Quercetin and Coumarin Inhibit Dipeptidyl Peptidase-IV and Exhibits Antioxidant Properties: In Silico, In Vitro, Ex Vivo

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Experimental Animals
2.3. In Silico and Molecular Docking Studies
2.4. DPP-IV Inhibition Activity
2.5. β-Bleaching Inhibition Assay
2.6. Lipid Peroxidation Inhibition Activity
2.7. Erythrocyte Haemolysis Inhibition Activity
2.8. Statistical Analyses
3. Results
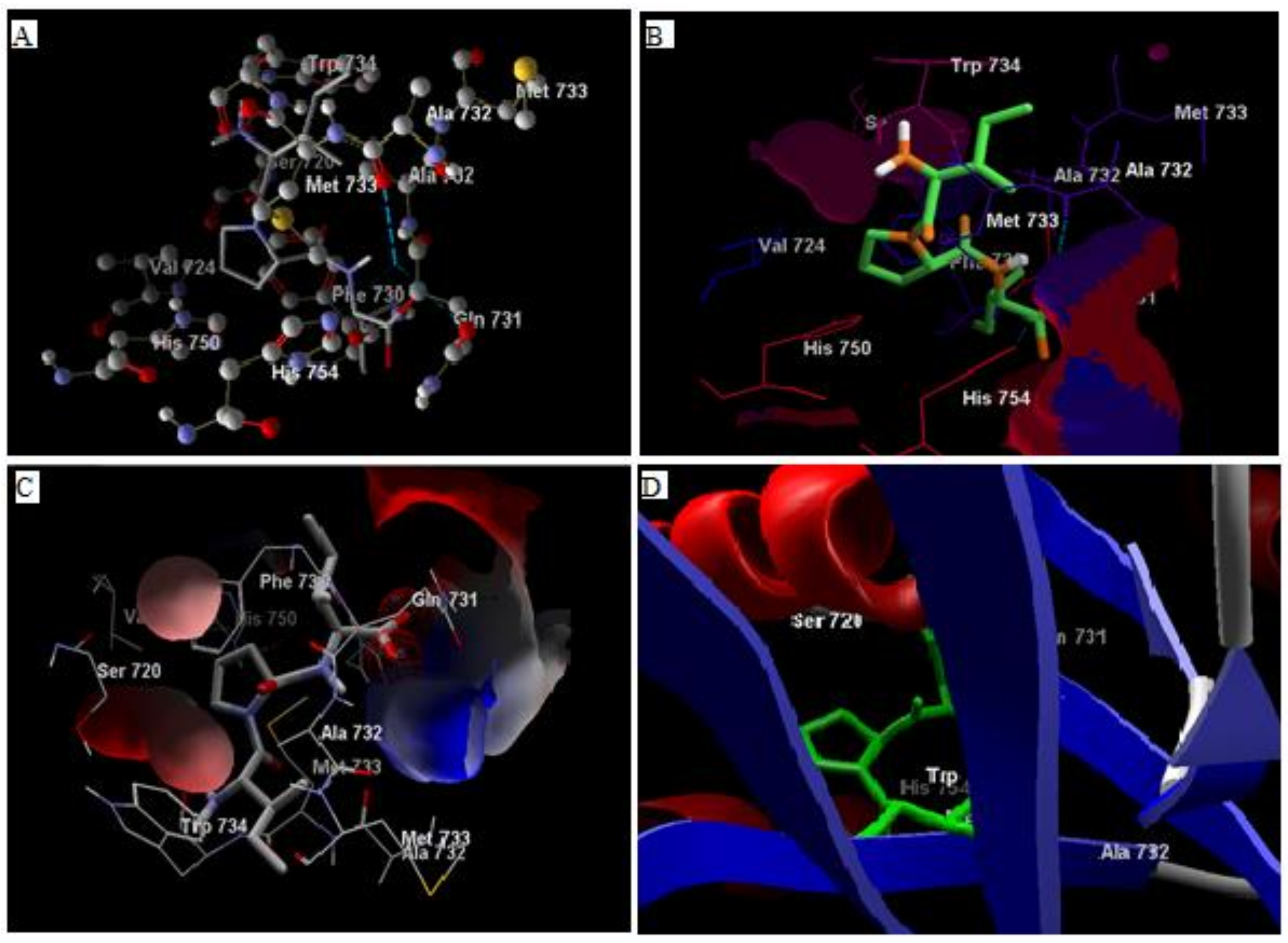
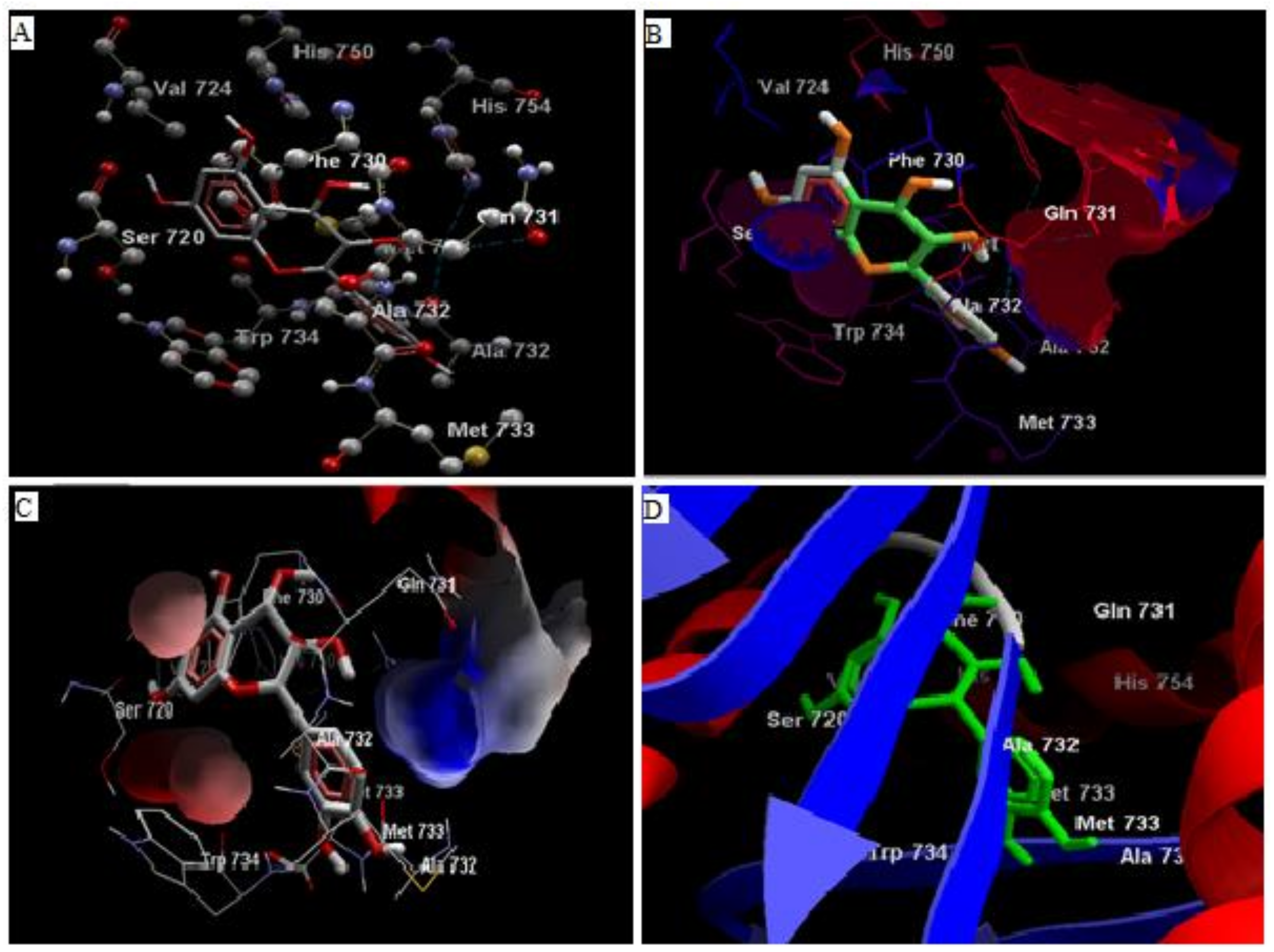
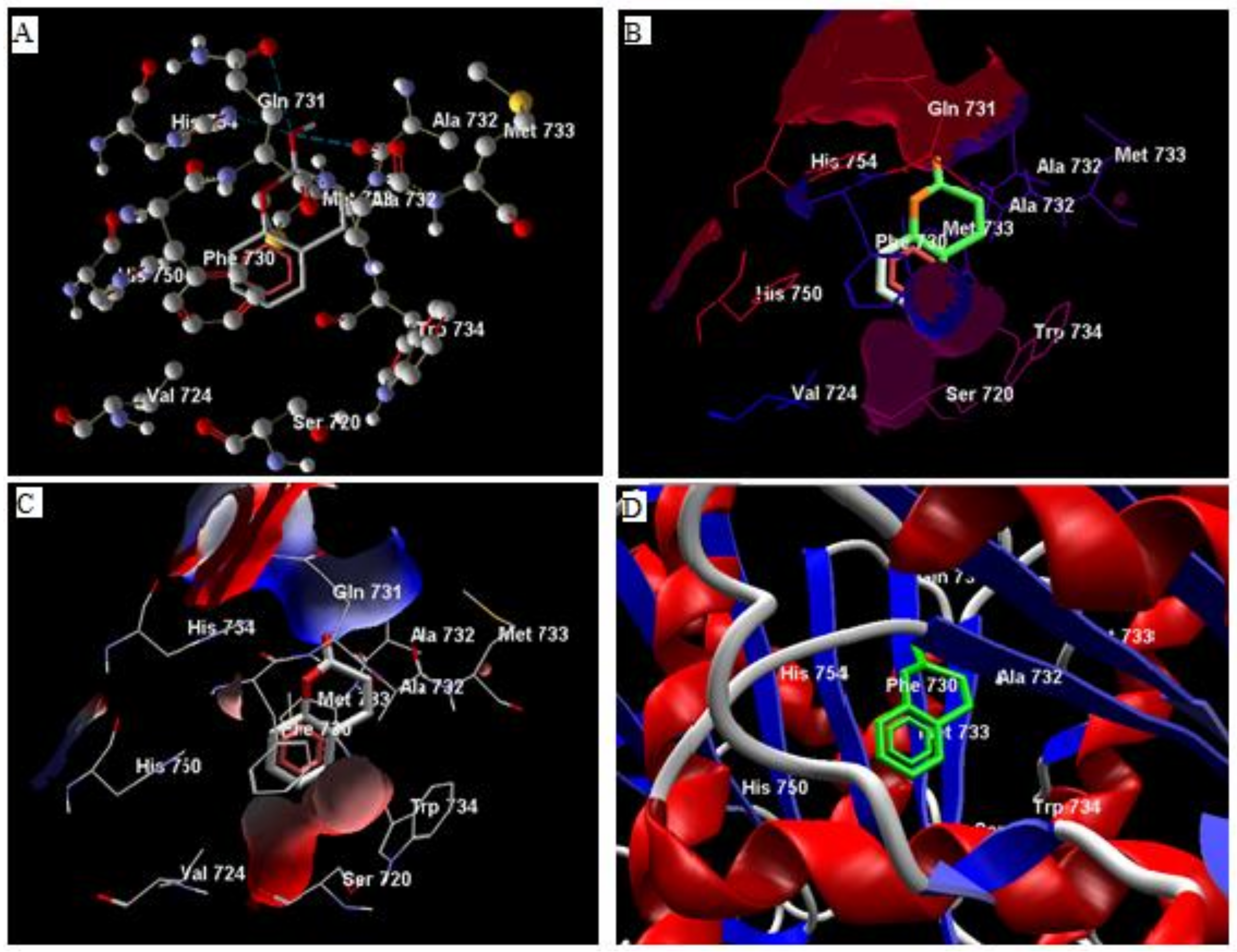
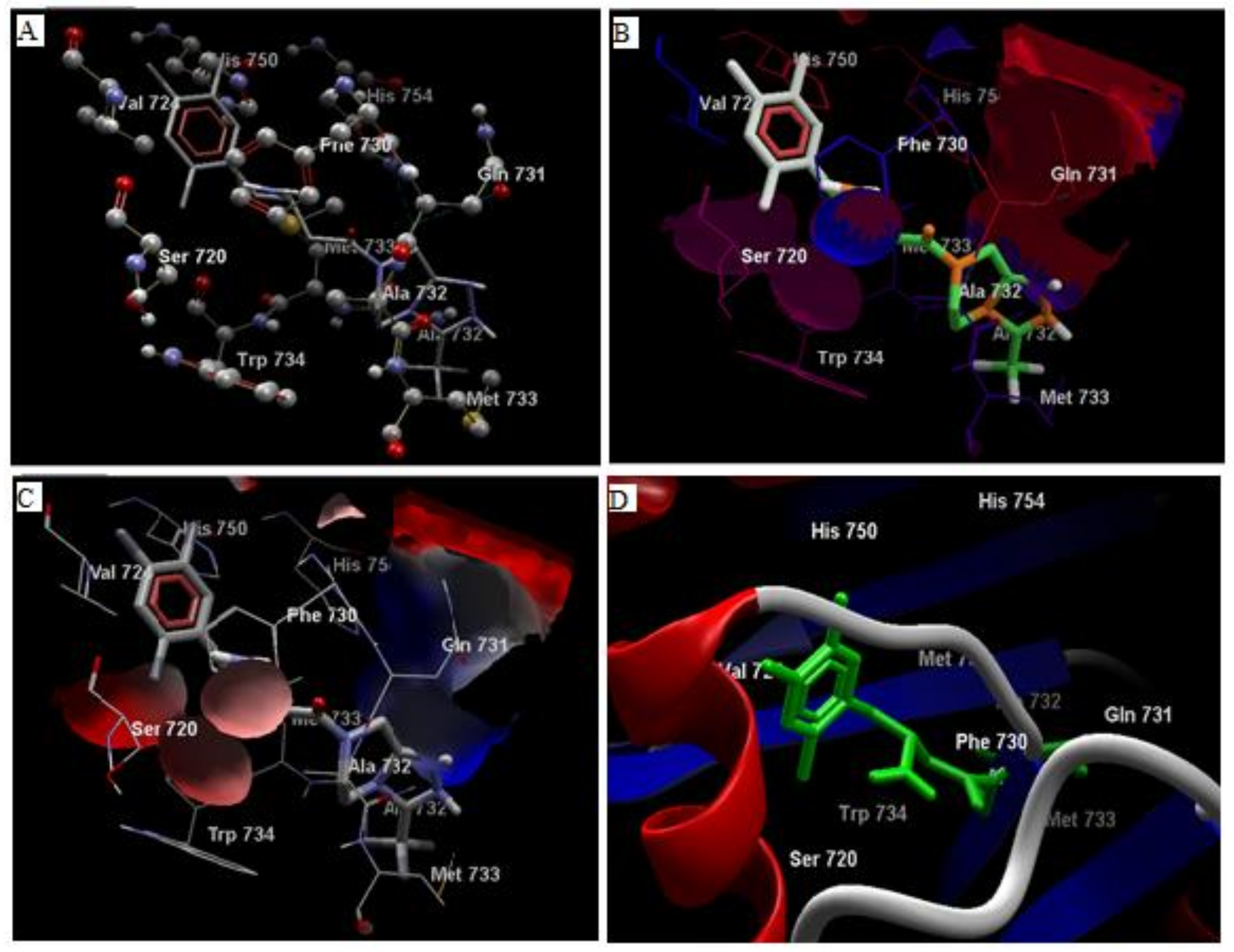
3.1. In Silico Studies
3.2. Analysis of Ligand Binding Affinities
3.3. DPP-IV Inhibition Activity
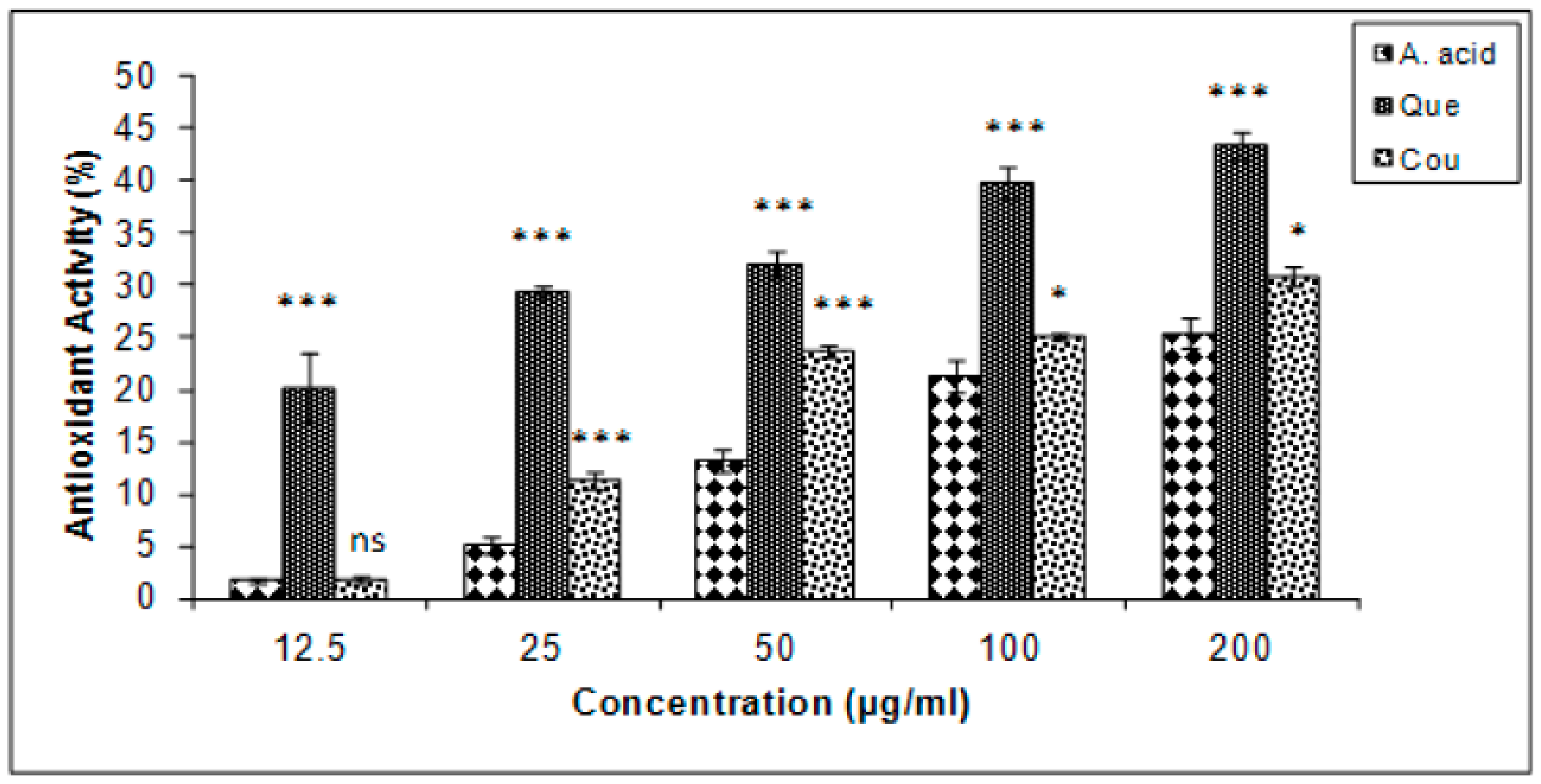
3.4. β-Carotene Bleaching Inhibition
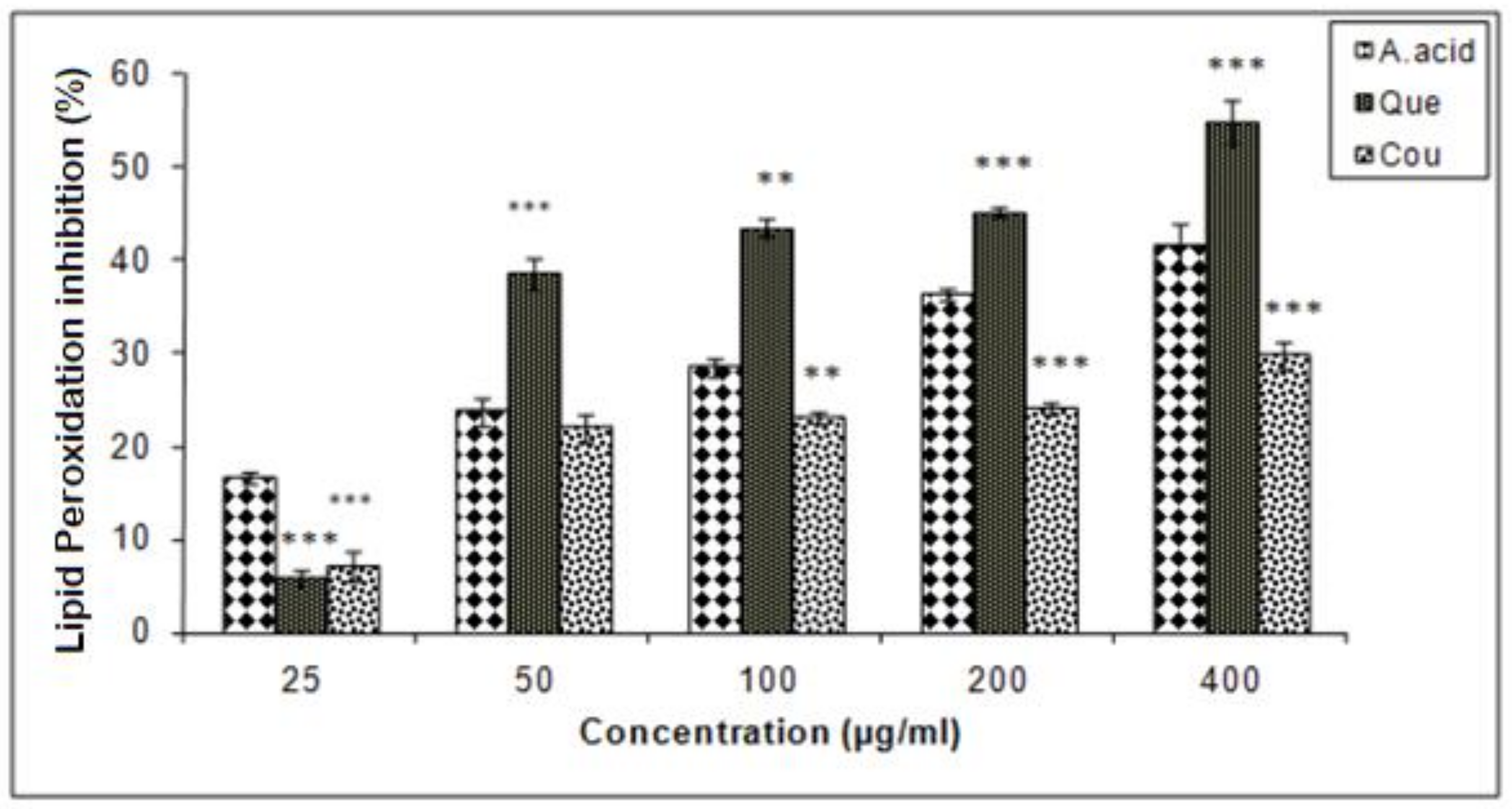
3.5. Hepatic Lipid Peroxidation Inhibition
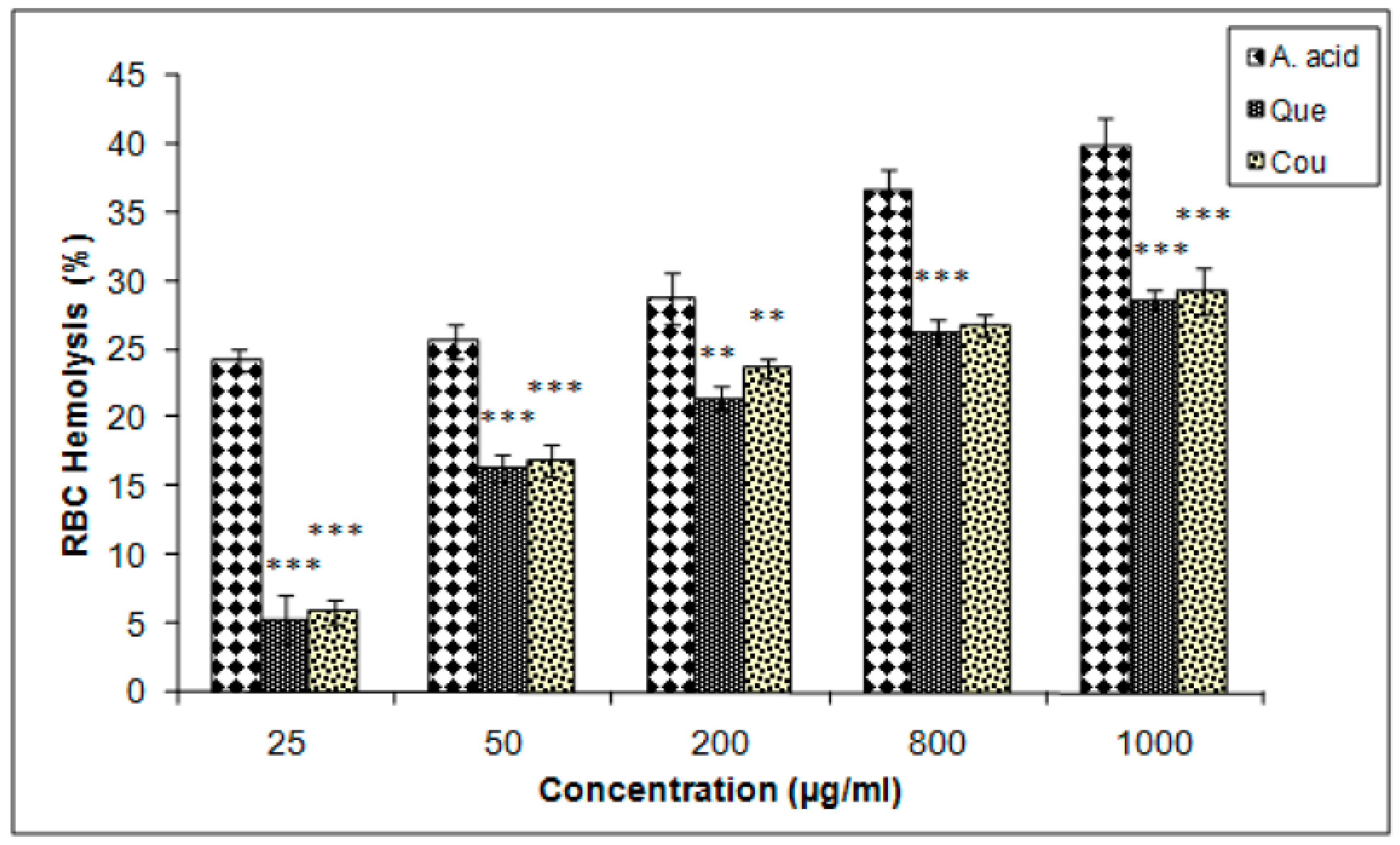
3.6. Erythrocyte Hemolysis Inhibition
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Nyenwe, E.A.; Jerkins, T.W.; Umpierrez, G.E.; Kitabchi, A.E. Management of type 2 diabetes: Evolving strategies for the treatment of patients with type 2 diabetes. Metabolism 2011, 60, 1–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaudhury, A.; Duvoor, C.; Dendi, R.; Sena, V.; Kraleti, S.; Chada, A.; Ravilla, R.; Marco, A.; Shekhawat, N.S.; Montales, M.T. Clinical review of antidiabetic drugs: Implications for type 2 diabetes mellitus management. Front. Endocrinol. (Lausanne) 2017, 8, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holstein, A.; Hammer, C.; Hahn, M.; Kulamadayil, N.-S.-A.; Kovacs, P. Severe sulfonylurea-induced hypoglycemia: A problem of uncritical prescription and deficiencies of diabetes care in geriatric patients. Expert Opin. Drug Saf. 2010, 9, 675–681. [Google Scholar] [CrossRef] [PubMed]
- Amiel, S.; Dixon, T.; Mann, R.; Jameson, K. Hypoglycaemia in type 2 diabetes. Diabet. Med. 2008, 25, 245–254. [Google Scholar] [CrossRef] [Green Version]
- Monnier, L.; Lapinski, H.; Colette, C. Contributions of fasting and postprandial plasma glucose increments to the overall diurnal hyperglycemia of type 2 diabetic patients: Variations with increasing levels of hba1c. Diabetes Care 2003, 26, 881–885. [Google Scholar] [CrossRef] [Green Version]
- Ng, V.W.; Kong, A.P. Dipeptidyl peptidase (dpp)-iv inhibitor: A novel class of oral anti-hyperglycemic agents. Drug Rev. 2007, 12, 33–34. [Google Scholar]
- Nongonierma, A.B.; FitzGerald, R.J. Susceptibility of milk protein-derived peptides to dipeptidyl peptidase iv (dpp-iv) hydrolysis. Food Chem. 2014, 145, 845–852. [Google Scholar] [CrossRef] [Green Version]
- Srinivasan, B.T.; Jarvis, J.; Khunti, K.; Davies, M.J. Recent advances in the management of type 2 diabetes mellitus: A review. Postgrad. Med. J. 2008, 84, 524–531. [Google Scholar] [CrossRef]
- Patil, P.; Mandal, S.; Tomar, S.K.; Anand, S. Food protein-derived bioactive peptides in management of type 2 diabetes. Eur. J. Nutr. 2015, 54, 863–880. [Google Scholar] [CrossRef]
- Kumar, B.; Gupta, S.K.; Nag, T.C.; Srivastava, S.; Saxena, R.; Jha, K.A.; Srinivasan, B.P. Retinal neuroprotective effects of quercetin in streptozotocin-induced diabetic rats. Exp. Eye Res. 2014, 125, 193–202. [Google Scholar] [CrossRef]
- Yokoyama, A.; Yokoyama, A.; Sakakibara, H.; Yokoyama, A.; Sakakibara, H.; Crozier, A.; Kawai, Y.; Matsui, A.; Terao, J.; Kumazawa, S. Quercetin metabolites and protection against peroxynitrite-induced oxidative hepatic injury in rats. Free Radic. Res. 2009, 43, 913–921. [Google Scholar] [CrossRef]
- Yousef, M.I.; Omar, S.A.; El-Guendi, M.I.; Abdelmegid, L.A. Potential protective effects of quercetin and curcumin on paracetamol-induced histological changes, oxidative stress, impaired liver and kidney functions and haematotoxicity in rat. Food Chem. Toxicol. 2010, 48, 3246–3261. [Google Scholar] [CrossRef] [PubMed]
- Arya, A.; Al-Obaidi, M.M.J.; Shahid, N.; Noordin, M.I.B.; Looi, C.Y.; Wong, W.F.; Khaing, S.L.; Mustafa, M.R. Synergistic effect of quercetin and quinic acid by alleviating structural degeneration in the liver, kidney and pancreas tissues of stz-induced diabetic rats: A mechanistic study. Food Chem. Toxicol. 2014, 71, 183–196. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Joshi, J.; Jatwa, R. Dipeptidyl peptidase iv (dpp-iv/cd26) inhibitory and free radical scavenging potential of w. Somnifera and t. Foenum-graecum extract. Int. J. Phytomed. 2014, 5, 503–509. [Google Scholar]
- Gupta, R.; Walunj, S.S.; Tokala, R.K.; Parsa, K.V.; Singh, S.K.; Pal, M. Emerging drug candidates of dipeptidyl peptidase iv (dpp iv) inhibitor class for the treatment of type 2 diabetes. Curr. Drug Targets 2009, 10, 71–87. [Google Scholar] [CrossRef] [PubMed]
- Longenecker, K.L.; Stewart, K.D.; Madar, D.J.; Jakob, C.G.; Fry, E.H.; Wilk, S.; Lin, C.W.; Ballaron, S.J.; Stashko, M.A.; Lubben, T.H. Crystal structures of dpp-iv (cd26) from rat kidney exhibit flexible accommodation of peptidase-selective inhibitors. Biochemistry 2006, 45, 7474–7482. [Google Scholar] [CrossRef]
- Gehlhaar, D.K.; Verkhivker, G.M.; Rejto, P.A.; Sherman, C.J.; Fogel, D.R.; Fogel, L.J.; Freer, S.T. Molecular recognition of the inhibitor ag-1343 by hiv-1 protease: Conformationally flexible docking by evolutionary programming. Chem. Biol. 1995, 2, 317–324. [Google Scholar] [CrossRef] [Green Version]
- Ballester, P.J.; Mitchell, J.B. A machine learning approach to predicting protein–ligand binding affinity with applications to molecular docking. Bioinformatics 2010, 26, 1169–1175. [Google Scholar] [CrossRef] [Green Version]
- Bandaru, S.; Hema Prasad, M.; Jyothy, A.; Nayarisseri, A.; Yadav, M. Binding modes and pharmacophoric features of muscarinic antagonism and β2 agonism (maba) conjugates. Curr. Top. Med. Chem. 2013, 13, 1650–1655. [Google Scholar] [CrossRef]
- Parmar, H.S.; Jain, P.; Chauhan, D.S.; Bhinchar, M.K.; Munjal, V.; Yusuf, M.; Choube, K.; Tawani, A.; Tiwari, V.; Manivannan, E. Dpp-iv inhibitory potential of naringin: An in silico, in vitro and in vivo study. Diabetes Res. Clin. Pract. 2012, 97, 105–111. [Google Scholar] [CrossRef]
- Singh, A.K.; Jatwa, R.; Joshi, J. Cytoprotective and dipeptidyl peptidase iv (dpp-iv/cd26) inhibitory roles of ocimum sanctum and momordicacharantia extract. Asian J. Pharm Clin Res. 2014, 7, 115–120. [Google Scholar]
- Javanmardi, J.; Stushnoff, C.; Locke, E.; Vivanco, J. Antioxidant activity and total phenolic content of iranianocimum accessions. Food Chem. 2003, 83, 547–550. [Google Scholar] [CrossRef]
- Ananthi, S.; Raghavendran, H.R.B.; Sunil, A.G.; Gayathri, V.; Ramakrishnan, G.; Vasanthi, H.R. In vitro antioxidant and in vivo anti-inflammatory potential of crude polysaccharide from turbinariaornata (marine brown alga). Food Chem. Toxicol. 2010, 48, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Miki, M.; Tamai, H.; Mino, M.; Yamamoto, Y.; Niki, E. Free-radical chain oxidation of rat red blood cells by molecular oxygen and its inhibition by α-tocopherol. Arch. Biochem. Biophys. 1987, 258, 373–380. [Google Scholar] [CrossRef]
- Gayathri, P.; Gayathri, D.; Srinivasan, S.; Saroja, S. Screening and quantitation of phytochemicals and nutritional components of the fruit and bark of helicteresisora. Hygeia J. D. Med. 2010, 2, 57–62. [Google Scholar]
- Nongonierma, A.B.; FitzGerald, R.J. Inhibition of dipeptidyl peptidase iv (dpp-iv) by proline containing casein-derived peptides. J. Funct. Foods 2013, 5, 1909–1917. [Google Scholar] [CrossRef] [Green Version]
- Varona, A.; Blanco, L.; Perez, I.; Gil, J.; Irazusta, J.; López, J.I.; Candenas, M.L.; Pinto, F.M.; Larrinaga, G. Expression and activity profiles of dpp iv/cd26 and nep/cd10 glycoproteins in the human renal cancer are tumor-type dependent. BMC Cancer 2010, 10, 193. [Google Scholar] [CrossRef] [Green Version]
- Deacon, C.F. Therapeutic strategies based on glucagon-like peptide 1. Diabetes 2004, 53, 2181–2189. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.K.; Jatwa, R.; Purohit, A.; Ram, H. Synthetic and phytocompounds based dipeptidyl peptidase-iv (dpp-iv) inhibitors for therapeutics of diabetes. J. Asian Nat. Prod. Res. 2017, 19, 1036–1045. [Google Scholar] [CrossRef]
- Ahmed, R.H.; Huri, H.Z.; Al-Hamodi, Z.; Salem, S.D.; Muniandy, S. Serum levels of soluble cd26/dipeptidyl peptidase-iv in type 2 diabetes mellitus and its association with metabolic syndrome and therapy with antidiabetic agents in malaysian subjects. PLoS ONE 2015, 10, e0140618. [Google Scholar] [CrossRef] [Green Version]
- Freeman, J.S. Role of the incretin pathway in the pathogenesis of type 2 diabetes mellitus. Cleve. Clin. J. Med. 2009, 76 (Suppl. 5), S12–S19. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Manson, J.E.; Buring, J.E.; Sesso, H.D.; Liu, S. Associations of dietary flavonoids with risk of type 2 diabetes, and markers of insulin resistance and systemic inflammation in women: A prospective study and cross-sectional analysis. J. Am. Coll. Nutr. 2005, 24, 376–384. [Google Scholar] [CrossRef]
- Treadway, J.L.; Mendys, P.; Hoover, D.J. Glycogen phosphorylase inhibitors for treatment of type 2 diabetes mellitus. Expert Opin. Investig. Drugs 2001, 10, 439–454. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, P.; Vijayakumar, S.; Kothandaraman, S.; Palani, M. Anti-diabetic activity of quercetin extracted from phyllanthusemblica l. Fruit: In silico and in vivo approaches. J. Pharm. Anal. 2018, 8, 109–118. [Google Scholar] [CrossRef]
- Jadhav, R.; Puchchakayala, G.J.G. Hypoglycemic and antidiabetic activity of flavonoids: Boswellic acid, ellagic acid, quercetin, rutin on streptozotocin-nicotinamide induced type 2 diabetic rats. Int. J. Pharm. Pharm. Sci. 2012, 4, 251–256. [Google Scholar]
- Li, Y.Q.; Zhou, F.C.; Gao, F.; Bian, J.S.; Shan, F. Comparative evaluation of quercetin, isoquercetin and rutin as inhibitors of alpha-glucosidase. J. Agric. Food Chem. 2009, 57, 11463–11468. [Google Scholar] [CrossRef]
- Adisakwattana, S.; Roengsamran, S.; Hsu, W.H.; Yibchok-anun, S. Mechanisms of antihyperglycemic effect of p-methoxycinnamic acid in normal and streptozotocin-induced diabetic rats. Life Sci. 2005, 78, 406–412. [Google Scholar] [CrossRef]
- Bharti, S.K.; Krishnan, S.; Kumar, A.; Rajak, K.K.; Murari, K.; Bharti, B.K.; Gupta, A.K. Antidiabetic activity and molecular docking of fructooligosaccharides produced by aureobasidium pullulans in poloxamer-407-induced T2DM rats. Food Chem. 2013, 136, 813–821. [Google Scholar] [CrossRef]
- Lacroix, I.M.E.; Li-Chan, E.C.Y. Evaluation of the potential of dietary proteins as precursors of dipeptidyl peptidase (dpp)-iv inhibitors by an in silico approach. J. Funct. Foods 2012, 4, 403–422. [Google Scholar] [CrossRef]
- Nisha, J. Molecular Docking Analysis of Potential Dipeptidyl peptidase-4 (DPP-4) Inhibitors from Siddha formulation PungampooChooranam for treating Diabetes mellitus. Int. J. Adv. Res. Biol. Sci 2017, 4, 78–85. [Google Scholar] [CrossRef]
- Bucolo, C.; Ward, K.W.; Mazzon, E.; Cuzzocrea, S.; Drago, F. Protective effects of a coumarin derivative in diabetic rats. Invest. Ophthalmol. Vis. Sci. 2009, 50, 3846–3852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manna, P.; Jain, S.K. Obesity, oxidative stress, adipose tissue dysfunction, and the associated health risks: Causes and therapeutic strategies. Metab. Syndr. Relat. Disord. 2015, 13, 423–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mortensen, A.; Skibsted, L.H.; Sampson, J.; Rice-Evans, C.; Everett, S.A. Comparative mechanisms and rates of free radical scavenging by carotenoid antioxidants. FEBS Lett. 1997, 418, 91–97. [Google Scholar] [CrossRef]
- Kuo, C.T.; Liu, T.H.; Hsu, T.H.; Lin, F.Y.; Chen, H.Y. Antioxidant and antiglycation properties of different solvent extracts from chinese olive (canarium album l.) fruit. Asian Pac. J. Trop. Med. 2015, 8, 1013–1021. [Google Scholar] [CrossRef]
- Khan, R.A. Evaluation of flavonoids and diverse antioxidant activities of sonchusarvensis. Chem. Cent. J. 2012, 6, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayaprakasha, G.K.; Singh, R.P.; Sakariah, K.K. Antioxidant activity of grape seed (vitisvinifera) extracts on peroxidation models in vitro. Food Chem. 2001, 73, 285–290. [Google Scholar] [CrossRef]
- Davies, K.J.; Goldberg, A.L. Oxygen radicals stimulate intracellular proteolysis and lipid peroxidation by independent mechanisms in erythrocytes. J. Biol. Chem. 1987, 262, 8220–8226. [Google Scholar]
- Kumar, P.; Sharma, S.; Khanna, M.; Raj, H.G. Effect of quercetin on lipid peroxidation and changes in lung morphology in experimental influenza virus infection. Int. J. Exp. Pathol. 2003, 84, 127–133. [Google Scholar] [CrossRef]
- Duangnin, N.; Phitak, T.; Pothacharoen, P.; Kongtawelert, P. In vitro and in vivo investigation of natural compounds from seed extract of mucunapruriens lacking l-dopa for the treatment of erectile dysfunction. Asian Pac. J. Trop. Med. 2017, 10, 238–252. [Google Scholar] [CrossRef]
- Meir, S.; Kanner, J.; Akiri, B.; Philosoph-Hadas, S. Determination and involvement of aqueous reducing compounds in oxidative defense systems of various senescing leaves. J. Agric. Food Chem. 1995, 43, 1813–1819. [Google Scholar] [CrossRef]
- Kruckeberg, W.C.; Doorenbos, D.I.; Brown, P.O. Genetic differences in hemoglobin influence on erythrocyte oxidative stress hemolysis. Blood 1987, 70, 909–914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizvi, S.I.; Pandey, K.B. Activation of the erythrocyte plasma membrane redox system by resveratrol: A possible mechanism for antioxidant properties. Pharmacol. Rep. 2010, 62, 726–732. [Google Scholar] [CrossRef]
- Turdu, G.; Gao, H.; Jiang, Y.; Kabas, M. Plant dipeptidyl peptidase-iv inhibitors as antidiabetic agents: A brief review. Future Med. Chem. 2018, 10, 1229–1239. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S.No | Name of Molecule | Mol Dock Score | Rerank Score | H Bond |
|---|---|---|---|---|
| 1 | Diprotin-A | −107.70 | −6.68 | −6.58 |
| 2 | Quercetin | −85.49 | −72.54 | −11.21 |
| 3 | Coumarin | −54.17 | −48.50 | −3.180 |
| 4 | Sitagliptin | −108.25 | −77.68 | −2.29 |
| Sample | Concentration (nmol/mL) | Inhibition (%) | IC50 (nmol/mL) |
|---|---|---|---|
| Diprotin-A (Ile-Pro-Ile) Positive control | 0.19 | 22.72 | 0.653 |
| 0.37 | 40.61 | ||
| 0.73 | 62.82 | ||
| 1.46 | 74.36 | ||
| Coumarin | 3.42 | 16.91 | 54.83 |
| 13.68 | 28.39 | ||
| 27.37 | 47.64 | ||
| 136.85 | 62.75 | ||
| Quercetin | 1.65 | 21.93 | 4.02 |
| 6.62 | 58.15 | ||
| 13.23 | 80.88 | ||
| 66.17 | 84.09 | ||
| Sitagliptin | 3.32 | 48.6 | 5.49 |
| 49.10 | 77.9 | ||
| 98.20 | 81.4 | ||
| 147.32 | 86.4 | ||
| 196.41 | 90.4 | ||
| 245.51 | 95.7 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, A.-K.; Patel, P.K.; Choudhary, K.; Joshi, J.; Yadav, D.; Jin, J.-O. Quercetin and Coumarin Inhibit Dipeptidyl Peptidase-IV and Exhibits Antioxidant Properties: In Silico, In Vitro, Ex Vivo. Biomolecules 2020, 10, 207. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10020207
Singh A-K, Patel PK, Choudhary K, Joshi J, Yadav D, Jin J-O. Quercetin and Coumarin Inhibit Dipeptidyl Peptidase-IV and Exhibits Antioxidant Properties: In Silico, In Vitro, Ex Vivo. Biomolecules. 2020; 10(2):207. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10020207
Chicago/Turabian StyleSingh, Anand-Krishna, Pankaj Kumar Patel, Komal Choudhary, Jaya Joshi, Dhananjay Yadav, and Jun-O Jin. 2020. "Quercetin and Coumarin Inhibit Dipeptidyl Peptidase-IV and Exhibits Antioxidant Properties: In Silico, In Vitro, Ex Vivo" Biomolecules 10, no. 2: 207. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10020207