Alpha-Amylase and Alpha-Glucosidase Enzyme Inhibition and Antioxidant Potential of 3-Oxolupenal and Katononic Acid Isolated from Nuxia oppositifolia

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Plant Material
Extraction and Isolation
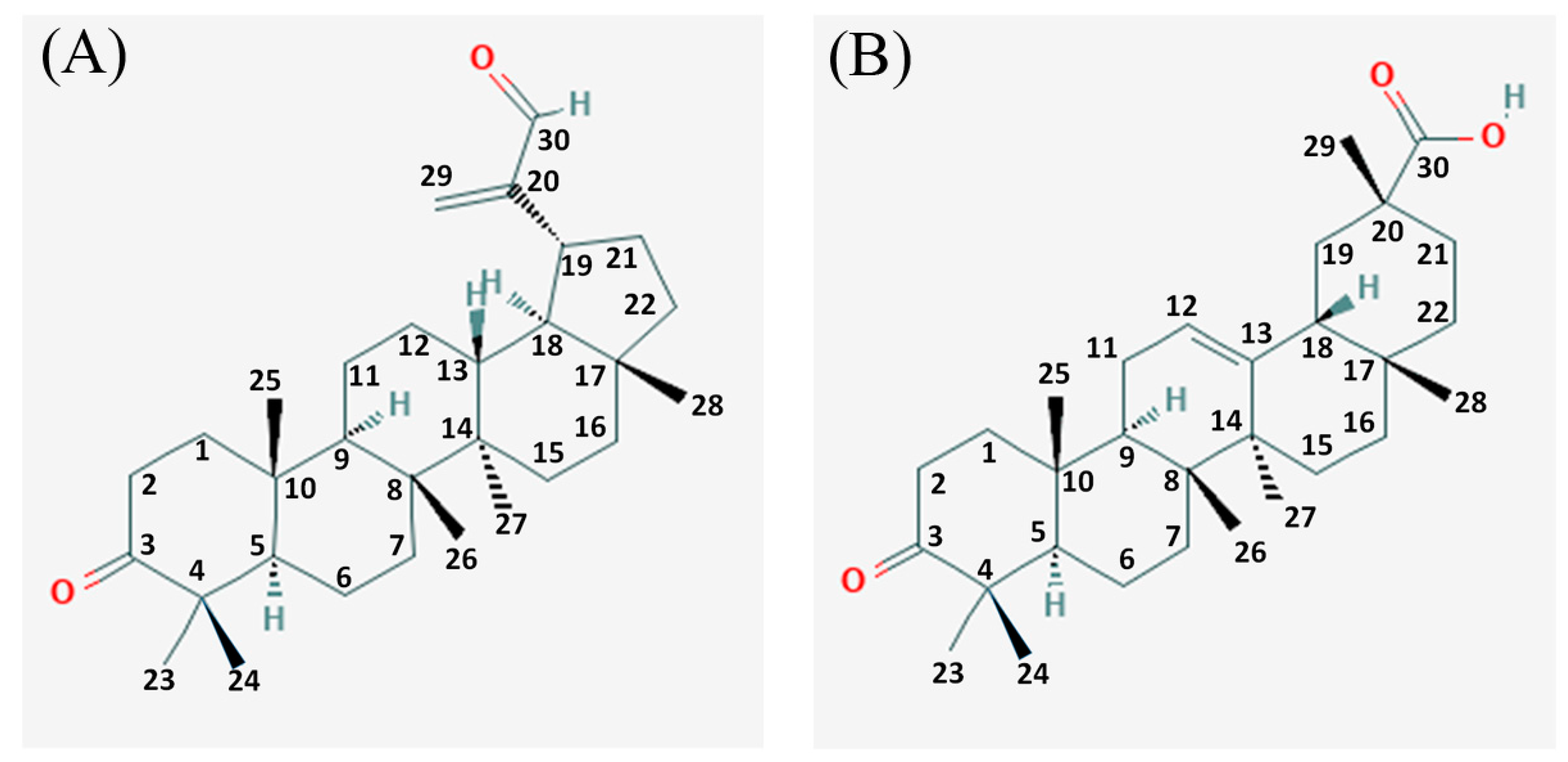
2.3. Isolation and Identification of 3-Oxolupenal and Katononic Acid
2.4. Inhibition Assay for Amylase Activity
2.5. Inhibition Assay for Glucosidase Activity
2.6. DPPH Radical Scavenging Activity
2.7. ABTS Radical Cation Scavenging Activity
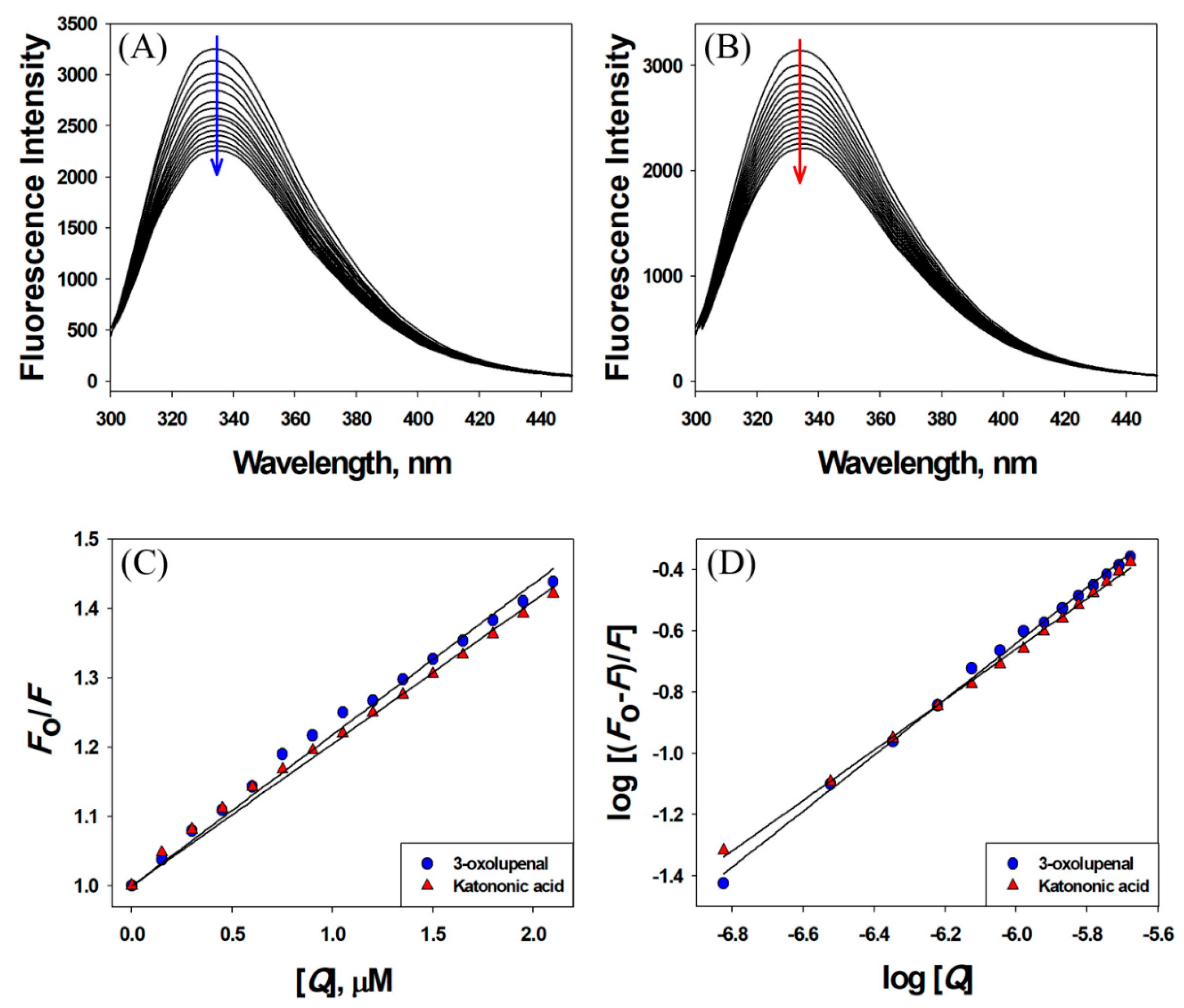
2.8. Fluorescence Quenching-Based Binding Studies
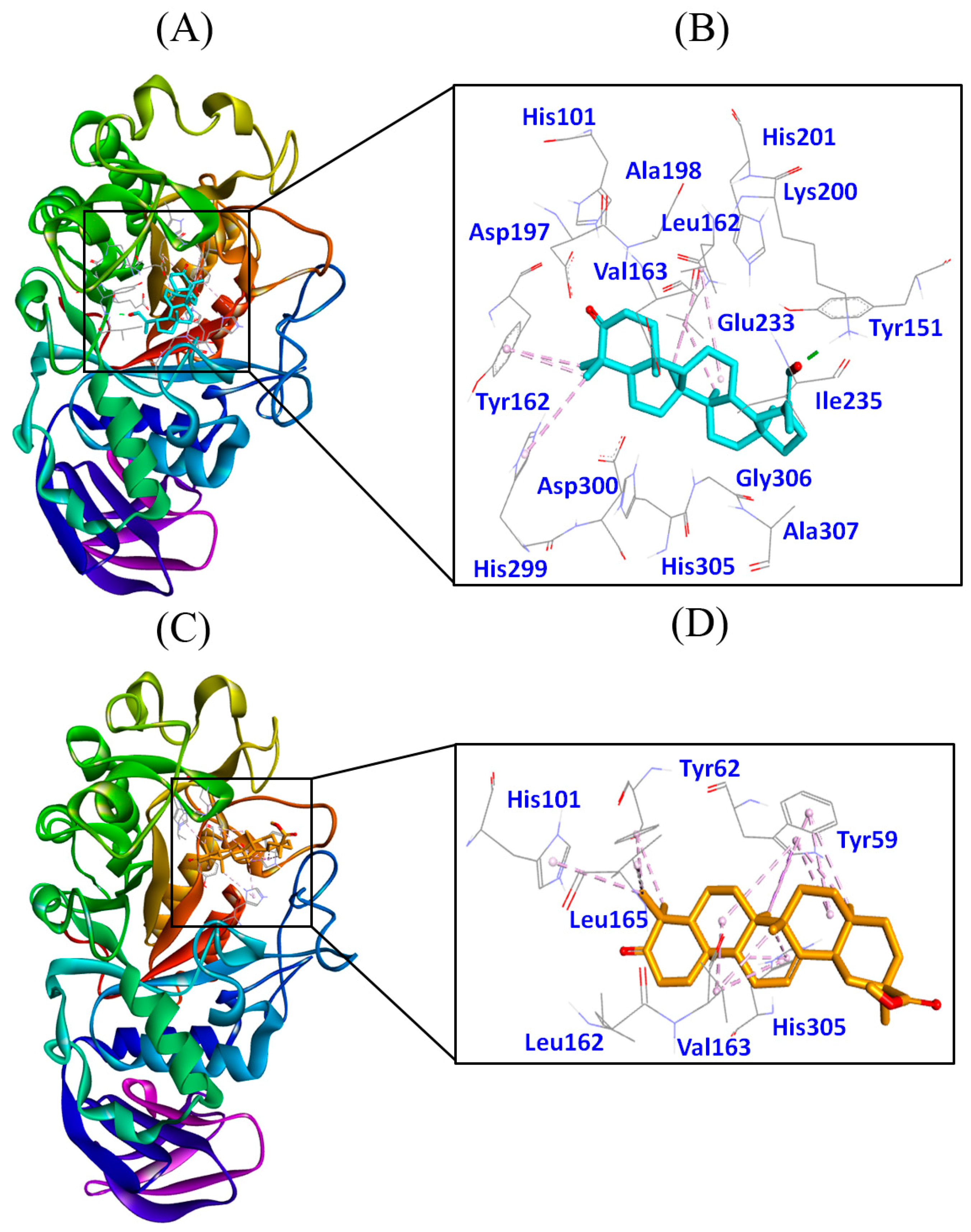
2.9. Molecular Docking Studies
2.10. Statistical Analysis
3. Results and Discussion
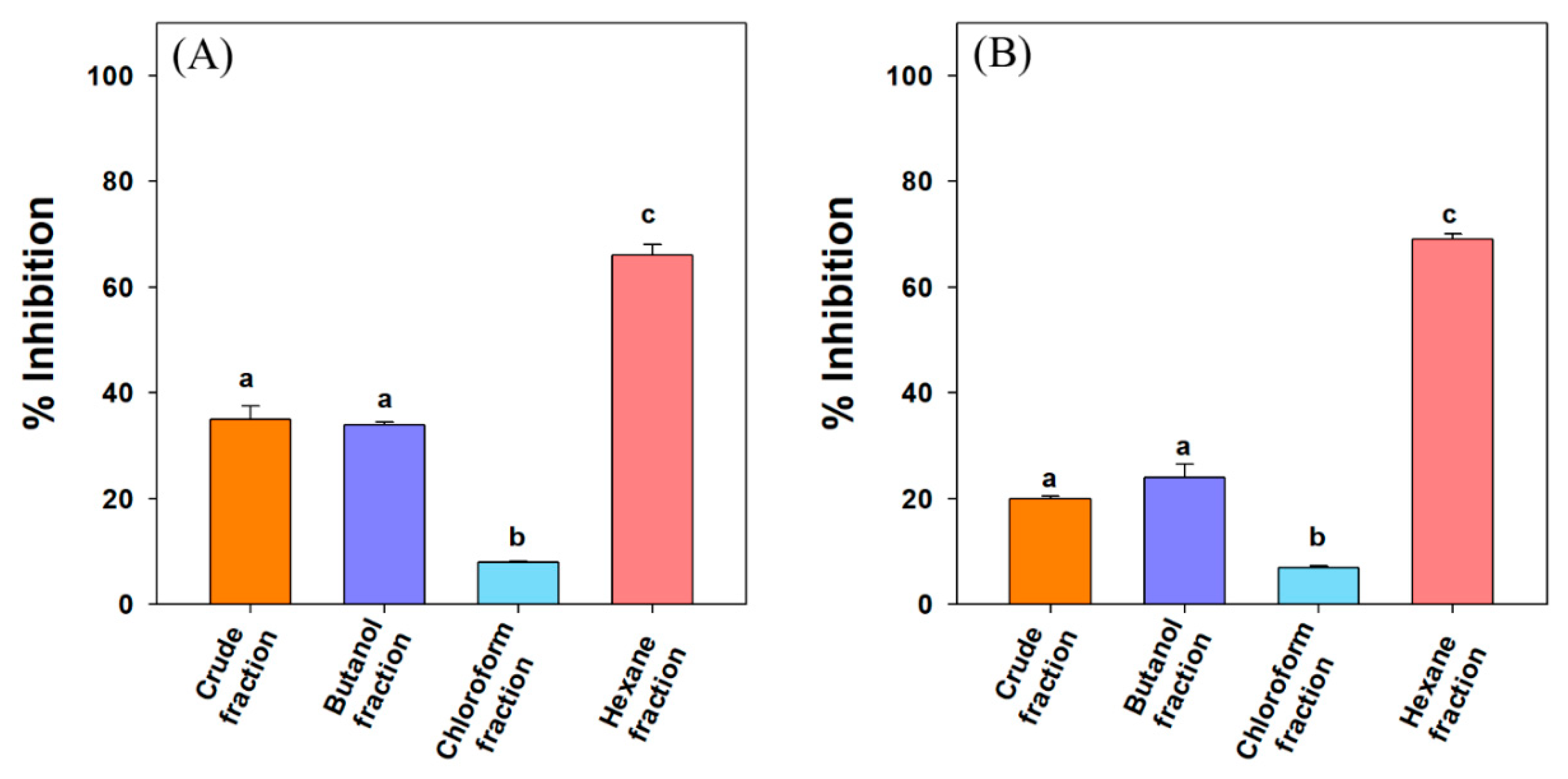
3.1. α-Amylase and α-Glucosidase Inhibition Studies
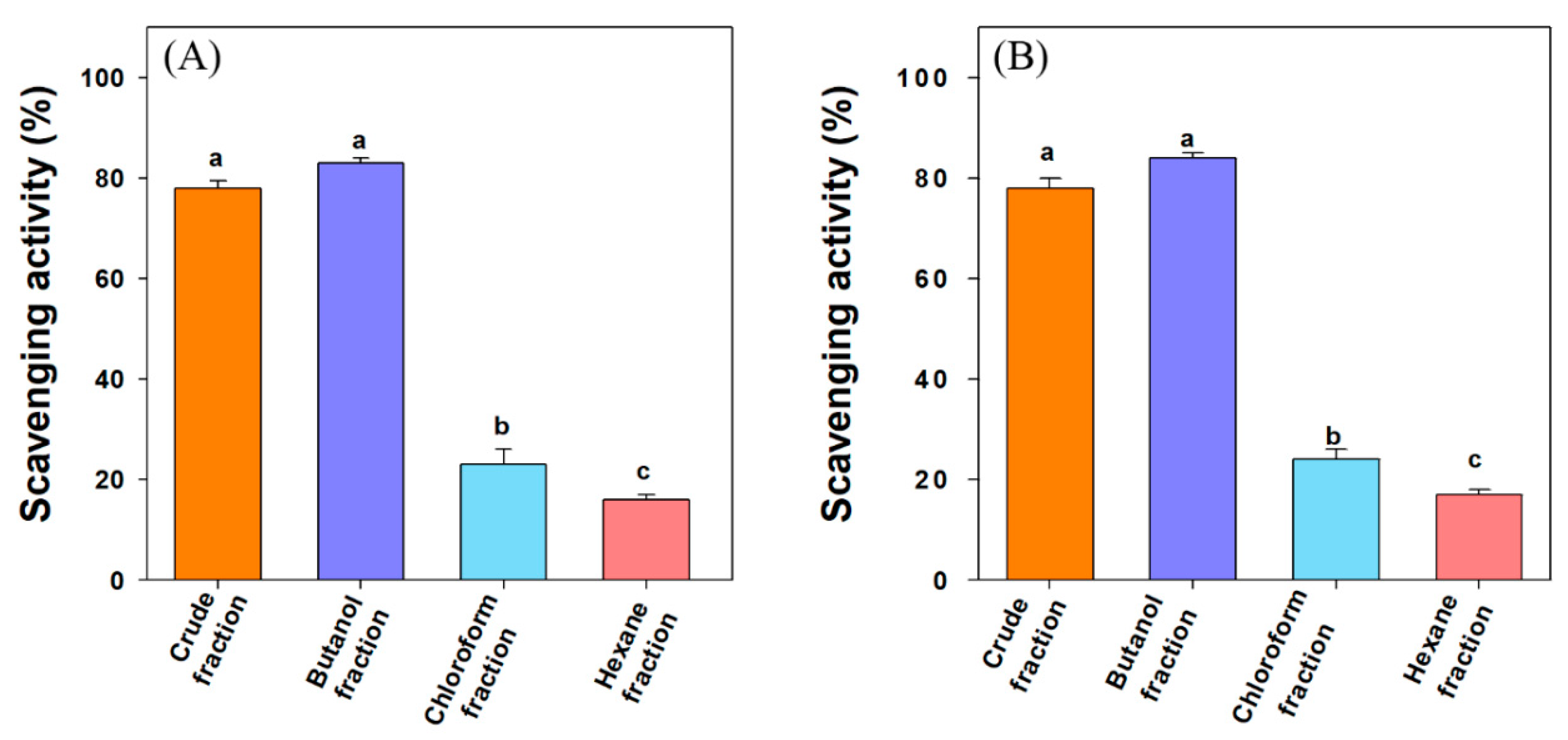
3.2. DPPH and ABTS Scavenging Activity
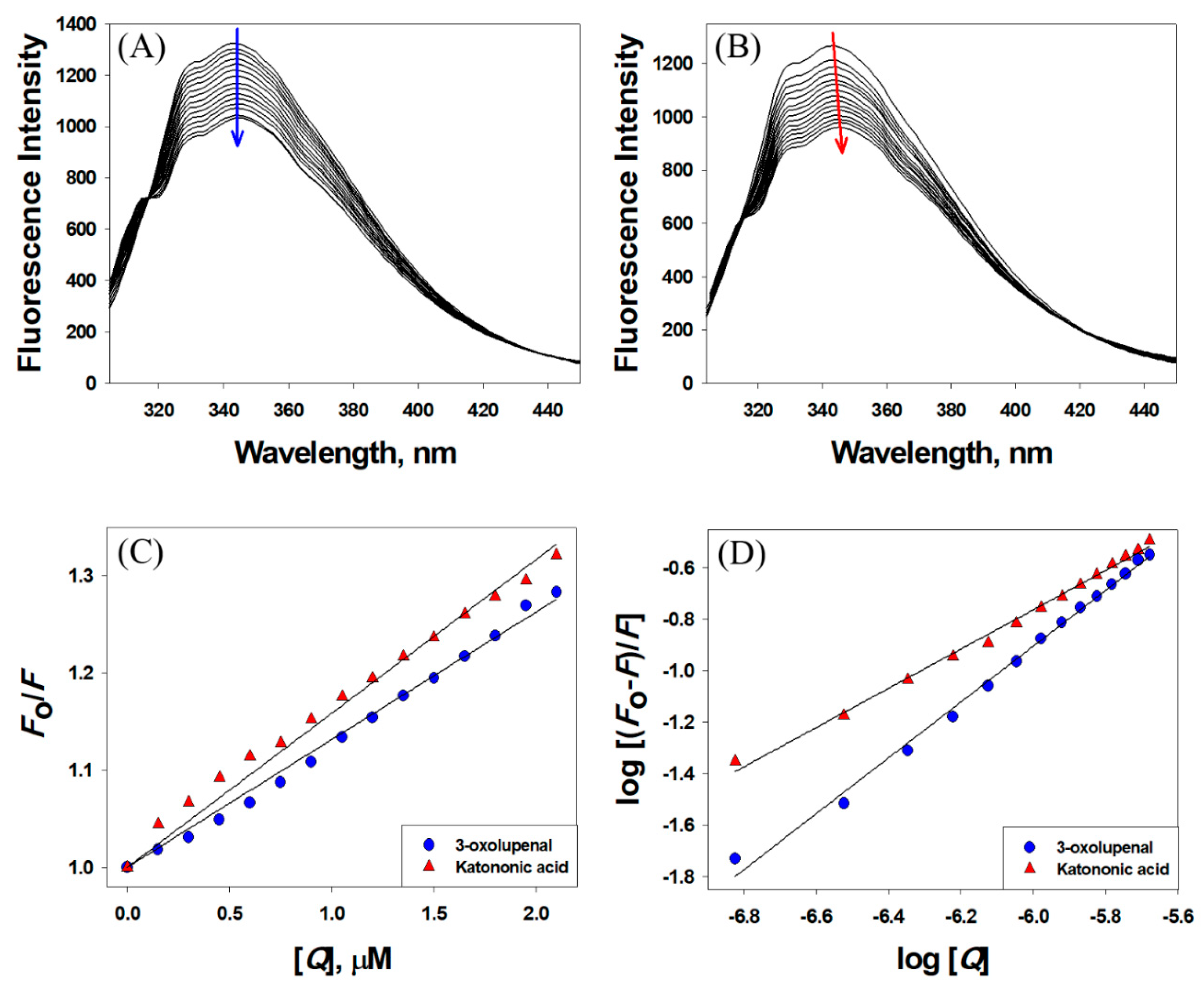
3.3. Interaction between 3-Oxolupenal and Katononic Acid with α-Amylase and α-Glucosidase
3.4. Molecular Docking Studies
3.4.1. Authentication of Molecular Docking Method
3.4.2. Molecular Docking of 3-Oxolupenal and Katononic Acid with α-Amylase
3.4.3. Molecular Docking of 3-Oxolupenal and Katononic Acid with α-Glucosidase
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pecoits-Filho, R.; Abensur, H.; Betônico, C.C.R.; Machado, A.D.; Parente, E.B.; Queiroz, M.; Salles, J.E.N.; Titan, S.; Vencio, S. Interactions between kidney disease and diabetes: Dangerous liaisons. Diabetol. Metab. Syndr. 2016, 8, 50. [Google Scholar] [CrossRef] [PubMed]
- Canivell, S.; Gomis, R. Diagnosis and classification of autoimmune diabetes mellitus. Autoimmun. Rev. 2014, 13, 403–407. [Google Scholar] [CrossRef] [PubMed]
- Adeloye, D.; Ige, J.O.; Aderemi, A.V.; Adeleye, N.; Amoo, E.O.; Auta, A.; Oni, G. Estimating the prevalence, hospitalisation and mortality from type 2 diabetes mellitus in Nigeria: A systematic review and meta-analysis. BMJ Open 2017, 7, e015424. [Google Scholar] [CrossRef] [PubMed]
- Kajaria, D.; Tiwari, S.; Tripathi, J.; Tripathi, Y. Ranjana In-vitro α amylase and glycosidase inhibitory effect of ethanolic extract of antiasthmatic drug-Shirishadi. J. Adv. Pharm. Technol. Res. 2013, 4, 206–209. [Google Scholar] [CrossRef]
- Derosa, G.; Maffioli, P. Mini-special issue paper management of diabetic patients with hypoglycemic agents α-Glucosidase inhibitors and their use in clinical practice. Arch. Med. Sci. 2012, 5, 899–906. [Google Scholar] [CrossRef]
- Matough, F.A.; Budin, S.B.; Hamid, Z.A.; Alwahaibi, N.; Mohamed, J. The role of oxidative stress and antioxidants in diabetic complications. Sultan Qaboos Univ. Med. J. 2012, 12, 5–18. [Google Scholar] [CrossRef]
- Al-Abbasi, T.M.; Al-Farhan, A.; Al-Khulaidi, A.W.; Hall, M.; Llewellyn, O.A.; Miller, A.G.; Patzelt, A. Important plant areas in the arabian peninsula. Edinb. J. Bot. 2010, 67, 25–35. [Google Scholar] [CrossRef]
- Mambu, L.; Grellier, P.; Florent, L.; Joyeau, R.; Ramanitrahasimbola, D.; Rasoanaivo, P.; Frappier, F. Clerodane and labdane diterpenoids from Nuxia sphaerocephala. Phytochemistry 2006, 67, 444–451. [Google Scholar] [CrossRef]
- Boaduo, N.K.K.; Katerere, D.; Eloff, J.N.; Naidoo, V. Evaluation of six plant species used traditionally in the treatment and control of diabetes mellitus in South Africa using in vitro methods. Pharm. Biol. 2014, 52, 756–761. [Google Scholar] [CrossRef] [Green Version]
- Jensen, S.R. Chemotaxonomy of the genus Nuxia (Buddlejaceae). Stud. Plant Sci. 1999, 6, 379–382. [Google Scholar]
- Al-Massarani, S.; El-Gamal, A.; Parvez, M.; Al-Dosari, M.; Al-Said, M.; Abdel-Kader, M.; Basudan, O. New cytotoxic seco-type triterpene and labdane-type diterpenes from Nuxia oppositifolia. Molecules 2017, 22, 389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nazaruk, J.; Borzym-Kluczyk, M. The role of triterpenes in the management of diabetes mellitus and its complications. Phytochem. Rev. 2015, 14, 675–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Lou, G.; Meng, Z.; Huang, W. TGR5: A novel target for weight maintenance and glucose metabolism. Exp. Diabetes Res. 2011, 2011. [Google Scholar] [CrossRef] [PubMed]
- Sato, H.; Genet, C.; Strehle, A.; Thomas, C.; Lobstein, A.; Wagner, A.; Mioskowski, C.; Auwerx, J.; Saladin, R. Anti-hyperglycemic activity of a TGR5 agonist isolated from Olea europaea. Biochem. Biophys. Res. Commun. 2007, 362, 793–798. [Google Scholar] [CrossRef] [PubMed]
- Genet, C.; Strehle, A.; Schmidt, C.; Boudjelal, G.; Lobstein, A.; Schoonjans, K.; Souchet, M.; Auwerx, J.; Saladin, R.; Wagner, A. Structure-Activity relationship study of betulinic acid, a novel and selective TGR5 agonist, and its synthetic derivatives: Potential impact in diabetes. J. Med. Chem. 2010, 53, 178–190. [Google Scholar] [CrossRef]
- Ramachandran, S.; Prasad, N.R. Effect of ursolic acid, a triterpenoid antioxidant, on ultraviolet-B radiation-induced cytotoxicity, lipid peroxidation and DNA damage in human lymphocytes. Chem. Biol. Interact. 2008, 176, 99–107. [Google Scholar] [CrossRef]
- Yamaguchi, Y.; Yamada, K.; Yoshikawa, N.; Nakamura, K.; Haginaka, J.; Kunitomo, M. Corosolic acid prevents oxidative stress, inflammation and hypertension in SHR/NDmcr-cp rats, a model of metabolic syndrome. Life Sci. 2006, 79, 2474–2479. [Google Scholar] [CrossRef]
- Manna, P.; Sinha, M.; Sil, P.C. Prophylactic role of arjunolic acid in response to streptozotocin mediated diabetic renal injury: Activation of polyol pathway and oxidative stress responsive signaling cascades. Chem. Biol. Interact. 2009, 181, 297–308. [Google Scholar] [CrossRef]
- Ghosh, T.; Maity, T.K.; Singh, J. Antihyperglycemic activity of bacosine, a triterpene from bacopa monnieri, in alloxan-induced diabetic rats. Planta Med. 2011, 77, 804–808. [Google Scholar] [CrossRef]
- Xiang, M.; Wang, J.; Zhang, Y.; Ling, J.; Xu, X. Attenuation of aortic injury by ursolic acid through RAGE-Nox-NFκB pathway in streptozocin-induced diabetic rats. Arch. Pharm. Res. 2012, 35, 877–886. [Google Scholar] [CrossRef]
- Kwon, Y.-I.; Apostolidis, E.; Shetty, K. Inhibitory potential of wine and tea against α-amylase and α-glucosidase for management of hyperglycemia linked to type 2 diabetes. J. Food Biochem. 2008, 32, 15–31. [Google Scholar] [CrossRef]
- Elya, B.; Basah, K.; Mun’im, A.; Yuliastuti, W.; Bangun, A.; Septiana, E.K. Screening of α-Glucosidase inhibitory activity from some plants of Apocynaceae, Clusiaceae, Euphorbiaceae, and Rubiaceae. J. Biomed. Biotechnol. 2012, 2012, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braca, A.; De Tommasi, N.; Di Bari, L.; Pizza, C.; Politi, M.; Morelli, I. Antioxidant principles from Bauhinia t arapotensis. J. Nat. Prod. 2001, 64, 892–895. [Google Scholar] [CrossRef] [PubMed]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Rehman, M.T.; Shamsi, H.; Khan, A.U. Insight into the binding mechanism of imipenem to human serum albumin by spectroscopic and computational approaches. Mol. Pharm. 2014, 11, 1785–1797. [Google Scholar] [CrossRef]
- Khan, M.S.; Rehman, M.T.; Bhat, S.A.; Tabrez, S.; Hussain, A.; Husain, F.M.; AlAjmi, M.F.; Alamery, S.F.; Sumbul, S. Food additive dye (quinoline yellow) promotes unfolding and aggregation of myoglobin: A spectroscopic and molecular docking analysis. Spectrochim. Acta Part. A Mol. Biomol. Spectrosc. 2019, 214, 216–226. [Google Scholar] [CrossRef]
- Rehman, M.T.; Dey, P.; Hassan, M.I.; Ahmad, F.; Batra, J.K. Functional role of Glutamine 28 and Arginine 39 in double stranded RNA cleavage by human pancreatic ribonuclease. PLoS ONE 2011, 6, e17159. [Google Scholar] [CrossRef]
- Williams, L.K.; Li, C.; Withers, S.G.; Brayer, G.D. Order and Disorder: Differential structural impacts of myricetin and ethyl caffeate on human amylase, an antidiabetic target. J. Med. Chem. 2012, 55, 10177–10186. [Google Scholar] [CrossRef]
- Roig-Zamboni, V.; Cobucci-Ponzano, B.; Iacono, R.; Ferrara, M.C.; Germany, S.; Bourne, Y.; Parenti, G.; Moracci, M.; Sulzenbacher, G. Structure of human lysosomal acid α-glucosidase–a guide for the treatment of Pompe disease. Nat. Commun. 2017, 8, 1111. [Google Scholar] [CrossRef] [Green Version]
- Sudha, P.; Zinjarde, S.S.; Bhargava, S.Y.; Kumar, A.R. Potent α-amylase inhibitory activity of Indian Ayurvedic medicinal plants. BMC Complement. Altern. Med. 2011, 11, 5. [Google Scholar] [CrossRef] [Green Version]
- De, D.; Bera, T.K.; Ali, K.M.; Mandal, S.; Barik, B.; Ghosh, D. Inhibitory effects of n-hexane fraction from hydro-methanolic (2:3) extract on α-amylase and α-glucosidase activities of sepal of Salmalia malabarica: An approach through in vitro study. Biomark. Genom. Med. 2013, 5, 48–53. [Google Scholar] [CrossRef]
- Pak, W.-M.; Kim, K.-B.-W.R.; Kim, M.-J.; Cho, J.-Y.; Ahn, D.-H. Inhibitory effect of hexane fraction from Myagropsis myagroides on pancreatic α-amylase in vitro. J. Microbiol. Biotechnol. 2015, 25, 328–333. [Google Scholar] [CrossRef] [PubMed]
- Salehi, P.; Asghari, B.; Esmaeili, M.A.; Dehghan, H.; Ghazi, I. α-glucosidase and α-amylase inhibitory effect and antioxidant activity of ten plant extracts traditionally used in Iran for diabetes. J. Med. Plants Res. 2013, 7, 257–266. [Google Scholar]
- Alqahtani, A.; Hamid, K.; Kam, A.; Wong, K.H.; Abdelhak, Z.; Razmovski-Naumovski, V.; Chan, K.; Li, K.M.; Groundwater, P.W.; Li, G.Q. The pentacyclic triterpenoids in herbal medicines and their pharmacological activities in diabetes and diabetic complications. Curr. Med. Chem. 2013, 20, 908–931. [Google Scholar]
- Wang, X.-L.; Jiao, F.-R.; Yu, M.; Lin, L.-B.; Xiao, J.; Zhang, Q.; Wang, L.; Duan, D.-Z.; Xie, G. Constituents with potent α-glucosidase inhibitory activity from Pueraria lobata (Willd.) ohwi. Bioorg. Med. Chem. Lett. 2017, 27, 1993–1998. [Google Scholar] [CrossRef]
- He, K.; Song, S.; Zou, Z.; Feng, M.; Wang, D.; Wang, Y.; Li, X.; Ye, X. The hypoglycemic and synergistic effect of loganin, morroniside, and ursolic acid isolated from the fruits of Cornus officinalis. Phyther. Res. 2016, 30, 283–291. [Google Scholar] [CrossRef]
- Hou, W.; Li, Y.; Zhang, Q.; Wei, X.; Peng, A.; Chen, L.; Wei, Y. Triterpene acids isolated from Lagerstroemia speciosa leaves as α -glucosidase inhibitors. Phyther. Res. 2009, 23, 614–618. [Google Scholar] [CrossRef]
- Zhang, B.; Xing, Y.; Wen, C.; Yu, X.; Sun, W.; Xiu, Z.; Dong, Y. Pentacyclic triterpenes as α-glucosidase and α-amylase inhibitors: Structure-activity relationships and the synergism with acarbose. Bioorg. Med. Chem. Lett. 2017, 27, 5065–5070. [Google Scholar] [CrossRef]
- Kumar, V.; Prakash, O.; Kumar, S.; Narwal, S. α-glucosidase inhibitors from plants: A natural approach to treat diabetes. Pharmacogn. Rev. 2011, 5, 19–29. [Google Scholar] [CrossRef] [Green Version]
- Benalla, W.; Bellahcen, S.; Bnouham, M. Antidiabetic medicinal plants as a source of alpha glucosidase inhibitors. Curr. Diabetes Rev. 2010, 6, 247–254. [Google Scholar] [CrossRef]
- Karakaya, S.; Gözcü, S.; Güvenalp, Z.; Özbek, H.; Yuca, H.; Dursunoğlu, B.; Kazaz, C.; Kılıç, C.S. The α-amylase and α-glucosidase inhibitory activities of the dichloromethane extracts and constituents of Ferulago bracteata roots. Pharm. Biol. 2018, 56, 18–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adjimani, J.P.; Asare, P. Antioxidant and free radical scavenging activity of iron chelators. Toxicol. Rep. 2015, 2, 721–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amorati, R.; Valgimigli, L. Advantages and limitations of common testing methods for antioxidants. Free Radic. Res. 2015, 49, 633–649. [Google Scholar] [CrossRef] [PubMed]
- Adewusi, E.A.; Moodley, N.; Steenkamp, V. Antioxidant and acetylcholinesterase inhibitory activity of selected southern African medicinal plants. S. Afr. J. Bot. 2011, 77, 638–644. [Google Scholar] [CrossRef] [Green Version]
- Amoo, S.O.; Aremu, A.O.; Moyo, M.; Van Staden, J. Antioxidant and acetylcholinesterase-inhibitory properties of long-term stored medicinal plants. BMC Complement. Altern. Med. 2012, 12, 1060. [Google Scholar] [CrossRef] [Green Version]
- Gangwar, M.; Gautam, M.K.; Sharma, A.K.; Tripathi, Y.B.; Goel, R.K.; Nath, G. Antioxidant capacity and radical scavenging effect of polyphenol rich mallotus philippenensis fruit extract on human erythrocytes: An in vitro study. Sci. World J. 2014, 2014, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Rehman, M.T.; Ahmed, S.; Khan, A.U. Interaction of meropenem with ‘N’ and ‘B’ isoforms of human serum albumin: A spectroscopic and molecular docking study. J. Biomol. Struct. Dyn. 2016, 34, 1849–1864. [Google Scholar] [CrossRef]
- Ware, W.R. Oxygen quenching of fluorescence in solution: An experimental study of the diffusion process. J. Phys. Chem. 1962, 66, 455–458. [Google Scholar] [CrossRef]
- AlAjmi, M.; Hussain, A.; Rehman, M.; Khan, A.; Shaikh, P.; Khan, R. Design, synthesis, and biological evaluation of benzimidazole-derived biocompatible Copper(II) and Zinc(II) complexes as anticancer chemotherapeutics. Int. J. Mol. Sci. 2018, 19, 1492. [Google Scholar] [CrossRef] [Green Version]
- Rydberg, E.H.; Li, C.; Maurus, R.; Overall, C.M.; Brayer, G.D.; Withers, S.G. Mechanistic analyses of catalysis in human pancreatic α-amylase: Detailed kinetic and structural studies of mutants of three conserved carboxylic acids. Biochemistry 2002, 41, 4492–4502. [Google Scholar] [CrossRef]
- Hermans, M.M.P.; Kroos, M.A.; Van Beeumen, J.; Oostra, B.A.; Reuser, A.J.J. Human lysosomal α-glucosidase: Characterization of the catalytic site. J. Biol. Chem. 1991, 266, 13507–13512. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fractions/Compounds | Inhibition of α-Amylase (µg/mL) | Inhibition of α-Glucosidase (µg/mL) |
|---|---|---|
| Crude extract | 90.3 ± 3.2 a | 98.4 ± 5.2 e |
| n-butanol fraction | ND | ND |
| Chloroform fraction | ND | ND |
| n-hexane fraction | 73.4 ± 7.3 b | 80.3 ± 8.9 b |
| 3-oxolupenal | 46.2 ± 5.2 c (105.3 µM) | 62.3 ± 2.4 c (141.8 µM) |
| Katononic acid | 52.4 ± 3.7 c (115.2 µM) | 88.6 ± 6.2 a (194.8 µM) |
| Acarbose (control) | 27.3 ± 7.3 d (42.3 µM) | 38.1 ± 3.1 d (59.0 µM) |
| Fractions/Compounds | DPPH Scavenging Activity (µg/mL) | ABTS Scavenging Activity (µg/mL) |
|---|---|---|
| Crude extract | 62.3 ± 5.3 a | 66.8 ± 8.3 a |
| n-butanol fraction | 47.6 ± 4.7 b | 43.1 ± 5.2 b |
| Chloroform fraction | NA | NA |
| n-hexane fraction | NA | NA |
| 3-oxolupenal | 82.7 ± 6.7 c (188.5 µM) | 84.2 ± 5.1 c (191.9 µM) |
| Katononic acid | 87.5 ± 5.4 c (192.4 µM) | 82.7 ± 4.7 c (181.9 µM) |
| BHT (control) | 16.6 ± 6.2 d (75.3 µM) | 28.0 ± 1.9 d (127.2 µM) |
| Compounds | Stern–Volmer Constant (KSV) M−1 | Bimolecular Quenching Rate Constant (kq) M−1 s−1 | Number of Binding Sites n | Binding Affinity (Kb) M−1 |
|---|---|---|---|---|
| α-Amylase | ||||
| 3-oxolupenal | 1.31 × 105 | 2.29 × 1013 | 1.0865 | 4.11 × 105 |
| Katononic Acid | 1.58 × 105 | 2.77 × 1013 | 0.7646 | 0.67 × 104 |
| α-Glucosidase | ||||
| 3-oxolupenal | 2.18 × 105 | 3.82 × 1013 | 0.9133 | 0.69 × 105 |
| Katononic Acid | 2.05 × 105 | 3.59 × 1013 | 0.8239 | 1.92 × 104 |
| Compounds | Interacting Residues | Type of Interaction | Distance (Å) | Binding Energy, kcal/mol |
|---|---|---|---|---|
| α-Amylase | ||||
| 3-oxolupenal | Lys200:HZ2-Lig:O | Hydrogen Bond | 1.8982 | −9.1 |
| Leu162-Lig | Hydrophobic (Alkyl) | 5.3493 | ||
| Lig:C-Val163 | Hydrophobic (Alkyl) | 4.5101 | ||
| Lig:C-Leu162 | Hydrophobic (Alkyl) | 5.1043 | ||
| Lig:C-Val163 | Hydrophobic (Alkyl) | 3.8819 | ||
| Tyr62-Lig:C | Hydrophobic (Pi-Alkyl) | 4.1690 | ||
| Tyr62-Lig:C | Hydrophobic (Pi-Alkyl) | 4.0323 | ||
| His299-Lig:C | Hydrophobic (Pi-Alkyl) | 4.7817 | ||
| Katononic acid | Lig:C-Trp59 | Hydrophobic (Pi-Sigma) | 3.7042 | −8.6 |
| Val163-Lig | Hydrophobic (Alkyl) | 4.4902 | ||
| Val163-Lig | Hydrophobic (Alkyl) | 4.2520 | ||
| Lig:C-Val163 | Hydrophobic (Alkyl) | 3.4001 | ||
| Lig:C-Leu165 | Hydrophobic (Alkyl) | 4.9166 | ||
| Trp59-Lig | Hydrophobic (Pi-Alkyl) | 5.4839 | ||
| Trp59-Lig | Hydrophobic (Pi-Alkyl) | 4.9116 | ||
| Trp59-Lig:C | Hydrophobic (Pi-Alkyl) | 4.1964 | ||
| Trp59-Lig | Hydrophobic (Pi-Alkyl) | 4.5411 | ||
| Trp59-Lig:C | Hydrophobic (Pi-Alkyl) | 4.7649 | ||
| Trp59-Lig:C | Hydrophobic (Pi-Alkyl) | 4.3555 | ||
| Tyr62-Lig:C | Hydrophobic (Pi-Alkyl) | 5.3466 | ||
| Tyr62-Lig:C | Hydrophobic (Pi-Alkyl) | 4.3793 | ||
| His101-Lig:C | Hydrophobic (Pi-Alkyl) | 4.7060 | ||
| His305-Lig:C | Hydrophobic (Pi-Alkyl) | 4.3501 | ||
| His305-Lig:C | Hydrophobic (Pi-Alkyl) | 4.8747 | ||
| α-Glucosidase | ||||
| 3-oxolupenal | Arg608:HH21-Lig:O | Hydrogen Bond | 2.4589 | −7.9 |
| Glu866:HN-Lig:O | Hydrogen Bond | 2.5890 | ||
| Val867:HN-Lig:O | Hydrogen Bond | 2.4786 | ||
| Arg585-Lig | Hydrophobic (Alkyl) | 5.2399 | ||
| Val588-Lig | Hydrophobic (Alkyl) | 4.7955 | ||
| Leu865-Lig | Hydrophobic (Alkyl) | 4.9869 | ||
| Lig:C-Val588 | Hydrophobic (Alkyl) | 4.4625 | ||
| Lig:C-Leu865 | Hydrophobic (Alkyl) | 3.6528 | ||
| Lig:C-Arg594 | Hydrophobic (Alkyl) | 4.1884 | ||
| Lig:C-Leu865 | Hydrophobic (Alkyl) | 4.8546 | ||
| His717-Lig:C | Hydrophobic (Pi-Alkyl) | 5.4121 | ||
| His717-Lig:C | Hydrophobic (Pi-Alkyl) | 4.1431 | ||
| Katononic acid | Glu866:HN-Lig:O | Hydrogen Bond | 2.2888 | −7.7 |
| Lys589:CE-Lig:O | Hydrogen Bond | 3.6549 | ||
| Val588-Lig | Hydrophobic (Alkyl) | 4.9374 | ||
| Lig:C-Arg585 | Hydrophobic (Alkyl) | 3.7005 | ||
| Lig:C-Val588 | Hydrophobic (Alkyl) | 4.0007 | ||
| Lig:C-Arg585 | Hydrophobic (Alkyl) | 4.2464 | ||
| Lig:C-Arg594 | Hydrophobic (Alkyl) | 4.0545 | ||
| Lig:C-Leu865 | Hydrophobic (Alkyl) | 5.0477 | ||
| His584-Lig | Hydrophobic (Pi-Alkyl) | 5.2947 | ||
| His584-Lig:C | Hydrophobic (Pi-Alkyl) | 4.2068 | ||
| His584-Lig:C | Hydrophobic (Pi-Alkyl) | 4.4082 | ||
| His717-Lig | Hydrophobic (Pi-Alkyl) | 5.1446 | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alqahtani, A.S.; Hidayathulla, S.; Rehman, M.T.; ElGamal, A.A.; Al-Massarani, S.; Razmovski-Naumovski, V.; Alqahtani, M.S.; El Dib, R.A.; AlAjmi, M.F. Alpha-Amylase and Alpha-Glucosidase Enzyme Inhibition and Antioxidant Potential of 3-Oxolupenal and Katononic Acid Isolated from Nuxia oppositifolia. Biomolecules 2020, 10, 61. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10010061
Alqahtani AS, Hidayathulla S, Rehman MT, ElGamal AA, Al-Massarani S, Razmovski-Naumovski V, Alqahtani MS, El Dib RA, AlAjmi MF. Alpha-Amylase and Alpha-Glucosidase Enzyme Inhibition and Antioxidant Potential of 3-Oxolupenal and Katononic Acid Isolated from Nuxia oppositifolia. Biomolecules. 2020; 10(1):61. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10010061
Chicago/Turabian StyleAlqahtani, Ali S., Syed Hidayathulla, Md Tabish Rehman, Ali A. ElGamal, Shaza Al-Massarani, Valentina Razmovski-Naumovski, Mohammed S. Alqahtani, Rabab A. El Dib, and Mohamed F. AlAjmi. 2020. "Alpha-Amylase and Alpha-Glucosidase Enzyme Inhibition and Antioxidant Potential of 3-Oxolupenal and Katononic Acid Isolated from Nuxia oppositifolia" Biomolecules 10, no. 1: 61. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10010061