“What You Need, Baby, I Got It”: Transposable Elements as Suppliers of Cis-Operating Sequences in Drosophila


Abstract
:1. Introduction
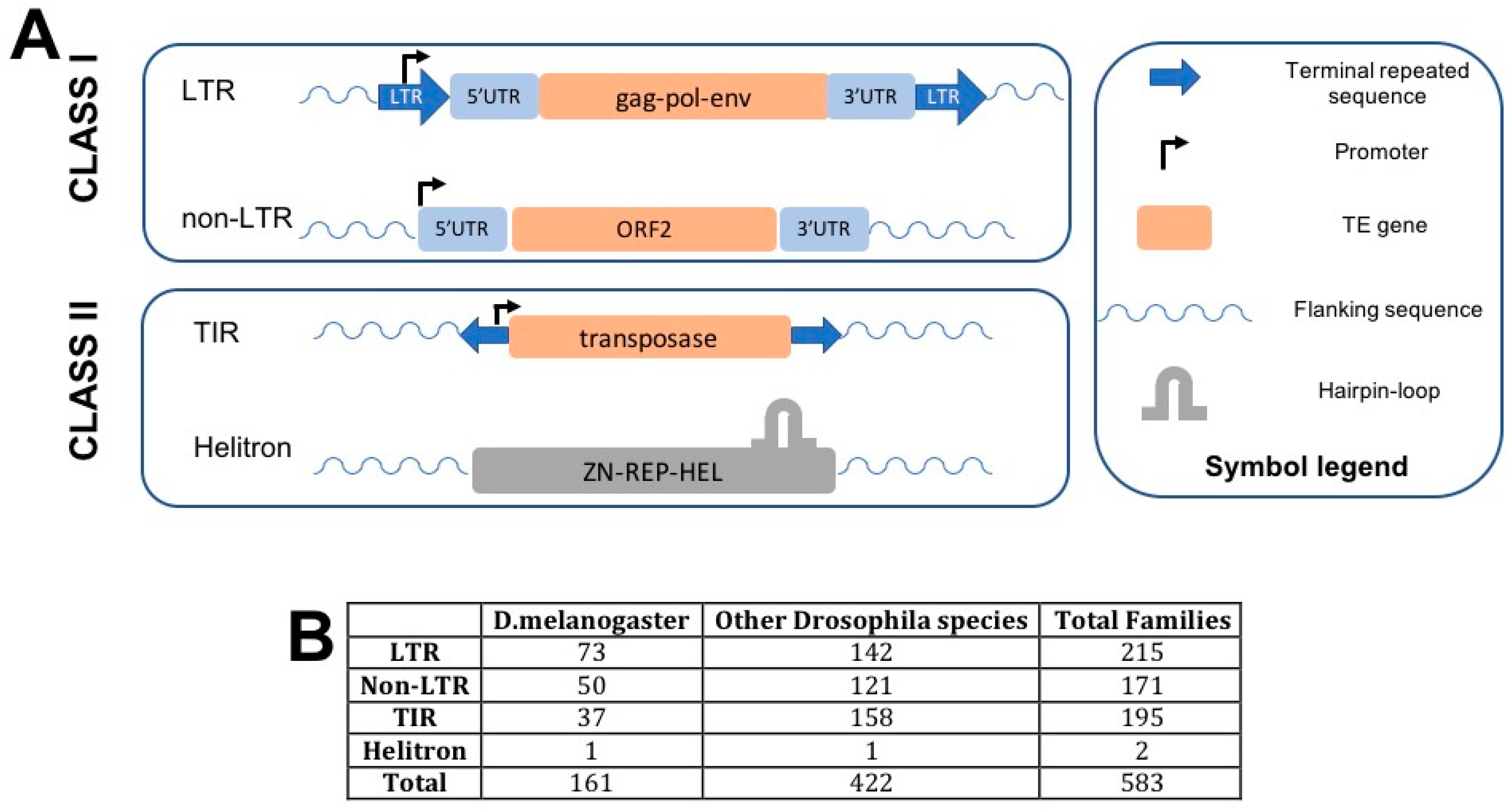
2. Drosophila TEs: A Brief Overview
3. TEs as Promoter Suppliers
3.1. Enhancers, Silencers and Insulators within TEs
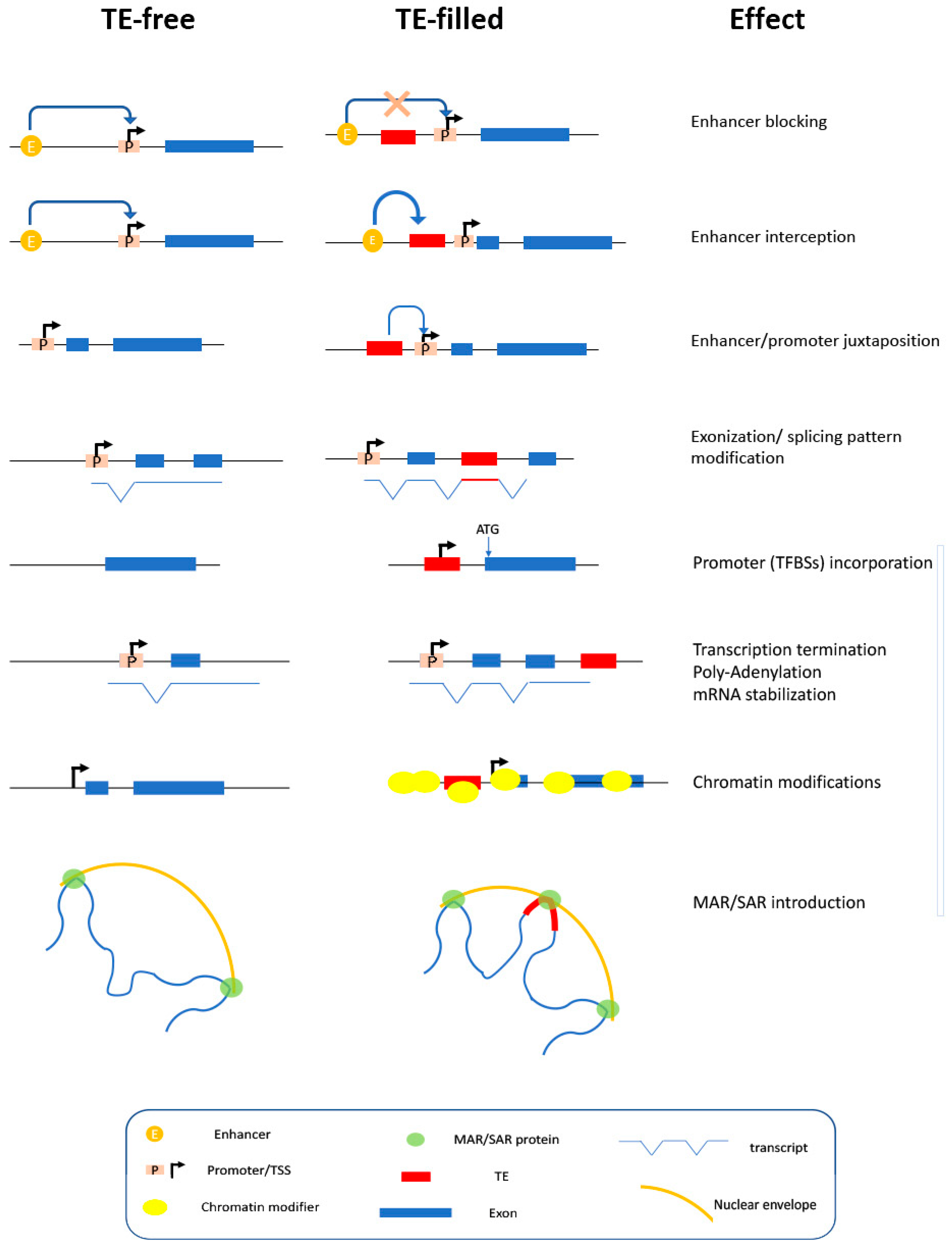
3.2. Additional Cis-Regulatory Transcriptional Signals within TE
3.3. Structural Role of Cis-Operating Sequences within TEs
4. Conclusions and Future Directions: What Can We Still Learn from Drosophila?
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- McClintock, B. The origin and behavior of mutable loci in maize. Proc. Natl. Acad. Sci. USA 1950, 36, 344–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orgel, L.E.; Crick, F.H. Selfish DNA: The ultimate parasite. Nature 1980, 284, 604–607. [Google Scholar] [CrossRef] [PubMed]
- Doolittle, W.F.; Sapienza, C. Selfish genes, the phenotype paradigm and genome evolution. Nature 1980, 284, 601–603. [Google Scholar] [CrossRef] [PubMed]
- Britten, R.J.; Kohne, D.E. Repeated sequences in DNA. Hundreds of thousands of copies of DNA sequences have been incorporated into the genomes of higher organisms. Science 1968, 161, 529–540. [Google Scholar] [CrossRef]
- Cosby, R.L.; Chang, N.C.; Feschotte, C. Host-transposon interactions: Conflict, cooperation, and cooption. Genes. Dev. 2019, 33, 1098–1116. [Google Scholar] [CrossRef] [Green Version]
- Arkhipova, I.R.; Mazo, A.M.; Cherkasova, V.A.; Gorelova, T.V.; Schuppe, N.G.; Llyin, Y.V. The steps of reverse transcription of Drosophila mobile dispersed genetic elements and U3-R-U5 structure of their LTRs. Cell 1986, 44, 555–563. [Google Scholar] [CrossRef]
- Luan, D.D.; Korman, M.H.; Jakubczak, J.L.; Eickbush, T.H. Reverse transcription of R2Bm RNA is primed by a nick at the chromosomal target site: A mechanism for non-LTR retrotransposition. Cell 1993, 72, 595–605. [Google Scholar] [CrossRef]
- St Johnston, D. The art and design of genetic screens: Drosophila melanogaster. Nat. Rev. Genet. 2002, 3, 176–188. [Google Scholar] [CrossRef]
- Hales, K.G.; Korey, C.A.; Larracuente, A.M.; Roberts, D.M. Genetics on the Fly: A Primer on the Drosophila Model System. Genetics 2015, 201, 815–842. [Google Scholar] [CrossRef] [Green Version]
- Adams, M.D.; Celniker, S.E.; Holt, R.A.; Evans, C.A.; Gocayne, J.D.; Amanatides, P.G.; Scherer, S.E.; Li, P.W.; Hoskins, R.A.; Galle, R.F.; et al. The genome sequence of Drosophila melanogaster. Science 2000, 287, 2185–2195. [Google Scholar] [CrossRef] [Green Version]
- Hoskins, R.A.; Smith, C.D.; Carlson, J.W.; Carvalho, A.B.; Halpern, A.; Kaminker, J.S.; Kennedy, C.; Mungall, C.J.; Sullivan, B.A.; Sutton, G.G.; et al. Heterochromatic sequences in a Drosophila whole-genome shotgun assembly. Genome. Biol. 2002, 3, RESEARCH0085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoskins, R.A.; Carlson, J.W.; Wan, K.H.; Park, S.; Mendez, I.; Galle, S.E.; Booth, B.W.; Pfeiffer, B.D.; George, R.A.; Svirskas, R.; et al. The Release 6 reference sequence of the Drosophila melanogaster genome. Genome Res. 2015, 25, 455–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Signor, S.; Seher, T.; Kopp, A. Genomic resources for multiple species in the Drosophila ananassae species group. Fly 2013, 7, 47–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilbert, D.G. DroSpeGe: Rapid access database for new Drosophila species genomes. Nucleic Acids. Res. 2007, 35, D480–D485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Celniker, S.E.; Dillon, L.A.L.; Gerstein, M.B.; Gunsalus, K.C.; Henikoff, S.; Karpen, G.H.; Kellis, M.; Lai, E.C.; Lieb, J.D.; MacAlpine, D.M.; et al. Unlocking the secrets of the genome. Nature 2009, 459, 927–930. [Google Scholar] [CrossRef] [Green Version]
- Agapite, J.; Albou, L.P.; Aleksander, S.; Argasinska, J.; Arnaboldi, V.; Attrill, H.; Bello, S.M.; Blake, J.A.; Blodgett, O.; Bradford, Y.M. Alliance of Genome Resources C. Alliance of Genome Resources Portal: Unified model organism research platform. Nucleic Acids Res. 2020, 48, D650–D658. [Google Scholar]
- Thurmond, J.; Goodman, J.L.; Strelets, V.B.; Attrill, H.; Gramates, L.S.; Marygold, S.J.; Matthews, B.B.; Millburn, G.; Antonazzo, G.; Trovisco, V.; et al. FlyBase 2.0: The next generation. Nucleic Acids Res. 2018, 47, D759–D765. [Google Scholar] [CrossRef] [Green Version]
- Potter, S.S.; Brorein, W.J., Jr.; Dunsmuir, P.; Rubin, G.M. Transposition of elements of the 412, copia and 297 dispersed repeated gene families in Drosophila. Cell 1979, 17, 415–427. [Google Scholar] [CrossRef]
- Strobel, E.; Dunsmuir, P.; Rubin, G.M. Polymorphisms in the chromosomal locations of elements of the 412, copia and 297 dispersed repeated gene families in Drosophila. Cell 1979, 17, 429–439. [Google Scholar] [CrossRef]
- Engels, W.R. Extrachromosomal control of mutability in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 1979, 76, 4011–4015. [Google Scholar] [CrossRef] [Green Version]
- Rubin, G.M.; Kidwell, M.G.; Bingham, P.M. The molecular basis of P-M hybrid dysgenesis: The nature of induced mutations. Cell 1982, 29, 987–994. [Google Scholar] [CrossRef]
- Bingham, P.M.; Kidwell, M.G.; Rubin, G.M. The molecular basis of P-M hybrid dysgenesis: The role of the P element, a P-strain-specific transposon family. Cell 1982, 29, 995–1004. [Google Scholar] [CrossRef]
- Rasmuson, B.; Montell, I.; Rasmuson, A.; Svahlin, H.; Westerberg, B.M. Genetic instability in Drosophila melanogaster: Evidence for regulation, excision and transposition at the white locus. Mol. Gen. Genet. 1980, 177, 567–570. [Google Scholar] [CrossRef] [PubMed]
- Ilyin, Y.V.; Tchurikov, N.A.; Ananiev, E.V.; Ryskov, A.P.; Yenikolopov, G.N.; Limborska, S.A.; Maleeva, N.E.; Gvozdev, V.A.; Georgiev, G.P. Studies on the DNA fragments of mammals and Drosophila containing structural genes and adjacent sequences. Cold Spring Harb. Symp. Quant. Biol. 1978, 42, 959–969. [Google Scholar] [CrossRef] [PubMed]
- Spradling, A.C.; Rubin, G.M. Transposition of cloned P elements into Drosophila germ line chromosomes. Science 1982, 218, 341–347. [Google Scholar] [CrossRef] [Green Version]
- Rubin, G.M.; Spradling, A.C. Genetic transformation of Drosophila with transposable element vectors. Science 1982, 218, 348–353. [Google Scholar] [CrossRef] [Green Version]
- Pavlopoulos, A.; Oehler, S.; Kapetanaki, M.G.; Savakis, C. The DNA transposon Minos as a tool for transgenesis and functional genomic analysis in vertebrates and invertebrates. Genome Biol. 2007, 8 (Suppl. 1), S2. [Google Scholar] [CrossRef] [Green Version]
- Kaminker, J.S.; Bergman, C.M.; Kronmiller, B.; Carlson, J.; Svirskas, R.; Patel, S.; Frise, E.; Wheeler, D.A.; Lewis, S.E.; Rubin, G.M.; et al. The transposable elements of the Drosophila melanogaster euchromatin: A genomics perspective. Genome Biol. 2002, 3, RESEARCH0084. [Google Scholar] [CrossRef] [Green Version]
- Marsano, R.M.; Marconi, S.; Moschetti, R.; Barsanti, P.; Caggese, C.; Caizzi, R. MAX, a novel retrotransposon of the BEL-Pao family, is nested within the Bari1 cluster at the heterochromatic h39 region of chromosome 2 in Drosophila melanogaster. Mol. Genet. Genomics 2004, 270, 477–484. [Google Scholar] [CrossRef]
- Zanni, V.; Eymery, A.; Coiffet, M.; Zytnicki, M.; Luyten, I.; Quesneville, H.; Vaury, C.; Jensen, S. Distribution, evolution, and diversity of retrotransposons at the flamenco locus reflect the regulatory properties of piRNA clusters. Proc. Natl. Acad. Sci. USA 2013, 110, 19842–19847. [Google Scholar] [CrossRef] [Green Version]
- Desset, S.; Conte, C.; Dimitri, P.; Calco, V.; Dastugue, B.; Vaury, C. Mobilization of two retroelements, ZAM and Idefix, in a novel unstable line of Drosophila melanogaster. Mol. Biol. Evol. 1999, 16, 54–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drosophila 12 Genomes Consortium. Evolution of genes and genomes on the Drosophila phylogeny. Nature 2007, 450, 203–218. [Google Scholar] [CrossRef] [PubMed]
- Seetharam, A.S.; Stuart, G.W. Whole genome phylogeny for 21 Drosophila species using predicted 2b-RAD fragments. PeerJ 2013, 1, e226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palazzo, A.; Lovero, D.; D’Addabbo, P.; Caizzi, R.; Marsano, R.M. Identification of Bari Transposons in 23 Sequenced Drosophila Genomes Reveals Novel Structural Variants, MITEs and Horizontal Transfer. PLoS ONE 2016, 11, e0156014. [Google Scholar] [CrossRef]
- Bargues, N.; Lerat, E. Evolutionary history of LTR-retrotransposons among 20 Drosophila species. Mob. DNA 2017, 8, 7. [Google Scholar] [CrossRef] [Green Version]
- Loreto, E.L.S.; Depra, M.; Diesel, J.F.; Panzera, Y.; Valente-Gaiesky, V.L.S. Drosophila relics hobo and hobo-MITEs transposons as raw material for new regulatory networks. Genet Mol. Biol. 2018, 41, 198–205. [Google Scholar] [CrossRef] [Green Version]
- Gagnier, L.; Belancio, V.P.; Mager, D.L. Mouse germ line mutations due to retrotransposon insertions. Mob. DNA 2019, 10, 15. [Google Scholar] [CrossRef] [Green Version]
- Pritham, E.J.; Feschotte, C. Massive amplification of rolling-circle transposons in the lineage of the bat Myotis lucifugus. Proc. Natl. Acad. Sci. USA 2007, 104, 1895–1900. [Google Scholar] [CrossRef] [Green Version]
- Ray, D.A.; Feschotte, C.; Pagan, H.J.; Smith, J.D.; Pritham, E.J.; Arensburger, P.; Atkinson, P.W.; Craig, N.L. Multiple waves of recent DNA transposon activity in the bat, Myotis lucifugus. Genome Res. 2008, 18, 717–728. [Google Scholar] [CrossRef] [Green Version]
- Ray, D.A.; Pagan, H.J.; Thompson, M.L.; Stevens, R.D. Bats with hATs: Evidence for recent DNA transposon activity in genus Myotis. Mol. Biol. Evol. 2007, 24, 632–639. [Google Scholar] [CrossRef]
- Pagan, H.J.; Macas, J.; Novak, P.; McCulloch, E.S.; Stevens, R.D.; Ray, D.A. Survey sequencing reveals elevated DNA transposon activity, novel elements, and variation in repetitive landscapes among vesper bats. Genome Biol. Evol. 2012, 4, 575–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, H.; Bogwitz, M.R.; McCart, C.; Andrianopoulos, A.; Ffrench-Constant, R.H.; Batterham, P.; Daborn, P.J. Cis-regulatory elements in the Accord retrotransposon result in tissue-specific expression of the Drosophila melanogaster insecticide resistance gene Cyp6g1. Genetics 2007, 175, 1071–1077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlenke, T.A.; Begun, D.J. Strong selective sweep associated with a transposon insertion in Drosophila simulans. Proc. Natl. Acad. Sci. USA 2004, 101, 1626–1631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clemmons, A.W.; Wasserman, S.A. Combinatorial effects of transposable elements on gene expression and phenotypic robustness in Drosophila melanogaster development. G3 Genes Genomes Genet. 2013, 3, 1531–1538. [Google Scholar]
- Zhang, L.; Beaucher, M.; Cheng, Y.; Rong, Y.S. Coordination of transposon expression with DNA replication in the targeting of telomeric retrotransposons in Drosophila. EMBO J. 2014, 33, 1148–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- George, J.A.; Traverse, K.L.; DeBaryshe, P.G.; Kelley, K.J.; Pardue, M.L. Evolution of diverse mechanisms for protecting chromosome ends by Drosophila TART telomere retrotransposons. Proc. Natl. Acad. Sci. USA 2010, 107, 21052–21057. [Google Scholar] [CrossRef] [Green Version]
- Abad, J.P.; De Pablos, B.; Osoegawa, K.; De Jong, P.J.; Martin-Gallardo, A.; Villasante, A. TAHRE, a novel telomeric retrotransposon from Drosophila melanogaster, reveals the origin of Drosophila telomeres. Mol. Biol. Evol. 2004, 21, 1620–1624. [Google Scholar] [CrossRef]
- Conte, C.; Dastugue, B.; Vaury, C. Coupling of enhancer and insulator properties identified in two retrotransposons modulates their mutagenic impact on nearby genes. Mol. Cell Biol. 2002, 22, 1767–1777. [Google Scholar] [CrossRef] [Green Version]
- Caggese, C.; Barsanti, P.; Viggiano, L.; Bozzetti, M.P.; Caizzi, R. Genetic, molecular and developmental analysis of the glutamine synthetase isozymes of Drosophila melanogaster. Genetica 1994, 94, 275–281. [Google Scholar] [CrossRef]
- Minervini, C.F.; Marsano, R.M.; Casieri, P.; Fanti, L.; Caizzi, R.; Pimpinelli, S.; Rocchi, M.; Viggiano, L. Heterochromatin protein 1 interacts with 5’UTR of transposable element ZAM in a sequence-specific fashion. Gene 2007, 393, 1–10. [Google Scholar] [CrossRef]
- Wilson, S.; Matyunina, L.V.; McDonald, J.F. An enhancer region within the copia untranslated leader contains binding sites for Drosophila regulatory proteins. Gene 1998, 209, 239–246. [Google Scholar] [CrossRef]
- Ding, D.; Lipshitz, H.D. Spatially regulated expression of retrovirus-like transposons during Drosophila melanogaster embryogenesis. Genet. Res. 1994, 64, 167–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zatsepina, O.G.; Velikodvorskaia, V.V.; Molodtsov, V.B.; Garbuz, D.; Lerman, D.N.; Bettencourt, B.R.; Feder, M.E.; Evgenev, M.B. A Drosophila Melanogaster Strain From Sub-Equatorial Africa Has Exceptional Thermotolerance But Decreased Hsp70 Expression. J. Exp. Biol. 2001, 204, 1869. [Google Scholar] [PubMed]
- Lerman, D.N.; Feder, M.E. Naturally occurring transposable elements disrupt hsp70 promoter function in Drosophila melanogaster. Mol. Biol. Evol. 2005, 22, 776–783. [Google Scholar] [CrossRef] [PubMed]
- Kimbrell, D.A.; Tojo, S.J.; Alexander, S.; Brown, E.E.; Tobin, S.L.; Fristrom, J.W. Regulation of larval cuticle protein gene expression in Drosophila melanogaster. Dev. Genet. 1989, 10, 198–209. [Google Scholar] [CrossRef] [PubMed]
- Kimbrell, D.A.; Berger, E.; King, D.S.; Wolfgang, W.J.; Fristrom, J.W. Cuticle protein gene expression during the third instar of Drosophila melanogaster. Insect Biochem. 1988, 18, 229–235. [Google Scholar] [CrossRef]
- Gruber, J.D.; Genissel, A.; Macdonald, S.J.; Long, A.D. How repeatable are associations between polymorphisms in achaete-scute and bristle number variation in Drosophila? Genetics 2007, 175, 1987–1997. [Google Scholar] [CrossRef] [Green Version]
- Long, A.D.; Lyman, R.F.; Morgan, A.H.; Langley, C.H.; Mackay, T.F. Both naturally occurring insertions of transposable elements and intermediate frequency polymorphisms at the achaete-scute complex are associated with variation in bristle number in Drosophila melanogaster. Genetics 2000, 154, 1255–1269. [Google Scholar]
- Gonzalez, J.; Lenkov, K.; Lipatov, M.; Macpherson, J.M.; Petrov, D.A. High rate of recent transposable element-induced adaptation in Drosophila melanogaster. PLoS Biol. 2008, 6, e251. [Google Scholar] [CrossRef] [Green Version]
- Spana, C.; Harrison, D.A.; Corces, V.G. The Drosophila melanogaster suppressor of Hairy-wing protein binds to specific sequences of the gypsy retrotransposon. Genes. Dev. 1988, 2, 1414–1423. [Google Scholar] [CrossRef] [Green Version]
- Roseman, R.R.; Swan, J.M.; Geyer, P.K. A Drosophila insulator protein facilitates dosage compensation of the X chromosome min-white gene located at autosomal insertion sites. Development 1995, 121, 3573–3582. [Google Scholar] [PubMed]
- Cai, H.N.; Levine, M. The gypsy insulator can function as a promoter-specific silencer in the Drosophila embryo. EMBO J. 1997, 16, 1732–1741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brönner, G.; Taubert, H.; Jäckle, H. Mesoderm-specific B104 expression in the Drosophila embryo is mediated by internal cis-acting elements of the transposon. Chromosoma 1995, 103, 669–675. [Google Scholar] [PubMed]
- Batut, P.; Dobin, A.; Plessy, C.; Carninci, P.; Gingeras, T.R. High-fidelity promoter profiling reveals widespread alternative promoter usage and transposon-driven developmental gene expression. Genome Res. 2013, 23, 169–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, Y.; Berrocal, A.; Morita, T.; Longden, K.D.; Stern, D.L. Natural courtship song variation caused by an intronic retroelement in an ion channel gene. Nature 2016, 536, 329–332. [Google Scholar] [CrossRef]
- Merenciano, M.; Ullastres, A.; de Cara, M.A.; Barron, M.G.; Gonzalez, J. Multiple Independent Retroelement Insertions in the Promoter of a Stress Response Gene Have Variable Molecular and Functional Effects in Drosophila. PLoS Genet. 2016, 12, e1006249. [Google Scholar] [CrossRef]
- Tanda, S.; Corces, V.G. Retrotransposon-induced overexpression of a homeobox gene causes defects in eye morphogenesis in Drosophila. EMBO J. 1991, 10, 407–417. [Google Scholar] [CrossRef]
- Cherkasova, V.A.; IuV, I. The leader region of the Drosophila transposon MDG1 contains transcription termination sites. Genetika 1990, 26, 1893–1904. [Google Scholar]
- Nabirochkin, S.; Ossokina, M.; Heidmann, T. A nuclear matrix/scaffold attachment region co-localizes with the gypsy retrotransposon insulator sequence. J. Biol. Chem. 1998, 273, 2473–2479. [Google Scholar] [CrossRef] [Green Version]
- Mamillapalli, A.; Pathak, R.U.; Garapati, H.S.; Mishra, R.K. Transposable element ‘roo’ attaches to nuclear matrix of the Drosophila melanogaster. J. Insect. Sci. 2013, 13, 111. [Google Scholar] [CrossRef]
- Ellison, C.E.; Bachtrog, D. Dosage compensation via transposable element mediated rewiring of a regulatory network. Science 2013, 342, 846–850. [Google Scholar] [CrossRef] [Green Version]
- Mateo, L.; Ullastres, A.; Gonzalez, J. A transposable element insertion confers xenobiotic resistance in Drosophila. PLoS Genet. 2014, 10, e1004560. [Google Scholar] [CrossRef] [Green Version]
- Guio, L.; Barron, M.G.; Gonzalez, J. The transposable element Bari-Jheh mediates oxidative stress response in Drosophila. Mol Ecol. 2014, 23, 2020–2030. [Google Scholar] [CrossRef] [PubMed]
- Guio, L.; Vieira, C.; Gonzalez, J. Stress affects the epigenetic marks added by natural transposable element insertions in Drosophila melanogaster. Sci. Rep. 2018, 8, 12197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Steensel, B.; Delrow, J.; Henikoff, S. Chromatin profiling using targeted DNA adenine methyltransferase. Nat. Genet. 2001, 27, 304–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marsano, R.M.; Caizzi, R.; Moschetti, R.; Junakovic, N. Evidence for a functional interaction between the Bari1 transposable element and the cytochrome P450 cyp12a4 gene in Drosophila melanogaster. Gene 2005, 357, 122–128. [Google Scholar] [CrossRef]
- Sentmanat, M.F.; Elgin, S.C. Ectopic assembly of heterochromatin in Drosophila melanogaster triggered by transposable elements. Proc. Natl. Acad. Sci. USA 2012, 109, 14104–14109. [Google Scholar] [CrossRef] [Green Version]
- Deprá, M.; da Silva Valente, V.L.; Margis, R.; Loreto, E.L.S. The hobo transposon and hobo-related elements are expressed as developmental genes in Drosophila. Gene 2009, 448, 57–63. [Google Scholar] [CrossRef]
- Maside, X.; Bartolome, C.; Charlesworth, B. S-element insertions are associated with the evolution of the Hsp70 genes in Drosophila melanogaster. Curr. Biol. 2002, 12, 1686–1691. [Google Scholar] [CrossRef] [Green Version]
- Schilbach, S.; Hantsche, M.; Tegunov, D.; Dienemann, C.; Wigge, C.; Urlaub, H.; Cramer, P. Structures of transcription pre-initiation complex with TFIIH and Mediator. Nature 2017, 551, 204–209. [Google Scholar] [CrossRef]
- Palazzo, A.; Caizzi, R.; Viggiano, L.; Marsano, R.M. Does the Promoter Constitute a Barrier in the Horizontal Transposon Transfer Process? Insight from Bari Transposons. Genome Biol. Evol. 2017, 9, 1637–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palazzo, A.; Lorusso, P.; Miskey, C.; Walisko, O.; Gerbino, A.; Marobbio, C.M.T.; Ivics, Z.; Marsano, R.M. Transcriptionally promiscuous “blurry” promoters in Tc1/mariner transposons allow transcription in distantly related genomes. Mob. DNA 2019, 10, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russo, J.; Harrington, A.W.; Steiniger, M. Antisense Transcription of Retrotransposons in Drosophila: An Origin of Endogenous Small Interfering RNA Precursors. Genetics 2016, 202, 107–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrington, A.W.; Steiniger, M. Bioinformatic analyses of sense and antisense expression from terminal inverted repeat transposons in Drosophila somatic cells. Fly 2016, 10, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Villanueva-Canas, J.L.; Horvath, V.; Aguilera, L.; Gonzalez, J. Diverse families of transposable elements affect the transcriptional regulation of stress-response genes in Drosophila melanogaster. Nucleic Acids. Res. 2019, 47, 6842–6857. [Google Scholar] [CrossRef] [Green Version]
- Marsano, R.M.; Giordano, E.; Messina, G.; Dimitri, P. A New Portrait of Constitutive Heterochromatin: Lessons from Drosophila melanogaster. Trends Genet. 2019, 35, 615–631. [Google Scholar] [CrossRef]
- Yasuhara, J.C.; DeCrease, C.H.; Wakimoto, B.T. Evolution of heterochromatic genes of Drosophila. Proc. Natl. Acad. Sci. USA 2005, 102, 10958–10963. [Google Scholar] [CrossRef] [Green Version]
- Sneddon, A.; Flavell, A.J. The transcriptional control regions of the copia retrotransposon. Nucleic Acids Res. 1989, 17, 4025–4035. [Google Scholar] [CrossRef] [Green Version]
- Holdridge, C.; Dorsett, D. Repression of hsp70 heat shock gene transcription by the suppressor of hairy-wing protein of Drosophila melanogaster. Mol Cell Biol. 1991, 11, 1894–1900. [Google Scholar] [CrossRef] [Green Version]
- Scott, K.C.; Taubman, A.D.; Geyer, P.K. Enhancer blocking by the Drosophila gypsy insulator depends upon insulator anatomy and enhancer strength. Genetics 1999, 153, 787–798. [Google Scholar]
- Daborn, P.J.; Yen, J.L.; Bogwitz, M.R.; Le Goff, G.; Feil, E.; Jeffers, S.; Tijet, N.; Perry, T.; Heckel, D.; Batterham, P.; et al. A single p450 allele associated with insecticide resistance in Drosophila. Science 2002, 297, 2253–2256. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Bai, S.; Cass, B.N. Accord insertion in the 5’ flanking region of CYP6G1 confers nicotine resistance in Drosophila melanogaster. Gene 2012, 502, 1–8. [Google Scholar] [CrossRef]
- Bire, S.; Casteret, S.; Piegu, B.; Beauclair, L.; Moire, N.; Arensbuger, P.; Bigot, Y. Mariner Transposons Contain a Silencer: Possible Role of the Polycomb Repressive Complex 2. PLoS Genet. 2016, 12, e1005902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minervini, C.F.; Ruggieri, S.; Traversa, M.; D’Aiuto, L.; Marsano, R.M.; Leronni, D.; Centomani, I.; Giovanni, D.C.; Viggiano, L. Evidences for insulator activity of the 5’UTR of the Drosophila melanogaster LTR-retrotransposon ZAM. Mol. Genet. Genom. 2010, 283, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Sorek, R. The birth of new exons: Mechanisms and evolutionary consequences. RNA 2007, 13, 1603–1608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Treiber, C.D.; Waddell, S. Transposon expression in the Drosophila brain is driven by neighboring genes and diversifies the neural transcriptome. bioRxiv 2019, 838045. [Google Scholar] [CrossRef] [Green Version]
- Laski, F.A.; Rio, D.C.; Rubin, G.M. Tissue specificity of Drosophila P element transposition is regulated at the level of mRNA splicing. Cell 1986, 44, 7–19. [Google Scholar] [CrossRef]
- Palazzo, A.; Moschetti, R.; Caizzi, R.; Marsano, R.M. The Drosophila mojavensis Bari3 transposon: Distribution and functional characterization. Mob. DNA 2014, 5, 21. [Google Scholar] [CrossRef] [Green Version]
- Palazzo, A.; Marconi, S.; Specchia, V.; Bozzetti, M.P.; Ivics, Z.; Caizzi, R.; Massimiliano, R.M. Functional characterization of the Bari1 transposition system. PLoS ONE 2013, 8, e79385. [Google Scholar] [CrossRef] [Green Version]
- Dorer, D.R.; Henikoff, S. Expansions of transgene repeats cause heterochromatin formation and gene silencing in Drosophila. Cell 1994, 77, 993–1002. [Google Scholar] [CrossRef]
- Tartof, K.D.; Hobbs, C.; Jones, M. A structural basis for variegating position effects. Cell 1984, 37, 869–878. [Google Scholar] [CrossRef]
- Ryu, H.W.; Lee, D.H.; Florens, L.; Swanson, S.K.; Washburn, M.P.; Kwon, S.H. Analysis of the heterochromatin protein 1 (HP1) interactome in Drosophila. J. Proteom. 2014, 102, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Greil, F.; van der Kraan, I.; Delrow, J.; Smothers, J.F.; de Wit, E.; Bussemaker, H.J.; van Driel, R.; Henikoff, S.; van Steensel, B. Distinct HP1 and Su(var)3-9 complexes bind to sets of developmentally coexpressed genes depending on chromosomal location. Genes Dev. 2003, 17, 2825–2838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinemann, M.; Steinemann, S. Degenerating Y chromosome of Drosophila miranda: A trap for retrotransposons. Proc. Natl. Acad. Sci. USA 1992, 89, 7591–7595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frydrychova, R.C.; Biessmann, H.; Mason, J.M. Regulation of telomere length in Drosophila. Cytogenet. Genome Res. 2008, 122, 356–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.H.; Chavan, A.; Palladino, J.; Wei, X.; Martins, N.M.C.; Santinello, B.; Chen, C.C.; Erceg, J.; Wu, C.T.; Larracuente, A.M.; et al. Islands of retroelements are major components of Drosophila centromeres. PLoS Biol. 2019, 17, e3000241. [Google Scholar] [CrossRef] [Green Version]
- Brennecke, J.; Aravin, A.A.; Stark, A.; Dus, M.; Kellis, M.; Sachidanandam, R.; Hannon, G.J. Discrete small RNA-generating loci as master regulators of transposon activity in Drosophila. Cell 2007, 128, 1089–1103. [Google Scholar] [CrossRef] [Green Version]
- Fingerhut, J.M.; Moran, J.V.; Yamashita, Y.M. Satellite DNA-containing gigantic introns in a unique gene expression program during Drosophila spermatogenesis. PLoS Genet. 2019, 15, e1008028. [Google Scholar] [CrossRef] [Green Version]
- Perrat, P.N.; DasGupta, S.; Wang, J.; Theurkauf, W.; Weng, Z.; Rosbash, M.; Waddell, S. Transposition-driven genomic heterogeneity in the Drosophila brain. Science 2013, 340, 91–95. [Google Scholar] [CrossRef] [Green Version]
- Treiber, C.D.; Waddell, S. Resolving the prevalence of somatic transposition in Drosophila. Elife 2017, 6, e28297. [Google Scholar] [CrossRef] [Green Version]
- Baillie, J.K.; Barnett, M.W.; Upton, K.R.; Gerhardt, D.J.; Richmond, T.A.; De Sapio, F.; Bernnan, P.M.; Rizzu, P.; Smith, S.; Fell, M.; et al. Somatic retrotransposition alters the genetic landscape of the human brain. Nature 2011, 479, 534–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Upton, K.R.; Gerhardt, D.J.; Jesuadian, J.S.; Richardson, S.R.; Sanchez-Luque, F.J.; Bodea, G.O.; Ewing, A.D.; Salvador-Palomeque, C.; van der Knaap, M.S.; Brennan, P.M.; et al. Ubiquitous L1 mosaicism in hippocampal neurons. Cell 2015, 161, 228–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.I.; Lyttle, T.W.; Wu, M.L.; Lin, G.F. Association between a satellite DNA sequence and the Responder of Segregation Distorter in D. melanogaster. Cell 1988, 54, 179–189. [Google Scholar] [CrossRef]
- Caggese, C.; Pimpinelli, S.; Barsanti, P.; Caizzi, R. The distribution of the transposable element Bari-1 in the Drosophila melanogaster and Drosophila simulans genomes. Genetica 1995, 96, 269–283. [Google Scholar] [CrossRef]
- Port, F.; Strein, C.; Stricker, M.; Rauscher, B.; Heigwer, F.; Zhou, J.; Beyersdörffer, C.; Frei, J.; Hess, A.; Kern, K.; et al. A large-scale resource for tissue-specific CRISPR mutagenesis in Drosophila. bioRxiv. 2019, 636076. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species | Affected Gene/Locus | TE | Transposon Type | Cis-Regulatory Activity | Effect | Evidences | Reference |
|---|---|---|---|---|---|---|---|
| Dmel | Cyp6g1 | Accord | LTR | Enhancer | Increased xenobiotic resistance | Reporter Assay | [42] |
| Dsim | Cyp6g1 | Doc | non-LTR | Enhancer(?) | Increased xenobiotic resistance | DDT resistance and gene over-expression | [43] |
| Dmel | bxd1 | gypsy | LTR | enhancer | Development of thoracic segment | Phenotype assay | [44] |
| Dmel | Telomeres | HeT-A | non-LTR | Telomere elongation | Telomere maintainance | In vivo assay | [45] |
| Dmel Dvir | Telomeres | TART | non-LTR | Telomere elongation | Telomere maintainance | In vivo assay | [46] |
| Dmel | Telomeres | TAHRE | non-LTR | Telomere elongation | Telomere maintainance | DNA sequencing | [47] |
| Dmel | white | Idefix | LTR | insulator | Eye pigmentation | Phenotype assay | [48] |
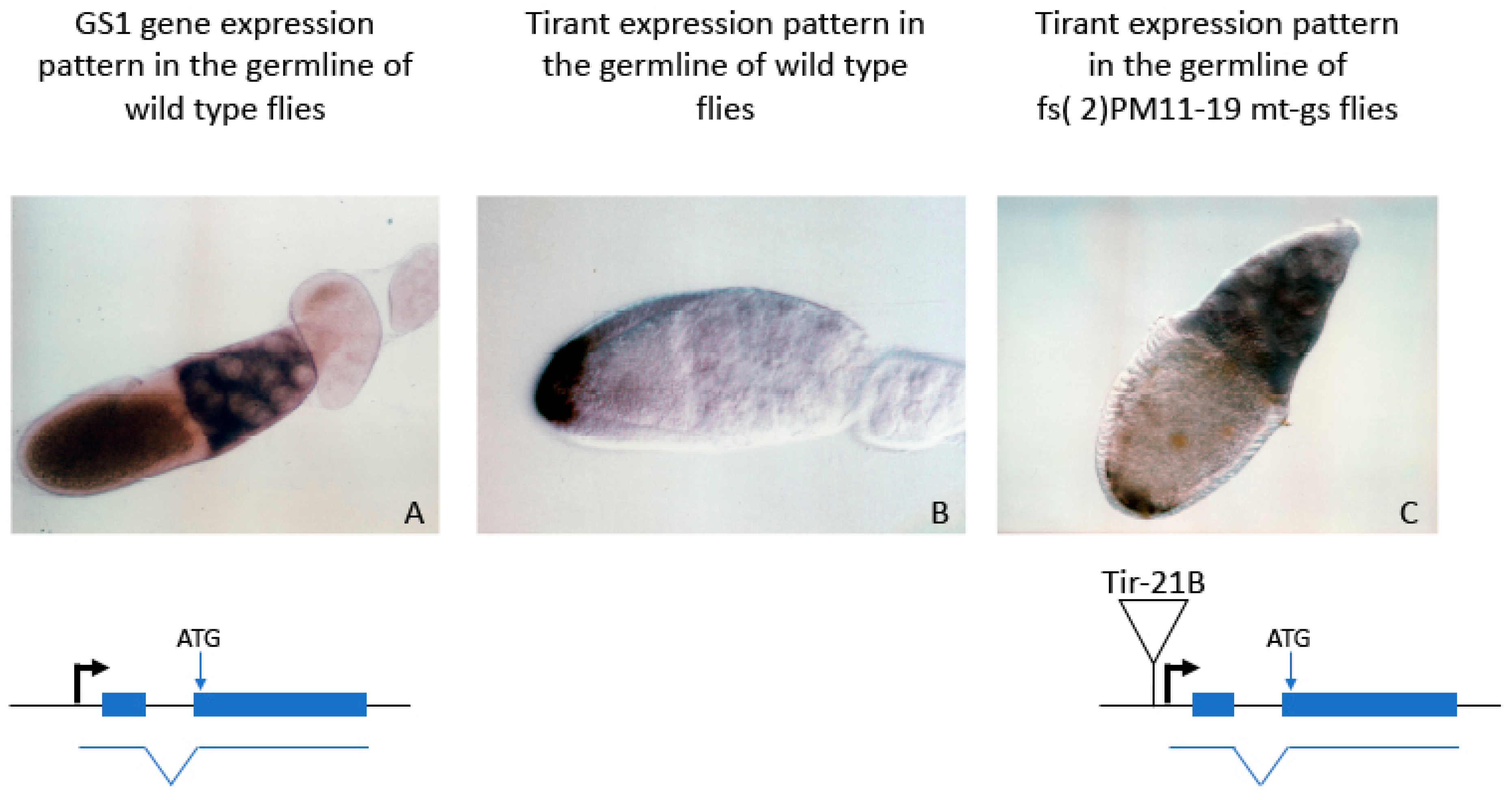
| Dmel | GS1 | Tirant | LTR | ND | ND | Direct assay | [49] |
| Dmel | NA | ZAM | LTR | HP1 binding | Chromatin state determination | In vitro assay | [50] |
| Dmel | NA | copia | LTR | Enhancer | Reporter expression | Direct assay | [51] |
| Dmel | development genes | 17.6, 297, 412, 1731, 3S18, blood, copia, gypsy, HMS Beagle Kermit/fleamdg1 mdg3 opus B104/roo springer | LTR | Cis-regulatory sequences | alterations of gene expression during embryogenesis | Expression analyses | [52] |
| Dmel | Hsp70Ba | jockey | LTR | Cis-regulatory sequences | suppression of the deleterious phenotypes of Hsp70. | Phenotypic assay, expression analysis | [53,54] |
| Dmel | 87A7 hsp70 | H.M.S. Beagle | LTR | Unknown | suppression of the deleterious phenotypes of Hsp70. | Phenotypic assay, expression analysis | [53] |
| Dmel | LCP-1 LCP-f2 | H.M.S. Beagle | LTR | enhancer-like elements | Transcriptional activation of LCP genes | Genetic variants analyses | [55] [56] |
| Dmel | achaete-scute complex | transpac | LTR | enhancer-like elements | variation in bristle number | Genetic variants analyses | [57] [58] |
| Dmel | kuz | F-element | non-LTR | cis-regulatory | Gene up- regulation | Population analyses | [59] |
| Dmel | yellow | gypsy | LTR | insulator | Yellow phenotype | In vivo analyses | [60] [61] [62] |
| Dmel | NA | B104/roo | LTR | promoter | NA | Inferred from in vivo assay | [63] |
| Dmel | TM4SF | 297 | LTR | promoter | NA | RNA ligase-mediated 5′-RACE | [64] |
| Dmel | 152 annotated genes | roo gypsy Pao | LTR | Promoter | NA | RNA ligase-mediated 5′-RACE | [64] |
| Dsim | Slowpoke | Shellder | LTR | Altered splicing | Courtship song variation | Trait mapping, in vivo CRISPR knockout | [65] |
| Dmel | CG18446 | roo | LTR | alternative transcription start site | increased expression | 5′-RACE | [66] |
| Dana | Om(10) | TOM | LTR | enhancers | Eye morphogenesis | In vivo assay | [67] |
| Dmel | NA | MDG1 | LTR | Transcription termination | NA | Transcriptional analysis | [68] |
| Dmel | NA | gypsy | LTR | MAR | NA | In vivo assay | [69] |
| Dmel | NA | roo | LTR | MAR | NA | In vivo assay | [70] |
| Dmir | Neo X | ISY | Helitron | MSL binding site | Dosage compensation | Direct assay | [71] |
| Dmel | HSP70BA | P-element | DNA | Silencer | Reduction of Hsp70 expression level. | Direct phenotypic assay | [54] [53] |
| Dmel | CG11699 | POGON1 | TIR | Poly-A signal | Increased xenobiotic resistance | 3′ RACE | [72] |
| Dmel | Jheh1, Jheh2 | Bari1 | TIR | HP1 seeding | Antioxidant response | Phenotypic assay | [73] [74] |
| Dmel | h39 region | Bari1 | TIR | HP1 binding | Chromatin state determination | Direct assay | [75] |
| Dmel | Cyp12a4 | Bari1 | TIR | polyA signal | detoxification | 3′ RACE | [76] |
| Dmel | NA | 1360/hoppel | TIR | Hp1 recruitment | Heterochromatin formation | In vivo assay | [77] |
| Dsim | hunchback even-skipped | hoboVA | TIR | Promoter, transcription factor binding sites (TFBSs) | new phenotype | Expression and in situ analyses | [36,78] |
| Dmel | Hsp70Bb | S-element | TIR | cis-regulatory | NA | population genetics study | [79] |
| Dmel | rdx | S-element | TIR | cis-regulatory | down-regulation | Population analysis | [59] |
| Dmel | 152 annotated genes | Tc1 P hAT Helitron | TIR Helitron | Promoter TSS clusters | NA | RNA ligase-mediated 5′-RACE | [64] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moschetti, R.; Palazzo, A.; Lorusso, P.; Viggiano, L.; Massimiliano Marsano, R. “What You Need, Baby, I Got It”: Transposable Elements as Suppliers of Cis-Operating Sequences in Drosophila. Biology 2020, 9, 25. https://0-doi-org.brum.beds.ac.uk/10.3390/biology9020025
Moschetti R, Palazzo A, Lorusso P, Viggiano L, Massimiliano Marsano R. “What You Need, Baby, I Got It”: Transposable Elements as Suppliers of Cis-Operating Sequences in Drosophila. Biology. 2020; 9(2):25. https://0-doi-org.brum.beds.ac.uk/10.3390/biology9020025
Chicago/Turabian StyleMoschetti, Roberta, Antonio Palazzo, Patrizio Lorusso, Luigi Viggiano, and René Massimiliano Marsano. 2020. "“What You Need, Baby, I Got It”: Transposable Elements as Suppliers of Cis-Operating Sequences in Drosophila" Biology 9, no. 2: 25. https://0-doi-org.brum.beds.ac.uk/10.3390/biology9020025