Advances in Omics Approaches for Abiotic Stress Tolerance in Tomato

 , , and
, , and
Abstract
:1. Introduction
2. Tomato Genomics for Abiotic Stress Tolerance
Whole Genome Sequencing and Resequencing
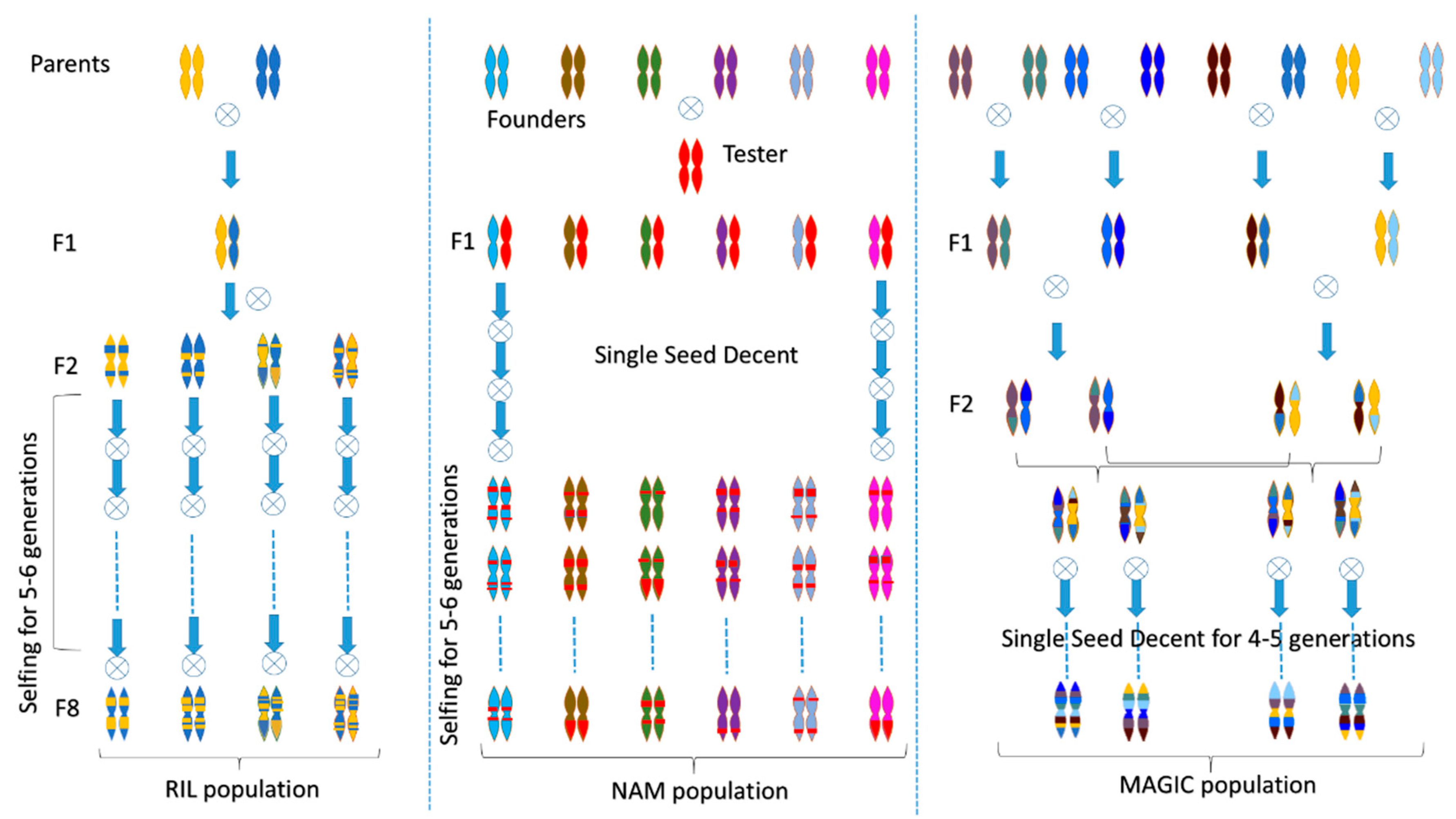
3. Molecular Markers Resources in Tomato
4. Identification of Loci Governing Abiotic Stress through QTL Mapping and GWAS
5. Genomic Selection (GS) for Abiotic Stress in Tomato
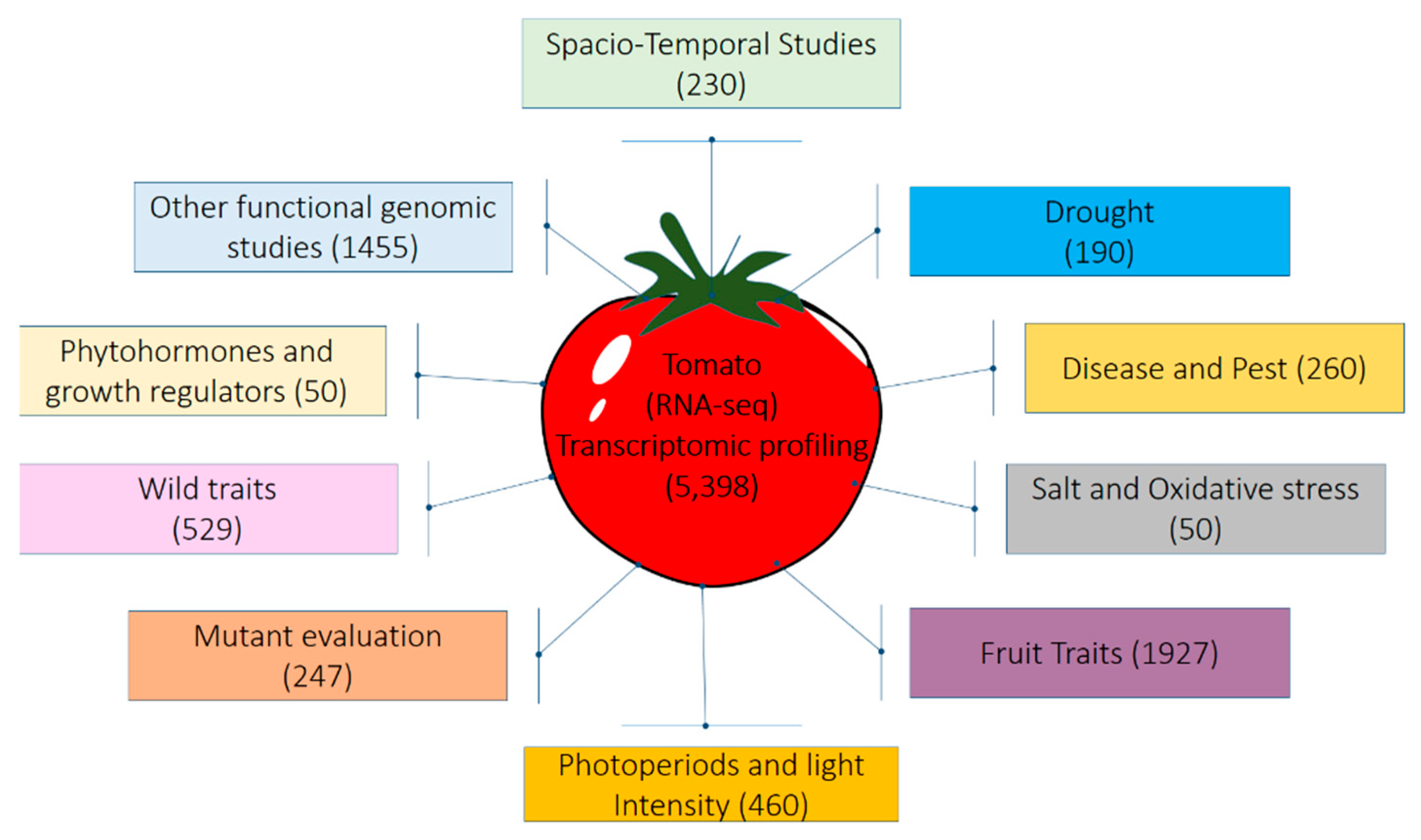
6. Advances in Transcriptomics
7. Tomato Proteomics: Applicability and Challenges
8. Tomato Metabolomics
9. Tomato Ionomics
10. Phenomic Advances for Abiotic Stress Tolerance in Tomato
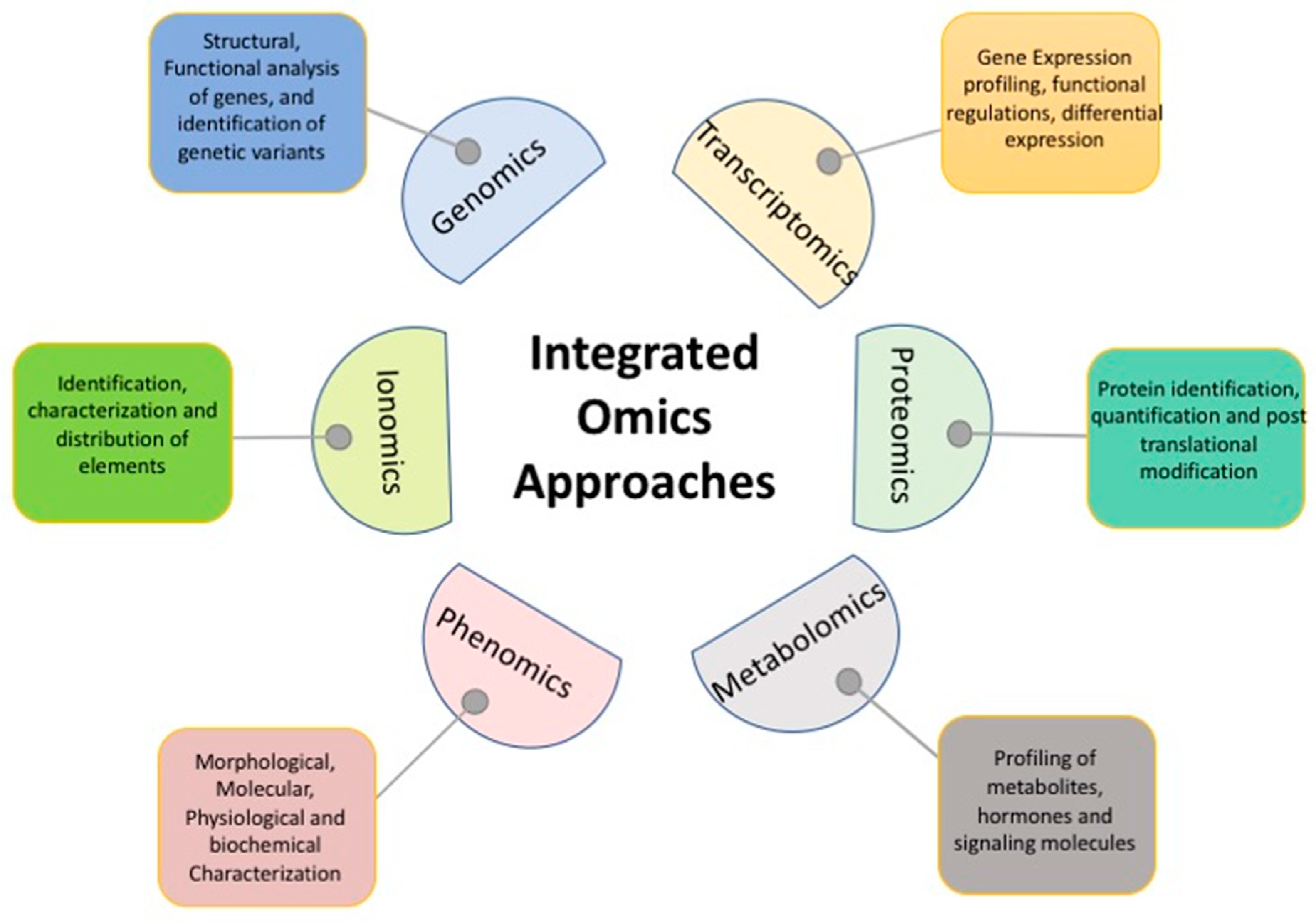
11. Integration of Omics Technologies
12. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chaudhary, J.; Alisha, A.; Bhatt, V.; Chandanshive, S.; Kumar, N.; Mir, Z.; Kumar, A.; Yadav, S.K.; Shivaraj, S.M.; Sonah, H.; et al. Mutation Breeding in Tomato: Advances, Applicability and Challenges. Plants 2019, 8, 128. [Google Scholar] [CrossRef]
- Patil, G.; Do, T.; Vuong, T.D.; Valliyodan, B.; Lee, J.D.; Chaudhary, J.; Shannon, J.G.; Nguyen, H.T. Genomic-assisted haplotype analysis and the development of high-throughput SNP markers for salinity tolerance in soybean. Sci. Rep. 2016, 6, 19199. [Google Scholar] [CrossRef]
- Rizhsky, L.; Liang, H.; Shuman, J.; Shulaev, V.; Davletova, S.; Mittler, R. When Defense Pathways Collide. The Response of Arabidopsis to a Combination of Drought and Heat Stress. Plant Physiol. 2004, 134, 1683–1696. [Google Scholar] [CrossRef]
- Lin, T.; Zhu, G.; Zhang, J.; Xu, X.; Yu, Q.; Zheng, Z.; Zhang, Z.; Lun, Y.; Li, S.; Wang, X.; et al. Genomic analyses provide insights into the history of tomato breeding. Nat. Genet. 2014, 46, 1220–1226. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, J.; Patil, G.B.; Sonah, H.; Deshmukh, R.K.; Vuong, T.D.; Valliyodan, B.; Nguyen, H.T. Expanding Omics Resources for Improvement of Soybean Seed Composition Traits. Front. Plant Sci. 2015, 6, 504. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Kailasrao Deshmukh, R.; Jain, N.; Kumar Singh, N. Combining qtl mapping and transcriptome profiling for an insight into genes for grain number in rice (Oryza sativa L.). Indian J. Genet. Plant Breed. 2011, 71, 115. [Google Scholar]
- Chopperla, R.; Singh, S.; Tomar, R.; Mohanty, S.; Khan, S.; Reddy, N.; Padaria, J.C.; Solanke, A.U. Isolation and allelic characterization of finger millet (Eleusine coracana L.) small heat shock protein echsp17. 8 for stress tolerance. Indian J. Genet. Plant Breed. 2018, 78, 95–103. [Google Scholar] [CrossRef]
- Sonah, H.; Deshmukh, R.K.; Sharma, A.; Singh, V.P.; Gupta, D.K.; Gacche, R.N.; Rana, J.C.; Singh, N.K.; Sharma, T.R. Genome-wide distribution and organization of microsatellites in plants: An insight into marker development in brachypodium. PLoS ONE 2011, 6, e21298. [Google Scholar] [CrossRef] [PubMed]
- Zargar, S.M.; Raatz, B.; Sonah, H.; Nazir, M.; Bhat, J.A.; Dar, Z.A.; Agrawal, G.K.; Rakwal, R. Recent advances in molecular marker techniques: Insight into QTL mapping, GWAS and genomic selection in plants. J. Crop. Sci. Biotechnol. 2015, 18, 293–308. [Google Scholar] [CrossRef]
- Francia, E.; Tacconi, G.; Crosatti, C.; Barabaschi, D.; Bulgarelli, D.; Dall’Aglio, E.; Valè, G. Marker assisted selection in crop plants. Plant Cell Tissue Organ Cult. 2005, 82, 317–342. [Google Scholar] [CrossRef]
- Kumari, S.; Mir, R.R.; Tyagi, S.; Balyan, H.S.; Gupta, P.K. Validation of QTL for grain weight using MAS-derived pairs of NILs in bread wheat (Triticum aestivum L.). J. Plant Biochem. Biotechnol. 2019, 28, 336–344. [Google Scholar] [CrossRef]
- Collard, B.C.Y.; Jahufer, M.Z.Z.; Brouwer, J.B.; Pang, E.C.K. An introduction to markers, quantitative trait loci (QTL) mapping and marker-assisted selection for crop improvement: The basic concepts. Euphytica 2005, 142, 169–196. [Google Scholar] [CrossRef]
- Agarwal, G.; Clevenger, J.; Pandey, M.K.; Wang, H.; Shasidhar, Y.; Chu, Y.; Fountain, J.C.; Choudhary, D.; Culbreath, A.K.; Liu, X.; et al. High-density genetic map using whole-genome resequencing for fine mapping and candidate gene discovery for disease resistance in peanut. Plant Biotechnol. J. 2018, 16, 1954–1967. [Google Scholar] [CrossRef] [PubMed]
- Foolad, M.R. Current Status of Breeding Tomatoes for Salt and Drought Tolerance; Springer Science and Business Media LLC: Berlin, Germany, 2007; pp. 669–700. [Google Scholar]
- Giancola, S.; McKhann, H.I.; Bérard, A.; Camilleri, C.; Durand, S.; Libeau, P.; Roux, F.; Reboud, X.; Gut, I.G.; Brunel, D. Utilization of the three high-throughput SNP genotyping methods, the GOOD assay, Amplifluor and TaqMan, in diploid and polyploid plants. Theor. Appl. Genet. 2006, 112, 1115–1124. [Google Scholar] [CrossRef]
- Sonah, H.; Bastien, M.; Iquira, E.; Tardivel, A.; Légaré, G.; Boyle, B.; Normandeau, E.; Laroche, J.; LaRose, S.; Jean, M.; et al. An Improved Genotyping by Sequencing (GBS) Approach Offering Increased Versatility and Efficiency of SNP Discovery and Genotyping. PLoS ONE 2013, 8, e54603. [Google Scholar] [CrossRef]
- 100 Tomato Genome Sequencing Consortium; Aflitos, S.; Schijlen, E.; de Jong, H.; de Ridder, D.; Smit, S.; Finkers, R.; Wang, J.; Zhang, G.; Li, N.; et al. Exploring genetic variation in the tomato (Solanum section Lycopersicon) clade by whole-genome sequencing. Plant J. 2014, 80, 136–148. [Google Scholar]
- Fukushima, A.; Kusano, M.; Redestig, H.; Arita, M.; Saito, K. Integrated omics approaches in plant systems biology. Curr. Opin. Chem. Biol. 2009, 13, 532–538. [Google Scholar] [CrossRef]
- Shah, T.; Xu, J.; Zou, X.; Cheng, Y.; Nasir, M.; Zhang, X. Omics Approaches for Engineering Wheat Production under Abiotic Stresses. Int. J. Mol. Sci. 2018, 19, 2390. [Google Scholar] [CrossRef]
- Chaudhary, J.; Deshmukh, R.; Mir, Z.A.; Bhat, J.A. Metabolomics: An emerging technology for soybean improvement. In Biotechnology Products in Everyday Life; Springer: Berlin, Germany, 2019; pp. 175–186. [Google Scholar]
- Hong, J.; Yang, L.; Zhang, D.; Shi, J. Plant Metabolomics: An Indispensable System Biology Tool for Plant Science. Int. J. Mol. Sci. 2016, 17, 767. [Google Scholar] [CrossRef]
- Berardini, T.Z.; Reiser, L.; Li, D.; Mezheritsky, Y.; Muller, R.; Strait, E.; Huala, E. The Arabidopsis information resource: Making and mining the “gold standard” annotated reference plant genome. Genesis 2015, 53, 474–485. [Google Scholar] [CrossRef]
- Goodwin, S.; McPherson, J.D.; McCombie, W.R. Coming of age: Ten years of next-generation sequencing technologies. Nat. Rev. Genet. 2016, 17, 333–351. [Google Scholar] [CrossRef]
- Varshney, R.K.; Nayak, S.N.; May, G.D.; Jackson, S.A. Next-generation sequencing technologies and their implications for crop genetics and breeding. Trends Biotechnol. 2009, 27, 522–530. [Google Scholar] [CrossRef]
- Consortium, T.G. The tomato genome sequence provides insights into fleshy fruit evolution. Nature 2012, 485, 635. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Khurana, A. Functional genomics of tomato: Opportunities and challenges in post-genome NGS era. J. Biosci. 2014, 39, 917–929. [Google Scholar] [CrossRef] [PubMed]
- Causse, M.; Desplat, N.; Pascual, L.; Le Paslier, M.C.; Sauvage, C.; Bauchet, G.; Bérard, A.; Bounon, R.; Tchoumakov, M.; Brunel, D.; et al. Whole genome resequencing in tomato reveals variation associated with introgression and breeding events. BMC Genom. 2013, 14, 791. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.; Scossa, F.; Bolger, M.E.; Lanz, C.; Maumus, F.; Tohge, T.; Quesneville, H.; Alseekh, S.; Sørensen, I.; Lichtenstein, G.; et al. The genome of the stress-tolerant wild tomato species Solanum pennellii. Nat. Genet. 2014, 46, 1034–1038. [Google Scholar] [CrossRef] [PubMed]
- Tranchida-Lombardo, V.; Cigliano, R.A.; Anzar, I.; Landi, S.; Palombieri, S.; Colantuono, C.; Bostan, H.; Termolino, P.; Aversano, R.; Batelli, G.; et al. Whole-genome re-sequencing of two Italian tomato landraces reveals sequence variations in genes associated with stress tolerance, fruit quality and long shelf-life traits. DNA Res. 2017, 25, 149–160. [Google Scholar] [CrossRef]
- Patil, V.U.; Girimalla, V.; Sagar, V.; Bhardwaj, V.; Chakrabarti, S. Draft genome sequencing of rhizoctonia solani anastomosis group 3 (ag3-pt) causing stem canker and black scurf of potato. Am. J. Potato Res. 2018, 95, 87–91. [Google Scholar] [CrossRef]
- Yang, H.; Zhao, T.; Jiang, J.; Wang, S.; Wang, A.; Li, J.; Xu, X. Mapping and screening of the tomato Stemphylium lycopersici resistance gene, Sm, based on bulked segregant analysis in combination with genome resequencing. BMC Plant Boil. 2017, 17, 266. [Google Scholar] [CrossRef]
- Arsenault-Labrecque, G.; Sonah, H.; Lebreton, A.; Labbé, C.; Marchand, G.; Xue, A.; Belzile, F.; Knaus, B.J.; Grünwald, N.J.; Bélanger, R.R. Stable predictive markers for Phytophthora sojae avirulence genes that impair infection of soybean uncovered by whole genome sequencing of 31 isolates. BMC Boil. 2018, 16, 80. [Google Scholar] [CrossRef]
- Gao, L.; Gonda, I.; Sun, H.; Ma, Q.; Bao, K.; Tieman, D.M.; Burzynski-Chang, E.A.; Fish, T.L.; Stromberg, K.A.; Sacks, G.L.; et al. The tomato pan-genome uncovers new genes and a rare allele regulating fruit flavor. Nat. Genet. 2019, 51, 1044–1051. [Google Scholar] [CrossRef] [PubMed]
- Elbasyoni, I.S.; Lorenz, A.; Guttieri, M.; Frels, K.; Baenziger, P.; Poland, J.; Akhunov, E. A comparison between genotyping-by-sequencing and array-based scoring of SNPs for genomic prediction accuracy in winter wheat. Plant Sci. 2018, 270, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Eltaher, S.; Sallam, A.; Belamkar, V.; Emara, H.A.; Nower, A.A.; Salem, K.F.M.; Poland, J.; Baenziger, P.S. Genetic Diversity and Population Structure of F3:6 Nebraska Winter Wheat Genotypes Using Genotyping-By-Sequencing. Front. Genet. 2018, 9, 76. [Google Scholar] [CrossRef] [PubMed]
- Poland, J.A.; Rife, T.W. Genotyping-by-Sequencing for Plant Breeding and Genetics. Plant Genome 2012, 5, 92. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.; Guo, H.; Kong, W.; Chandnani, R.; Shuang, L.S.; Paterson, A.H. Application of genotyping by sequencing technology to a variety of crop breeding programs. Plant Sci. 2016, 242, 14–22. [Google Scholar] [CrossRef] [Green Version]
- Sim, S.C.; Durstewitz, G.; Plieske, J.; Wieseke, R.; Ganal, M.W.; Van Deynze, A.; Hamilton, J.P.; Buell, C.R.; Causse, M.; Wijeratne, S.; et al. Development of a Large SNP Genotyping Array and Generation of High-Density Genetic Maps in Tomato. PLoS ONE 2012, 7, e40563. [Google Scholar] [CrossRef]
- Cockram, J.; Mackay, I. Genetic Mapping Populations for Conducting High-Resolution Trait Mapping in Plants In Plant Genetics and Molecular Biology; Springer: Cham, Germany, 2018; pp. 109–138. [Google Scholar]
- Pascual, L.; Desplat, N.; Huang, B.E.; Desgroux, A.; Bruguier, L.; Bouchet, J.P.; Le, Q.H.; Chauchard, B.; Verschave, P.; Causse, M. Potential of a tomato magic population to decipher the genetic control of quantitative traits and detect causal variants in the resequencing era. Plant Biotechnol. J. 2015, 13, 565–577. [Google Scholar] [CrossRef]
- Liu, Y.; Zhou, T.; Ge, H.; Pang, W.; Gao, L.; Ren, L.; Chen, H. SSR Mapping of QTLs Conferring Cold Tolerance in an Interspecific Cross of Tomato. Int. J. Genom. 2016, 2016, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Foolad, M.R.; Subbiah, P.; Zhang, L. Common qtl affect the rate of tomato seed germination under different stress and nonstress conditions. Int. J. Plant Genom. 2007, 2007, 97–106. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Liu, L.; Bai, Y.; Zhang, P.; Finkers, R.; Du, Y.; Visser, R.G.; van Heusden, A.W.J.E. Seedling salt tolerance in tomato. Euphytica 2011, 178, 403–414. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Driedonks, N.; Rutten, M.J.; Vriezen, W.H.; de Boer, G.J.; Rieu, I.J.M.B. Mapping quantitative trait loci for heat tolerance of reproductive traits in tomato (Solanum lycopersicum). Mol. Breed. 2017, 37, 58. [Google Scholar] [CrossRef]
- Pasam, R.K.; Sharma, R.; Malosetti, M.; Van Eeuwijk, F.A.; Haseneyer, G.; Kilian, B.; Graner, A. Genome-wide association studies for agronomical traits in a world wide spring barley collection. BMC Plant Boil. 2012, 12, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Zhao, J.; Xu, Y.; Liang, J.; Chang, P.; Yan, F.; Li, M.; Liang, Y.; Zou, Z. Genome-Wide Association Mapping for Tomato Volatiles Positively Contributing to Tomato Flavor. Front. Plant Sci. 2015, 6, 617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crossa, J.; Pérez-Rodríguez, P.; Cuevas, J.; Montesinos-López, O.; Jarquín, D.; Campos, G.D.L.; Burgueño, J.; González-Camacho, J.M.; Pérez-Elizalde, S.; Beyene, Y.; et al. Genomic Selection in Plant Breeding: Methods, Models, and Perspectives. Trends Plant Sci. 2017, 22, 961–975. [Google Scholar] [CrossRef]
- Deshmukh, R.; Sonah, H.; Patil, G.; Chen, W.; Prince, S.; Mutava, R.; Vuong, T.; Valliyodan, B.; Nguyen, H.T. Integrating omic approaches for abiotic stress tolerance in soybean. Front. Plant Sci. 2014, 5, 244. [Google Scholar] [CrossRef]
- Shikha, M.; Kanika, A.; Rao, A.R.; Mallikarjuna, M.G.; Gupta, H.S.; Nepolean, T. Genomic Selection for Drought Tolerance Using Genome-Wide SNPs in Maize. Front. Plant Sci. 2017, 8, 63. [Google Scholar] [CrossRef]
- Yamamoto, E.; Matsunaga, H.; Onogi, A.; Kajiya-Kanegae, H.; Minamikawa, M.; Suzuki, A.; Shirasawa, K.; Hirakawa, H.; Nunome, T.; Yamaguchi, H.; et al. A simulation-based breeding design that uses whole-genome prediction in tomato. Sci. Rep. 2016, 6, 19454. [Google Scholar] [CrossRef]
- Duangjit, J.; Causse, M.; Sauvage, C. Efficiency of genomic selection for tomato fruit quality. Mol. Breed. 2016, 36, 29. [Google Scholar] [CrossRef]
- Shinde, S.; Behpouri, A.; McElwain, J.C.; Ng, C.K.Y. Genome-wide transcriptomic analysis of the effects of sub-ambient atmospheric oxygen and elevated atmospheric carbon dioxide levels on gametophytes of the moss, Physcomitrella patens. J. Exp. Bot. 2015, 66, 4001–4012. [Google Scholar] [CrossRef] [Green Version]
- Wirta, V. Mining the Transcriptome-Methods and Applications; KTH: Stockholm, Sweden, 2006. [Google Scholar]
- Albert, E.; Duboscq, R.; Latreille, M.; Santoni, S.; Beukers, M.; Bouchet, J.P.; Bitton, F.; Gricourt, J.; Poncet, C.; Gautier, V.; et al. Allele-specific expression and genetic determinants of transcriptomic variations in response to mild water deficit in tomato. Plant J. 2018, 96, 635–650. [Google Scholar] [CrossRef]
- Iovieno, P.; Punzo, P.; Guida, G.; Mistretta, C.; Van Oosten, M.J.; Nurcato, R.; Bostan, H.; Colantuono, C.; Costa, A.; Bagnaresi, P.; et al. Transcriptomic Changes Drive Physiological Responses to Progressive Drought Stress and Rehydration in Tomato. Front. Plant Sci. 2016, 7, 371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fei, Z. Tomato Expression Database (TED): A suite of data presentation and analysis tools. Nucleic Acids Res. 2006, 34, D766–D770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oono, Y.; Yazawa, T.; Kanamori, H.; Sasaki, H.; Mori, S.; Handa, H.; Matsumoto, T. Genome-Wide Transcriptome Analysis of Cadmium Stress in Rice. BioMed Res. Int. 2016, 2016, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sa, K.J.; Choi, I.Y.; Park, D.H.; Lee, J.K. Comparative Gene Expression Analysis of Seed Development in Waxy and Dent Corn (Zea mays L.). Plant Breed. Biotechnol. 2018, 6, 337–353. [Google Scholar] [CrossRef]
- Garcia, B.J.; Labbé, J.L.; Jones, P.; Abraham, P.E.; Hodge, I.; Climer, S.; Jawdy, S.; Gunter, L.; Tuskan, G.A.; Yang, X.; et al. Phytobiome and Transcriptional Adaptation of Populus deltoides to Acute Progressive Drought and Cyclic Drought. Phytobiomes J. 2018, 2, 249–260. [Google Scholar] [CrossRef] [Green Version]
- Gong, P.; Zhang, J.; Li, H.; Yang, C.; Zhang, C.; Zhang, X.; Khurram, Z.; Zhang, Y.; Wang, T.; Fei, Z.; et al. Transcriptional profiles of drought-responsive genes in modulating transcription signal transduction, and biochemical pathways in tomato. J. Exp. Bot. 2010, 61, 3563–3575. [Google Scholar] [CrossRef]
- Ashrafi-Dehkordi, E.; Alemzadeh, A.; Tanaka, N.; Razi, H. Meta-analysis of transcriptomic responses to biotic and abiotic stress in tomato. PeerJ 2018, 6, e4631. [Google Scholar] [CrossRef]
- Sharma, M.K.; Kumar, R.; Solanke, A.U.; Sharma, R.; Tyagi, A.K.; Sharma, A.K. Identification, phylogeny, and transcript profiling of ERF family genes during development and abiotic stress treatments in tomato. Mol. Genet. Genom. 2010, 284, 455–475. [Google Scholar] [CrossRef]
- Wang, Y.; Tao, X.; Tang, X.M.; Xiao, L.; Sun, J.L.; Yan, X.F.; Li, D.; Deng, H.Y.; Ma, X.R. Comparative transcriptome analysis of tomato (Solanum lycopersicum) in response to exogenous abscisic acid. BMC Genom. 2013, 14, 841. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Shim, D.; Moon, S.; Kim, H.; Bae, W.; Kim, K.; Kim, Y.H.; Rhee, S.K.; Hong, C.P.; Hong, S.Y.; et al. Genome-wide transcriptomic analysis of BR-deficient Micro-Tom reveals correlations between drought stress tolerance and brassinosteroid signaling in tomato. Plant Physiol. Biochem. 2018, 127, 553–560. [Google Scholar] [CrossRef]
- Keshishian, E.A.; Hallmark, H.T.; Ramaraj, T.; Plačková, L.; Sundararajan, A.; Schilkey, F.; Novák, O.; Rashotte, A.M. Salt and oxidative stresses uniquely regulate tomato cytokinin levels and transcriptomic response. Plant Direct 2018, 2, e00071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, Q.; Geng, L.; Lu, M.; Jin, W.; Nan, X.; He, P.A.; Yao, Y. Comparative transcriptome analysis of the different tissues between the cultivated and wild tomato. PLoS ONE 2017, 12, 0172411. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Pozo, N.; Zheng, Y.; Snyder, S.I.; Nicolas, P.; Shinozaki, Y.; Fei, Z.; Catala, C.; Giovannoni, J.J.; Rose, J.K.; Mueller, L.A. The tomato expression atlas. Bioinformatics 2017, 33, 2397–2398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukushima, A.; Nishizawa, T.; Hayakumo, M.; Hikosaka, S.; Saito, K.; Goto, E.; Kusano, M. Exploring tomato gene functions based on coexpression modules using graph clustering and differential coexpression approaches. Plant Physiol. 2012, 158, 1487–1502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, L.; Dong, C.; Shang, Q. Gene co-expression network analysis reveals pathways associated with graft healing by asymmetric profiling in tomato. BMC Plant Boil. 2019, 19, 373. [Google Scholar] [CrossRef] [PubMed]
- Thagun, C.; Imanishi, S.; Kudo, T.; Nakabayashi, R.; Ohyama, K.; Mori, T.; Kawamoto, K.; Nakamura, Y.; Katayama, M.; Nonaka, S.; et al. Jasmonate-responsive ERF transcription factors regulate steroidal glycoalkaloid biosynthesis in tomato. Plant Cell Physiol. 2016, 57, 961–975. [Google Scholar] [CrossRef] [Green Version]
- Cárdenas, P.D.; Sonawane, P.D.; Pollier, J.; Bossche, R.V.; Dewangan, V.; Weithorn, E.; Tal, L.; Meir, S.; Rogachev, I.; Malitsky, S.; et al. GAME9 regulates the biosynthesis of steroidal alkaloids and upstream isoprenoids in the plant mevalonate pathway. Nat. Commun. 2016, 7, 10654. [Google Scholar] [CrossRef] [Green Version]
- Itkin, M.; Rogachev, I.; Alkan, N.; Rosenberg, T.; Malitsky, S.; Masini, L.; Meir, S.; Iijima, Y.; Aoki, K.; De Vos, R.; et al. GLYCOALKALOID METABOLISM1 Is Required for Steroidal Alkaloid Glycosylation and Prevention of Phytotoxicity in Tomato. Plant Cell 2011, 23, 4507–4525. [Google Scholar] [CrossRef] [Green Version]
- Klee, H.J.; Giovannoni, J.J. Genetics and Control of Tomato Fruit Ripening and Quality Attributes. Annu. Rev. Genet. 2011, 45, 41–59. [Google Scholar] [CrossRef]
- Neilson, K.A.; Ali, N.A.; Muralidharan, S.; Mirzaei, M.; Mariani, M.; Assadourian, G.; Lee, A.; Van Sluyter, S.C.; Haynes, P.A. Less label, more free: Approaches in label-free quantitative mass spectrometry. Proteomics 2011, 11, 535–553. [Google Scholar] [CrossRef]
- Mora, L.; Bramley, P.M.; Fraser, P.D.; Soler, L.M. Development and optimisation of a label-free quantitative proteomic procedure and its application in the assessment of genetically modified tomato fruit. Proteomics 2013, 13, 2016–2030. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Sauvé, R.; Thannhauser, T.W. Proteome changes induced by aluminium stress in tomato roots. Plant Signal. Behav. 2009, 60, 1849–1857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muneer, S.; Jeong, B.R. Proteomic analysis of salt-stress responsive proteins in roots of tomato (Lycopersicon esculentum L.) plants towards silicon efficiency. Plant Growth Regul. 2015, 77, 133–146. [Google Scholar] [CrossRef]
- Ahsan, N.; Lee, D.G.; Lee, S.H.; Kang, K.Y.; Bahk, J.D.; Choi, M.S.; Lee, I.J.; Renaut, J.; Lee, B.H. A comparative proteomic analysis of tomato leaves in response to waterlogging stress. Physiol. Plant. 2007, 131, 555–570. [Google Scholar] [CrossRef]
- Salazar-Salas, N.Y.; Valenzuela-Ponce, L.; Vega-Garcia, M.O.; Pineda-Hidalgo, K.V.; Vega-Alvarez, M.; Chavez-Ontiveros, J.; Delgado-Vargas, F.; Lopez-Valenzuela, J.A. Protein changes associated with chilling tolerance in tomato fruit with hot water pre-treatment. Postharvest Boil. Technol. 2017, 134, 22–30. [Google Scholar] [CrossRef]
- Gong, B.; Zhang, C.; Li, X.; Wen, D.; Wang, S.; Shi, Q.; Wang, X. Identification of nacl and nahco3 stress responsive proteins in tomato roots using itraq-based analysis. Biochem. Biophys. Res. Commun. 2014, 446, 417–422. [Google Scholar] [CrossRef]
- Muneer, S.; Ko, C.H.; Wei, H.; Chen, Y.; Jeong, B.R. Physiological and Proteomic Investigations to Study the Response of Tomato Graft Unions under Temperature Stress. PLoS ONE 2016, 11, e0157439. [Google Scholar] [CrossRef] [Green Version]
- Tamburino, R.; Vitale, M.; Ruggiero, A.; Sassi, M.; Sannino, L.; Arena, S.; Costa, A.; Batelli, G.; Zambrano, N.; Scaloni, A.; et al. Chloroplast proteome response to drought stress and recovery in tomato (Solanum lycopersicum L.). BMC Plant Boil. 2017, 17, 40. [Google Scholar] [CrossRef] [Green Version]
- Sang, Q.; Shan, X.; An, Y.; Shu, S.; Sun, J.; Guo, S. Proteomic Analysis Reveals the Positive Effect of Exogenous Spermidine in Tomato Seedlings’ Response to High-Temperature Stress. Front. Plant Sci. 2017, 8, 555. [Google Scholar] [CrossRef] [Green Version]
- Alam Khan, T.; Yusuf, M.; Ahmad, A.; Bashir, Z.; Saeed, T.; Fariduddin, Q.; Hayat, S.; Mock, H.P.; Wu, T.; Khan, T.A. Proteomic and physiological assessment of stress sensitive and tolerant variety of tomato treated with brassinosteroids and hydrogen peroxide under low-temperature stress. Food Chem. 2019, 289, 500–511. [Google Scholar] [CrossRef]
- Kaspar, S.; Peukert, M.; Mock, H.P.; Svatos, A.; Matros, A.; Mock, H. MALDI-imaging mass spectrometry—An emerging technique in plant biology. Proteomics 2011, 11, 1840–1850. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J.; Perdian, D.C.; Song, Z.; Yeung, E.S.; Nikolau, B.J. Use of mass spectrometry for imaging metabolites in plants. Plant J. 2012, 70, 81–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schripsema, J. Application of nmr in plant metabolomics: Techniques, problems and prospects. Hytochem. Anal. Int. J. Plant Chem. Biochem. Tech. 2010, 21, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Rodríguez, E.; Moreno, D.A.; Ferreres, F.; Rubio-Wilhelmi, M.D.M.; Ruiz, J.M. Differential responses of five cherry tomato varieties to water stress: Changes on phenolic metabolites and related enzymes. Phytochemistry 2011, 72, 723–729. [Google Scholar] [CrossRef] [PubMed]
- Ampofo-Asiama, J.; Baiye, V.; Hertog, M.; Waelkens, E.; Geeraerd, A.; Nicolai, B.J.P.B. The metabolic response of cultured tomato cells to low oxygen stress. Plant Biol. 2014, 16, 594–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, G.; Wang, S.; Huang, Z.; Zhang, S.; Liao, Q.; Zhang, C.; Lin, T.; Qin, M.; Peng, M.; Yang, C.; et al. Rewiring of the Fruit Metabolome in Tomato Breeding. Cell 2018, 172, 249–261. [Google Scholar] [CrossRef] [Green Version]
- Nunes-Nesi, A.; Alseekh, S.; Silva, F.M.D.O.; Omranian, N.; Lichtenstein, G.; Mirnezhad, M.; González, R.R.R.; Garcia, J.S.Y.; Conte, M.; Leiss, K.A.; et al. Identification and characterization of metabolite quantitative trait loci in tomato leaves and comparison with those reported for fruits and seeds. Metabolomics 2019, 15, 46. [Google Scholar] [CrossRef] [Green Version]
- Salt, D.E.; Baxter, I.; Lahner, B. Ionomics and the Study of the Plant Ionome. Annu. Rev. Plant Boil. 2008, 59, 709–733. [Google Scholar] [CrossRef] [Green Version]
- Bhat, J.A.; Shivaraj, S.M.; Singh, P.; Navadagi, D.B.; Tripathi, D.K.; Dash, P.K.; Solanke, A.U.; Sonah, H.; Deshmukh, R. Role of Silicon in Mitigation of Heavy Metal Stresses in Crop Plants. Plants 2019, 8, 71. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, P.; Tripathi, D.K.; Deshmukh, R.; Singh, V.P.; Corpas, F.J. Revisiting the role of ROS and RNS in plants under changing environment. Environ. Exp. Bot. 2019, 161, 1–3. [Google Scholar] [CrossRef]
- Kim, Y.H.; Khan, A.L.; Waqas, M.; Lee, I.J. Silicon Regulates Antioxidant Activities of Crop Plants under Abiotic-Induced Oxidative Stress: A Review. Front. Plant Sci. 2017, 8, 282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Zhu, Y.; Hu, Y.; Han, W.; Gong, H. Beneficial effects of silicon in alleviating salinity stress of tomato seedlings grown under sand culture. Acta Physiol. Plant. 2015, 37, 71. [Google Scholar] [CrossRef]
- Haghighi, M.; Pessarakli, M. Influence of silicon and nano-silicon on salinity tolerance of cherry tomatoes (Solanum lycopersicum L.) at early growth stage. Sci. Hortic. 2013, 161, 111–117. [Google Scholar] [CrossRef]
- Marmiroli, M.; Pigoni, V.; Savo-Sardaro, M.; Marmiroli, N. The effect of silicon on the uptake and translocation of arsenic in tomato (Solanum lycopersicum L.). Environ. Exp. Bot. 2014, 99, 9–17. [Google Scholar] [CrossRef]
- Ichihashi, Y.; Sinha, N.R. From genome to phenome and back in tomato. Curr. Opin. Plant Boil. 2014, 18, 9–15. [Google Scholar] [CrossRef] [PubMed]
- White, J.W.; Andrade-Sanchez, P.; Gore, M.A.; Bronson, K.F.; Coffelt, T.A.; Conley, M.M.; Feldmann, K.A.; French, A.N.; Heun, J.T.; Hunsaker, D.J.; et al. Field-based phenomics for plant genetics research. Field Crops Res. 2012, 133, 101–112. [Google Scholar] [CrossRef]
- Ubbens, J.R.; Stavness, I. Deep Plant Phenomics: A Deep Learning Platform for Complex Plant Phenotyping Tasks. Front. Plant Sci. 2017, 8, 8. [Google Scholar] [CrossRef] [Green Version]
- Tardieu, F.; Cabrera-Bosquet, L.; Pridmore, T.; Bennett, M. Plant Phenomics, From Sensors to Knowledge. Curr. Boil. 2017, 27, R770–R783. [Google Scholar] [CrossRef]
- Laxman, R.H.; Hemamalini, P.; Bhatt, R.M.; Sadashiva, A.T. Non-invasive quantification of tomato (Solanum lycopersicum L.) plant biomass through digital imaging using phenomics platform. Indian J. Plant Physiol. 2018, 23, 369–375. [Google Scholar] [CrossRef]
- Cohen, H.; Aharoni, A.; Szymanski, J.; Dominguez, E. Assimilation of ‘omics’ strategies to study the cuticle layer and suberin lamellae in plants. J. Exp. Bot. 2017, 68, 5389–5400. [Google Scholar] [CrossRef]
- Chaudhary, J.; Shivaraj, S.; Khatri, P.; Ye, H.; Zhou, L.; Klepadlo, M.; Dhakate, P.; Kumawat, G.; Patil, G.; Sonah, H.; et al. Approaches, Applicability, and Challenges for Development of Climate-Smart Soybean. In Genomic Designing of Climate-Smart Oilseed Crops; Springer Science and Business Media LLC: Berlin, Germany, 2019; pp. 1–74. [Google Scholar]
- Shivaraj, S.M.; Dhakate, P.; Sonah, H.; Vuong, T.; Nguyen, H.T.; Deshmukh, R. Progress Toward Development of Climate-Smart Flax: A Perspective on Omics-Assisted Breeding. In Genomic Designing of Climate-Smart Oilseed Crops; Springer Science and Business Media LLC: Berlin, Germany, 2019; pp. 239–274. [Google Scholar]
- Sonah, H.; O’Donoughue, L.; Cober, E.; Rajcan, I.; Belzile, F. Identification of loci governing eight agronomic traits using a gbs-gwas approach and validation by qtl mapping in soya bean. Plant Biotechnol. J. 2015, 13, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Yao, Q.; Patil, G.B.; Agarwal, G.; Deshmukh, R.K.; Lin, L.; Wang, B.; Wang, Y.; Prince, S.J.; Song, L.; et al. Identification and Comparative Analysis of Differential Gene Expression in Soybean Leaf Tissue under Drought and Flooding Stress Revealed by RNA-Seq. Front. Plant Sci. 2016, 7, 827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonah, H.; Zhang, X.; Deshmukh, R.K.; Borhan, M.H.; Fernando, W.G.D.; Bélanger, R.R. Comparative Transcriptomic Analysis of Virulence Factors in Leptosphaeria maculans during Compatible and Incompatible Interactions with Canola. Front. Plant Sci. 2016, 7, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deshmukh, R.; Singh, A.; Jain, N.; Anand, S.; Gacche, R.; Singh, A.; Gaikwad, K.; Sharma, T.; Mohapatra, T.; Singh, N. Identification of candidate genes for grain number in rice (Oryza sativa L.). Funct. Integr. Genom. 2010, 10, 339–347. [Google Scholar] [CrossRef]
- Guo, T.; Yang, J.; Li, D.; Sun, K.; Luo, L.; Xiao, W.; Wang, J.; Liu, Y.; Wang, S.; Wang, H.; et al. Integrating GWAS, QTL, mapping and RNA-seq to identify candidate genes for seed vigor in rice (Oryza sativa L.). Mol. Breed. 2019, 39, 87. [Google Scholar] [CrossRef]
- Albert, E.; Sauvage, C.; Bouchet, J.P.; Bitton, F.; Beukers, M.; Carretero, Y.; Causse, M. Integration of qtl, eqtl and allele specific expression to unravel genotype by watering regime interaction in cultivated tomato. In Proceedings of the Plant and Animal Genome Conference (PAG), San Diego, CA, USA, 14–18 January 2017. [Google Scholar]
- Aoki, K.; Yano, K.; Suzuki, A.; Kawamura, S.; Sakurai, N.; Suda, K.; Kurabayashi, A.; Suzuki, T.; Tsugane, T.; Watanabe, M.; et al. Large-scale analysis of full-length cDNAs from the tomato (Solanum lycopersicum) cultivar Micro-Tom, a reference system for the Solanaceae genomics. BMC Genom. 2010, 11, 210. [Google Scholar] [CrossRef] [Green Version]
- Yano, K.; Watanabe, M.; Yamamoto, N.; Tsugane, T.; Aoki, K.; Sakurai, N.; Shibata, D. MiBASE: A database of a miniature tomato cultivar Micro-Tom. Plant Biotechnol. 2006, 23, 195–198. [Google Scholar] [CrossRef] [Green Version]
- Shikata, M.; Hoshikawa, K.; Ariizumi, T.; Fukuda, N.; Yamazaki, Y.; Ezura, H. TOMATOMA Update: Phenotypic and Metabolite Information in the Micro-Tom Mutant Resource. Plant Cell Physiol. 2015, 57, e11. [Google Scholar] [CrossRef] [Green Version]
- Kudo, T.; Kobayashi, M.; Terashima, S.; Katayama, M.; Ozaki, S.; Kanno, M.; Saito, M.; Yokoyama, K.; Ohyanagi, H.; Aoki, K.; et al. TOMATOMICS: A Web Database for Integrated Omics Information in Tomato. Plant Cell Physiol. 2017, 58, e8. [Google Scholar] [CrossRef] [Green Version]
- Suresh, B.V.; Roy, R.; Sahu, K.; Misra, G.; Chattopadhyay, D. Tomato Genomic Resources Database: An Integrated Repository of Useful Tomato Genomic Information for Basic and Applied Research. PLoS ONE 2014, 9, e86387. [Google Scholar] [CrossRef]
- Fei, Z.; Joung, J.G.; Tang, X.; Zheng, Y.; Huang, M.; Lee, J.M.; McQuinn, R.; Tieman, D.M.; Alba, R.; Klee, H.J.; et al. Tomato Functional Genomics Database: A comprehensive resource and analysis package for tomato functional genomics. Nucleic Acids Res. 2010, 39, D1156–D1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirasawa, K.; Hirakawa, H. DNA marker applications to molecular genetics and genomics in tomato. Breed. Sci. 2013, 63, 21–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moco, S.; Bino, R.J.; Vorst, O.; Verhoeven, H.A.; De Groot, J.; Van Beek, T.A.; Vervoort, J.; De Vos, C.R. A Liquid Chromatography-Mass Spectrometry-Based Metabolome Database for Tomato1. Plant Physiol. 2006, 141, 1205–1218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narise, T.; Sakurai, N.; Obayashi, T.; Ohta, H.; Shibata, D. Co-expressed Pathways DataBase for Tomato: A database to predict pathways relevant to a query gene. BMC Genom. 2017, 18, 437. [Google Scholar] [CrossRef]
- Shinozaki, Y.; Nicolas, P.; Fernandez-Pozo, N.; Ma, Q.; Evanich, D.J.; Shi, Y.; Xu, Y.; Zheng, Y.; Snyder, S.I.; Martin, L.B.B.; et al. High-resolution spatiotemporal transcriptome mapping of tomato fruit development and ripening. Nat. Commun. 2018, 9, 364. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Sr.No. | Stress | Trait | QTL | Chromosome | Position (cM) | LOD Score | R (%) | References |
|---|---|---|---|---|---|---|---|---|
| 1 | Cold Tolerance | RGR (Relative Germination Ratio) | qRGI-1-1 | 1 | 40.4 | 5.45 | 19.55 | [41] |
| qRGI-1-2 | 1 | 47.2 | 2.53 | 8.52 | ||||
| qRGI-4-1 | 4 | 10.4 | 2.06 | 6.02 | ||||
| qRGI-9-1 | 9 | 7.8 | 2.12 | 5.95 | ||||
| qRGI-12-1 | 12 | 8 | 4.26 | 11.33 | ||||
| CI (Chilling index) | qCI-1-1 | 1 | 9.8 | 3.25 | 0.95 | |||
| qCI-2-1 | 2 | 18 | 2.96 | 10.34 | ||||
| qCI-3-1 | 3 | 0 | 3.01 | 10.31 | ||||
| qCI-9-1 | 9 | 26.8 | 2.35 | 7.31 | ||||
| 2 | cold stress | Seed Germination | cld1.1 | 1 | 7 | 7.41 | 30.95 | [42] |
| cld1.2 | 1 | 8.8 | 4.27 | 17.24 | ||||
| cld1.3 | 1 | 15.4 | 2.27 | 9.78 | ||||
| cld4.1 | 4 | 13.8 | 2.06 | 9.13 | ||||
| cld4.1 | 4 | 20.7 | 2.98 | 13.15 | ||||
| cld8.1 | 8 | 2.5 | 1.26 | 5.76 | ||||
| 3 | Salt Stress | Seed Germination | slt1.1 | 1 | 7 | 2.66 | 10.86 | |
| slt1.2 | 1 | 8.8 | 3.53 | 13.58 | ||||
| slt2.1 | 2 | 18.7 | 1.2 | 6.4 | ||||
| slt5.1 | 5 | 16 | 1.52 | 8.32 | ||||
| slt7.1 | 7 | 4.5 | 1.52 | 7.16 | ||||
| slt9.1 | 9 | 21.4 | 2.01 | 6.3 | ||||
| slt12.1 | 12 | 7.1 | 2.4 | 12.41 | ||||
| 4 | Salt Tolerance | Seedling Stage | Stlq4 | 4 | 63.6 | [43] | ||
| Stlq6 | 6 | 64.8 | ||||||
| Stlq9a | 9 | 61 | ||||||
| Stlq9b | 9 | 63.6 | ||||||
| Stlq12a | 12 | 63.3 | ||||||
| Stlq12a | 12 | 61 | ||||||
| Stlq12b | 12 | 64.4 | ||||||
| 5 | Heat tolerance | Pollen viability | qPV11 | 11 | 19.4 | 36.3 | [44] | |
| Pollen Number | qPN7 | 7 | 134.7 | 18.6 | ||||
| Style protrusion | qSP1 | 1 | 16 | 19.5 | ||||
| qSP3 | 3 | 80.4 | 28 | |||||
| Anther length | qAL1 | 1 | 70 | 15.5 | ||||
| qAL2 | 2 | 80.8 | 11.6 | |||||
| qAL7 | 7 | 134.7 | 25.2 | |||||
| Style length | qSL1 | 1 | 16 | 22.7 | ||||
| qSL2 | 2 | 80.8 | 10.5 | |||||
| qSL3 | 3 | 75.8 | 15.8 | |||||
| Flowers per | qFPI1 | 1 | 40 | 38.7 | ||||
| inflorescence | ||||||||
| Inflorescence number | qIN1 | 1 | 39 | 21.9 | ||||
| qIN8 | 8 | 95.3 | 13.4 | |||||
| Trait | Platform | DEG | Key Point | References |
|---|---|---|---|---|
| Microarray gene expression data of tomato to study meta-analysis of stress response | Affymetrix tomato Genome Array | 835 | Expression profile of different genes under different conditions, Meta-analysis to characterize the candidate genes for abiotic stress. | [61] |
| Temporal stage of fruit development To study the transcriptome profiling of ERF family genes. | Tomato Gene Chip arrays | 57 | Over expression of ERF family genes in tomato has been shown to confer increased resistance to abiotic stresses. | [62] |
| Tomato leaf responses to exogenous ABA | Illumina RNA-sequencing | 2787 | Exogenous ABA has potential to up- regulate many genes related to stress tolerance. | [63] |
| Solanum lycopersicum cultivars (WT) and MT (Micro-Tom) | RNA-sequencing | 619 | BR-deficient (Brassinosteroids) Micro-Tom showed lower drought and osmotic stress tolerance. BR signaling is tightly connected with gene networks related to abiotic stress and development | [64] |
| Micro-Tom seedling | RNA sequencing IlluminaGAIIx Platform | 6643 | Salt and oxidative stresses regulate tomato cytokinin level and transcriptomic responses. | [65] |
| Different stages of cultivated and wild tomato (Root, stem, leaf, flower, fruit and seedling) | RNA sequencing Illumina high-throughput sequencing | Upregulated- 126 Downregulated-87 | These DEG associated with salt resistance, drought resistance and fruit nutrition. | [66] |
| Sr.No | Database | URL | Description/Applications | References |
|---|---|---|---|---|
| 1. | KaFtom | www.pgb.kazusa.or.jp | Database for Micro Tom full length cDNA clones, Full length cDNA libraries for EST sequencing. | [113] |
| 2. | MiBASE | http://omictools.com | Database for Micro Tom ESTs and tomato Unigenes EST Sequencing, ESTAnnotations, SNPs, SSRs, Gene ontology, Metabolic pathways of Gene expressions And Sequence similarities. | [114] |
| 3. | Tomatoma (Micro Tom Database) | http://tomatoma.nbrp.jp | Micro Tom mutant Resources, Metabolite information, Phenotype information, TILLING. | [115] |
| 4. | Tomatomics | http://omictool.com/tomatomics-tool | Full length mRNA sequences, Gene structures, Expression Profiles and functional annotations of genes. | [116] |
| 5. | TGRD (Tomato Genomic Resources Database) | http://omictool.com/trgd-tool | Interactive browsing of tomato genes, micro RNAs, simple sequence repeats (SSRs), important quantitative trait loci. | [117] |
| 6. | TFGD (Tomato Functional Genomic Database) | http://ted.bti.cornell.edu | Microarray Expression Database, Metabolite profile Data analysis, RNA Seq. Data | [118] |
| 7. | KaTomics DB (Kazusa Tomato Genomic Database) | www.kazusa.or.jp | Database for DNA markers, SNP annotations, and genome sequences | [119] |
| 8. | MoToDB Metabolome Database | http://appliedbioinformatics.wur.nl/moto/ | LC-MS (Liquid Chromatography Mass Spectrometry) | [120] |
| 9. | CoxPathDB | http://cox-path-db.kazusa.or.jp/tomato | To predict function of tomato genes from result of functional enrichment analyses of co-expressed genes. | [121] |
| 10. | Sol Genomics Network | http://solgenomics.net | Browse the tomato genome, Find the sequence similarity, and Download annotations. | [122] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chaudhary, J.; Khatri, P.; Singla, P.; Kumawat, S.; Kumari, A.; R, V.; Vikram, A.; Jindal, S.K.; Kardile, H.; Kumar, R.; et al. Advances in Omics Approaches for Abiotic Stress Tolerance in Tomato. Biology 2019, 8, 90. https://0-doi-org.brum.beds.ac.uk/10.3390/biology8040090
Chaudhary J, Khatri P, Singla P, Kumawat S, Kumari A, R V, Vikram A, Jindal SK, Kardile H, Kumar R, et al. Advances in Omics Approaches for Abiotic Stress Tolerance in Tomato. Biology. 2019; 8(4):90. https://0-doi-org.brum.beds.ac.uk/10.3390/biology8040090
Chicago/Turabian StyleChaudhary, Juhi, Praveen Khatri, Pankaj Singla, Surbhi Kumawat, Anu Kumari, Vinaykumar R, Amit Vikram, Salesh Kumar Jindal, Hemant Kardile, Rahul Kumar, and et al. 2019. "Advances in Omics Approaches for Abiotic Stress Tolerance in Tomato" Biology 8, no. 4: 90. https://0-doi-org.brum.beds.ac.uk/10.3390/biology8040090