The Unique Lipidomic Signatures of Saccharina latissima Can Be Used to Pinpoint Their Geographic Origin

 , , ,
, , ,  and
and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Biochemical and Elemental Compositional Analysis
2.3. Lipid Extraction
2.4. Fatty Acid Analysis Using Gas Chromatography–Mass Spectrometry (GC-MS)
2.5. Hydrophilic Interaction Liquid Chromatography–Mass Spectrometry (HILIC-LC-MS)
2.6. Statistical Analysis
3. Results
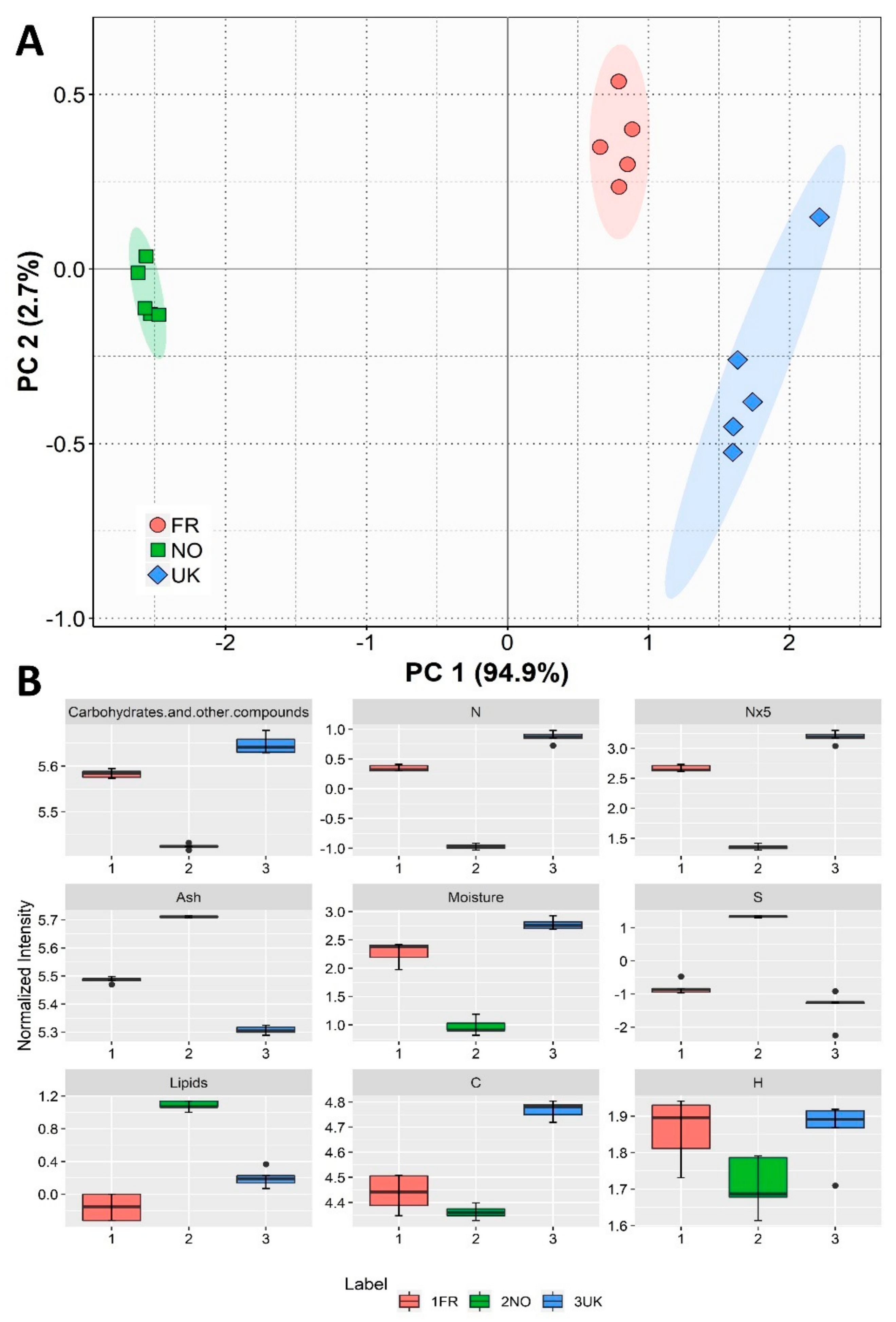
3.1. Biochemical and Elemental Compositional Analysis
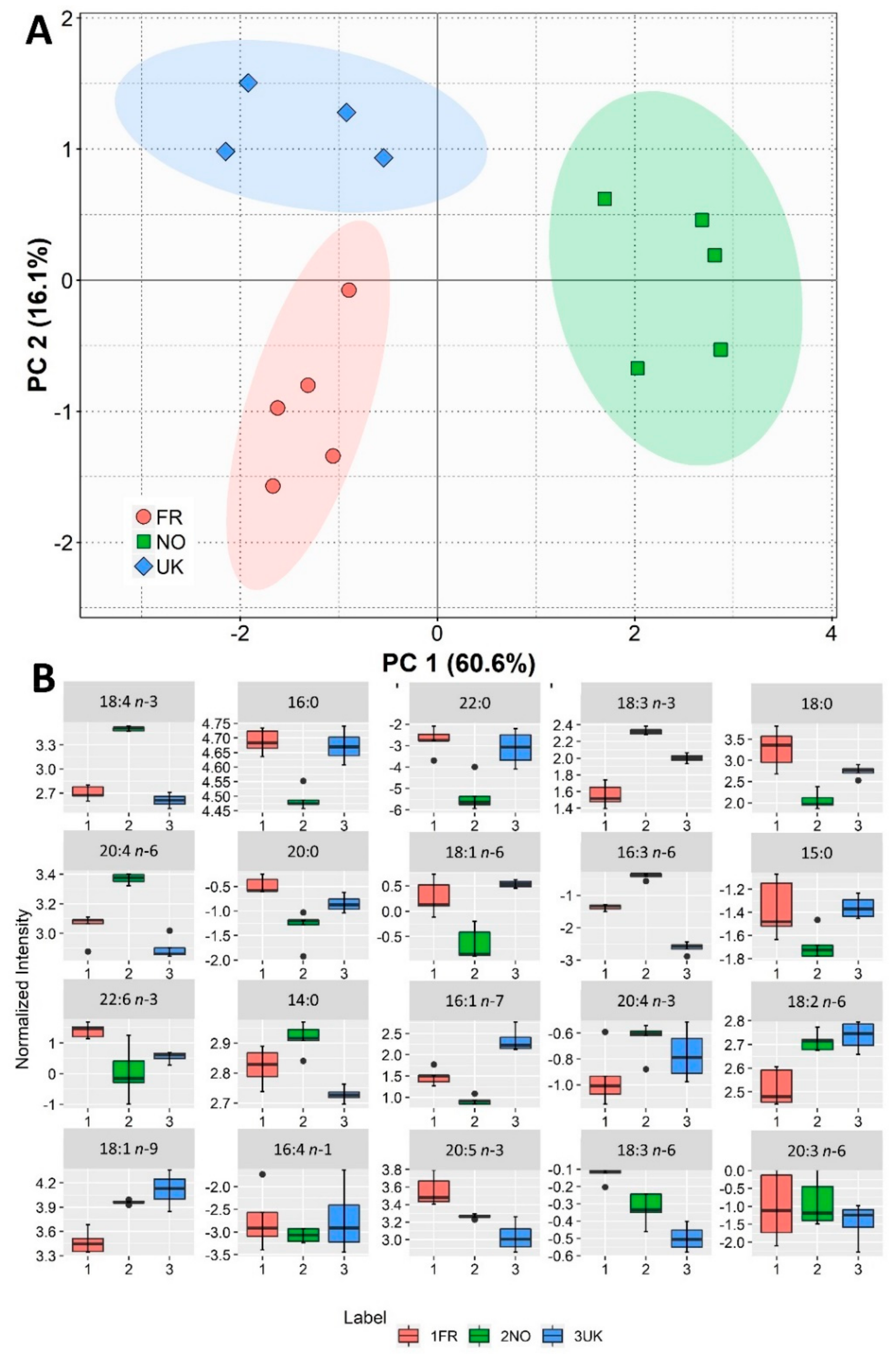
3.2. Fatty Acid Composition
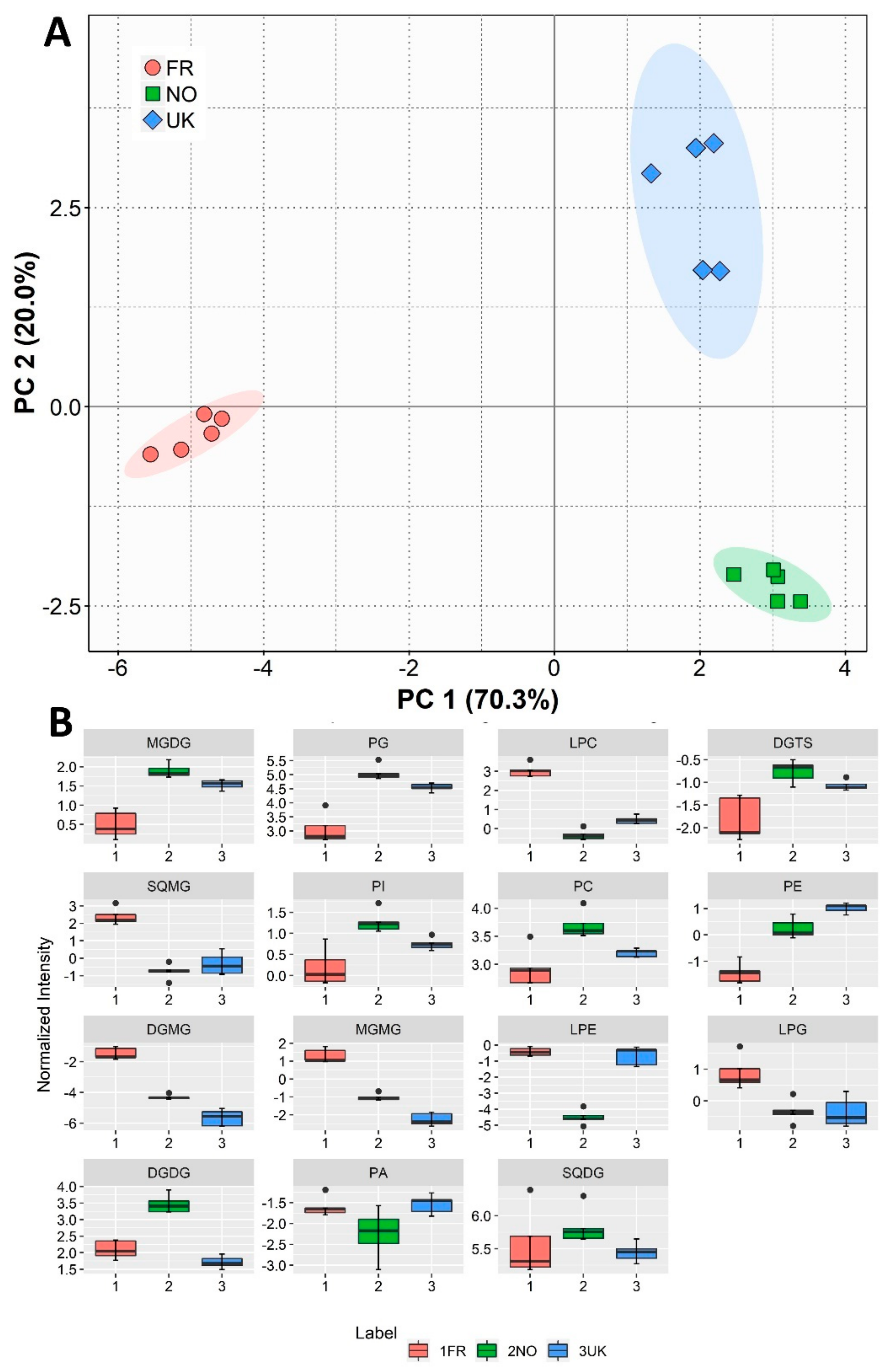
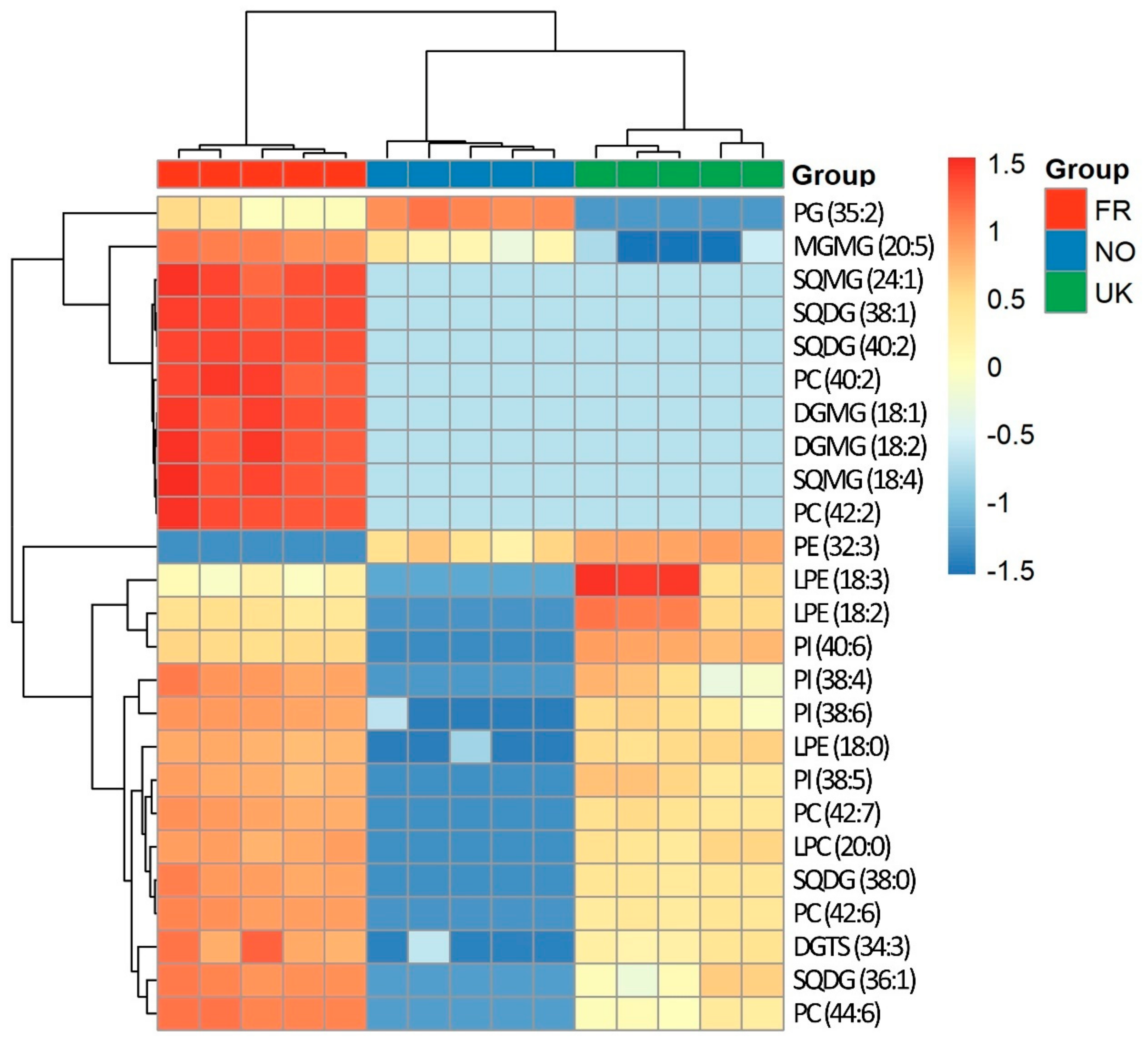
3.3. Polar Lipidome Analysis by HILIC-LC-MS
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Marinho-Soriano, E.; Fonseca, P.C.; Carneiro, M.A.A.; Moreira, W.S.C. Seasonal variation in the chemical composition of two tropical seaweeds. Bioresour. Technol. 2006, 97, 2402–2406. [Google Scholar] [CrossRef]
- Pérez, M.J.; Falqué, E.; Domínguez, H. Antimicrobial action of compounds from marine seaweed. Mar. Drugs 2016, 14, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leal, M.C.; Munro, M.H.G.; Blunt, J.W.; Puga, J.; Jesus, B.; Calado, R.; Rosa, R.; Madeira, C. Biogeography and biodiscovery hotspots of macroalgal marine natural products. Nat. Prod. Rep. 2013, 30, 1380–1390. [Google Scholar] [CrossRef]
- Kim, J.K.; Yarish, C.; Hwang, E.K.; Park, M.; Kim, Y. Seaweed aquaculture: Cultivation technologies, challenges and its ecosystem services. ALGAE 2017, 32, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Neiva, J.; Paulino, C.; Nielsen, M.M.; Krause-Jensen, D.; Saunders, G.W.; Assis, J.; Bárbara, I.; Tamigneaux, É.; Gouveia, L.; Aires, T.; et al. Glacial vicariance drives phylogeographic diversification in the amphi-boreal kelp Saccharina latissima. Sci. Rep. 2018, 8, 1112. [Google Scholar] [CrossRef]
- Broch, O.J.; Alver, M.O.; Bekkby, T.; Gundersen, H.; Forbord, S.; Handå, A.; Skjermo, J.; Hancke, K. The kelp cultivation potential in coastal and offshore regions of Norway. Front. Mar. Sci. 2019, 5. [Google Scholar] [CrossRef]
- Marinho, G.S.; Alvarado-Morales, M.; Angelidaki, I. Valorization of macroalga Saccharina latissima as novel feedstock for fermentation-based succinic acid production in a biorefinery approach and economic aspects. Algal Res. 2016, 16, 102–109. [Google Scholar] [CrossRef] [Green Version]
- Olischläger, M.; Iñiguez, C.; Gordillo, F.J.L.; Wiencke, C. Biochemical composition of temperate and Arctic populations of Saccharina latissima after exposure to increased pCO2 and temperature reveals ecotypic variation. Planta 2014, 240, 1213–1224. [Google Scholar] [CrossRef] [Green Version]
- Ehrig, K.; Alban, S. Sulfated galactofucan from the brown alga Saccharina latissima—Variability of yield, structural composition and bioactivity. Mar. Drugs 2015, 13, 76–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tibbetts, S.M.; Milley, J.E.; Lall, S.P. Nutritional quality of some wild and cultivated seaweeds: Nutrient composition, total phenolic content and in vitro digestibility. J. Appl. Phycol. 2016, 28, 3575–3585. [Google Scholar] [CrossRef]
- Schiener, P.; Black, K.D.; Stanley, M.S.; Green, D.H. The seasonal variation in the chemical composition of the kelp species Laminaria digitata, Laminaria hyperborea, Saccharina latissima and Alaria esculenta. J. Appl. Phycol. 2015, 27, 363–373. [Google Scholar] [CrossRef]
- Marinho, G.S.; Holdt, S.L.; Jacobsen, C.; Angelidaki, I. Lipids and composition of fatty acids of Saccharina latissima cultivated year-round in integrated multi-trophic aquaculture. Mar. Drugs 2015, 13, 4357–4374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rey, F.; Lopes, D.; Maciel, E.; Monteiro, J.; Skjermo, J.; Funderud, J.; Raposo, D.; Domingues, P.; Calado, R.; Domingues, M.R. Polar lipid profile of Saccharina latissima, a functional food from the sea. Algal Res. 2019, 39, 101473. [Google Scholar] [CrossRef]
- Barbosa, M.; Fernandes, F.; Pereira, D.M.; Azevedo, I.C.; Sousa-Pinto, I.; Andrade, P.B.; Valentão, P. Fatty acid patterns of the kelps Saccharina latissima, Saccorhiza polyschides and Laminaria ochroleuca: Influence of changing environmental conditions. Arab. J. Chem. 2017. [Google Scholar] [CrossRef]
- Schmid, M.; Guihéneuf, F.; Stengel, D.B. Fatty acid contents and profiles of 16 macroalgae collected from the Irish Coast at two seasons. J. Appl. Phycol. 2014, 26, 451–463. [Google Scholar] [CrossRef]
- Stengel, D.B.; Connan, S.; Popper, Z.A. Algal chemodiversity and bioactivity: Sources of natural variability and implications for commercial application. Biotechnol. Adv. 2011, 29, 483–501. [Google Scholar] [CrossRef]
- Shalaby, E.A. Influence of abiotic stress on biosynthesis of alga-chemicals and its relation to biological activities. Indian J. Geo Mar. Sci. 2017, 46, 23–32. [Google Scholar]
- Ji, Y.; Xu, Z.; Zou, D.; Gao, K. Ecophysiological responses of marine macroalgae to climate change factors. J. Appl. Phycol. 2016, 28, 2953–2967. [Google Scholar] [CrossRef]
- Singh, S.P.; Singh, P. Effect of temperature and light on the growth of algae species: A review. Renew. Sustain. Energy Rev. 2015, 50, 431–444. [Google Scholar] [CrossRef]
- Chung, I.-C.; Hwang, R.-L.; Lin, S.-H.; Wu, T.-M.; Wu, J.-Y.; Su, S.-W.; Chen, C.-S.; Lee, T.-M. Nutrients, temperature, and salinity as primary factors influencing the temporal dynamics of macroalgal abundance and assemblage structure on a reef of Du-Lang Bay in Taitung in southeastern Taiwan. Bot. Stud. 2007, 48, 419–433. [Google Scholar]
- Stengel, D.; Conde-Álvarez, R.; Connan, S.; Nitschke, U.; Arenas, F.; Abreu, H.; Barufi, J.B.; Chow, F.; Robledo, D.; Malta, E. Short-term effects of CO2, nutrients and temperature on three marine macroalgae under solar radiation. Aquat. Biol. 2014, 22, 159–176. [Google Scholar] [CrossRef] [Green Version]
- Manns, D.M.; Nielsen, M.M.; Bruhn, A.; Saake, B.; Meyer, A.S. Compositional variations of brown seaweeds Laminaria digitata and Saccharina latissima in Danish waters. J. Appl. Phycol. 2017, 29, 1493–1506. [Google Scholar] [CrossRef]
- Peteiro, C.; Freire, Ó. Biomass yield and morphological features of the seaweed Saccharina latissima cultivated at two different sites in a coastal bay in the Atlantic coast of Spain. J. Appl. Phycol. 2013, 25, 205–213. [Google Scholar] [CrossRef]
- Angell, A.R.; Mata, L.; de Nys, R.; Paul, N.A. The protein content of seaweeds: A universal nitrogen-to-protein conversion factor of five. J. Appl. Phycol. 2016, 28, 511–524. [Google Scholar] [CrossRef]
- Da Costa, E.; Melo, T.; Moreira, A.S.P.; Bernardo, C.; Helguero, L.; Ferreira, I.; Cruz, M.T.; Rego, A.M.; Domingues, P.; Calado, R.; et al. Valorization of lipids from Gracilaria sp. through lipidomics and decoding of antiproliferative and anti-inflammatory activity. Mar. Drugs 2017, 15, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, J.P.; Pereira, C.V.; Silva, A.M.; Maciel, E.; Baldeiras, I.; Peixoto, F.; Domingues, M.R.; Jurado, A.S.; Oliveira, P.J. Rapeseed oil-rich diet alters hepatic mitochondrial membrane lipid composition and disrupts bioenergetics. Arch. Toxicol. 2013, 87, 2151–2163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colombo, S.; Melo, T.; Martínez-López, M.; Carrasco, M.J.; Domingues, M.R.; Pérez-Sala, D.; Domingues, P. Phospholipidome of endothelial cells shows a different adaptation response upon oxidative, glycative and lipoxidative stress. Sci. Rep. 2018, 8, 12365. [Google Scholar] [CrossRef] [Green Version]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Orešič, M. MZmine 2: Modular framework for processing, visualizing, and analyzing mass spectrometry-based molecular profile data. BMC Bioinform. 2010, 11, 395. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- RStudio: Integrated Development Environment for R. Inc., Boston, USA, 2016, 14. Available online: https://rstudio.com/ (accessed on 1 May 2019).
- Xia, J.; Sinelnikov, I.V.; Han, B.; Wishart, D.S. MetaboAnalyst 3.0—Making metabolomics more meaningful. Nucleic Acids Res. 2015, 43, W251–W257. [Google Scholar] [CrossRef] [Green Version]
- Murdoch, D.; Chow, E.D. Ellipse: Functions for Drawing Ellipses and Ellipse-Like Confidence Regions. R Package Version 0.3-8 2018. Available online: https://rdrr.io/cran/ellipse/ (accessed on 1 May 2019).
- Kolde, R. Pheatmap: Pretty Heatmaps. R Package Version 1.0.12 2019. Available online: https://rdrr.io/cran/pheatmap/ (accessed on 1 May 2019).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Wickham, H. The split-apply-combine strategy for data analysis. J. Stat. Softw. 2011, 40, 29. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H.; Francois, R.; Henry, L.; Müller, K. dplyr: A Grammar of Data Manipulation. R Package Version 0.7.7. 2018. Available online: https://dplyr.tidyverse.org/ (accessed on 1 May 2019).
- Wickham, H.; Henry, L. tidyr: Easily Tidy Data with ‘spread ()’ and ‘gather ()’ Functions. R Package Version 0.8.1 2018, 3. Available online: https://tidyr.tidyverse.org/ (accessed on 1 May 2019).
- Schmid, M.; Guihéneuf, F.; Stengel, D.B. Ecological and commercial implications of temporal and spatial variability in the composition of pigments and fatty acids in five Irish macroalgae. Mar. Biol. 2017, 164, 158. [Google Scholar] [CrossRef]
- Mansour, H.; Emam, M.M.; Mostafa, N.H. Influence of different habitats on the chemical constituents of Codium tomentosum. Egypt. J. Bot. 2018, 58, 275–285. [Google Scholar] [CrossRef]
- Mišurcová, L.; Kráčmar, S.; Klejdus, B.; Vacek, J. Nitrogen content, dietary fiber, and digestibility in algal food products. Czech J. Food Sci. 2010, 28, 27–35. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, M.M.; Manns, D.; D’Este, M.; Krause-Jensen, D.; Rasmussen, M.B.; Larsen, M.M.; Alvarado-Morales, M.; Angelidaki, I.; Bruhn, A. Variation in biochemical composition of Saccharina latissima and Laminaria digitata along an estuarine salinity gradient in inner Danish waters. Algal Res. 2016, 13, 235–245. [Google Scholar] [CrossRef]
- Sharma, S.; Neves, L.; Funderud, J.; Mydland, L.T.; Øverland, M.; Horn, S.J. Seasonal and depth variations in the chemical composition of cultivated Saccharina latissima. Algal Res. 2018, 32, 107–112. [Google Scholar] [CrossRef]
- Anastasakis, K.; Ross, A.B. Hydrothermal liquefaction of the brown macro-alga Laminaria Saccharina: Effect of reaction conditions on product distribution and composition. Bioresour. Technol. 2011, 102, 4876–4883. [Google Scholar] [CrossRef]
- Khotimchenko, S.V.; Yakovleva, I.M. Lipid composition of the red alga Tichocarpus crinitus exposed to different levels of photon irradiance. Phytochemistry 2005, 66, 73–79. [Google Scholar] [CrossRef]
- Smith, K.L.; Harwood, J.L. Lipids and lipid metabolism in the brown alga, Fucus serratus. Phytochemistry 1984, 23, 2469–2473. [Google Scholar] [CrossRef]
- Jones, A.L.; Harwood, J.L. Lipid metabolism in the brown marine algae Fucus vesiculosus and Ascophyllum nodosum. J. Exp. Bot. 1993, 44, 1203–1210. [Google Scholar] [CrossRef]
- Floreto, E.A.T.; Hirata, H.; Ando, S.; Yamasaki, S. Effects of temperature, light intensity, salinity and source of nitrogen on the growth, total lipid and fatty acid composition of Ulva pertusa Kjellman (Chlorophyta). Bot. Mar. 1993, 36, 149. [Google Scholar] [CrossRef]
- Levy, I.; Maxim, C.; Friedlander, M. Fatty acid distribution among some red algal macrophytes. J. Phycol. 1992, 28, 299–304. [Google Scholar] [CrossRef]
- Floreto, E.A.T.; Teshima, S. The fatty acid composition of seaweeds exposed to different levels of light intensity and salinity. Bot. Mar. 1998, 41, 467–482. [Google Scholar] [CrossRef]
- Dawes, C.J.; Kovach, C.; Friedlander, M. Exposure of Gracilaria to Various Environmental Conditions. II. The Effect on Fatty Acid Composition. Bot. Mar. 1993, 36, 289–296. [Google Scholar] [CrossRef]
- Pinto, E.; Carvalho, A.P.; Cardozo, K.H.M.; Malcata, F.X.; Anjos, F.M.d.; Colepicolo, P. Effects of heavy metals and light levels on the biosynthesis of carotenoids and fatty acids in the macroalgae Gracilaria tenuistipitata (var. liui Zhang & Xia). Rev. Bras. Farmacogn. 2011, 21, 349–354. [Google Scholar]
- Grzelczyk, A.; Gendaszewska-Darmach, E. Novel bioactive glycerol-based lysophospholipids: New data—New insight into their function. Biochimie 2013, 95, 667–679. [Google Scholar] [CrossRef]
- Lee, S.; Suh, S.; Kim, S.; Crain, R.C.; Kwak, J.M.; Nam, H.-G.; Lee, Y. Systemic elevation of phosphatidic acid and lysophospholipid levels in wounded plants. Plant. J. 1997, 12, 547–556. [Google Scholar] [CrossRef]
- Welti, R.; Li, W.; Li, M.; Sang, Y.; Biesiada, H.; Zhou, H.-E.; Rajashekar, C.B.; Williams, T.D.; Wang, X. Profiling membrane lipids in plant stress responses: Role of phospholipase Dalpha in freezing-induced lipid changes in Arabidopsis. J. Biol. Chem. 2002, 277, 31994–32002. [Google Scholar] [CrossRef] [Green Version]
- Khotimchenko, S.V.; Yakovleva, I.M. Effect of solar irradiance on lipids of the green alga Ulva fenestrata Postels et Ruprecht. Bot. Mar. 2004, 47, 395–401. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mean ± SEM | q | |||||
|---|---|---|---|---|---|---|
| Fatty Acid | FR | NO | UK | FR vs. NO | FR vs. UK | NO vs. UK |
| 12:0 | 0.07 ± 0.05 | 0.00 ± 0.00 | 0.00 ± 0.00 | |||
| 14:0 | 7.08 ± 0.13 | 7.55 ± 0.11 | 6.75 ± 0.14 | 0.009 | ||
| 15:0 | 0.39 ± 0.03 | 0.30 ± 0.01 | 0.37 ± 0.02 | |||
| 16:0 | 25.79 ± 0.32 | 22.46 ± 0.26 | 24.99 ± 0.65 | 0.014 | 0.036 | |
| 16:1 n-7 | 2.82 ± 0.17 | 1.90 ± 0.06 | 4.52 ± 0.76 | 0.007 | ||
| 16:3 n-6 | 0.39 ± 0.01 | 0.76 ± 0.02 | 0.16 ± 0.01 | 0.001 | ||
| 16:4 n-1 | 0.16 ± 0.04 | 0.12 ± 0.01 | 0.12 ± 0.01 | |||
| 18:0 | 10.07 ± 1.37 | 4.19 ± 0.28 | 6.11 ± 0.66 | 0.007 | ||
| 18:1 n-9 | 11.10 ± 0.49 | 15.59 ± 0.12 | 17.63 ± 1.03 | 0.004 | ||
| 18:1 n-6 | 1.24 ± 0.14 | 0.65 ± 0.07 | 1.45 ± 0.04 | 0.029 | 0.013 | |
| 18:2 n-6 | 5.73 ± 0.14 | 6.56 ± 0.08 | 6.57 ± 0.16 | 0.013 | 0.013 | |
| 18:3 n-6 | 0.93 ± 0.00 | 0.80 ± 0.02 | 0.72 ± 0.02 | 0.004 | ||
| 18:3 n-3 | 2.95 ± 0.13 | 5.00 ± 0.07 | 4.18 ± 0.19 | 0.002 | ||
| 18:4 n-3 | 6.52 ± 0.17 | 11.32 ± 0.09 | 6.11 ± 0.18 | 0.043 | 0.007 | |
| 20:0 | 0.72 ± 0.04 | 0.41 ± 0.04 | 0.54 ± 0.03 | 0.002 | ||
| 20:3 n-6 | 0.58 ± 0.16 | 0.58 ± 0.13 | 0.45 ± 0.08 | |||
| 20:4 n-6 | 8.51 ± 0.05 | 10.34 ± 0.11 | 7.89 ± 0.47 | 0.006 | ||
| 20:4 n-3 | 0.52 ± 0.04 | 0.64 ± 0.03 | 0.60 ± 0.04 | |||
| 20:5 n-3 | 11.83 ± 0.64 | 9.60 ± 0.07 | 8.42 ± 0.44 | 0.003 | ||
| 22:0 | 0.16 ± 0.03 | 0.02 ± 0.00 | 0.11 ± 0.04 | 0.019 | ||
| 22:6 n-3 | 2.68 ± 0.19 | 1.19 ± 0.33 | 1.64 ± 0.18 | 0.018 | ||
| Indexes | ||||||
| SFA | 44.28 ± 1.71 | 34.95 ± 0.58 | 38.88 ± 1.10 | 0.004 | ||
| MUFA | 15.16 ± 0.67 | 18.14 ± 0.13 | 23.48 ± 1.01 | 0.001 | ||
| PUFA | 40.56 ± 1.37 | 46.91 ± 0.52 | 37.64 ± 1.81 | 0.007 | ||
| PUFA n-3 | 24.50 ± 1.13 | 27.75 ± 0.39 | 21.56 ± 1.29 | 0.007 | ||
| PUFA n-6 | 17.13 ± 0.40 | 19.69 ± 0.22 | 17.25 ± 0.51 | 0.016 | 0.016 | |
| n-6/n-3 | 0.70 ± 0.02 | 0.71 ± 0.01 | 0.81 ± 0.03 | |||
| Sat/Unsat | 0.80 ± 0.05 | 0.54 ± 0.01 | 0.64 ± 0.03 | 0.004 | ||
| ACL | 17.67 ± 0.03 | 17.66 ± 0.02 | 17.55 ± 0.04 | |||
| DBI | 178.3 ± 6.33 | 197.5 ± 2.43 | 167.0 ± 6.91 | 0.018 | ||
| PI | 170.1 ± 6.78 | 178.1 ± 2.82 | 143.6 ± 8.20 | 0.033 | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monteiro, J.P.; Rey, F.; Melo, T.; Moreira, A.S.P.; Arbona, J.-F.; Skjermo, J.; Forbord, S.; Funderud, J.; Raposo, D.; Kerrison, P.D.; et al. The Unique Lipidomic Signatures of Saccharina latissima Can Be Used to Pinpoint Their Geographic Origin. Biomolecules 2020, 10, 107. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10010107
Monteiro JP, Rey F, Melo T, Moreira ASP, Arbona J-F, Skjermo J, Forbord S, Funderud J, Raposo D, Kerrison PD, et al. The Unique Lipidomic Signatures of Saccharina latissima Can Be Used to Pinpoint Their Geographic Origin. Biomolecules. 2020; 10(1):107. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10010107
Chicago/Turabian StyleMonteiro, João P., Felisa Rey, Tânia Melo, Ana S. P. Moreira, Jean-François Arbona, Jorunn Skjermo, Silje Forbord, Jon Funderud, Diogo Raposo, Philip D. Kerrison, and et al. 2020. "The Unique Lipidomic Signatures of Saccharina latissima Can Be Used to Pinpoint Their Geographic Origin" Biomolecules 10, no. 1: 107. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10010107