Fluorescein Derivatives as Antibacterial Agents Acting via Membrane Depolarization


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
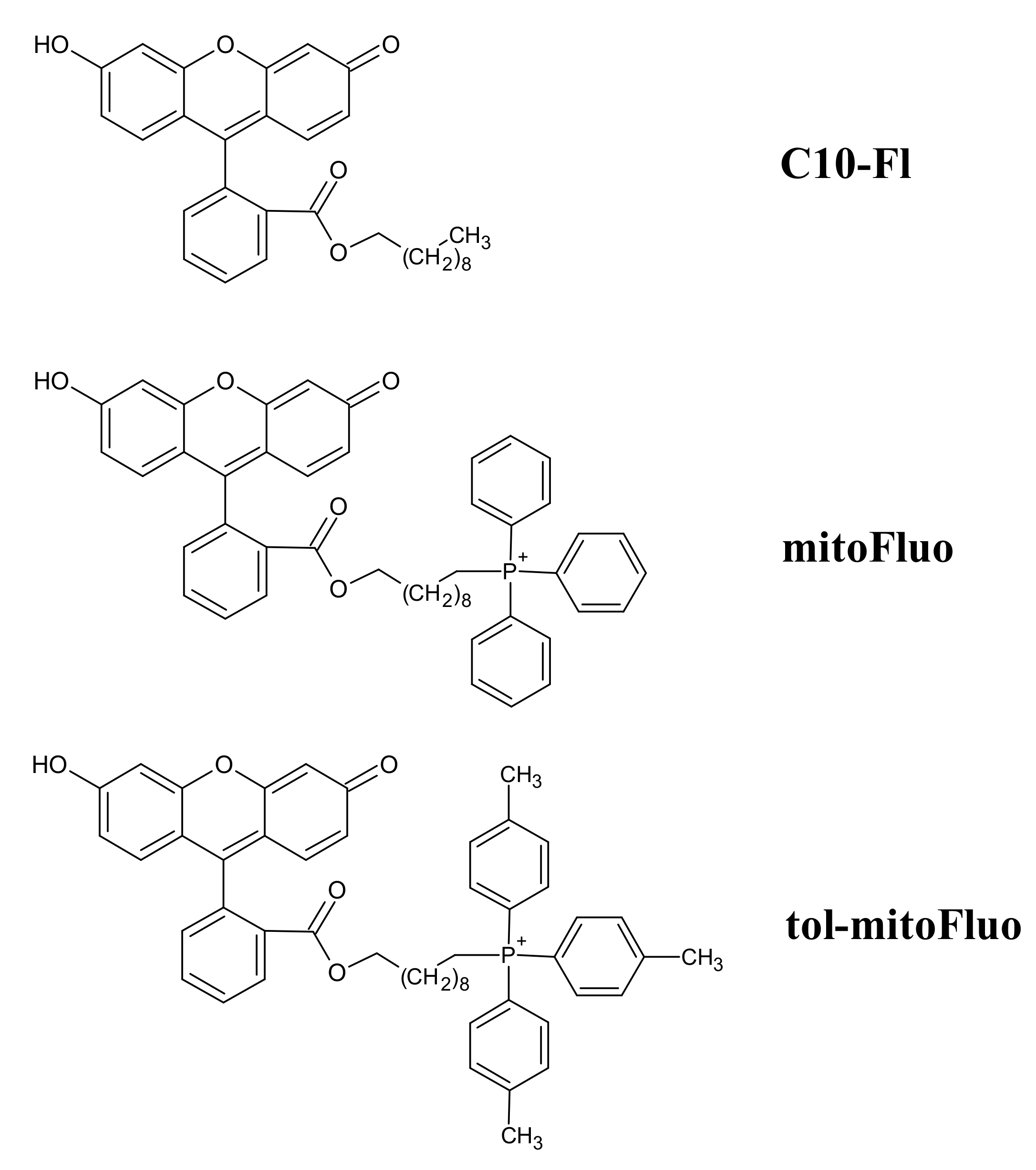
2.2. Synthesis of mitoFluo, tol-mitoFluo and C10-FL
2.2.1. Synthesis of 10-Bromodecyl(tri-p-tolyl)phosphonium Bromide
2.2.2. Synthesis of 10-[2-(3-Hydroxy-6-oxo-xanthen-9-yl)benzoyl]oxydecyl(tri-p-tolyl)phosphonium Bromide (tol-mitoFluo)
2.3. Isolation of Rat Liver Mitochondria
2.4. Measurement of Mitochondrial Membrane Potential and Respiration
2.5. Bacterial Strains
2.6. Growth Suppression Assay and MIC Determination
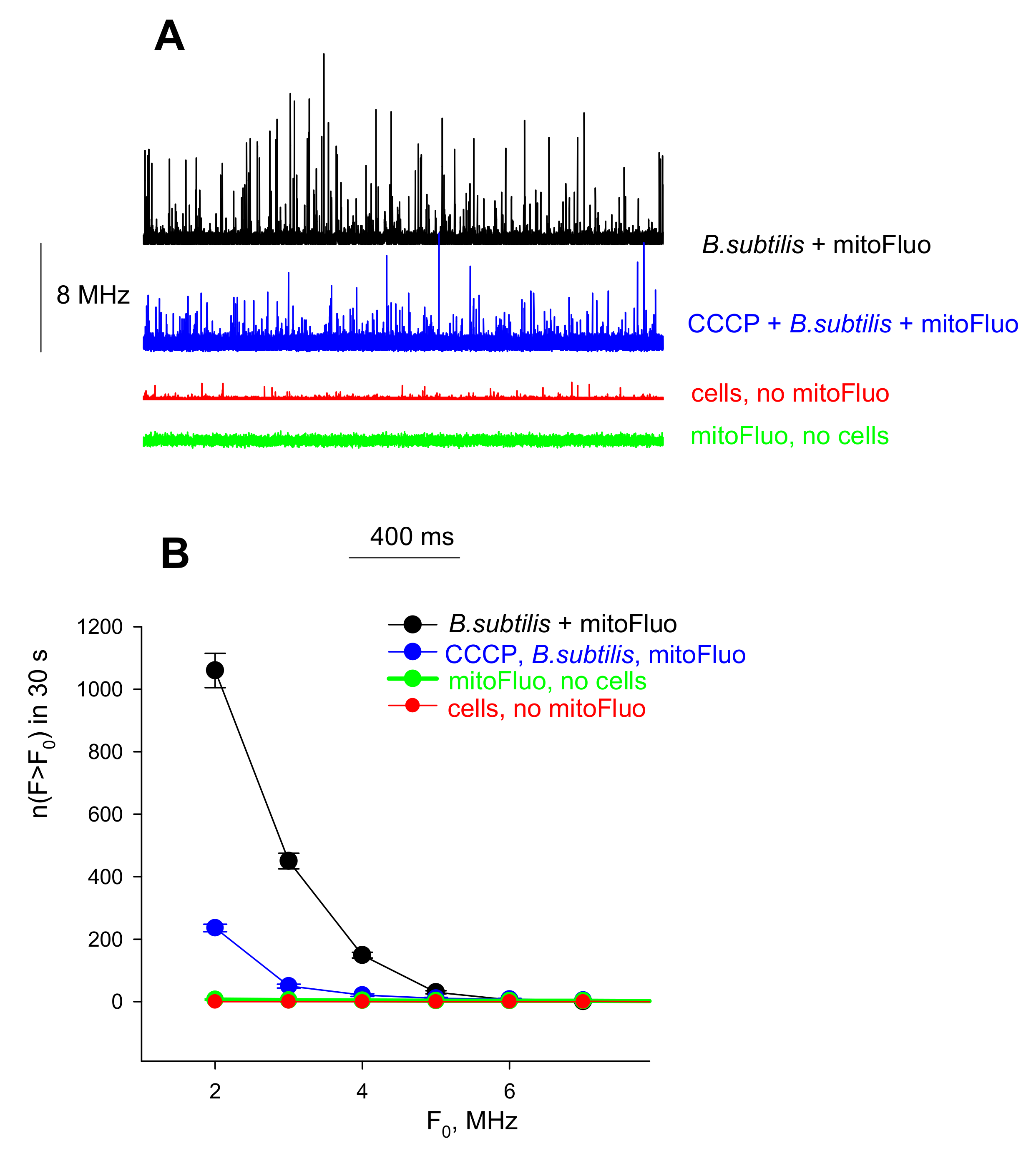
2.7. Bacterial Depolarization
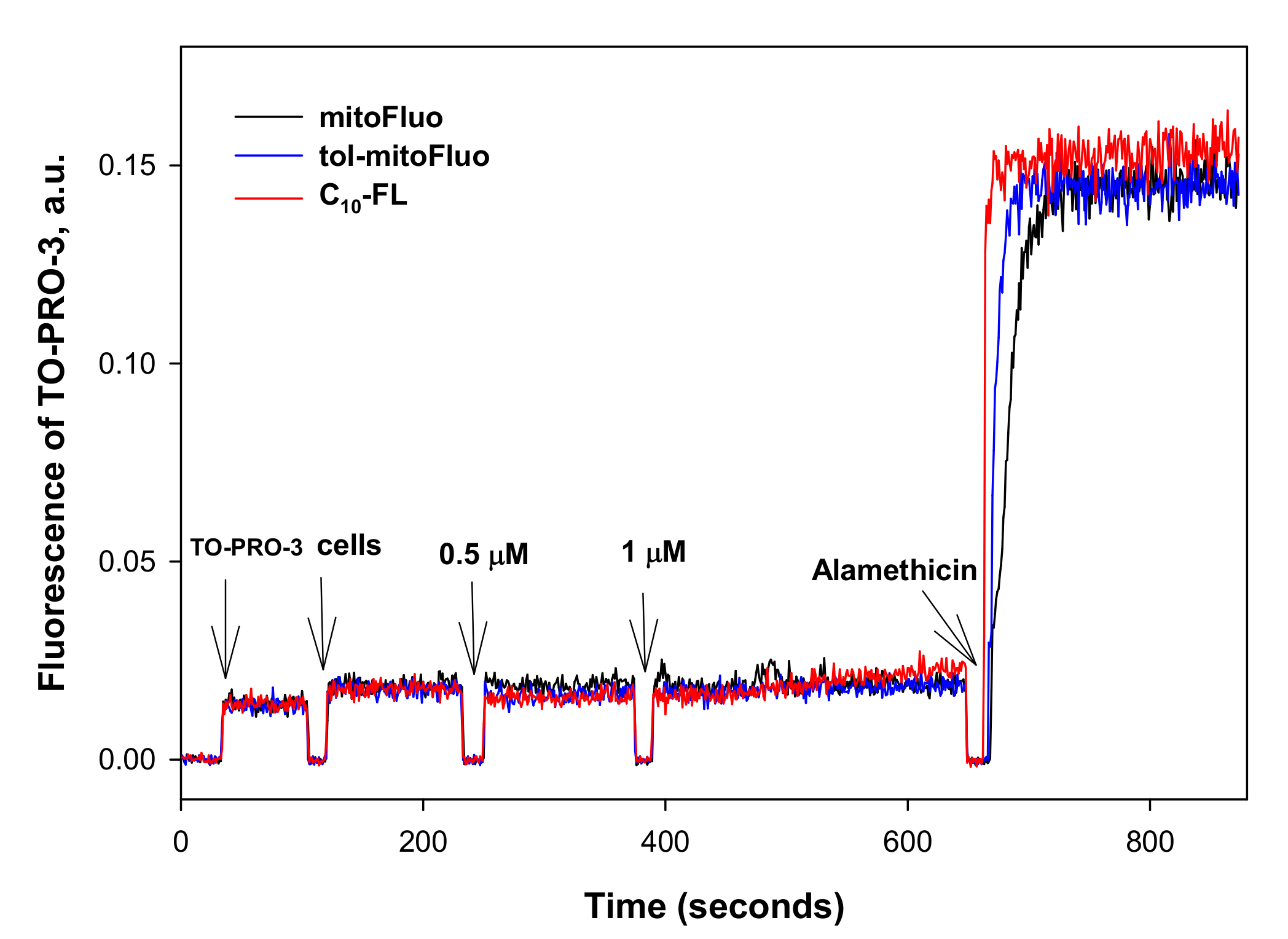
2.8. Bacterial Permeabilization
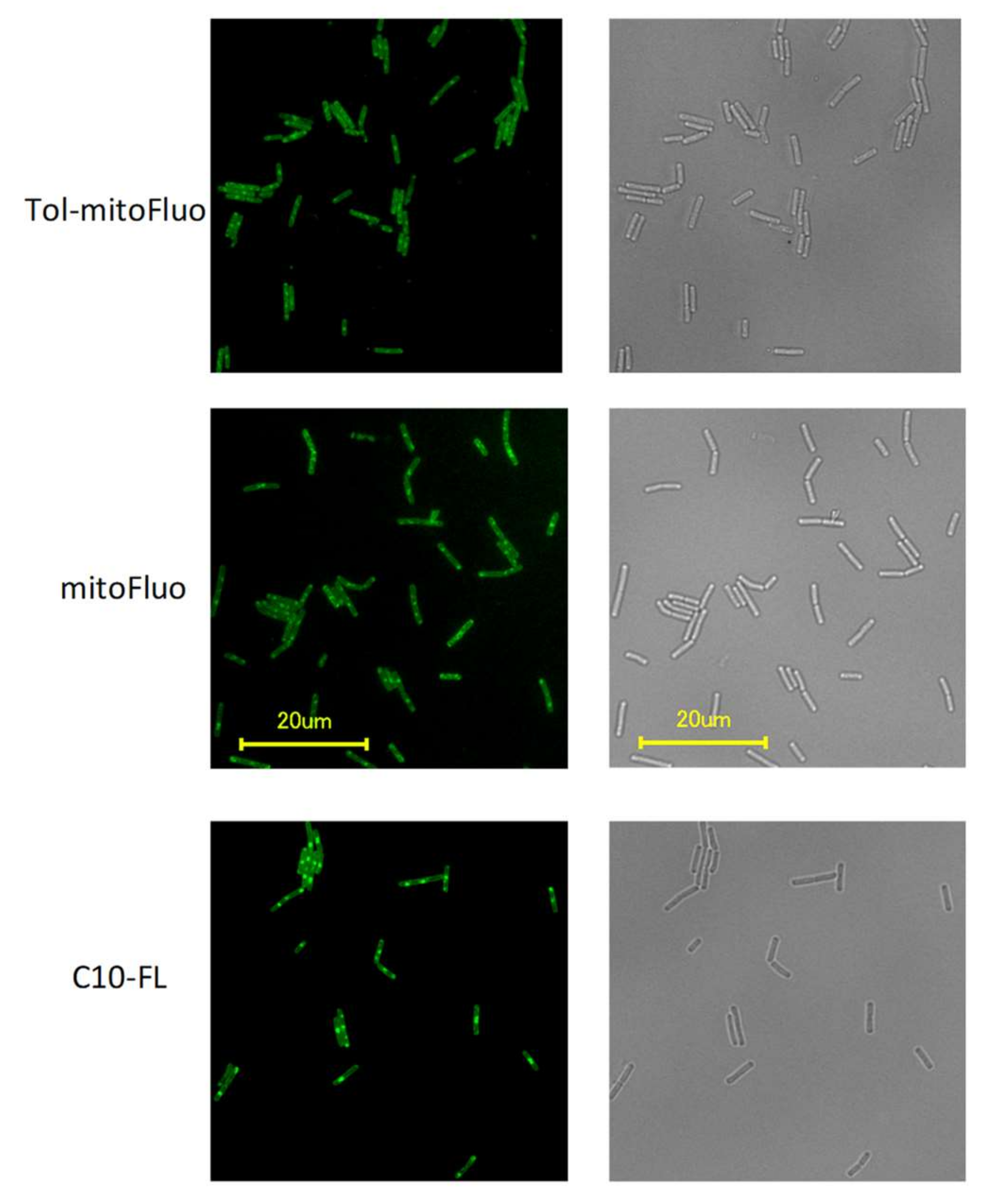
2.9. Fluorescence Microscopy
2.9.1. Agarose Pads Preparation
2.9.2. Sample Pads Preparation
2.9.3. Equipment Setup
2.10. Fluorescence Correlation Spectroscopy
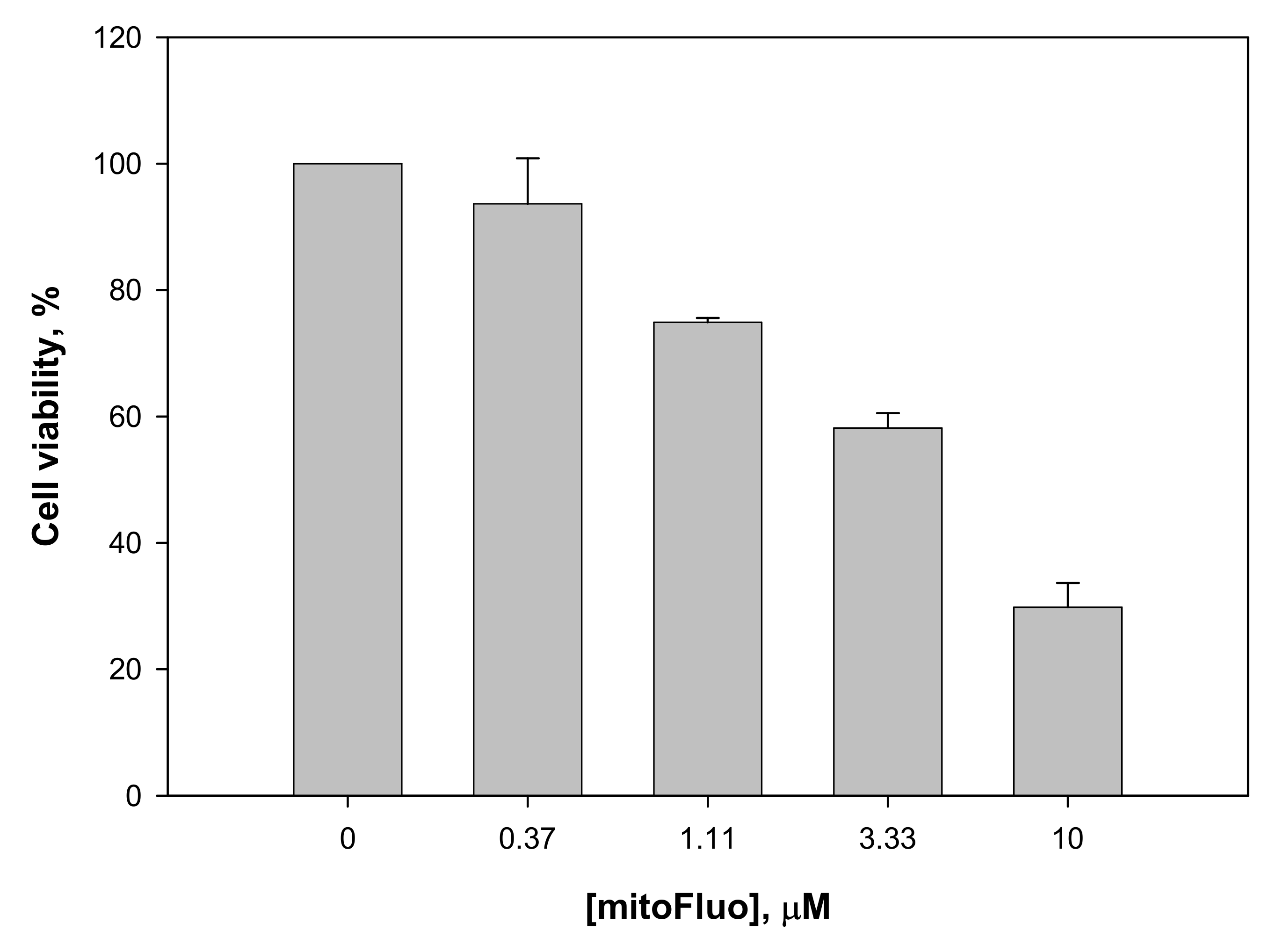
2.11. Experiments with Human Cell Line Rko
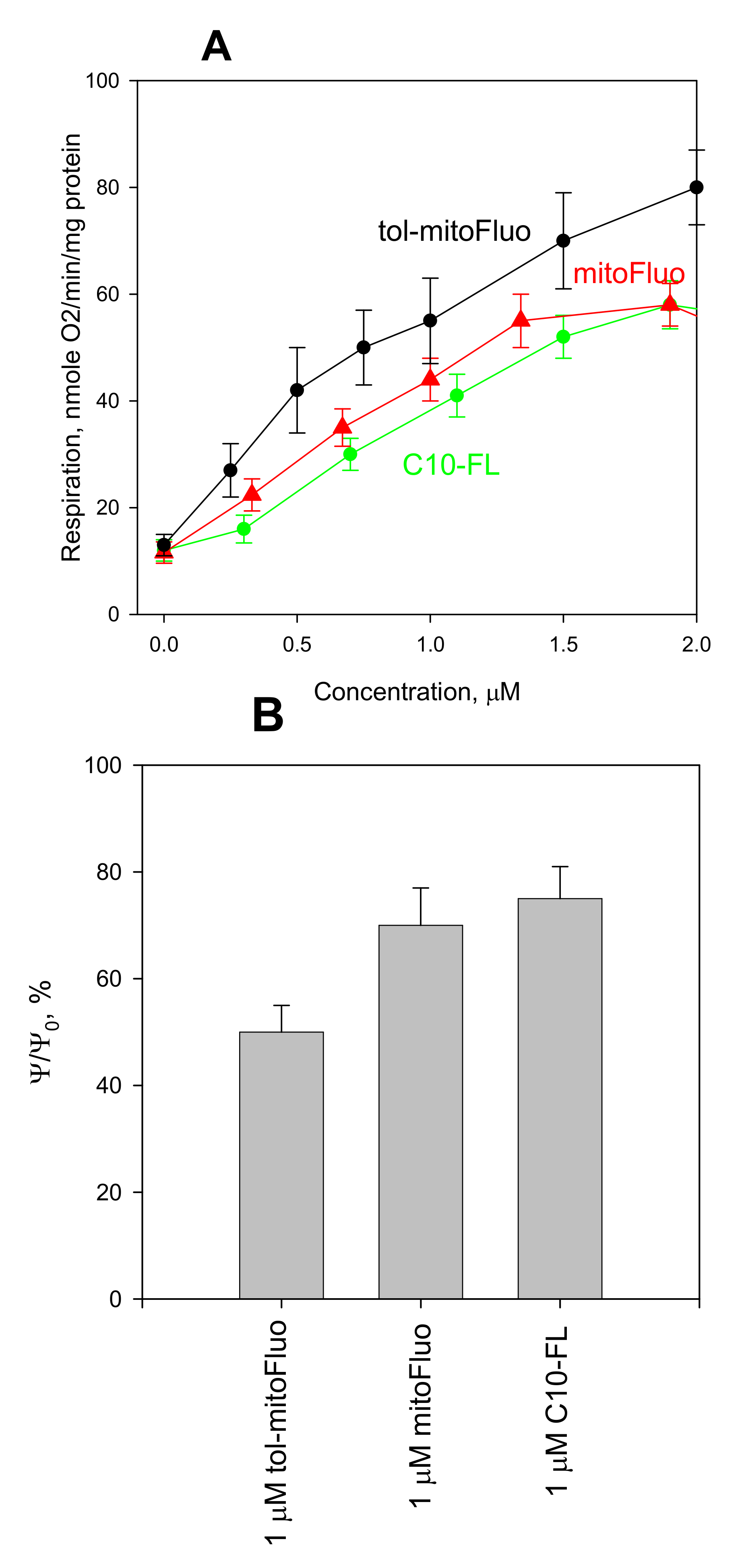
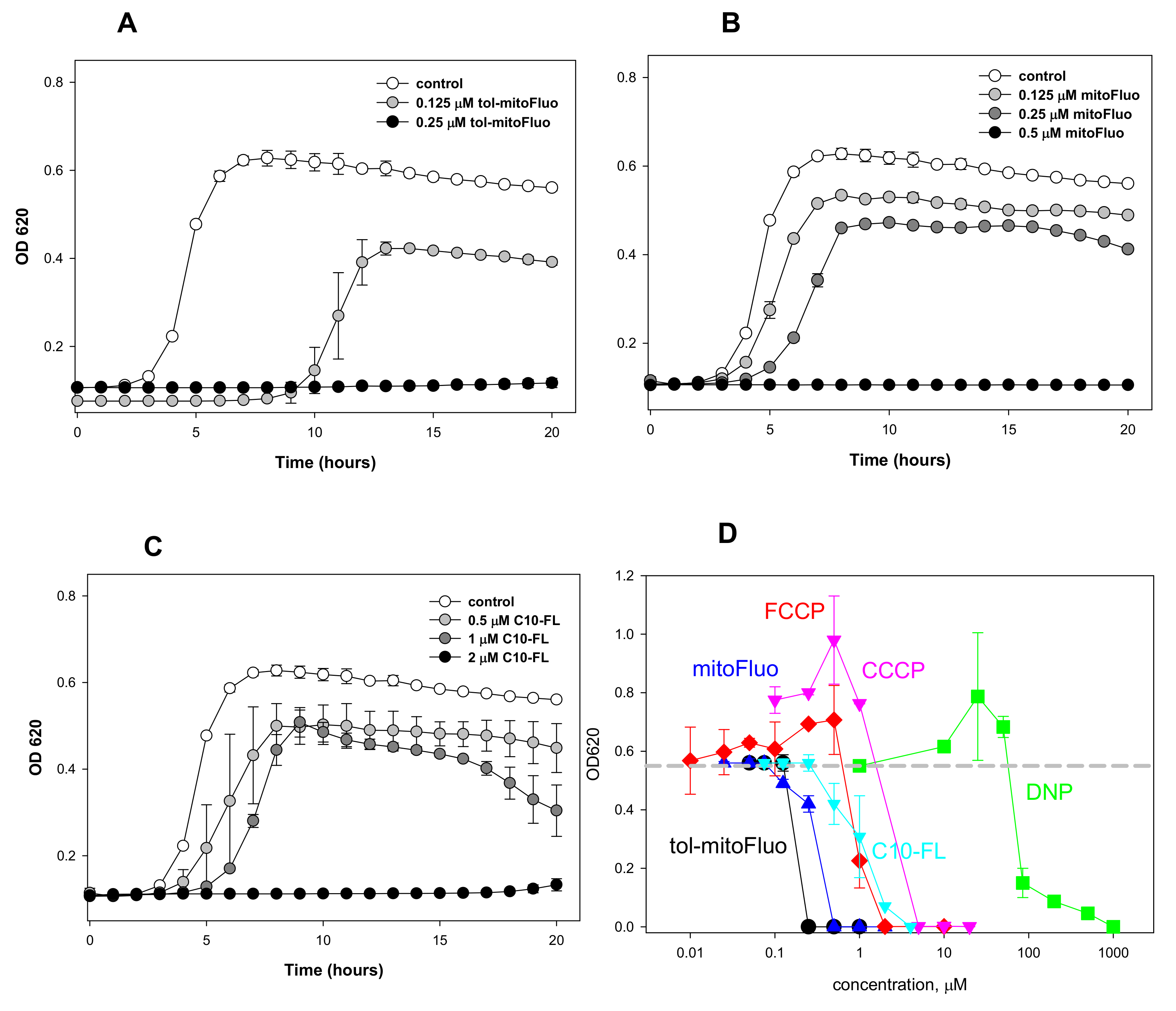
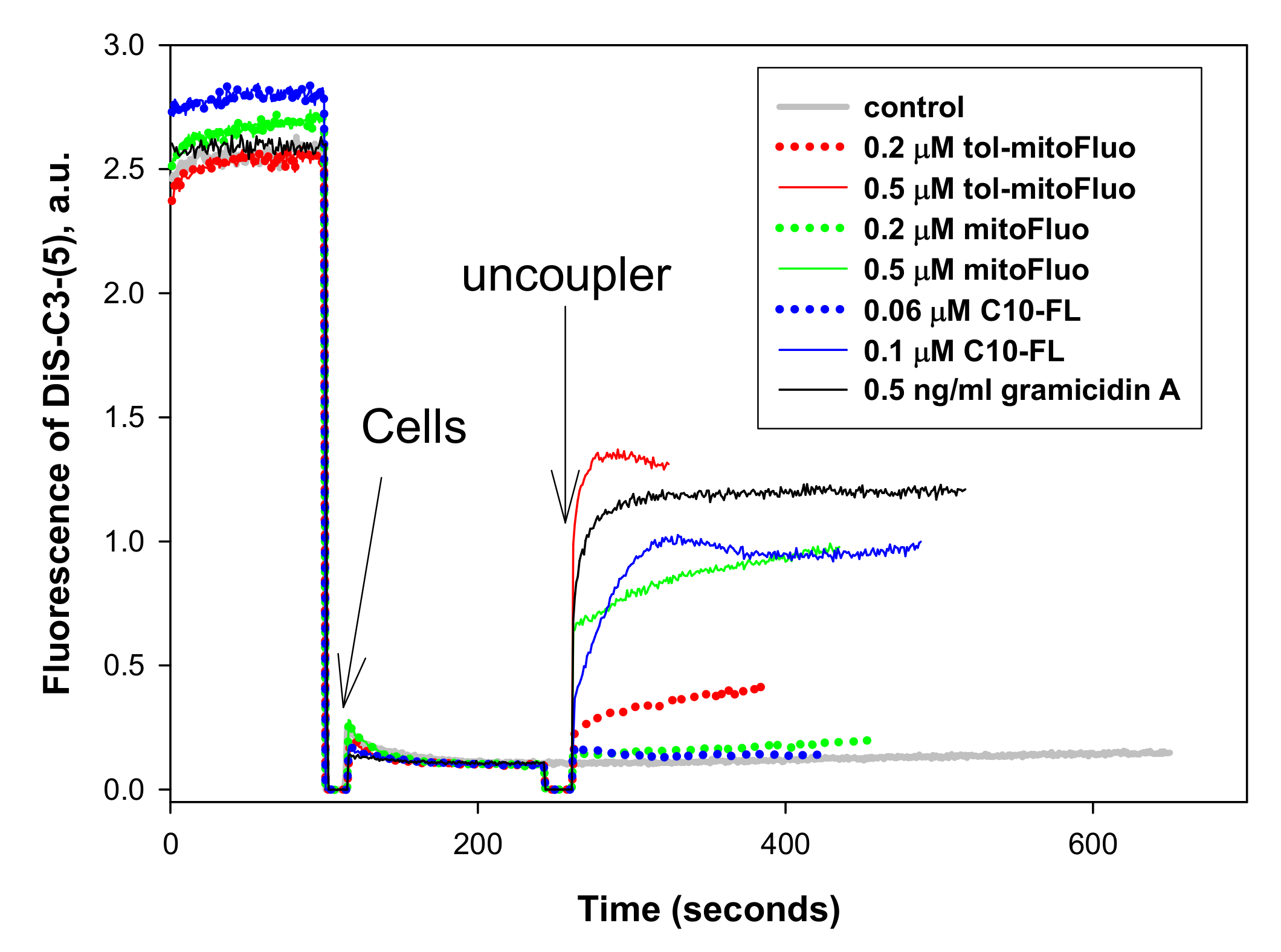
3. Results and Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Cunha, F.M.; Caldeira da Silva, C.C.; Cerqueira, F.M.; Kowaltowski, A.J. Mild mitochondrial uncoupling as a therapeutic strategy. Curr. Drug Targets 2011, 12, 783–789. [Google Scholar] [CrossRef] [PubMed]
- Perry, R.J.; Zhang, D.; Zhang, X.M.; Boyer, J.L.; Shulman, G.I. Controlled-release mitochondrial protonophore reverses diabetes and steatohepatitis in rats. Science 2015, 347, 1253–1256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ost, M.; Keipert, S.; Klaus, S. Targeted mitochondrial uncoupling beyond UCP1-the fine line between death and metabolic health. Biochimie 2017, 134, 77–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tam, J.; Hamza, T.; Ma, B.; Chen, K.; Beilhartz, G.L.; Ravel, J.; Feng, H.; Melnyk, R.A. Host-targeted niclosamide inhibits C. difficile virulence and prevents disease in mice without disrupting the gut microbiota. Nat. Commun. 2018, 9, 5233. [Google Scholar] [CrossRef] [PubMed]
- Childress, E.S.; Alexopoulos, S.J.; Hoehn, K.L.; Santos, W.L. Small molecule mitochondrial uncouplers and their therapeutic potential. J. Med. Chem. 2018, 61, 4641–4655. [Google Scholar] [CrossRef]
- Kumar, R.; Coronel, L.; Somalanka, B.; Raju, A.; Aning, O.A.; An, O.; Ho, Y.S.; Chen, S.; Mak, S.Y.; Hor, P.Y.; et al. Mitochondrial uncoupling reveals a novel therapeutic opportunity for p53-defective cancers. Nat. Commun. 2018, 9, 3931. [Google Scholar] [CrossRef] [Green Version]
- Geisler, J.G. 2,4 Dinitrophenol as medicine. Cells 2019, 8, E280. [Google Scholar] [CrossRef] [Green Version]
- Demine, S.; Renard, P.; Arnould, T. Mitochondrial Uncoupling: A Key Controller of Biological Processes in Physiology and Diseases. Cells 2019, 8, E795. [Google Scholar] [CrossRef] [Green Version]
- Kanemoto, N.; Okamoto, T.; Tanabe, K.; Shimada, T.; Minoshima, H.; Hidoh, Y.; Aoyama, M.; Ban, T.; Kobayashi, Y.; Ando, H.; et al. Antidiabetic and cardiovascular beneficial effects of a liver-localized mitochondrial uncoupler. Nat. Commun. 2019, 10, 2172. [Google Scholar] [CrossRef]
- Hurdle, J.G.; O’Neill, A.J.; Chopra, I.; Lee, R.E. Targeting bacterial membrane function: An underexploited mechanism for treating persistent infections. Nat. Rev. Microbiol. 2011, 9, 62–75. [Google Scholar] [CrossRef] [Green Version]
- Goldberg, K.; Sarig, H.; Zaknoon, F.; Epand, R.F.; Epand, R.M.; Mor, A. Sensitization of gram-negative bacteria by targeting the membrane potential. FASEB J. 2013, 27, 3818–3826. [Google Scholar] [CrossRef] [PubMed]
- Farha, M.A.; Verschoor, C.P.; Bowdish, D.; Brown, E.D. Collapsing the proton motive force to identify synergistic combinations against Staphylococcus aureus. Chem. Biol. 2013, 20, 1168–1178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, X.; Zhu, W.; Schurig-Briccio, L.A.; Lindert, S.; Shoen, C.; Hitchings, R.; Li, J.; Wang, Y.; Baig, N.; Zhou, T.; et al. Antiinfectives targeting enzymes and the proton motive force. Proc. Natl. Acad. Sci. USA 2015, 112, E7073–E7082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaneti, G.; Meir, O.; Mor, A. Controlling bacterial infections by inhibiting proton-dependent processes. Biochim. Biophys. Acta 2016, 1858, 995–1003. [Google Scholar] [CrossRef] [PubMed]
- Foss, M.H.; Pou, S.; Davidson, P.M.; Dunaj, J.L.; Winter, R.W.; Pou, S.; Licon, M.H.; Doh, J.K.; Li, Y.; Kelly, J.X.; et al. Diphenylether-Modified 1,2-Diamines with Improved Drug Properties for Development against Mycobacterium tuberculosis. ACS Infect. Dis. 2016, 2, 500–508. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Nyantakyi, S.A.; Gopal, P.; Aziz, D.B.; Dick, T.; Go, M.L. Indolylalkyltriphenylphosphonium Analogues Are Membrane-Depolarizing Mycobactericidal Agents. ACS Med. Chem. Lett. 2017, 8, 1165–1170. [Google Scholar] [CrossRef]
- Tharmalingam, N.; Port, J.; Castillo, D.; Mylonakis, E. Repurposing the anthelmintic drug niclosamide to combat Helicobacter pylori. Sci. Rep. 2018, 8, 3701. [Google Scholar] [CrossRef] [Green Version]
- Jeon, A.B.; Ackart, D.F.; Li, W.; Jackson, M.; Melander, R.J.; Melander, C.; Abramovitch, R.B.; Chicco, A.J.; Basaraba, R.J.; Obregyn-Henao, A. 2-Aminoimidazoles collapse mycobacterial proton motive force and block the electron transport chain. Sci. Rep. 2019, 9, 1513. [Google Scholar] [CrossRef]
- Valderrama, K.; Pradel, E.; Firsov, A.M.; Drobecq, H.; Bauderlique-le Roy, H.; Villemagne, B.; Antonenko, Y.N.; Hartkoorn, R.C. Pyrrolomycins Are Potent Natural Protonophores. Antimicrob. Agents Chemother. 2019, 63, e01450-19. [Google Scholar] [CrossRef]
- Harold, F.M. Antimicrobial Agents and Membrane Function. In Advances in Microbial Physiology, 1st ed.; Rose, A.H., Wilkinson, J.F., Eds.; Academic Press: Cambridge, MA, USA, 1969; Volume 4, pp. 45–104. [Google Scholar]
- Shchepinova, M.M.; Denisov, S.S.; Kotova, E.A.; Khailova, L.S.; Knorre, D.A.; Korshunova, G.A.; Tashlitsky, V.N.; Severin, F.F.; Antonenko, Y.N. Dodecyl and octyl esters of fluorescein as protonophores and uncouplers of oxidative phosphorylation in mitochondria at submicromolar concentrations. Biochim. Biophys. Acta 2014, 1837, 149–158. [Google Scholar] [CrossRef] [Green Version]
- Denisov, S.S.; Kotova, E.A.; Plotnikov, E.Y.; Tikhonov, A.A.; Zorov, D.B.; Korshunova, G.A.; Antonenko, Y.N. A mitochondria-targeted protonophoric uncoupler derived from fluorescein. Chem. Commun. 2014, 50, 15366–15369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antonenko, Y.N.; Denisov, S.S.; Silachev, D.N.; Khailova, L.S.; Jankauskas, S.S.; Rokitskaya, T.I.; Danilina, T.I.; Kotova, E.A.; Korshunova, G.A.; Plotnikov, E.Y.; et al. A long-linker conjugate of fluorescein and triphenylphosphonium as mitochondria-targeted uncoupler and fluorescent neuro- and nephroprotector. Biochim. Biophys. Acta 2016, 1860, 2463–2473. [Google Scholar] [CrossRef] [PubMed]
- Rokitskaya, T.I.; Luzhkov, V.B.; Korshunova, G.A.; Tashlitsky, V.N.; Antonenko, Y.N. Effect of methyl and halogen substituents on the transmembrane movement of lipophilic ions. Phys. Chem. Chem. Phys. 2019, 21, 23355–23363. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Sim, Y.; Kon, O.L.; Ng, W.H.; Ribeiro, A.J.; Ramos, M.J.; Fernandes, P.A.; Ganguly, R.; Xing, B.; Garcia, F.; et al. Unique Triphenylphosphonium Derivatives for Enhanced Mitochondrial Uptake and Photodynamic Therapy. Bioconjug. Chem. 2017, 28, 590–599. [Google Scholar] [CrossRef]
- Nikitina, E.V.; Zeldi, M.I.; Vafina, R.M.; Pugachev, M.V.; Shtyrlin, N.V.; Kuznetsova, S.V.; Evtugyn, V.G.; Kayumov, A.R.; Shtyrlin, Y.G. Elucidation of the mode of action of new antibacterial bis-phosphonium salts of pyridine derivatives active against Staphylococcus aureus. Res. J. Pharm. Biol. Chem. Sci. 2017, 8, 1906–1915. [Google Scholar]
- Rokitskaya, T.I.; Sumbatyan, N.V.; Tashlitsky, V.N.; Korshunova, G.A.; Antonenko, Y.N.; Skulachev, V.P. Mitochondria-targeted penetrating cations as carriers of hydrophobic anions through lipid membranes. Biochim. Biophys. Acta 2010, 1798, 1698–1706. [Google Scholar] [CrossRef] [Green Version]
- Johnson, D.; Lardy, H. Isolation of liver or kidney mitochondria. Methods Enzymol. 1967, 10, 94–96. [Google Scholar]
- Antonenko, Y.N.; Khailova, L.S.; Rokitskaya, T.I.; Nosikova, E.S.; Nazarov, P.A.; Luzina, O.A.; Salakhutdinov, N.F.; Kotova, E.A. Mechanism of action of an old antibiotic revisited: Role of calcium ions in protonophoric activity of usnic acid. Biochim. Biophys. Acta Bioenerg. 2019, 1860, 310–316. [Google Scholar] [CrossRef]
- Akerman, K.E.; Wikstrom, M.K. Safranine as a probe of the mitochondrial membrane potential. FEBS Lett. 1976, 68, 191–197. [Google Scholar] [CrossRef] [Green Version]
- Nazarov, P.A.; Osterman, I.A.; Tokarchuk, A.V.; Karakozova, M.V.; Korshunova, G.A.; Lyamzaev, K.G.; Skulachev, M.V.; Kotova, E.A.; Skulachev, V.P.; Antonenko, Y.N. Mitochondria-targeted antioxidants as highly effective antibiotics. Sci. Rep. 2017, 7, 1394. [Google Scholar] [CrossRef]
- Khailova, L.S.; Nazarov, P.A.; Sumbatyan, N.V.; Korshunova, G.A.; Rokitskaya, T.I.; Dedukhova, V.I.; Antonenko, Y.N.; Skulachev, V.P. Uncoupling and Toxic Action of Alkyltriphenylphosphonium Cations on Mitochondria and the Bacterium Bacillus subtilis as a Function of Alkyl Chain Length. Biochemistry 2015, 80, 1589–1597. [Google Scholar] [CrossRef] [PubMed]
- Antonenko, Y.N.; Denisov, S.S.; Khailova, L.S.; Nazarov, P.A.; Rokitskaya, T.I.; Tashlitsky, V.N.; Firsov, A.M.; Korshunova, G.A.; Kotova, E.A. Alkyl-substituted phenylamino derivatives of 7-nitrobenz-2-oxa-1,3-diazole as uncouplers of oxidative phosphorylation and antibacterial agents: Involvement of membrane proteins in the uncoupling action. Biochim. Biophys. Acta Biomembr. 2017, 1859, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Popova, L.B.; Nosikova, E.S.; Kotova, E.A.; Tarasova, E.O.; Nazarov, P.A.; Khailova, L.S.; Balezina, O.P.; Antonenko, Y.N. Protonophoric action of triclosan causes calcium efflux from mitochondria, plasma membrane depolarization and bursts of miniature end-plate potentials. Biochim. Biophys. Acta Biomembr. 2018, 1860, 1000–1007. [Google Scholar] [CrossRef] [PubMed]
- McAuley, S.; Huynh, A.; Czarny, T.L.; Brown, E.D.; Nodwell, J.R. Membrane activity profiling of small molecule B. subtilis growth inhibitors utilizing novel duel-dye fluorescence assay. MedChemComm 2018, 9, 554–561. [Google Scholar] [CrossRef] [PubMed]
- Young, J.W.; Locke, J.C.; Altinok, A.; Rosenfeld, N.; Bacarian, T.; Swain, P.S.; Mjolsness, E.; Elowitz, M.B. Measuring single-cell gene expression dynamics in bacteria using fluorescence time-lapse microscopy. Nat. Protoc. 2011, 7, 80–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Schindelin, J.; Rueden, C.T.; Hiner, M.C.; Eliceiri, K.W. The ImageJ ecosystem: An open platform for biomedical image analysis. Mol. Reprod. Dev. 2015, 82, 518–529. [Google Scholar] [CrossRef] [Green Version]
- Perevoshchikova, I.V.; Zorov, D.B.; Antonenko, Y.N. Peak intensity analysis as a method for estimation of fluorescent probe binding to artificial and natural nanoparticles: Tetramethylrhodamine uptake by isolated mitochondria. Biochim. Biophys. Acta 2008, 1778, 2182–2190. [Google Scholar] [CrossRef] [Green Version]
- Nazarov, P.A.; Kotova, E.A.; Skulachev, V.P.; Antonenko, Y.N. Genetic Variability of the AcrAB-TolC Multidrug Efflux Pump Underlies SkQ1 Resistance in Gram-Negative Bacteria. Acta Nat. 2019, 11, 93–98. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| mitoFluo MIC (µM) | tol-mitoFluo MIC (µM) | C10-FL MIC (µM) | |
|---|---|---|---|
| B. subtilis Br151 | 0.5 | 0.25 | 2 |
| B. subtilis PY79 | 0.5 | 0.25 | 2 |
| B. pumilus NCTC 8241 | 0.5 | 0.25 | 2 |
| S. aureus | 0.6 | 0.3 | 2.4 |
| E. coli W3110 | >15 | >15 | >15 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nazarov, P.A.; Kirsanov, R.S.; Denisov, S.S.; Khailova, L.S.; Karakozova, M.V.; Lyamzaev, K.G.; Korshunova, G.A.; Lukyanov, K.A.; Kotova, E.A.; Antonenko, Y.N. Fluorescein Derivatives as Antibacterial Agents Acting via Membrane Depolarization. Biomolecules 2020, 10, 309. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10020309
Nazarov PA, Kirsanov RS, Denisov SS, Khailova LS, Karakozova MV, Lyamzaev KG, Korshunova GA, Lukyanov KA, Kotova EA, Antonenko YN. Fluorescein Derivatives as Antibacterial Agents Acting via Membrane Depolarization. Biomolecules. 2020; 10(2):309. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10020309
Chicago/Turabian StyleNazarov, Pavel A., Roman S. Kirsanov, Stepan S. Denisov, Ljudmila S. Khailova, Marina V. Karakozova, Konstantin G. Lyamzaev, Galina A. Korshunova, Konstantin A. Lukyanov, Elena A. Kotova, and Yuri N. Antonenko. 2020. "Fluorescein Derivatives as Antibacterial Agents Acting via Membrane Depolarization" Biomolecules 10, no. 2: 309. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10020309