Curcumin Regulates Anti-Inflammatory Responses by JAK/STAT/SOCS Signaling Pathway in BV-2 Microglial Cells

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Cultures and Treatments
2.2. Electrophoresis
2.3. Western Blotting
2.4. ELISA
2.5. Reverse Transcriptase–Polymerase Chain Reaction (RT-PCR) and Quantitative Real-Time PCR Analysis
2.6. Treatment of Cell Cultures with Small Interfering RNA
2.7. Statistical Analysis
3. Results
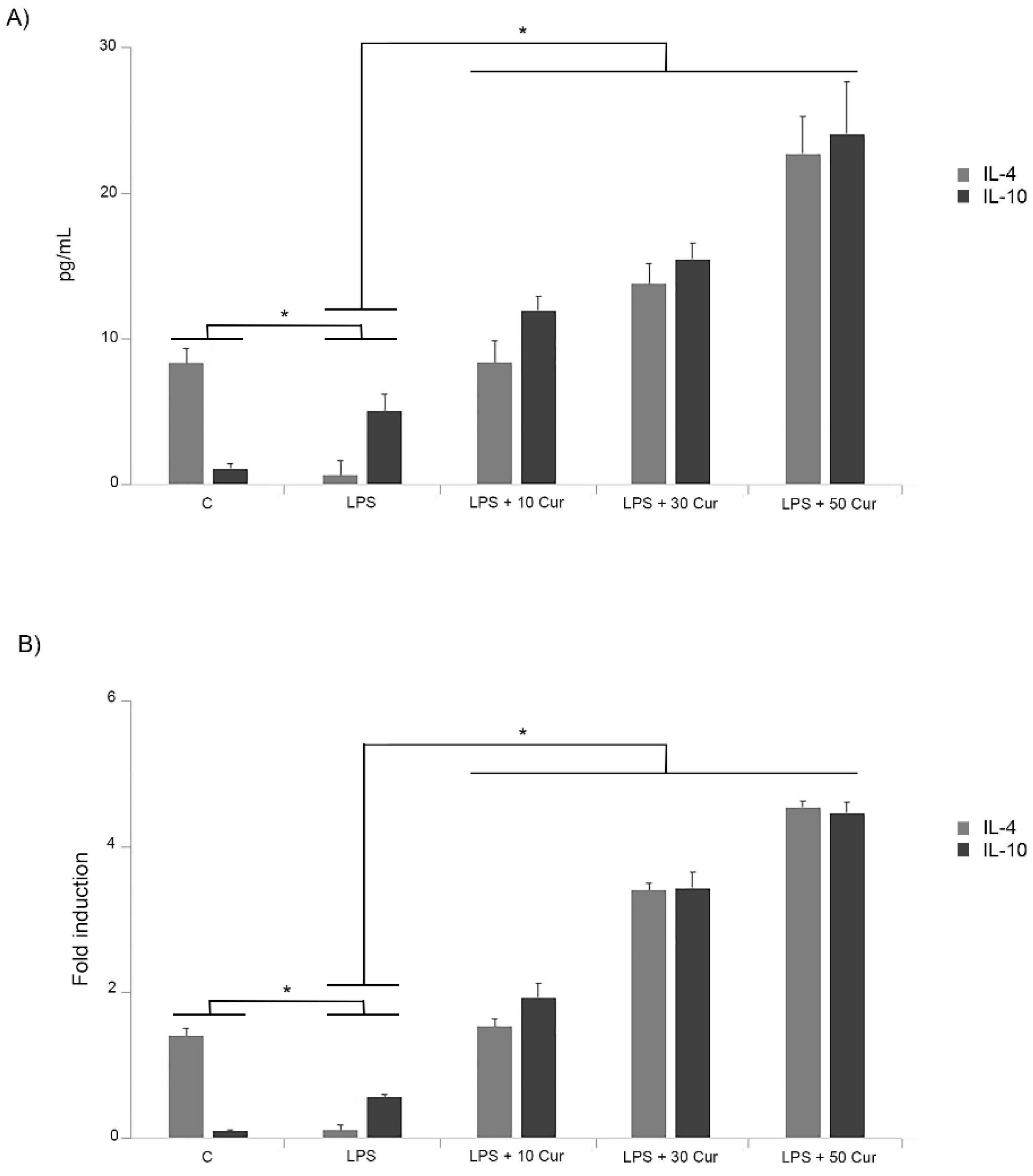
3.1. Curcumin Regulated Pro-Inflammatory Cytokine Expression in LPS-Treated Cells
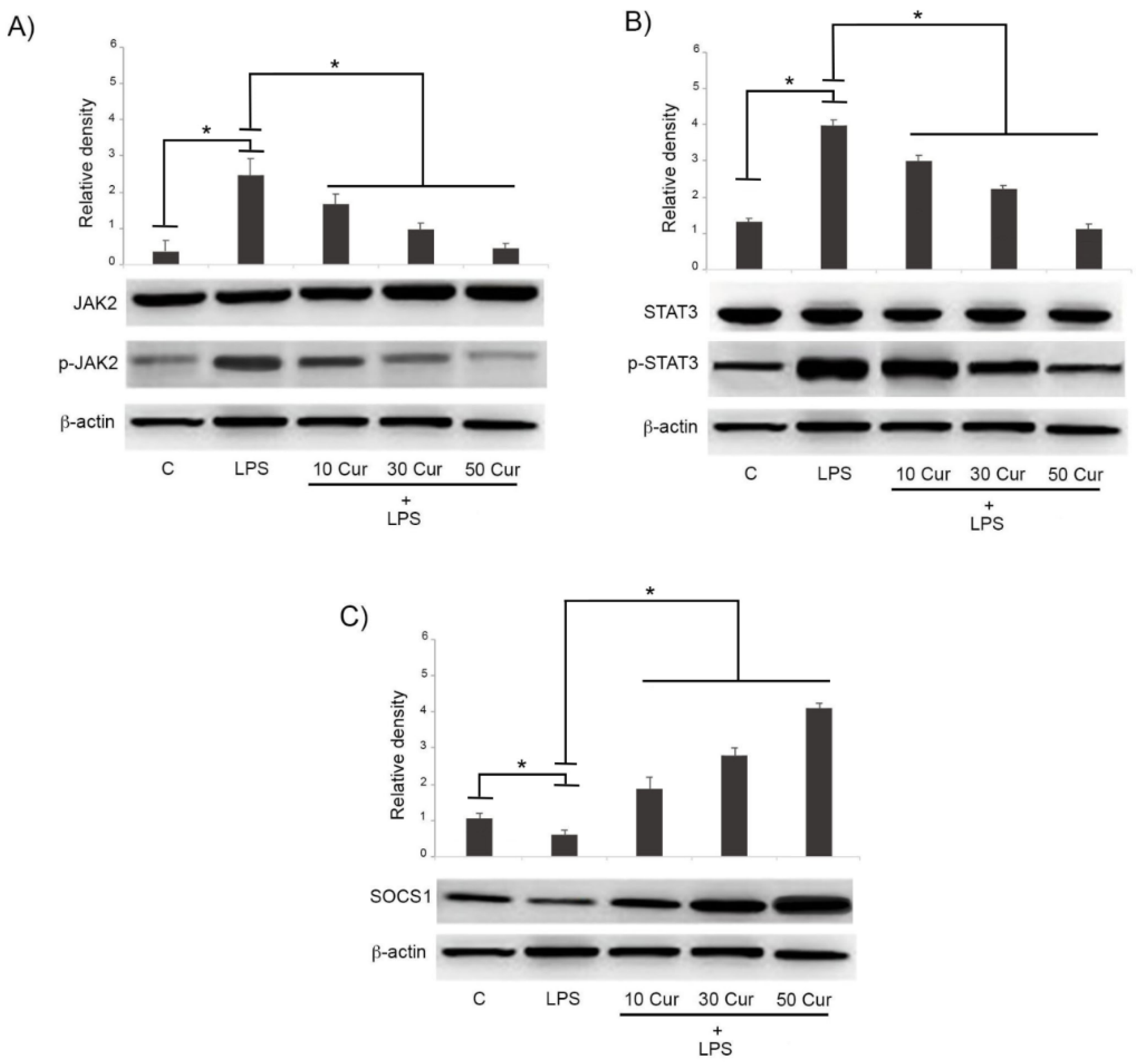
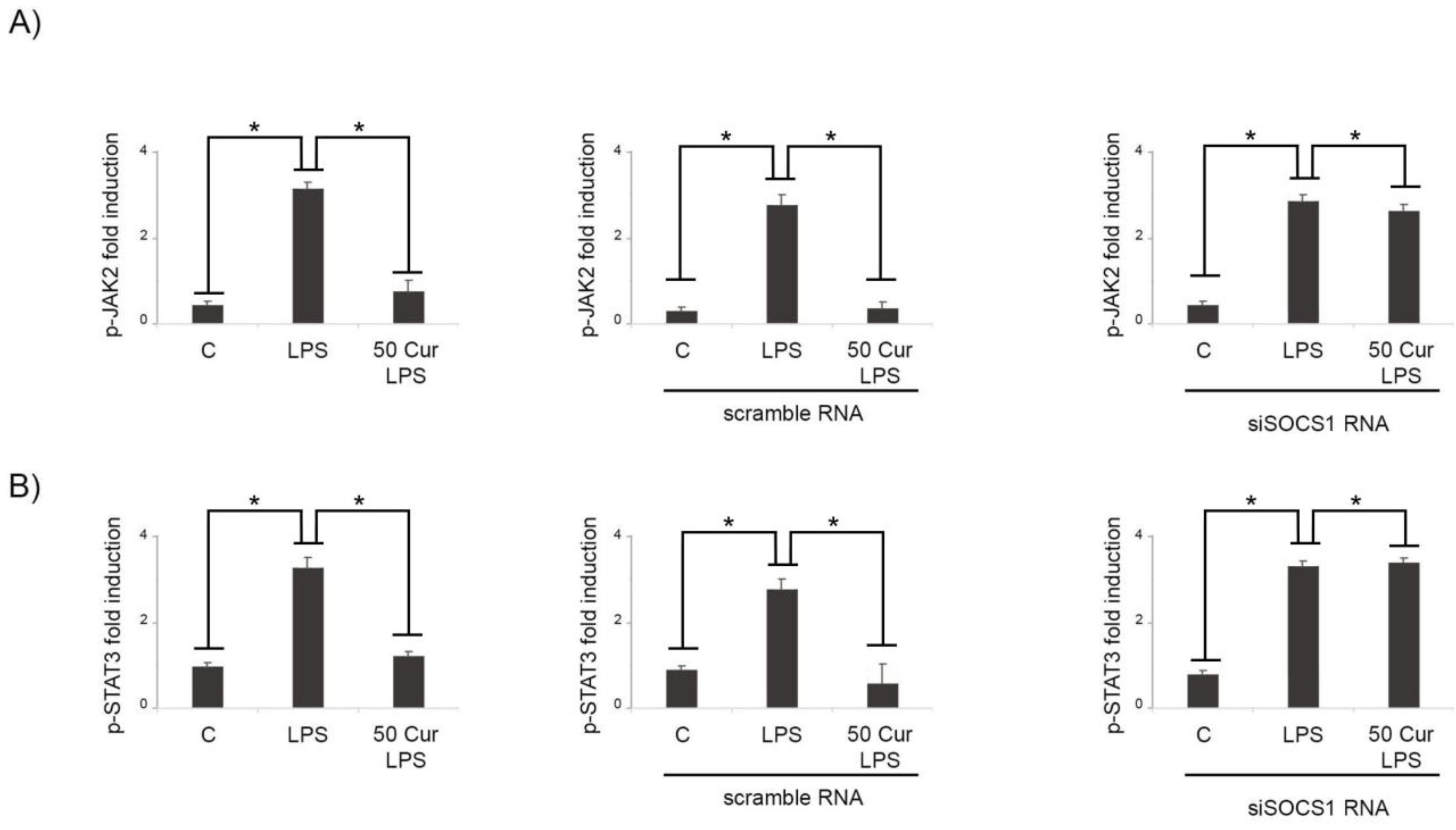
3.2. Effect of Curcumin on the Signaling Pathway of JAK/STAT/SOCS
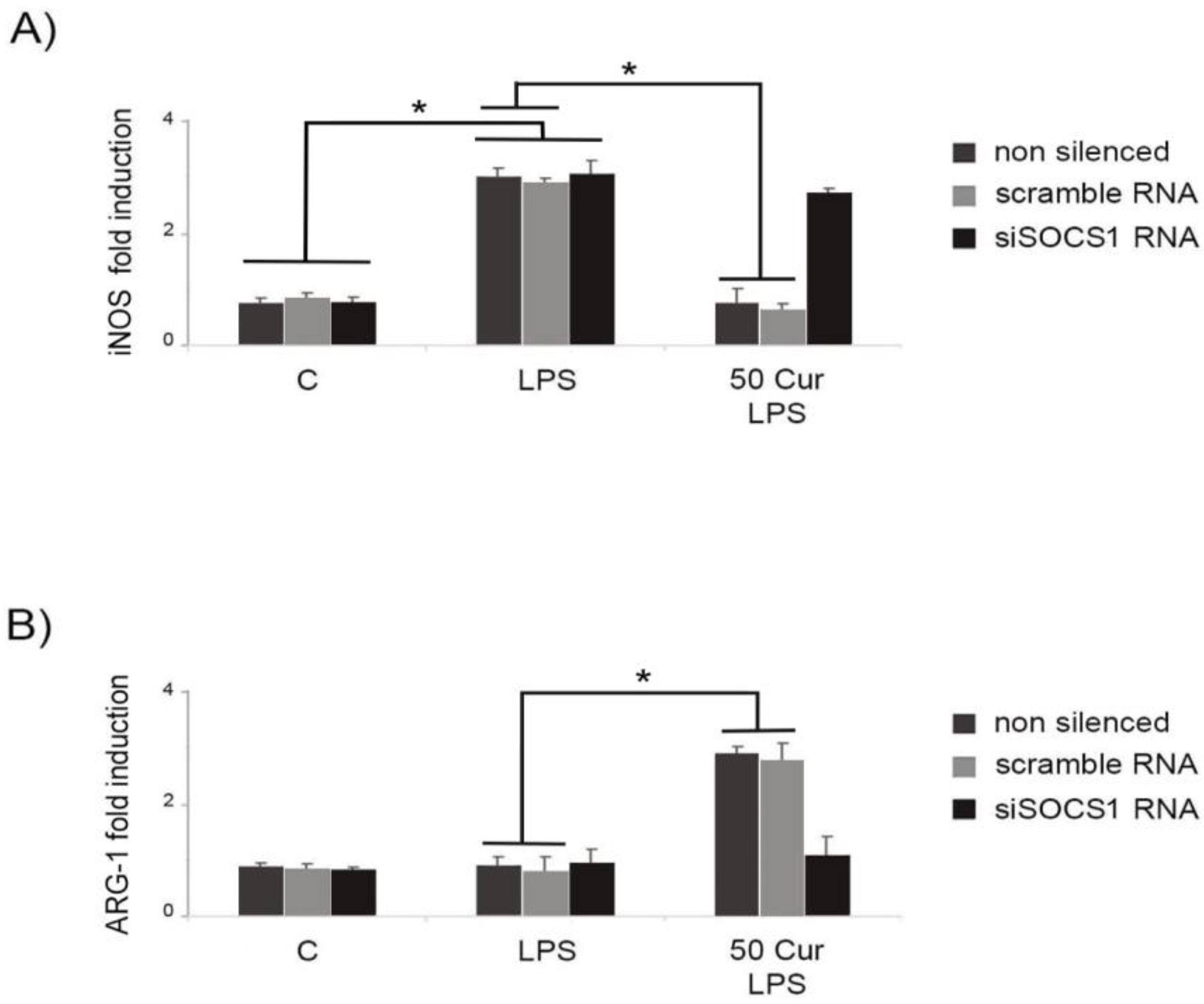
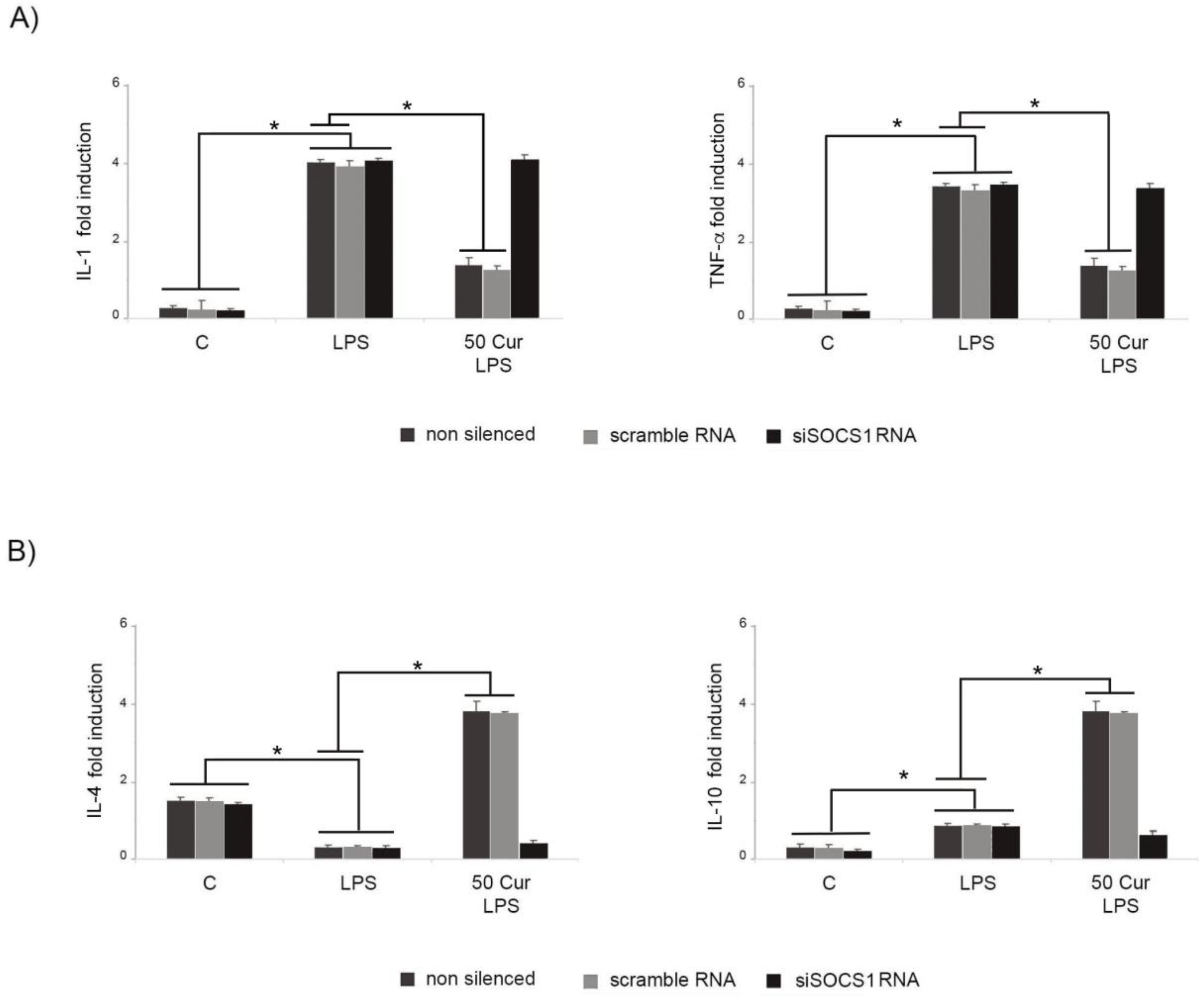
3.3. Influence of SOCS-1 Downregulation on Microglial Polarization
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lue, L.F.; Kuo, Y.M.; Beach, T.; Walker, D.G. Microglia activation and anti-inflammatory regulation in Alzheimer’s disease. Mol. Neurobiol. 2010, 41, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M.A. The impact of neuroimmune changes on development of amyloid pathology; relevance to Alzheimer’s disease. Immunology 2014, 141, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Kierdorf, K.; Prinz, M. Factors regulating microglia activation. Front. Cell Neurosci. 2013, 7, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kile, B.T.; Alexander, W.S. The suppressors of cytokine signalling (SOCS). Cell. Mol. Life. Sci. 2001, 58, 1627–1635. [Google Scholar] [CrossRef] [PubMed]
- Łabuzek, K.; Suchy, D.; Gabryel, B.; Pierzchała, O.; Okopień, B. Role of the SOCS in monocytes/macrophages-related pathologies. Are we getting closer to a new pharmacological target? Pharmacol. Rep. 2012, 64, 1038–1054. [Google Scholar]
- Aggarwal, B.B.; Sung, B. Pharmacological basis for the role of curcumin in chronic diseases: An age-old spice with modern targets. Trends Pharmacol. Sci. 2009, 30, 85–94. [Google Scholar] [CrossRef]
- Perrone, D.; Ardito, F.; Giannatempo, G.; Dioguardi, M.; Troiano, G.; Lo Russo, L.; De Lillo, A.; Laino, L.; Lo Muzio, L. Biological and therapeutic activities, and anticancer properties of curcumin. Exp. Ther. Med. 2015, 10, 1615–1623. [Google Scholar] [CrossRef] [Green Version]
- Amalraj, A.; Pius, A.; Gopi, S.; Gopi, S. Biological activities of curcuminoids, other biomolecules from turmeric and their derivatives—A review. J. Tradit. Complement. Med. 2016, 7, 205–233. [Google Scholar] [CrossRef]
- Cianciulli, A.; Calvello, R.; Porro, C.; Trotta, T.; Salvatore, R.; Panaro, M.A. PI3k/Akt signalling pathway plays a crucial role in the anti-inflammatory effects of curcumin in LPS-activated microglia. Int. Immunopharmacol. 2016, 36, 282–290. [Google Scholar] [CrossRef]
- Tajiri, K.; Imanaka‑Yoshida, K.; Matsubara, A.; Tsujimura, Y.; Hiroe, M.; Naka, T.; Shimojo, N.; Sakai, S.; Aonuma, K.; Yasutomi, Y. Suppressor of cytokine signaling 1 DNA administration inhibits inflammatory and pathogenic responses in autoimmune myocarditis. J. Immunol. 2012, 189, 2043–2053. [Google Scholar] [CrossRef]
- Hashimoto, M.; Ayada, T.; Kinjyo, I.; Hiwatashi, K.; Yoshida, H.; Okada, Y.; Kobayashi, T.; Yoshimura, A. Silencing of SOCS1 in macrophages suppresses tumor development by enhancing antitumor inflammation. Cancer Sci. 2009, 100, 730–736. [Google Scholar] [CrossRef] [PubMed]
- Capello, D.; Gloghini, A.; Baldanzi, G.; Martini, M.; Deambrogi, C.; Lucioni, M.; Piranda, D.; Famà, R.; Grazianim, A.; Spina, M.; et al. Alterations of negative regulators of cytokine signalling in immunodeficiency-related non-Hodgkin lymphoma. Hematol. Oncol. 2013, 31, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Souma, Y.; Nishida, T.; Serada, S.; Iwahori, K.; Takahashi, T.; Fujimoto, M.; Ripley, B.; Nakajima, K.; Miyazaki, Y.; Mori, M.; et al. Antiproliferative effect of SOCS-1 through the suppression of STAT3 and p38 MAPK activation in gastric cancer cells. Int. J. Cancer 2012, 131, 1287–1296. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.Y.; Park, E.J.; Joe, E.H.; Jou, I. Curcumin suppresses Janus kinase-STAT inflammatory signaling through activation of Src homology 2 domain-containing tyrosine phosphatase 2 in brain microglia. J. Immunol. 2003, 171, 6072–6079. [Google Scholar] [CrossRef] [PubMed]
- Shuai, K.; Liu, B. Regulation of JAK-STAT signalling in the immune system. Nat. Rev. Immunol. 2003, 3, 900–911. [Google Scholar] [CrossRef]
- Zhang, X.; Wu, J.; Ye, B.; Wang, Q.; Xie, X.; Shen, H. Protective effect of curcumin on TNBS-induced intestinal inflammation is mediated through the JAK/STAT pathway. BMC Complement Altern. Med. 2016, 16, 299. [Google Scholar] [CrossRef]
- Kinjyo, I.; Hanada, T.; Inagaki-Ohara, K.; Mori, H.; Aki, D.; Ohishi, M.; Yoshida, H.; Kubo, M.; Yoshimura, A. SOCS1/JAB is a negative regulator of LPS-induced macrophage activation. Immunity 2002, 17, 583–591. [Google Scholar] [CrossRef]
- Li, Y.; Chu, N.; Rostami, A.; Zhang, G.X. Dendritic cells transduced with SOCS-3 exhibit a tolerogenic/DC2 phenotype that directs type 2 Th cell differentiation in vitro and in vivo. J. Immunol. 2006, 177, 1679–1688. [Google Scholar] [CrossRef]
- Chainani-Wu, N. Safety and anti-inflammatory activity of curcumin: A component of tumeric (Curcumalonga). J. Altern. Complement. Med. 2003, 9, 161–168. [Google Scholar] [CrossRef]
- Zhang, D.M.; Li, Y.C.; Xu, D.; Ding, X.Q.; Kong, L.D. Protection of curcumin against fructose-induced hyperuricaemia andrenal endothelial dysfunction involves NO-mediated JAK-STAT signalling in rats. Food Chem. 2012, 134, 2184–2193. [Google Scholar] [CrossRef]
- Liu, L.; Liu, Y.L.; Liu, G.X.; Chen, X.; Yang, K.; Yang, Y.X.; Xie, Q.; Gan, H.K.; Huang, X.L.; Gan, H.T. Curcumin ameliorates dextran sulphate sodium-induced experimental colitis by blocking STAT3 signaling pathway. Int. Immunopharmacol. 2013, 17, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Trotta, T.; Panaro, M.A.; Prifti, E.; Porro, C. Modulation of biological activities in glioblastoma mediated by Curcumin. Nutr. Cancer 2019. [Google Scholar] [CrossRef] [PubMed]
- Duan, W.; Yang, Y.; Yan, J.; Yu, S.; Liu, J.; Zhou, J.; Zhang, J.; Jin, Z.; Yi, D. The effects of curcumin post-treatment against myocardial ischemia and reperfusion by activation of the JAK2/STAT3 signaling pathway. Basic Res. Cardiol. 2012, 107, 263. [Google Scholar] [CrossRef] [PubMed]
- Nelms, K.; Keegan, A.D.; Zamorano, J.; Ryan, J.J.; Paul, W.E. The IL-4 receptor: Signaling mechanisms and biologic functions. Annu. Rev. Immunol. 1999, 17, 701–738. [Google Scholar] [CrossRef] [PubMed]
- Butovsky, O.; Ziv, Y.; Schwartz, A.; Landa, G.; Talpalar, A.E.; Pluchino, S.; Martino, G.; Schwartz, M. Microglia activated by IL-4 or IFN-gamma differentially induce neurogenesis and oligodendrogenesis from adult stem/progenitor cells. Mol. Cell. Neurosci. 2006, 31, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Van der Poll, T.; Jansen, J.; Levi, M.; ten Cate, H.; ten Cate, J.W.; van Deventer, S.J. Regulation of interleukin 10 release by tumor necrosis factor in humans and chimpanzees. J. Exp. Med. 1994, 180, 1985–1988. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Wu, P.; Siegel, M.I.; Egan, R.W.; Billah, M.M. IL-10 inhibits transcription of cytokine genes in human peripheral blood mononuclear cells. J. Immunol. 1994, 153, 811–816. [Google Scholar] [PubMed]
- Gui, B.; Su, M.; Chen, J.; Jin, L.; Wan, R.; Qian, Y. Neuroprotective effects of pretreatment with propofol in LPS-induced BV-2 microglia cells: Role of TLR4 and GSK-3β. Inflammation 2012, 35, 1632–1640. [Google Scholar] [CrossRef] [PubMed]
- Bozic, I.; Savic, D.; Laketa, D.; Bjelobaba, I.; Milenkovic, I.; Pekovic, S.; Nedeljkovic, N.; Lavrnja, I. Benfotiamine attenuates inflammatory response in LPS stimulated BV-2 microglia. PLoS ONE 2015, 10, e0118372. [Google Scholar] [CrossRef]
- Chao, C.C.; Molitor, T.W.; Hu, S. Neuroprotective role of IL-4 against activated microglia. J. Immunol. 1993, 151, 1473–1481. [Google Scholar]
- Dooley, D.; Vidal, P.; Hendrix, S. Immunopharmacological intervention for successful neural stem cell therapy: New perspectives in CNS neurogenesis and repair. Pharmacol. Ther. 2014, 141, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Syed Hussein, S.S.; Kamarudin, M.N.; Kadir, H.A. (+)-Catechin Attenuates NF-κB Activation Through Regulation of Akt, MAPK, and AMPK Signaling Pathways in LPS-Induced BV-2 Microglial Cells. Am. J. Chin. Med. 2015, 43, 927–952. [Google Scholar] [CrossRef] [PubMed]
- Downer, E.J.; Cowley, T.R.; Lyons, A.; Mills, K.H.; Berezin, V.; Bock, E.; Lynch, M.A. A novel anti-inflammatory role of NCAM-derived mimetic peptide, FGL. Neurobiol. Aging 2010, 31, 118–128. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Park, H.J.; Starkweather, A.; An, K.; Shim, I. Decreased Interleukin-4 Release from the Neurons of the Locus Coeruleus in Response to Immobilization Stress. Mediators Inflamm. 2016, 2016, 3501905. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Chen, L.Y.; Papadimos, T.J.; Huang, S.; Zuraw, B.L.; Pan, Z.K. Lipopolysaccharide-driven Th2 cytokine production in macrophages is regulated by both MyD88 and TRAM. J. Biol. Chem. 2009, 284, 29391–29398. [Google Scholar] [CrossRef]
- Wilson, H.M. SOCS Proteins in Macrophage Polarization and Function. Front. Immunol. 2014, 5, 357. [Google Scholar] [CrossRef] [Green Version]
- Whyte, C.S.; Bishop, E.T.; Rückerl, D.; Gaspar-Pereira, S.; Barker, R.N.; Allen, J.E.; Rees, A.J.; Wilson, H.M. Suppressor of cytokine signaling (SOCS)1 is a key determinant of differential macrophage activation and function. J. Leukoc. Biol. 2011, 90, 845–854. [Google Scholar] [CrossRef]
- Liu, Y.; Stewart, K.N.; Bishop, E.; Marek, C.J.; Kluth, D.C.; Rees, A.J.; Wilson, H.M. Unique expression of suppressor of cytokine signaling 3 is essential for classical macrophage activation in rodents in vitro and in vivo. J. Immunol. 2008, 180, 6270–6278. [Google Scholar] [CrossRef]
- Cai, H.; Liang, Q.; Ge, G. Gypenoside Attenuates β Amyloid-Induced Inflammation in N9 Microglial Cells via SOCS1 Signaling. Neural Plast. 2016, 2016, 6362707. [Google Scholar] [CrossRef]
- Velmurugan, B.K.; Rathinasamy, B.; Lohanathan, B.P.; Thiyagarajan, V.; Weng, C.F. Neuroprotective Role of Phytochemicals. Molecules 2018, 23, 2485. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence 5’→ 3’ | Sequence Reference |
|---|---|---|
| IL-4 | Fw-ATCATCGGCATTTTGAACGAGGTC Rw-ACCTTGGAAGCCCTACAGACGA | NM_021283.2 |
| IL-10 | Fw-GCCAGTACAGCCGGGAAGACAATA Rw- GCCTTGTAGACACCTTGGTCTT | NM_012854.2 |
| TNF-α | Fw- GGCAGGTCTACTTTGGAGTCATTGC Rw- ACATTCGAGGCTCCAGTGAATTCGG | NM_013693.2 |
| IL-1β | Fw- GCAGCAGCACATCAACAAGAGC Rw- TGTCCTCATCCTGGAAGGTCCACG | NM_008361.2 |
| SOCS-1 | Fw- TGGGCACCTTCTTGGTGCGC Rw- GGCAGTCGAAGGTCTCGCGG | BC_132368.1 |
| iNOS | Fw- CAACAGGGAGAAAGCGCAAA Rw- TGATGGACCCCAAGCAAGAC | NM_001313921.1 |
| Arg-1 | Fw- TTTCAGGACTAGATATCATGGAAGTG Rw- CTTAGGTGGTTTAAGGTAGTCAGTCC | U_51805.1 |
| β-actin | Fw- GACCTCTATGCCAACACAGT Rw- AGTACTTGCGCTCAGGAGGA | NM_007393.5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Porro, C.; Cianciulli, A.; Trotta, T.; Lofrumento, D.D.; Panaro, M.A. Curcumin Regulates Anti-Inflammatory Responses by JAK/STAT/SOCS Signaling Pathway in BV-2 Microglial Cells. Biology 2019, 8, 51. https://0-doi-org.brum.beds.ac.uk/10.3390/biology8030051
Porro C, Cianciulli A, Trotta T, Lofrumento DD, Panaro MA. Curcumin Regulates Anti-Inflammatory Responses by JAK/STAT/SOCS Signaling Pathway in BV-2 Microglial Cells. Biology. 2019; 8(3):51. https://0-doi-org.brum.beds.ac.uk/10.3390/biology8030051
Chicago/Turabian StylePorro, Chiara, Antonia Cianciulli, Teresa Trotta, Dario Domenico Lofrumento, and Maria Antonietta Panaro. 2019. "Curcumin Regulates Anti-Inflammatory Responses by JAK/STAT/SOCS Signaling Pathway in BV-2 Microglial Cells" Biology 8, no. 3: 51. https://0-doi-org.brum.beds.ac.uk/10.3390/biology8030051