R. ACC deaminase from plant growth promoting bacteria affects crown gall development. Can. J. Microbiol. 2007, 53, 1291–1299. [Google Scholar] [CrossRef]
- Gamalero, E.; Berta, G.; Massa, N.; Glick, B.R.; Lingua, G. 2008. Synergistic interactions between the ACC deaminase-producing bacterium Pseudomonas putida UW4 and the AM fungus Gigaspora rosea positively affect cucumber plant growth. FEMS Microbiol. Ecol. 2008, 64, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.; Charles, T.C.; Glick, B.R. ACC deaminase increases A. tumefaciens-mediated transformation frequency of commercial canola cultivars. FEMS Microbiol. Lett. 2010, 307, 185–190. [Google Scholar] [CrossRef]
- Toklikishvili, N.; Dandurishvili, N.; Tediashvili, M.; Giorgobiani, N.; Szegedi, E.; Glick, B.R.; Vainstein, A.; Chernin, L. Inhibitory effect of ACC deaminase-producing bacteria on crown gall formation in tomato plants infected by Agrobacterium tumefaciens or A. vitis. Plant Pathol. 2010, 59, 1023–1030. [Google Scholar] [CrossRef]
- Hao, Y.; Charles, T.C.; Glick, B.R. An ACC deaminase containing A. tumefaciens strain D3 shows biocontrol activity to crown gall disease. Can. J. Microbiol. 2011, 57, 278–286. [Google Scholar] [CrossRef]
- Siddikee, M.A.; Glick, B.R.; Chauhan, P.S.; Yim, W.-J.; Sa, T. Enhancement of growth and salt tolerance of red pepper seedlings (Capsicum annuum L.) by regulating stress ethylene synthesis with halotolerant bacteria containing ACC deaminase activity. Plant Physiol. Biochem. 2011, 49, 427–434. [Google Scholar] [CrossRef]
- Chernin, L.; Glick, B.R. The use of ACC deaminase to increase the tolerance of plants to various phytopathogens. In Bacteria in Agrobiology: Stress Management; Maheshwari, D.K., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 279–299. [Google Scholar]
- Ali, S.; Charles, T.C.; Glick, B.R. Delay of carnation flower senescence by bacterial endophytes expressing ACC deaminase. J. Appl. Microbiol. 2012, 113, 1139–1144. [Google Scholar] [CrossRef]
- Bal, H.B.; Nayak, L.; Das, S.; Adhya, T.K. Isolation of ACC deaminase producing PGPR from rice rhizosphere and evaluating their plant growth promoting activity under salt stress. Plant Soil 2013, 366, 93–105. [Google Scholar] [CrossRef]
- Kong, Z.; Glick, B.R.; Duan, J.; Ding, S.; Tian, J.; McConkey, B.; Wei, G. Effects of 1-aminocyclopropane-1-carboxylate (ACC) deaminase-overproducing Sinorhizobium meliloti on plant growth and copper tolerance of Medicago lupulina. Plant Soil 2015, 391, 383–398. [Google Scholar] [CrossRef]
- Singh, R.P.; Shelke, G.M.; Kumar, A.; Jha, P.N. Biochemistry and genetics of ACC deaminase: A weapon to “stress ethylene” produced in plants. Front. Microbiol. 2015, 6, 937. [Google Scholar] [PubMed] [Green Version]
- Mendoza-Hernández, J.C.; Perea-Vélez, Y.S.; Arriola-Morales, J.; Martínez-Simón, S.M.; Pérez-Osorio, G. Assessing the effects of heavy metals in ACC deaminase and IAA production on plant growth-promoting bacteria. Microbiol. Res. 2016, 188–189, 53–61. [Google Scholar]
- Gamalero, E.; Marzachì, C.; Galetto, L.; Veratti, F.; Massa, N.; Bona, E.; Novello, G.; Glick, B.R.; Ali, S.; Cantamessa, S.; et al. An ACC deaminase-expressing endophyte increases plant tolerance to Flavescence dorée phytoplasma infection. Plant Biosys. 2016, 151, 331–340. [Google Scholar] [CrossRef]
- Nascimento, F.X.; Rossi, M.J.; Glick, B.R. Role of ACC deaminase in stress control of leguminous plants. In Plant Growth Promoting Actinobacteria; Subramanian, G., Arumugam, S., Rajendran, V., Eds.; Springer Science: Singapore, 2016; pp. 179–192. [Google Scholar]
- Nascimento, F.X.; Tavares, M.J.; Franck, J.; Ali, S.; Glick, B.R.; Rossi, M.J. ACC deaminase plays a major role in Pseudomonas fluorescens YsS6 ability to promote the nodulation of Alpha- and Betaproteobacteria rhizobial strains. Arch. Microbiol. 2019, 201, 817–822. [Google Scholar] [CrossRef]
- Gupta, S.; Pandey, S. ACC deaminase producing bacteria with multifarious plant growth promoting traits alleviates salinity stress in French bean (Phaseolus vulgaris) plants. Front. Microbiol. 2019, 10, 1506. [Google Scholar] [CrossRef] [PubMed]
- Orozco-Mosqueda, M.C.; Duan, J.; DiBernardo, M.; Zetter, E.; Campos-García, J.; Glick, B.R.; Santoyo, G. The production of ACC deaminase and trehalose by the plant growth promoting bacterium Pseudomonas sp. UW4 synergistically protect tomato plants against salt stress. Front. Microbiol. 2019, 10, 1392. [Google Scholar] [CrossRef] [Green Version]
- Chandra, D.; Srivastava, R.; Glick, B.R.; Sharma, A.K. Rhizobacteria producing ACC deaminase mitigate water-stress response in finger millet (Eleusine coracana (L.) Gaertn.). 3 Biotech 2020, 10, 65. [Google Scholar] [CrossRef]
- Orozco-Mosqueda, M.C.; Glick, B.R.; Santoyo, G. ACC deaminase in plant growth-promoting bacteria (PGPB): An efficient mechanism to counter salt in crops. Microbiol. Res. 2020, 235, 126439. [Google Scholar] [CrossRef]
- Paço, A.; da-Silva, J.R.; Denise Pereira Torres, D.; Glick, B.R.; Brígido, C. Exogenous ACC deaminase is key to improving the performance of pasture legume-rhizobial symbioses in the presence of a high manganese concentration. Plants 2020, 9, 1630. [Google Scholar] [CrossRef]
- Duan, B.; Li, L.; Chen, G.; Su-Zhou, C.; Li, Y.; Merkeryan, H.; Liu, W.; Liu, X. 1-Aminocyclopropane-1-carboxylate deaminase producing plant growth-promoting rhizobacteria improve drought stress tolerance in grapevine (Vitis vinifera L.). Front. Plant Sci. 2021, 12, 706990. [Google Scholar] [CrossRef]
- Nascimento, F.X.; Urón, P.; Glick, B.R.; Giachini, A.; Rossi, M.J. Genomic analysis of the ACC deaminase-producing Pseudomonas thivervalensis SC5 reveals its multifaceted roles in soil and beneficial interactions with plants. Front. Microbiol. 2021, 12, 752288. [Google Scholar] [CrossRef]
- Jacobson, C.B.; Pasternak, J.J.; Glick, B.R. Partial purification and characterization of ACC deaminase from the plant growth-promoting rhizobacterium Pseudomonas putida GR12-2. Can. J. Microbiol. 1994, 40, 1019–1025. [Google Scholar] [CrossRef]
- Honma, M. Chemically reactive sulfhydryl groups of 1-aminocyclopropane-1-carboxylate deaminase. Agric. Biol. Chem. 1985, 49, 567–571. [Google Scholar]
- Ali, S.; Glick, B.R. The biochemistry and molecular biology of the enzyme ACC deaminase. In Microbes: The Foundation Stone of the Biosphere; Hurst, C., Ed.; Springer: Cham, Switzerland, 2021; Chapter 19; pp. 365–390. [Google Scholar]
- Hontzeas, N.; Zoidakis, J.; Glick, B.R.; Abu-Omar, M.M. Expression and characterization of 1-aminocyclopropane-1-carboxylate deaminase from the rhizobacterium Pseudomonas putida UW4: A key enzyme in bacterial plant growth promotion. Biochim. Biophys. Acta 2004, 1703, 11–19. [Google Scholar] [CrossRef]
- Glick, B.R.; Cheng, Z.; Czarny, J.; Duan, J. Promotion of plant growth by ACC deaminase-containing soil bacteria. Eur. J. Plant Pathol. 2007, 119, 329–339. [Google Scholar] [CrossRef]
- Duca, D.; Lorv, J.; Patten, C.L.; Rose, D.; Glick, B.R. Indole-3-acetic acid in plant-microbe interactions. Antonie Leeuwenhoek 2014, 106, 85–125. [Google Scholar] [CrossRef]
- Duca, D.R.; Glick, B.R. Indole-3-acetic acid biosynthesis and its regulation in plant-associated bacteria. Appl. Microbiol. Biotechnol. 2020, 104, 8607–8619. [Google Scholar] [CrossRef]
- Stearns, J.C.; Woody, O.Z.; McConkey, B.J.; Glick, B.R. Effects of bacterial ACC deaminase on Brassica napus gene expression. Mol. Plant-Microbe Interact. 2012, 25, 668–676. [Google Scholar] [CrossRef] [Green Version]
- Peck, S.C.; Kende, H. A gene encoding 1-aminocyclopropane-1-carboxylate (ACC) synthase produces two transcripts: Elucidation of a conserved response. Plant J. 1998, 14, 573–581. [Google Scholar] [CrossRef] [Green Version]
- Majda, M.; Robert, S. The role of auxin in cell wall expansion. Int. J. Mol. Sci. 2018, 9, 951. [Google Scholar] [CrossRef] [Green Version]
- Cronin, J.R.; Pizzarello, S. Enantiomeric excesses in meteoritic amino acids. Science 1997, 14, 951–955. [Google Scholar] [CrossRef]
- Aponte, J.C.; Elsila, J.E.; Hein, J.E.; Dworkin, J.P.; Glavin, D.P.; McLain, H.L.; Parker, E.T.; Cao, T.; Berger, E.L.; Burton, A.S. Analysis of amino acids, hydroxy acids, and amines in CR chondrites. Meteorit. Planet Sci. 2020, 55, 2422–2439. [Google Scholar] [CrossRef] [PubMed]
- Elsila, J.E.; Aponte, J.C.; Blackmond, D.G.; Burton, A.S.; Dworkin, J.P.; Glavin, D.P. Meteoritic amino acids: Diversity in compositions reflects parent body histories. ACS Cent. Sci. 2016, 22, 370–379. [Google Scholar] [CrossRef]
- Ollivaux, C.; Soyez, D.; Toullec, J.Y. Biogenesis of D-amino acid containing peptides/proteins: Where, when and how? J. Pept. Sci. 2014, 20, 595–612. [Google Scholar] [CrossRef] [Green Version]
- Veiga, P.; Piquet, S.; Maisons, A.; Furlan, S.; Courtin, P.; Chapot-Chartier, M.P.; Kulakauskas, S. Identification of an essential gene responsible for D-Asp incorporation in the Lactococcus lactis peptidoglycan crossbridge. Mol. Microbiol. 2006, 62, 1713–1724. [Google Scholar] [CrossRef]
- Bruckner, H.; Westhauser, T. Chromatographic determination of L- and d-amino acids in plants. Amino Acids 2003, 24, 43–55. [Google Scholar] [CrossRef]
- Michard, E.; Lima, P.T.; Borges, F.; Silva, A.C.; Portes, M.T.; Carvalho, J.E.; Gilliham, M.; Liu, L.H.; Obermeyer, G.; Feijo, J.A. Glutamate receptor-like genes form Ca 2+ channels in pollen tubes and are regulated by pistil-serine. Science 2011, 332, 434–437. [Google Scholar] [CrossRef] [Green Version]
- Vranova, V.; Zahradnickova, H.; Janous, D.; Skene, K.R.; Matharu, A.S.; Rejsek, K.; Formanek, P. The significance of d-amino acids in soil, fate and utilization by microbes and plants: Review and identification of knowledge gaps. Plant Soil 2012, 354, 21–39. [Google Scholar] [CrossRef]
- Gordes, D.; Kolukisaoglu, U.; Thurow, K. Uptake and conversion of d-amino acids in Arabidopsis thaliana. Amino Acids 2011, 40, 553–563. [Google Scholar] [CrossRef]
- Serralta-Interian, A.A.; Miranda-Ham, M.D.L.; Echevarría-Machado, I. Stimulation of root growth and enhanced nitrogenous metabolite content in habanero pepper (Capsicum chinense Jacq.) treated with a d-amino acid mixture. Theor. Exp. Plant Physiol. 2020, 32, 31–47. [Google Scholar] [CrossRef]
- Kolukisaoglu, Ü. D-amino acids in plants: Sources, metabolism, and functions. Int. J. Mol. Sci. 2020, 30, 5421. [Google Scholar] [CrossRef] [PubMed]
- Strauch, R.C.; Svedin, E.; Dilkes, B.; Chapple, C.; Li, X. Discovery of a novel amino acid racemase through exploration of natural variation in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2015, 15, 11726–11731. [Google Scholar] [CrossRef] [PubMed]
- Hirner, A.; Ladwig, F.; Stransky, H.; Okumoto, S.; Keinath, M.; Harms, A.; Frommer, W.B.; Koch, W. Arabidopsis LHT1 is a high-affinity transporter for cellular amino acid uptake in both root epidermis and leaf mesophyll. Plant Cell 2006, 18, 1931–1946. [Google Scholar] [CrossRef] [Green Version]
- Kolukisaoglu, Ü.; Suarez, J. D-Amino acids in plants: New insights and aspects, but also more open questions. In Amino Acid-New Insights and Roles in Plant and Animal; Asao, T., Asaduzzaman, M., Eds.; IntechOpen: London, UK, 2017; pp. 155–164. [Google Scholar]
- Lehmann, S.; Gumy, C.; Blatter, E.; Boeffel, S.; Fricke, W.; Rentsch, D. In planta function of compatible solute transporters of the AtProT family. J. Exp. Bot. 2011, 62, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Hener, C.; Hummel, S.; Suarez, J.; Stahl, M.; Kolukisaoglu, Ü. d-Amino Acids Are Exuded by Arabidopsis thaliana Roots to the Rhizosphere. Int. J. Mol. Sci. 2018, 19, 1109. [Google Scholar] [CrossRef] [Green Version]
- Suarez, J.; Hener, C.; Lehnhardt, V.A.; Hummel, S.; Stahl, M.; Kolukisaoglu, Ü. AtDAT1 is a key enzyme of D-Amino Acid stimulated ethylene production in Arabidopsis thaliana. Front. Plant Sci. 2019, 12, 1609. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.G.; Min, X.; Zhou, Z.H. Hydrogen sulfide: A signal molecule in plant cross-adaptation. Front. Plant Sci. 2016, 7, 1621. [Google Scholar] [CrossRef] [Green Version]
- Hirano, T.; Tanidokoro, K.; Shimizu, Y.; Kawarabayasi, Y.; Ohshima, T.; Sato, M.; Tadano, S.; Ishikawa, H.; Takio, S.; Takechi, K.; et al. Moss chloroplasts are surrounded by a peptidoglycan wall containing d-amino acids. Plant Cell. 2016, 28, 1521–1532. [Google Scholar] [CrossRef] [Green Version]
- Takano, H.; Takechi, K. Plastid peptidoglycan. Biochim. Biophys. Acta 2010, 1800, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Li, N.; Kudo, H.; Zhang, Z.; Li, J.; Wang, L.; Zhang, W.; Takechi, K.; Takano, H. Genes sufficient for synthesizing peptidoglycan are retained in Gymnosperm genomes, and MurE from Larix gmelinii can rescue the albino phenotype of Arabidopsis MurE mutation. Plant Cell Physiol. 2017, 1, 587–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riemenschneider, A.; Wegele, R.; Schmidt, A.; Papenbrock, J. Isolation and characterization of a D-cysteine desulfhydrase protein from Arabidopsis thaliana. FEBS J. 2005, 272, 1291–1304. [Google Scholar] [CrossRef]
- Bharath, S.R.; Bisht, S.; Harijan, R.K.; Svithri, H.S.; Murthy, M.R.N. Structural and mutational studies on substrate specificity and catalysis of Salmonella typhimurium D-cysteine desulfhydrase. PLoS ONE 2012, 7, e36267. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, A. A cysteine desulfhydrase from spinach leaves specific for D-cysteine. Z. Pflanzenphysiol. 1982, 107, 301–312. [Google Scholar] [CrossRef]
- Schmidt, A.; Erdle, I. A cysteine desulfhydrase specific for D-cysteine from the green alga Chlorella fusca. Z. Naturforsch. C 1983, 38, 428–435. [Google Scholar] [CrossRef]
- Rennenberg, H. Cysteine desulfhydrase activity in cucurbit plants: Simulation by preincubation with L- and D-cysteine. Phytochemistry 1983, 22, 1557–1560. [Google Scholar] [CrossRef]
- Rennenberg, H.; Arabatzis, N.; Grundel, I. Cysteine desulphydrase activity in higher plants: Evidence for the action of L- and D-cysteine specific enzymes. Phytochemistry 1987, 26, 1583–1589. [Google Scholar] [CrossRef]
- Soutourina, J.; Blanquet, S.; Plateau, P. Role of D-cysteine desulfhydrase in the adaptation of Escherichia coli to D-cysteine. J. Biol. Chem. 2001, 276, 40864–40872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, J.; Jiang, W.; Cheng, Z.; Heikkila, J.J.; Glick, B.R. The complete genome sequence of the plant growth-promoting bacterium Pseudomonas putida UW4. PLoS ONE 2013, 8, e58640. [Google Scholar]
- Moreira, D.; Philippe, H. Sure facts and open questions about the origin and evolution of photosynthetic plastids. Res. Microbiol. 2001, 152, 771–780. [Google Scholar] [CrossRef]
- Chu, K.H.; Qi, J.; Yu, Z.G.; Anh, V. Origin and phylogeny of chloroplasts revealed by a simple correlation analysis of complete genomes. Mol. Biol. Evol. 2004, 21, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Ezpeleta, N.; Brinkmann, H.; Burey, S.C.; Roure, B.; Burger, G.; Löffelhardt, W.; Bohnert, H.J.; Philippe, H.; Lang, B.F. Monophyly of primary photosynthetic eukaryotes: Green plants, red algae, and glaucophytes. Curr. Biol. 2005, 26, 1325–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruhfel, B.R.; Gitzendanner, M.A.; Soltis, P.S.; Soltis, D.E.; Burleigh, J.G. From algae to angiosperms-inferring the phylogeny of green plants (Viridiplantae) from 360 plastid genomes. BMC Evol. Biol. 2014, 14, 23. [Google Scholar] [CrossRef] [Green Version]
- Gitzendanner, M.A.; Soltis, P.S.; Wong, G.K.-S.; Ruhfel, B.R.; Soltis, D.E. Plastid phylogenomic analysis of green plants: A billion years of evolutionary history. Am. J. Bot. 2018, 105, 291–301. [Google Scholar] [CrossRef]
- Sato, N. Are Cyanobacteria an Ancestor of Chloroplasts or Just One of the Gene Donors for Plants and Algae? Genes 2021, 12, 823. [Google Scholar] [CrossRef]
- Burki, F.; Roger, A.J.; Brown, M.W.; Simpson, A.G.B. The new tree of Eukaryotes. Trends Ecol. Evol. 2020, 35, 43–55. [Google Scholar] [CrossRef] [Green Version]
- One Thousand Plant Transcriptomes Initiative. One thousand plant transcriptomes and the phylogenomics of green plants. Nature 2019, 574, 679–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, L.A.; McCourt, R.M. Green algae and the origin of land plants. Am. J. Bot. 2004, 91, 1535–1556. [Google Scholar] [CrossRef] [Green Version]
- Becker, B.; Marin, B. Streptophyte algae and the origin of embryophytes. Ann. Bot. 2009, 103, 999–1004. [Google Scholar] [CrossRef]
- Wickett, N.J.; Mirarab, S.; Nguyen, N.; Warnow, T.; Carpenter, E.; Matasci, N.; Ayyampalayam, S.; Barker, M.S.; Burleigh, J.G.; Gitzendanner, M.A.; et al. Phylotranscriptomic analysis of the origin and early diversification of land plants. Proc. Natl Acad. Sci. USA 2014, 111, E4859–E4868. [Google Scholar] [CrossRef]
- Lacey, R.F.; Binder, B.M. Ethylene regulates the physiology of the cyanobacterium Synechocystis sp. PCC 6803 via an ethylene receptor. Plant Physiol. 2016, 171, 2798–2809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narikawa, R.; Suzuki, F.; Yoshihara, S.; Higashi, K.; Watanabe, A.; Ikeuchi, M. Novel photosensory two-component system (PixA-NixB-NixC) involved in the regulation of positive and negative phototaxis of cyanobacterium Synechocystis sp. PCC 6803. Plant Cell Physiol. 2011, 52, 2114–2224. [Google Scholar] [CrossRef] [PubMed]
- Song, J.-Y.; Cho, H.S.; Cho, J.-I.; Jeon, J.-S.; Lagarias, J.C.; Park, Y.-I. Near-UV cyanobacteriochrome signaling system elicits negative phototaxis in the cyanobacterium Synechocystis sp. PCC 6803. Proc. Natl. Acad. Sci. USA 2011, 108, 10780–10785. [Google Scholar] [CrossRef]
- Ulijasz, A.T.; Cornilescu, G.; von Stetten, D.; Cornilescu, C.; Escobar, F.V.; Zhang, J.; Vierstra, R.D. Cyanochromes are blue/green light photoreversible photoreceptors defined by a stable double cysteine linkage to a phycoviolobilin-type chromophore. J. Biol. Chem. 2009, 284, 29757–29772. [Google Scholar] [CrossRef] [Green Version]
- Lacey, R.F.; Allen, C.J.; Bakshi, A.; Binder, B.M. Ethylene causes transcriptomic changes in Synechocystis during phototaxis. Plant Direct. 2018, 2, 1–16. [Google Scholar] [CrossRef]
- Wang, W.; Esch, J.J.; Shiu, S.H.; Agula, H.; Binder, B.M.; Chang, C.; Patterson, S.E.; Bleecker, A.B. Identification of important regions for ethylene binding and signaling in the transmembrane domain of the ETR1 ethylene receptor of Arabidopsis. Plant Cell 2006, 18, 3429–3442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Jimenez, P.; Robaina, R.R. Effects of ethylene on tetrasporogenesis in Pterocladiella capillacea (Rhodophyta). J. Phycol. 2012, 48, 710–715. [Google Scholar] [CrossRef]
- Maillard, P.; Thepenier, C.; Gudin, C. Determination of an ethylene biosynthesis pathway in the unicellular green-alga, Haematococcus pluvialis—Relationship between growth and ethylene production. J. Appl. Phycol. 1993, 5, 93–98. [Google Scholar] [CrossRef]
- Huang, T.-C.; Chow, T.-J. Ethylene production by blue-green algae. Bot. Bull. Acad. Sin. 1984, 25, 81–86. [Google Scholar]
- Chernys, J.; Kende, H. Ethylene biosynthesis in Regnellidium diphyllum and Marsilea quadrifolia. Planta 1996, 200, 113–118. [Google Scholar] [CrossRef]
- Osborne, D.J.; Walters, J.; Milborrow, B.V.; Norville, A.; Stange, L.M.C. Evidence for a non-ACC ethylene biosynthesis pathway in lower plants. Phytochemistry 1996, 42, 51–60. [Google Scholar] [CrossRef]
- Rohwer, F.; Bopp, M. Ethylene synthesis in moss protonema. J. Plant Physiol. 1985, 117, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Li, F.W.; Brouwer, P.; Carretero-Paulet, L.; Cheng, S.; de Vries, J.; Delaux, P.M.; Eily, A.; Koppers, N.; Kuo, L.Y.; Li, Z.; et al. Fern genomes elucidate land plant evolution and cyanobacterial symbioses. Nat. Plants 2018, 4, 460–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Overview of the biosynthetic pathway for the synthesis of ACC and ethylene in plants. Abbreviations: MTA, 5′-methylthioadenosine; M-ACC, 1-(malonyl)-ACC; G-ACC, 1-(glutamyl)-ACC. The enzymes catalyzing some of these reactions are shown to the left of the arrow indicating a catalyzed reaction.
Figure 1.
Overview of the biosynthetic pathway for the synthesis of ACC and ethylene in plants. Abbreviations: MTA, 5′-methylthioadenosine; M-ACC, 1-(malonyl)-ACC; G-ACC, 1-(glutamyl)-ACC. The enzymes catalyzing some of these reactions are shown to the left of the arrow indicating a catalyzed reaction.

Figure 2.
When ethylene is absent (Case A), the receptors located in the endoplasmic reticulum membrane (in the diagram this is represented by ETHYLENE RESPONSE 1, ETR1) repress downstream ethylene responses. A serine–threonine protein kinase called CONSTITUTIVE TRIPLE RESPONSE 1 (CTR1) phosphorylates EIN2 (ETHYLENE-INSENSITIVE 2) protein at the C terminal. In this way, EIN2 becomes targeted for degradation. In the nucleus, proteins EBF1 (EIN3 BINDING F-BOX1) and EBF2 (EIN3 BINDING F-BOX2) cooperate to activate the degradation of two transcription factors: EIN3 (ETHYLENE-INSENSITIVE 3) and EIL1 (ETHYLENE-INSENSITIVE 3–like 1). Altogether, these steps lead to inhibition of downstream ethylene signaling.
Figure 2.
When ethylene is absent (Case A), the receptors located in the endoplasmic reticulum membrane (in the diagram this is represented by ETHYLENE RESPONSE 1, ETR1) repress downstream ethylene responses. A serine–threonine protein kinase called CONSTITUTIVE TRIPLE RESPONSE 1 (CTR1) phosphorylates EIN2 (ETHYLENE-INSENSITIVE 2) protein at the C terminal. In this way, EIN2 becomes targeted for degradation. In the nucleus, proteins EBF1 (EIN3 BINDING F-BOX1) and EBF2 (EIN3 BINDING F-BOX2) cooperate to activate the degradation of two transcription factors: EIN3 (ETHYLENE-INSENSITIVE 3) and EIL1 (ETHYLENE-INSENSITIVE 3–like 1). Altogether, these steps lead to inhibition of downstream ethylene signaling.

Figure 3.
The cleavage of ACC by the enzyme ACC deaminase.

Figure 4.
Model of ACC deaminase promoting plant growth.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gamalero, E.; Lingua, G.; Glick, B.R. Ethylene, ACC, and the Plant Growth-Promoting Enzyme ACC Deaminase. Biology 2023, 12, 1043. https://0-doi-org.brum.beds.ac.uk/10.3390/biology12081043
AMA Style
Gamalero E, Lingua G, Glick BR. Ethylene, ACC, and the Plant Growth-Promoting Enzyme ACC Deaminase. Biology. 2023; 12(8):1043. https://0-doi-org.brum.beds.ac.uk/10.3390/biology12081043
Chicago/Turabian StyleGamalero, Elisa, Guido Lingua, and Bernard R. Glick. 2023. "Ethylene, ACC, and the Plant Growth-Promoting Enzyme ACC Deaminase" Biology 12, no. 8: 1043. https://0-doi-org.brum.beds.ac.uk/10.3390/biology12081043
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.