Cell Membrane-Interrupting Antimicrobial Peptides from Isatis indigotica Fortune Isolated by a Bacillus subtilis Expression System

 , and
, and 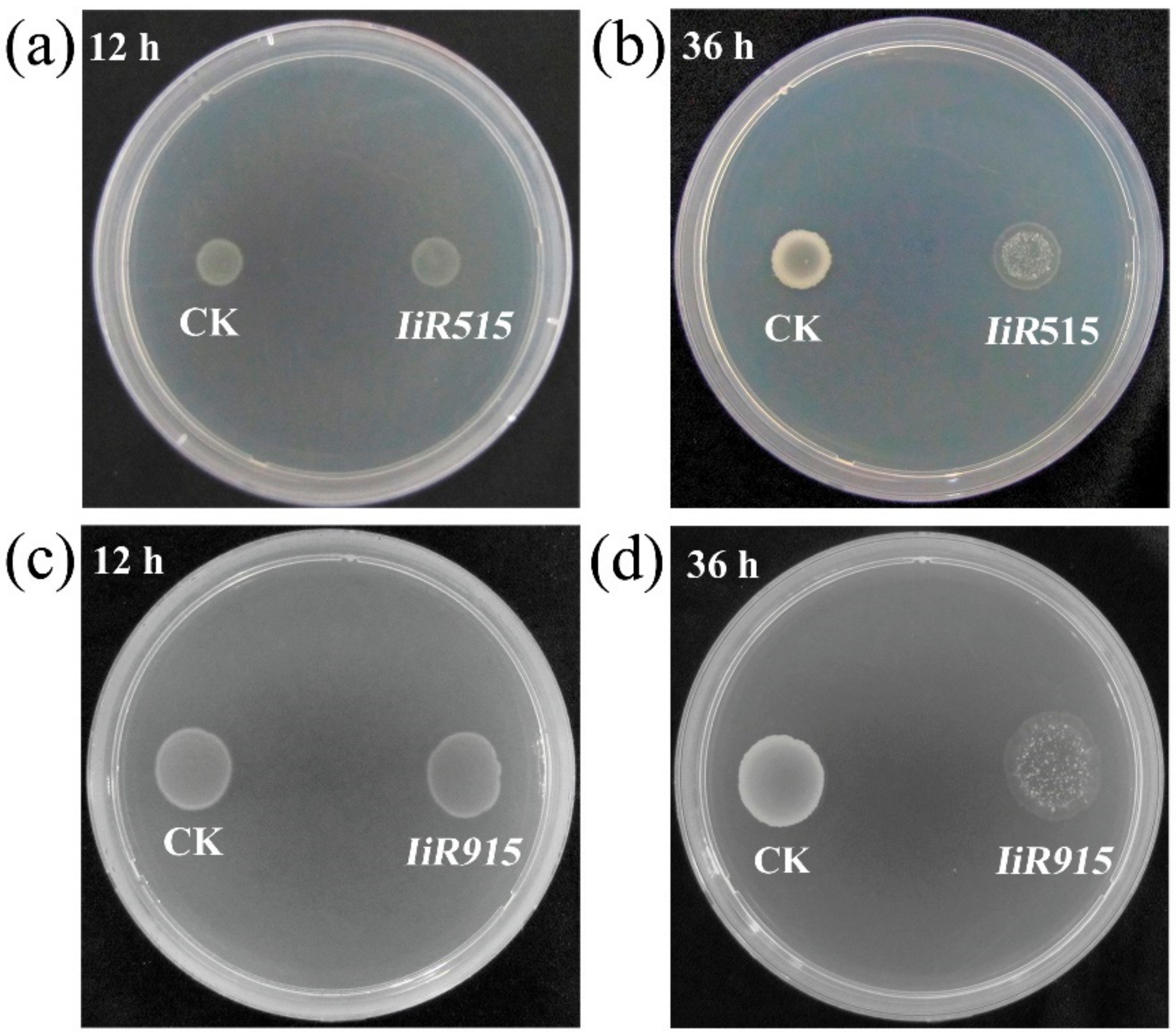{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Candidate Genes from an Isatis indigotica cDNA Library Exhibited Antimicrobial Potential
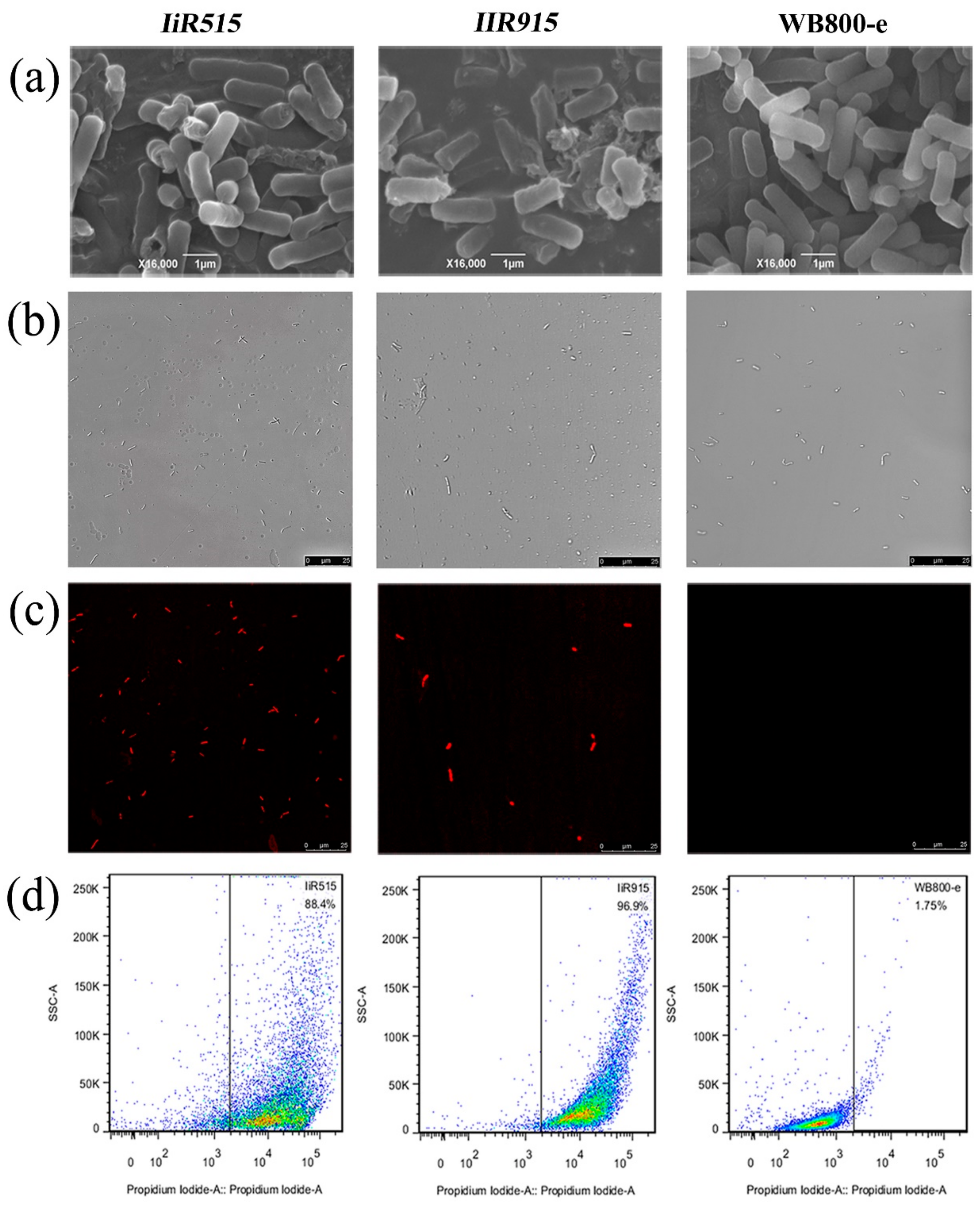
2.2. Candidate Antimicrobial Peptides Destroyed the Cell Membrane of B. subtilis
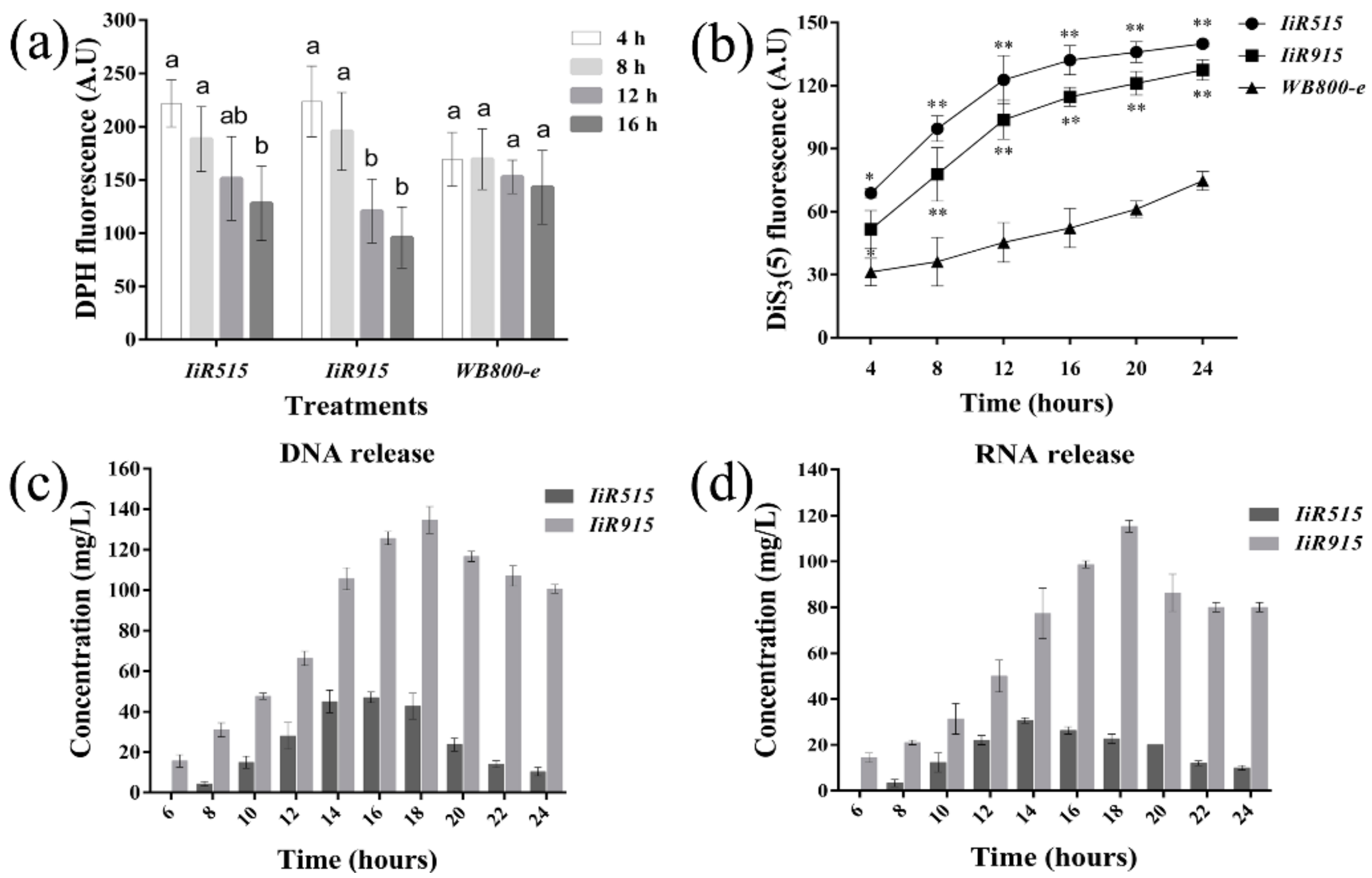
2.3. Candidate Antimicrobialpeptides Altered the Fluidity and Electrical Potential of the Bacillus subtilis Cytoplasmic Membrane
2.4. The Detection of DNA/RNA in Shaking Media Indicated Cell Membrane Breakage
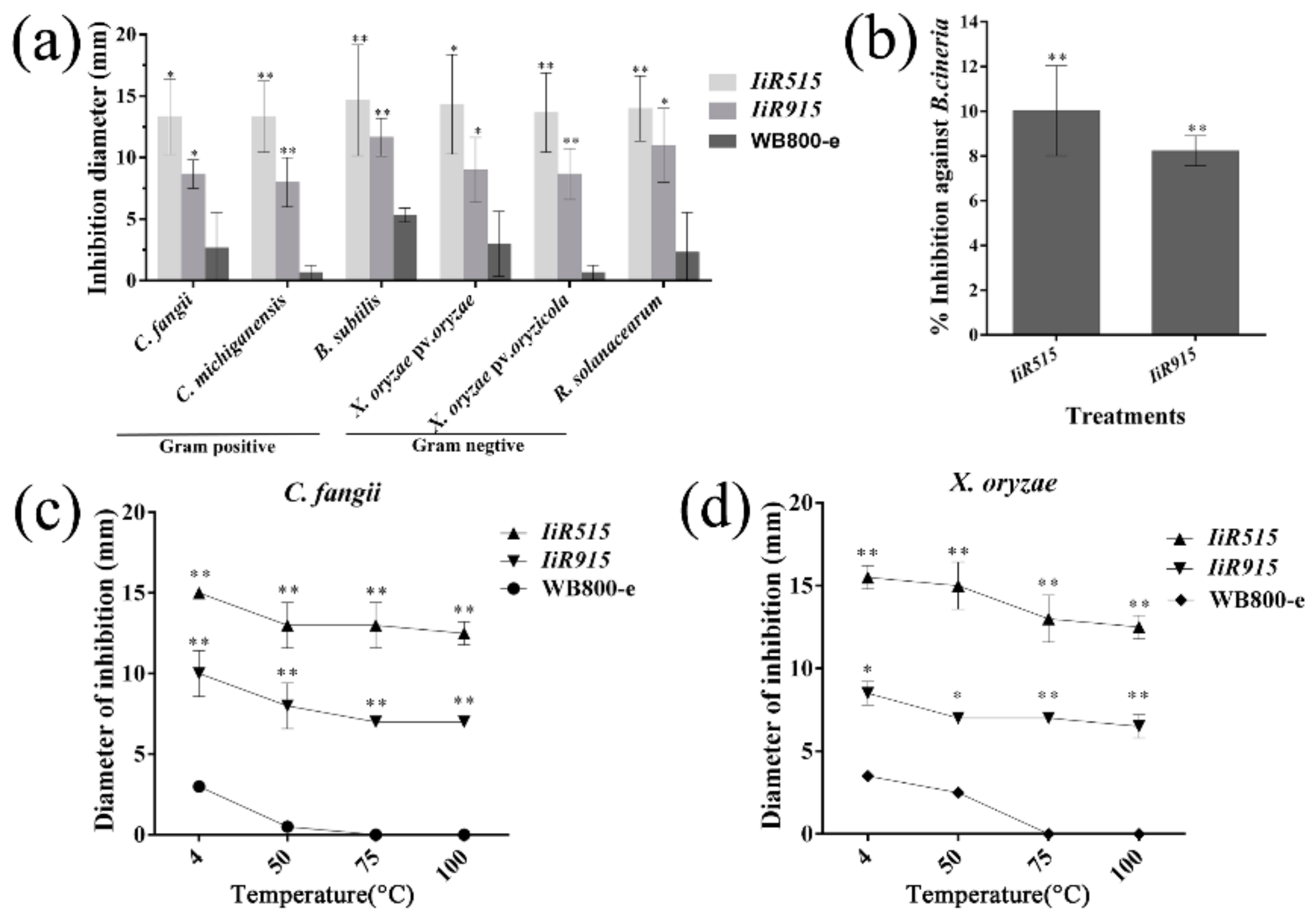
2.5. Extracellular Peptides of IiR515 and IiR915 Exhibited Antimicrobial Activities
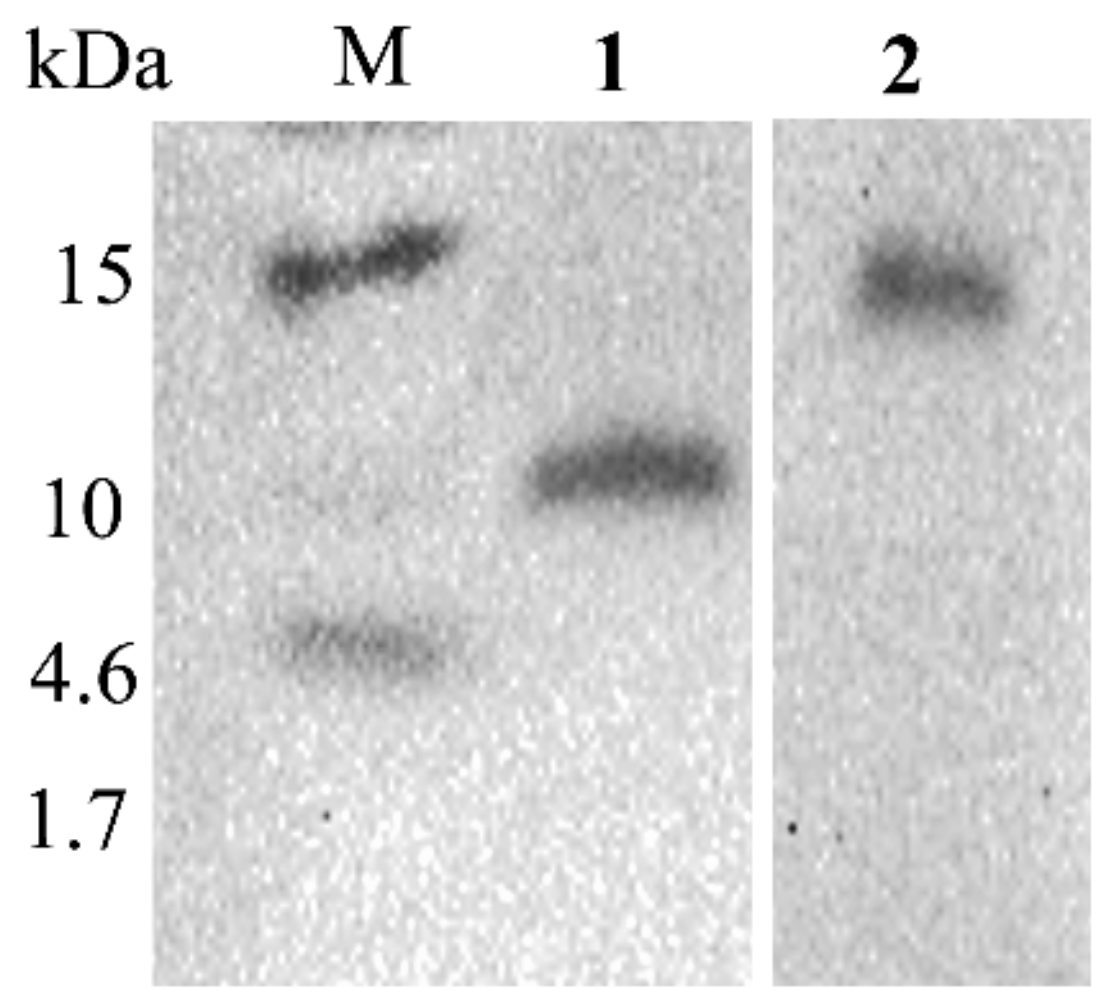
2.6. Western Blots Revealed the Expression and Size of the IiR515 and IiR915 Peptides
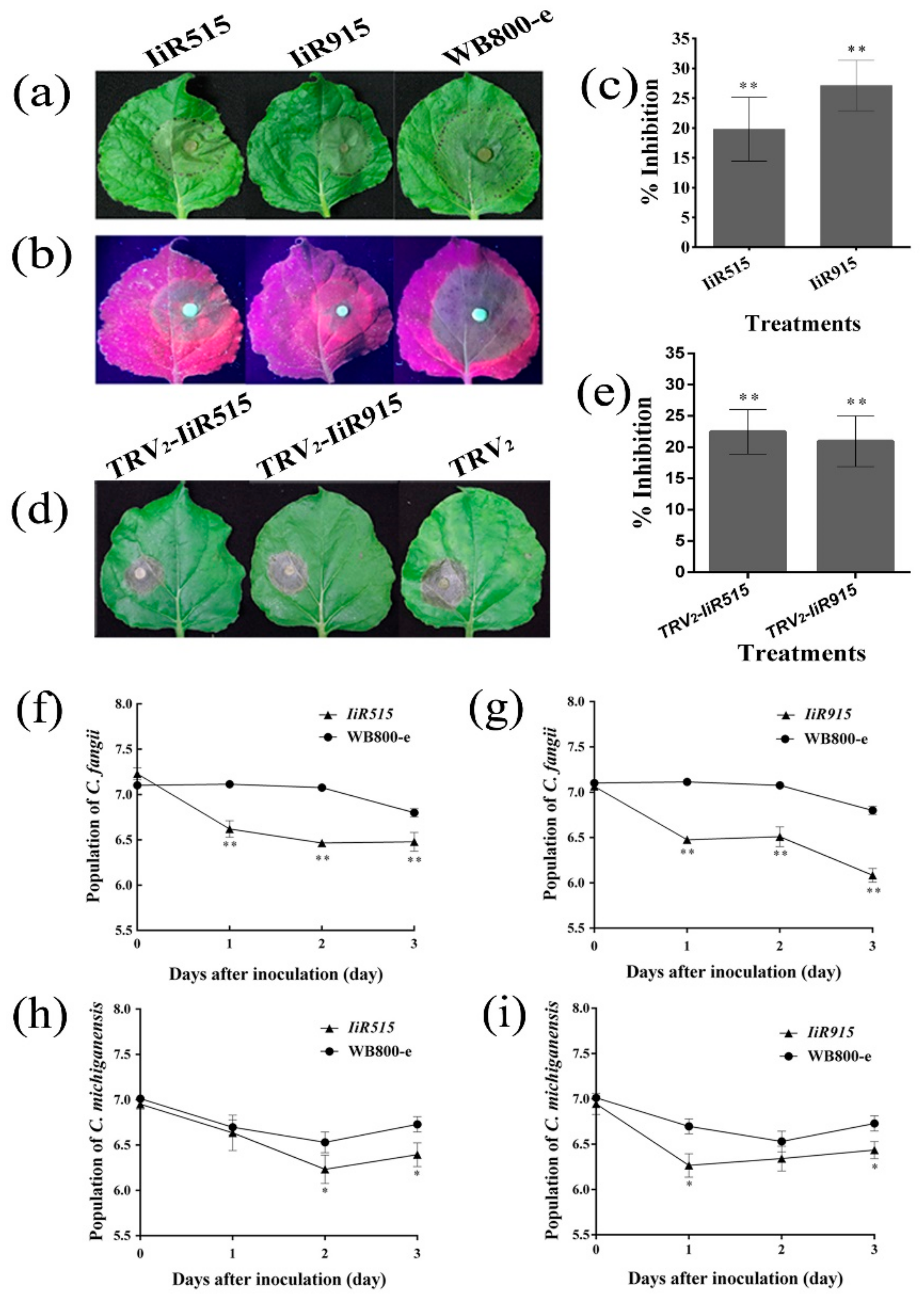
2.7. IiR515 and IiR915 Prevented Phytophthora capsici and Botrytis cinerea Infection on Detached Leaves of Nicotiana benthamiana
2.8. IiR515- and IiR915-Transformed Bacillus subtilis Inhibited the Growth of Soil-Borne Pathogens
2.9. IiR515 and IiR915 Peptides Showed No Significant Hemolytic Activity
3. Materials and Methods
3.1. Plant Materials and Pathogen Cultures
3.2. Isatis indigotica cDNA Library Construction
3.3. Candidate Gene Screening and Confirmation
3.4. Scanning Electron Microscopy and Cytometric Analysis
3.5. Analysis of Plasma Membrane Fluidity
3.6. Measurement of Membrane Potential
3.7. Detection of Cell Membrane Integrity
3.8. Expression of Crude Proteins
3.9. Antimicrobial Activity and Thermal Stability Assays
3.10. Generation of His-Tag Fusion Peptides
3.11. Purification of Extracellular Peptides
3.12. Tris-Tricine SDS-PAGE and Western Blotting
3.13. Resistance Determination Test
3.14. Anti-Soil-Borne Bacteria Assay
3.15. Hemolytic Activity of IiR515 and IiR915 Peptides
4. Discussion
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Ma, L.; Wang, Y.; Wang, M.; Tian, Y.; Kang, W.; Liu, H.; Wang, H.; Dou, J.; Zhou, C. Effective antimicrobial activity of cbf-14, derived from a cathelin-like domain, against penicillin-resistant bacteria. Biomaterials 2016, 87, 32–45. [Google Scholar] [CrossRef]
- Gill, E.E.; Franco, O.L.; Hancock, R.E.W. Antibiotic adjuvants: Diverse strategies for controlling drug-resistant pathogens. Chem. Biol. Drug Des. 2015, 85, 56–78. [Google Scholar] [CrossRef] [PubMed]
- Dawson, R.M.; Liu, C.-Q. Properties and applications of antimicrobial peptides in biodefense against biological warfare threat agents. Crit. Rev. Microbiol. 2008, 34, 89–107. [Google Scholar] [CrossRef] [PubMed]
- Davies, J.; Davies, D. Origins and evolution of antibiotic resistance. Microbiol. Mol. Biol. Rev. MMBR 2010, 74, 417–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganewatta, M.S.; Tang, C. Controlling macromolecular structures towards effective antimicrobial. Polymer 2015, 63, A1–A29. [Google Scholar] [CrossRef]
- Akaddar, A.; Doderer-Lang, C.; Marzahn, M.R.; Delalande, F.; Mousli, M.; Helle, K.; Van Dorsselaer, A.; Aunis, D.; Dunn, B.M.; Metz-Boutigue, M.H.; et al. Catestatin, an endogenous chromogranin a-derived peptide, inhibits in vitro growth of Plasmodium falciparum. Cell. Mol. Life Sci. CMLS 2010, 67, 1005–1015. [Google Scholar] [CrossRef] [Green Version]
- Liang, Q.; Chalamaiah, M.; Ren, X.; Ma, H.; Wu, J. Identification of new anti-inflammatory peptides from zein hydrolysate after simulated gastrointestinal digestion and transport in caco-2 cells. J. Agric. Food Chem. 2018, 66, 1114–1120. [Google Scholar] [CrossRef]
- Seel, W.; Flegler, A.; Zunabovic-Pichler, M.; Lipski, A. Increased isoprenoid quinone concentration modulates membrane fluidity in Listeria monocytogenes at low growth temperatures. J. Bacteriol. 2018, 200, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Schroeder, B.O.; Wu, Z.; Nuding, S.; Groscurth, S.; Marcinowski, M.; Beisner, J.; Buchner, J.; Schaller, M.; Stange, E.F.; Wehkamp, J. Reduction of disulphide bonds unmasks potent antimicrobial activity of human beta-defensin 1. Nature 2011, 469, 419–423. [Google Scholar] [CrossRef]
- Ramos, R.; Moreira, S.; Rodrigues, A.; Gama, M.; Domingues, L. Recombinant expression and purification of the antimicrobial peptide magainin-2. Biotechnol. Prog. 2013, 29, 17–22. [Google Scholar] [CrossRef] [Green Version]
- Ling, L.L.; Schneider, T.; Peoples, A.J.; Spoering, A.L.; Engels, I.; Conlon, B.P.; Mueller, A.; Schaberle, T.F.; Hughes, D.E.; Epstein, S.; et al. A new antibiotic kills pathogens without detectable resistance. Nature 2015, 517, 455–459. [Google Scholar] [CrossRef]
- Maria-Neto, S.; de Almeida, K.C.; Macedo, M.L.; Franco, O.L. Understanding bacterial resistance to antimicrobial peptides: From the surface to deep inside. Biochim. Biophys. Acta 2015, 1848, 3078–3088. [Google Scholar] [CrossRef] [Green Version]
- Bahar, A.A.; Ren, D. Antimicrobial peptides. Pharmaceuticals 2013, 6, 1543–1575. [Google Scholar] [CrossRef] [Green Version]
- Oumer, O.J.; Abate, D. Characterization of pectinase from Bacillus subtilis strain btk 27 and its potential application in removal of mucilage from coffee beans. Enzym. Res. 2017, 2017, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Kakeshita, H.; Kageyama, Y.; Endo, K.; Tohata, M.; Ara, K.; Ozaki, K.; Nakamura, K. Secretion of biologically-active human interferon-beta by Bacillus subtilis. Biotechnol. Lett. 2011, 33, 1847–1852. [Google Scholar] [CrossRef]
- Schallmey, M.; Singh, A.; Ward, O.P. Developments in the use of Bacillus species for industrial production. Can. J. Microbiol. 2004, 50, 1–17. [Google Scholar] [CrossRef]
- Shah, K.; Bhatt, S. Purification and characterization of lipase from Bacillus subtilis pa2. J. Biochem. Tech. 2011, 3, 292–295. [Google Scholar]
- Oyeleke, S.B.; Oyewole, O.A.; Egwim, E.C. Production of protease and amylase from Bacillus subtilis and Aspergillus niger using Parkia biglobossa (africa locust beans) as substrate in solid state fermentation. Adv. Life Sci. 2012, 1, 49–53. [Google Scholar]
- Li, J.; Zhou, B.; Li, C.; Chen, Q.; Wang, Y.; Li, Z.; Chen, T.; Yang, C.; Jiang, Z.; Zhong, N.; et al. Lariciresinol-4-o-beta-d-glucopyranoside from the root of Isatis indigotica inhibits influenza a virus-induced pro-inflammatory response. J. Ethnopharmacol. 2015, 174, 379–386. [Google Scholar] [CrossRef]
- Nguyen, T.K.; Jamali, A.; Grand, E.; Morreel, K.; Marcelo, P.; Gontier, E.; Dauwe, R. Phenylpropanoid profiling reveals a class of hydroxycinnamoyl glucaric acid conjugates in Isatis tinctoria leaves. Phytochemistry 2017, 144, 127–140. [Google Scholar] [CrossRef]
- Lin, C.-W.; Tsai, F.-J.; Tsai, C.-H.; Lai, C.-C.; Wan, L.; Ho, T.-Y.; Hsieh, C.-C.; Chao, P.-D.L. Anti-sars coronavirus 3c-like protease effects of Isatis indigotica root and plant-derived phenolic compounds. Antivir. Res. 2005, 68, 36–42. [Google Scholar] [CrossRef]
- Ahmad, I.; Fatima, I. Butyrylcholinesterase, lipoxygenase inhibiting and antifungal alkaloids from Isatis tinctoria. J. Enzym. Inhib. Med. Chem. 2008, 23, 313–316. [Google Scholar] [CrossRef]
- Camo, C.; Torne, M.; Besalu, E.; Roses, C.; Cirac, A.D.; Moiset, G.; Badosa, E.; Bardaji, E.; Montesinos, E.; Planas, M.; et al. Tryptophan-containing cyclic decapeptides with activity against plant pathogenic bacteria. Molecules 2017, 22, 1817. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Wang, Y.; Zheng, L.; Yang, X.; Liu, H.; Guo, J. Isolation and characterization of an antifungal protein from Bacillus licheniformis hs10. Biochem. Biophys. Res. Commun. 2014, 454, 48–52. [Google Scholar] [CrossRef]
- Kong, X.; Yang, M.; Abbas, H.M.K.; Wu, J.; Li, M.; Dong, W. Antimicrobial genes from Allium sativum and Pinellia ternata revealed by a Bacillus subtilis expression system. Sci. Rep. 2018, 8, 14514. [Google Scholar] [CrossRef]
- Abbas, H.M.K.; Xiang, J.; Ahmad, Z.; Wang, L.; Dong, W. Enhanced Nicotiana benthamiana immune responses caused by heterologous plant genes from Pinellia ternata. BMC Plant Biol. 2018, 18, 357. [Google Scholar] [CrossRef]
- Niu, X.; J.Guiltinan, M. DNA binding specificity of the wheat bzip protein embp-1. Nucleic Acids Res. 1994, 22, 4969–4978. [Google Scholar] [CrossRef] [Green Version]
- Davis, E.G.; Sang, Y.; Rush, B.; Zhang, G.; Blecha, F. Molecular cloning and characterization of equine nk-lysin. Vet. Immunol. Immunopathol. 2005, 105, 163–169. [Google Scholar] [CrossRef]
- Zheng, X.; Wang, X.; Teng, D.; Mao, R.; Hao, Y.; Yang, N.; Zong, L.; Wang, J. Mode of action of plectasin-derived peptides against gas gangrene-associated Clostridium perfringens type A. PLoS ONE 2017, 12, e0185215. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.; Hwang, J.S.; Lee, J.; Kim, J.I.; Lee, D.G. Scolopendin 2, a cationic antimicrobial peptide from centipede, and its membrane-active mechanism. Biochim. Biophys. Acta 2015, 1848, 634–642. [Google Scholar] [CrossRef] [Green Version]
- Teng, D.; Wang, X.; Xi, D.; Mao, R.; Zhang, Y.; Guan, Q.; Zhang, J.; Wang, J. A dual mechanism involved in membrane and nucleic acid disruption of AvBD103b, a new avian defensin from the king penguin, against Salmonella enteritidis CVCC3377. Appl. Microbiol. Biotechnol. 2014, 98, 8313–8325. [Google Scholar] [CrossRef]
- Jacquet, T.; Cailliez-Grimal, C.; Francius, G.; Borges, F.; Imran, M.; Duval, J.F.; Revol-Junelles, A.M. Antibacterial activity of class IIa bacteriocin Cbn BM1 depends on the physiological state of the target bacteria. Res. Microbiol. 2012, 163, 323–331. [Google Scholar] [CrossRef]
- Castellano, P.; Raya, R.; Vignolo, G. Mode of action of lactocin 705, a two-component bacteriocin from Lactobacillus casei CRL705. Int. J. Food Microbiol. 2003, 85, 35–43. [Google Scholar] [CrossRef]
- Patra, P.; Mitra, S.; Goswami, A.; Roy, S.; Pradhan, S.; Sarkar, S.; Debnath, N. Damage of lipopolysaccharides in outer cell membrane and production of ros-mediated stress within bacteria makes nano zinc oxide a bactericidal agent. Appl. Nanosci. 2015, 5, 857–866. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Yang, X.; Guo, L.; Zeng, H.; Qiu, D. Pebl1, a novel protein elicitor from Brevibacillus laterosporus strain a60, activates defense responses and systemic resistance in Nicotiana benthamiana. Appl. Environ. Microbiol. 2015, 81, 2706–2716. [Google Scholar] [CrossRef] [Green Version]
- Yin, M.; Liu, D.; Xu, F.; Xiao, L.; Wang, Q.; Wang, B.; Chang, Y.; Zheng, J.; Tao, X.; Liu, G.; et al. A specific antimicrobial protein CAP-1 from Pseudomonas sp. isolated from the jellyfish Cyanea capillata. Int. J. Biol. Macromol. 2016, 82, 488–496. [Google Scholar] [CrossRef]
- Zhou, F.; Zhang, X.-L.; Li, J.-L.; Zhu, F.-X. Dimethachlon resistance in Sclerotinia sclerotiorum in china. Plant Dis. 2014, 98, 1221–1226. [Google Scholar] [CrossRef] [Green Version]
- Anagnostopoulos, C.; Spizizen, J. Requirements for transformation in Bacillus subtilis. J. Bacteriol. 1961, 81, 741–746. [Google Scholar]
- Wen, L.S.; Philip, K.; Ajam, N. Purification, characterization and mode of action of plantaricin K25 produced by Lactobacillus plantarum. Food Control 2016, 60, 430–439. [Google Scholar] [CrossRef]
- Wang, S.; Wang, Q.; Zeng, X.; Ye, Q.; Huang, S.; Yu, H.; Yang, T.; Qiao, S. Use of the antimicrobial peptide sublancin with combined antibacterial and immunomodulatory activities to protect against methicillin-resistant Staphylococcus aureus infection in mice. J. Agric. Food Chem. 2017, 65, 8595–8605. [Google Scholar] [CrossRef]
- Xu, L.; Xiang, M.; White, D.; Chen, W. Ph dependency of sclerotial development and pathogenicity revealed by using genetically defined oxalate-minus mutants of Sclerotinia sclerotiorum. Environ. Microbiol. 2015, 17, 2896–2909. [Google Scholar] [CrossRef]
- Mohanram, H.; Bhattacharjya, S. ’Lollipop’-shaped helical structure of a hybrid antimicrobial peptide of temporin b-lipopolysaccharide binding motif and mapping cationic residues in antibacterial activity. Biochim. Biophys. Acta 2016, 1860, 1362–1372. [Google Scholar] [CrossRef]
- Tang, X.; Shi, J.; Dong, W. A yeast library-hybrid assay to screen maize-Rhizoctonia transcription factors and protein-protein interactions in one experimental pipeline. Agri Gene 2016, 1, 15–22. [Google Scholar] [CrossRef]
- Kumaresan, V.; Bhatt, P.; Ganesh, M.R.; Harikrishnan, R.; Arasu, M.; Al-Dhabi, N.A.; Pasupuleti, M.; Marimuthu, K.; Arockiaraj, J. A novel antimicrobial peptide derived from fish goose type lysozyme disrupts the membrane of Salmonella enterica. Mol. Immunol. 2015, 68, 421–433. [Google Scholar] [CrossRef]
- Westers, L.; Westers, H.; Quax, W.J. Bacillus subtilis as cell factory for pharmaceutical proteins: A biotechnological approach to optimize the host organism. Biochim. Biophys. Acta 2004, 1694, 299–310. [Google Scholar] [CrossRef]
- Seifert, K.; Unger, W. Insecticidal and fungicidal compounds from Isatis Tinctoria. Z. Nat. 1994, 49c, 44–48. [Google Scholar] [CrossRef] [Green Version]
- Xiao, P.; Huang, H.; Chen, J.; Li, X. In vitro antioxidant and anti-inflammatory activities of Radix isatidis extract and bioaccessibility of six bioactive compounds after simulated gastro-intestinal digestion. J. Ethnopharmacol. 2014, 157, 55–61. [Google Scholar] [CrossRef]
- Hao, G.; Shi, Y.-H.; Tang, Y.-L.; Le, G.-W. The intracellular mechanism of action on Escherichia coli of bf2-a/c, two analogues of the antimicrobial peptide buforin 2. J. Microbiol. 2013, 51, 200–206. [Google Scholar] [CrossRef]
- Haversen, L.; Kondori, N.; Baltzer, L.; Hanson, L.A.; Dolphin, G.T.; Duner, K.; Mattsby-Baltzer, I. Structure-microbicidal activity relationship of synthetic fragments derived from the antibacterial alpha-helix of human lactoferrin. Antimicrob. Agents Chemother. 2010, 54, 418–425. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.; Kim, H.J.; Hahm, K.S. Antibacterial synergism of novel antibiotic peptides with chloramphenicol. Biochem. Biophys. Res. Commun. 2004, 321, 109–115. [Google Scholar] [CrossRef]
- Ling Lin, F.; Zi Rong, X.; Wei Fen, L.; Jiang Bing, S.; Ping, L.; Chun Xia, H. Protein secretion pathways in Bacillus subtilis: Implication for optimization of heterologous protein secretion. Biotechnol. Adv. 2007, 25, 1–12. [Google Scholar] [CrossRef]
- Barbe, V.; Cruveiller, S.; Kunst, F.; Lenoble, P.; Meurice, G.; Sekowska, A.; Vallenet, D.; Wang, T.; Moszer, I.; Medigue, C.; et al. From a consortium sequence to a unified sequence: The Bacillus subtilis 168 reference genome a decade later. Microbiology 2009, 155, 1758–1775. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.C.; Yeung, J.C.; Duan, Y.; Ye, R.; Szarka, S.J.; Habibi, H.R.; Wong, S.L. Functional production and characterization of a fibrin-specific single-chain antibody fragment from Bacillus subtilis: Effects of molecular chaperones and a wall-bound protease on antibody fragment production. Appl. Environ. Microbiol. 2002, 68, 3261–3269. [Google Scholar] [CrossRef] [Green Version]
- Li, C.L.; Xu, T.T.; Chen, R.B.; Huang, X.X.; Zhao, Y.C.; Bao, Y.Y.; Zhao, W.D.; Zheng, Z.Y. Cloning, expression and characterization of antimicrobial porcine beta defensin 1 in Escherichia coli. Protein Exp. Purif. 2013, 88, 47–53. [Google Scholar] [CrossRef]
- Musta, A.C.; Riederer, K.; Shemes, S.; Chase, P.; Jose, J.; Johnson, L.B.; Khatib, R. Vancomycin MIC plus heteroresistance and outcome of methicillin-resistant Staphylococcus aureus bacteremia: Trends over 11 Years. J. Clin. Microbiol. 2009, 47, 1640–1644. [Google Scholar] [CrossRef] [Green Version]
- Lamour, K.H.; Stam, R.; Jupe, J.; Huitema, E. The oomycete broad-host-range pathogen Phytophthora capsici. Mol. Plant Pathol. 2012, 13, 329–337. [Google Scholar] [CrossRef]
- Lai, G.; Fu, P.; Liu, Y.; Xiang, J.; Lu, J. Molecular characterization and overexpression of VpRPW8s from Vitis pseudoreticulata enhances resistance to Phytophthora capsici in Nicotiana benthamiana. Int. J. Mol. Sci. 2018, 19, 839. [Google Scholar] [CrossRef] [Green Version]
- Zeitler, B.; Meyer, H.; Bernhard, A.; Sattler, M.; Lindermayr, C. Production of a de-novo designed antimicrobial peptide in Nicotiana benthamiana. Plant Mol. Biol. 2013, 81, 259–272. [Google Scholar] [CrossRef]
- Rammelkamp, C.H.; Weinstein, L. Toxic effects of tyrothricin, gramicidin and tyrocidine. J. Infect. Dis. 1942, 71, 166–173. [Google Scholar] [CrossRef]
- Ali, M.; Sun, Y.; Xie, L.; Yu, H.; Bashir, A.; Li, L. The pathogenicity of Pseudomonas syringae mb03 against Caenorhabditis elegans and the transcriptional response of nematicidal genes upon different nutritional conditions. Front. Microbiol. 2016, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, J.; Abbas, H.M.K.; Li, J.; Yuan, Y.; Liu, Y.; Wang, G.; Dong, W. Cell Membrane-Interrupting Antimicrobial Peptides from Isatis indigotica Fortune Isolated by a Bacillus subtilis Expression System. Biomolecules 2020, 10, 30. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10010030
Wu J, Abbas HMK, Li J, Yuan Y, Liu Y, Wang G, Dong W. Cell Membrane-Interrupting Antimicrobial Peptides from Isatis indigotica Fortune Isolated by a Bacillus subtilis Expression System. Biomolecules. 2020; 10(1):30. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10010030
Chicago/Turabian StyleWu, Jia, Hafiz Muhammad Khalid Abbas, Jiale Li, Yuan Yuan, Yunjun Liu, Guoying Wang, and Wubei Dong. 2020. "Cell Membrane-Interrupting Antimicrobial Peptides from Isatis indigotica Fortune Isolated by a Bacillus subtilis Expression System" Biomolecules 10, no. 1: 30. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10010030