Site-Specific Lipidomic Signatures of Sea Lettuce (Ulva spp., Chlorophyta) Hold the Potential to Trace Their Geographic Origin

 ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
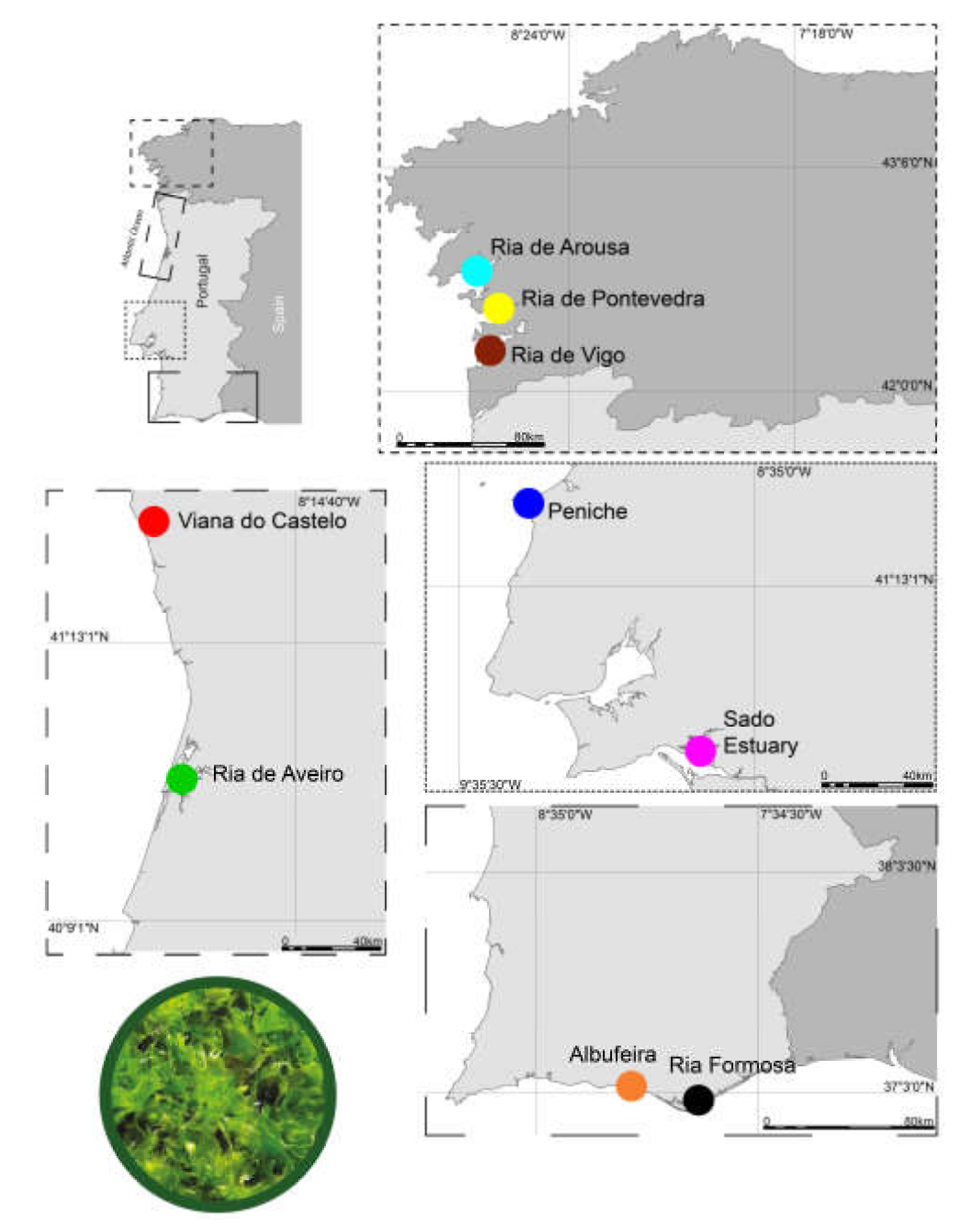
2.2. Sampling and Storage
2.3. Biochemical Composition
2.4. Lipid Extraction
2.5. Ultra High-Performance Liquid Chromatography-Mass Spectrometry
2.6. Data Analysis
3. Results
3.1. Biochemical Composition of Ulva spp
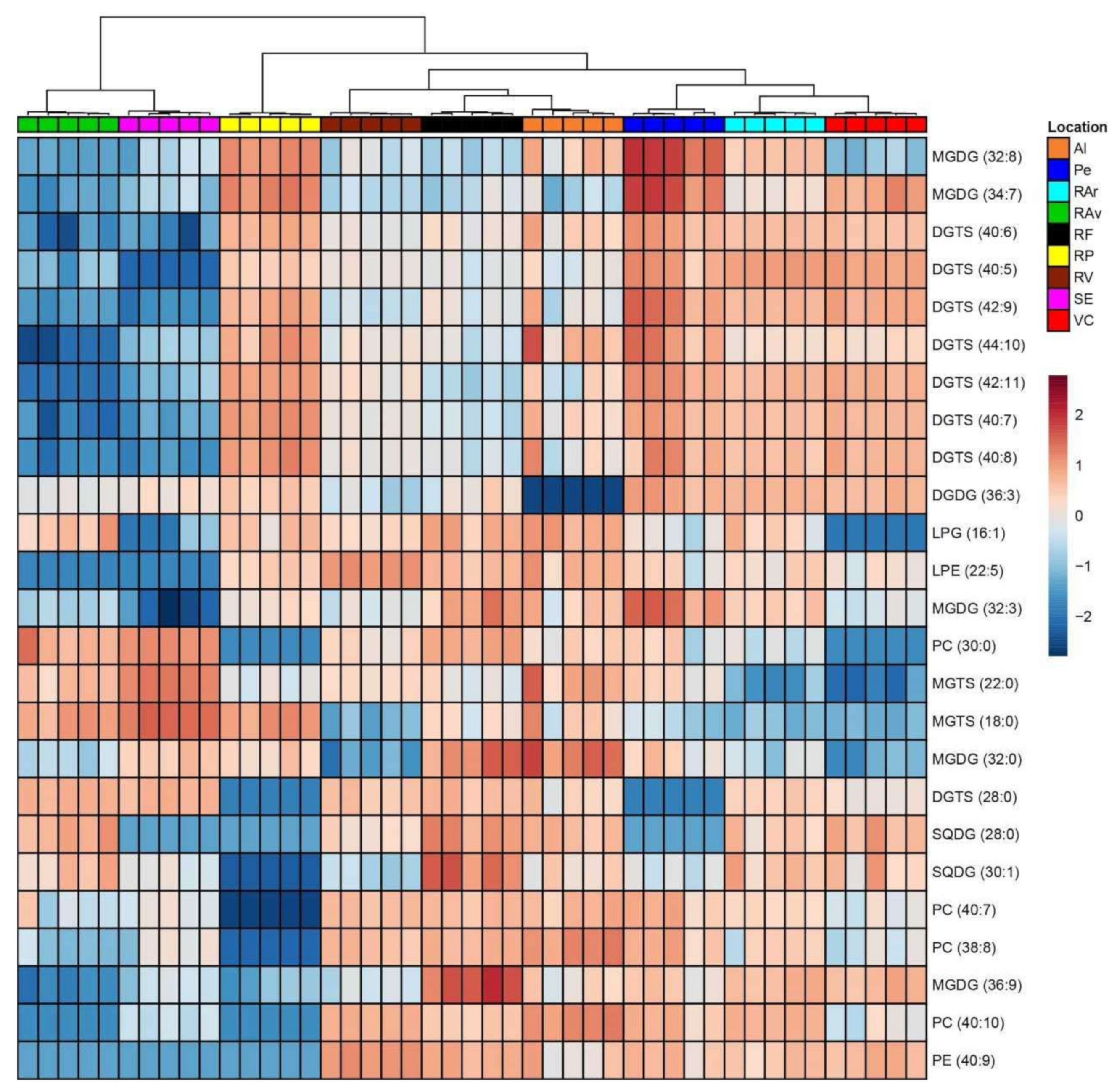
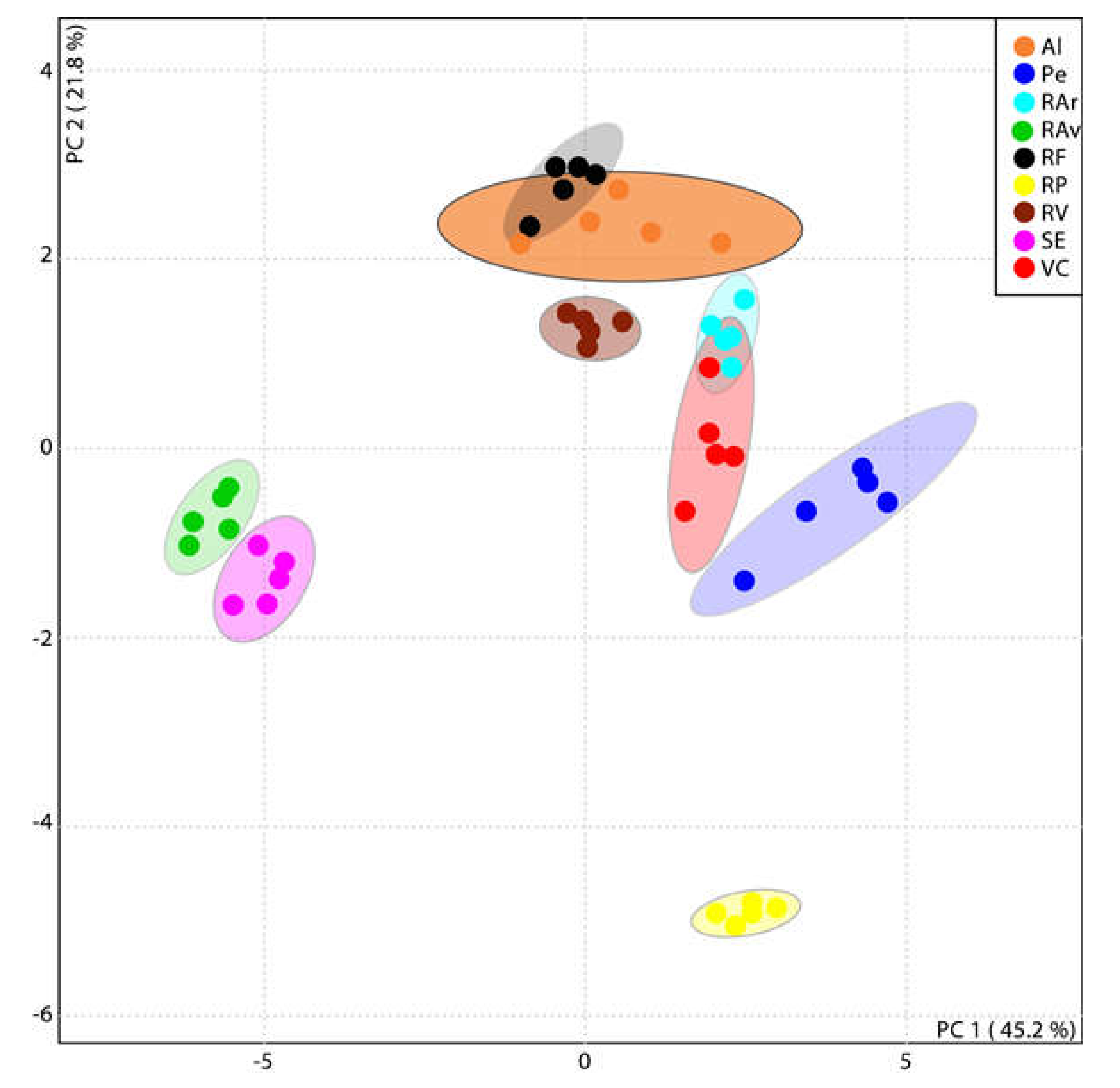
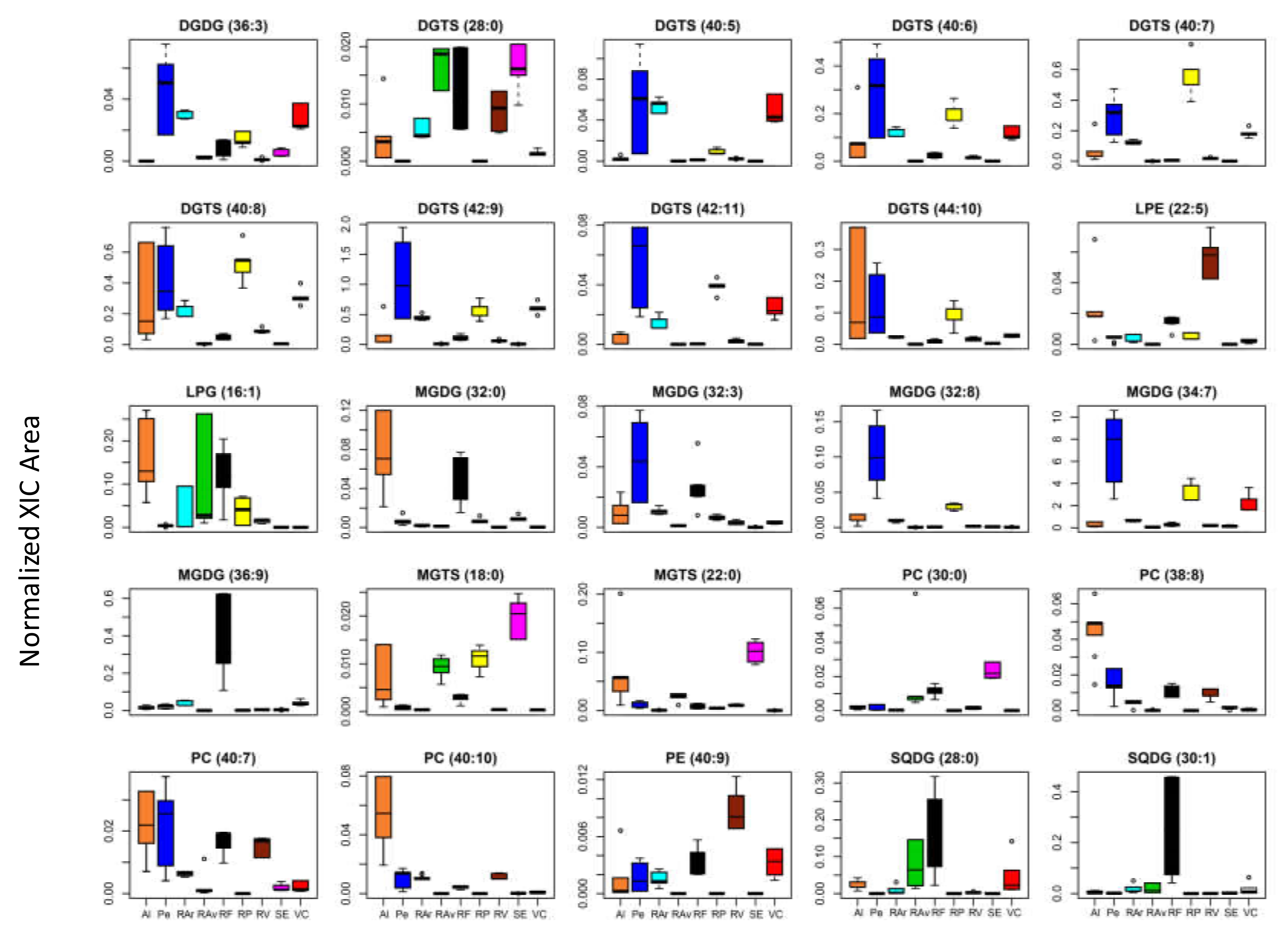
3.2. Lipidome Plasticity of Ulva spp. from Different Geographic Locations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Glossary
| Al | Albufeira |
| DGDG | Digalactosyl diacylglycerol |
| DGMG | Digalactosyl monoacylglycerol |
| DGTS | Diacylglyceryl-3-O-4-N,N,N-trimethyl-homoserine |
| DW | Dry Weight basis |
| HILIC | Hydrophilic interaction liquid chromatography |
| LPC | Lyso-phosphatidylcholine |
| LPE | Lyso-phosphatidylethanolamine |
| LPG | Lyso-phosphatidylglycerol |
| LPI | Lyso-phosphatidylinositol |
| m/z | Mass-to-charge ratio |
| MGDG | Monogalactosyl diacylglycerol |
| MGMG | Monogalactosyl monoacylglycerol |
| MGTS | Monocylglyceryl-3-O-4-(N,N,N-trimethyl) homoserine |
| MS | Mass spectrometry |
| MS/MS | Tandem mass spectrometry |
| PC | Phosphatidylcholine |
| Pe | Peniche |
| PE | Phosphatidylethanolamine |
| PG | Phosphatidylglycerol |
| PI | Phosphatidylinositol |
| Pi | Inorganic phosphate |
| PUFA | Polyunsaturated fatty acid |
| Rar | Ria Arousa |
| RAv | Ria de Aveiro |
| RF | Ria Formosa |
| RP | Ria de Pontevedra |
| RV | Ria de Vigo |
| SE | Sado Estuary |
| SFA | Saturated fatty acid |
| SQDG | Sulfoquinovosyl diacylglycerol |
| SQMG | Sulfoquinovosyl monoacylglycerol |
| VC | Viana do Castelo |
| TLs | Total lipids |
References
- Bolton, J.J.; Cyrus, M.D.; Brand, M.J.; Joubert, M.; Macey, B.M. Why grow Ulva? Its potential role in the future of aquaculture. Perspect. Phycol. 2016, 3, 113–120. [Google Scholar] [CrossRef]
- Kumari, P.; Kumar, M.; Reddy, C.R.K.R.K.; Jha, B. Nitrate and phosphate regimes induced lipidomic and biochemical changes in the intertidal macroalga Ulva lactuca (Ulvophyceae, Chlorophyta). Plant. Cell. Physiol. 2014, 55, 52–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dominguez, H.; Loret, E.P. Ulva lactuca, A source of troubles and potential riches. Mar. Drugs 2019, 1–20. [Google Scholar]
- Holdt, S.L.; Kraan, S. Bioactive compounds in seaweed: Functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–597. [Google Scholar] [CrossRef]
- Olsen, Y. How can mariculture better help feed humanity? Front. Mar. Sci. 2015, 2, 2009–2012. [Google Scholar] [CrossRef] [Green Version]
- Duarte, C.M.; Holmer, M.; Olsen, Y.; Soto, D.; Marbà, N.; Guiu, J.; Black, K.; Karakassis, I. Will the oceans help feed humanity? Bioscience 2009, 59, 967–976. [Google Scholar] [CrossRef]
- Leal, M.C.; Munro, M.H.G.G.; Blunt, J.W.; Puga, J.; Jesus, B.; Calado, R.; Rosa, R.; Madeira, C. Biogeography and biodiscovery hotspots of macroalgal marine natural products. Nat. Prod. Rep. 2013, 30, 1380–1390. [Google Scholar] [CrossRef]
- Kirkendale, L.; Saunders, G.W.; Winberg, P. A Molecular Survey of Ulva (Chlorophyta) in temperate Australia reveals enhanced levels of cosmopolitanism. J. Phycol. 2013, 49, 69–81. [Google Scholar] [CrossRef]
- Hayden, H.S.; Blomster, J.; Maggs, C.A.; Silva, P.C.; Stanhope, M.J.; Waaland, J.R. Linnaeus was right all along: Ulva and Enteromorpha are not distinct genera. Eur. J. Phycol. 2003, 38, 277–294. [Google Scholar] [CrossRef]
- Kraft, L.G.K.; Kraft, G.T.; Waller, R.F. Investigations into southern Australian Ulva (Ulvophyceae, Chlorophyta) taxonomy and molecular phylogeny indicate both cosmopolitanism and endemic cryptic species. J. Phycol. 2010, 46, 1257–1277. [Google Scholar] [CrossRef]
- Kostetsky, E.Y.; Goncharova, S.N.; Sanina, N.M.; Shnyrov, V.L. Season influence on lipid composition of marine macrophytes. Bot. Mar. 2004, 47, 134–139. [Google Scholar] [CrossRef]
- Castelar, B.; Reis, R.P.; dos Santos Calheiros, A.C. Ulva lactuca and U. flexuosa (Chlorophyta, Ulvophyceae) cultivation in Brazilian tropical waters: Recruitment, growth, and ulvan yield. J. Appl. Phycol. 2014, 26, 1989–1999. [Google Scholar] [CrossRef]
- Ghaderiardakani, F.; Califano, G.; Mohr, J.; Abreu, M.; Coates, J.; Wichard, T. Analysis of algal growth- and morphogenesis- promoting factors in an integrated multi-trophic aquaculture system for farming Ulva spp. Aquac. Environ. Interact. 2019, 11, 375–391. [Google Scholar] [CrossRef] [Green Version]
- Gadberry, B.A.; Colt, J.; Maynard, D.; Boratyn, D.C.; Webb, K.; Johnson, R.B.; Saunders, G.W.; Boyer, R.H. Intensive land-based production of red and green macroalgae for human consumption in the pacific northwest: An evaluation of seasonal growth, yield, nutritional composition, and contaminant levels. Algae 2018, 33, 109–125. [Google Scholar] [CrossRef] [Green Version]
- Fort, A.; Lebrault, M.; Allaire, M.; Esteves-Ferreira, A.A.; McHale, M.; Lopez, F.; Fariñas-Franco, J.M.; Alseekh, S.; Fernie, A.R.; Sulpice, R. Extensive variations in diurnal growth patterns and metabolism among Ulva spp stains. Plant. Phisiol. 2019, 180, 109–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leal, M.C.; Pimentel, T.; Ricardo, F.; Rosa, R.; Calado, R. Seafood traceability: Current needs, available tools, and biotechnological challenges for origin certification. Trends Biotechnol. 2015, 33, 331–336. [Google Scholar] [CrossRef]
- Galloway, A.W.E.; Britton-Simmons, K.H.; Duggins, D.O.; Gabrielson, P.W.; Brett, M.T. Fatty acid signatures differentiate marine macrophytes at ordinal and family ranks. J. Phycol. 2012, 48, 956–965. [Google Scholar] [CrossRef]
- Ricardo, F.; Pimentel, T.; Moreira, A.S.P.; Rey, F.; Coimbra, M.A.; Domingues, M.R.; Domingues, P.; Costa Leal, M.; Calado, R. Potential use of fatty acid profiles of the adductor muscle of cockles (Cerastoderma edule) for traceability of collection site. Sci. Rep. 2015, 5, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Floreto, E.A.T.; Teshima, S. The fatty acid composition of seaweed exposed to different levels of light intensity and salinity. Bot. Mar. 1998, 41, 467–481. [Google Scholar] [CrossRef]
- Cardoso, C.; Ripol, A.; Afonso, C.; Freire, M.; Varela, J.; Quental-Ferreira, H.; Pousão-Ferreira, P.; Bandarra, N. Fatty acid profiles of the main lipid classes of green seaweeds from fish pond aquaculture. Food Sci. Nutr. 2017, 5, 1186–1194. [Google Scholar] [CrossRef]
- Li-Beisson, Y.; Thelen, J.J.; Fedosejevs, E.; Harwood, J.L. The lipid biochemistry of eukaryotic algae. Prog. Lipid Res. 2019, 74, 31–68. [Google Scholar] [CrossRef] [PubMed]
- Schmid, M.; Guihéneuf, F.; Stengel, D.B. Plasticity and remodeling of lipids support acclimation potential in two species of low-intertidal macroalgae, Fucus serratus (Phaeophyceae) and Palmaria palmata (Rhodophyta). Algal Res. 2017, 26, 104–114. [Google Scholar] [CrossRef]
- Mæhre, H.K.; Malde, M.K.; Eilertsen, K.E.; Elvevoll, E.O. Characterization of protein, lipid and mineral contents in common Norwegian seaweeds and evaluation of their potential as food and feed. J. Sci. Food Agric. 2014, 94, 3281–3290. [Google Scholar] [CrossRef] [PubMed]
- Lopes, D.; Moreira, A.S.P.; Rey, F.; da Costa, E.; Melo, T.; Maciel, E.; Rego, A.; Abreu, M.H.; Domingues, P.; Calado, R.; et al. Lipidomic signature of the green macroalgae Ulva rigida farmed in a sustainable integrated multi-trophic aquaculture. J. Appl. Phycol. 2019, 31, 1369–1381. [Google Scholar] [CrossRef] [Green Version]
- Sanina, N.M.; Goncharova, S.N.; Kostetsky, E.Y. Fatty acid composition of individual polar lipid classes from marine macrophytes. Phytochemistry 2004, 65, 721–730. [Google Scholar] [CrossRef] [PubMed]
- Kendel, M.; Wielgosz-collin, G.; Bertrand, S.; Roussakis, C.; Bourgougnon, N.; Bedoux, G. Lipid Composition, fatty acids and sterols in the seaweeds Ulva armoricana, and Solieria chordalis from Brittany (France): An analysis from nutritional, chemotaxonomic, and antiproliferative activity perspectives. Mar. Drugs 2015, 5606–5628. [Google Scholar] [CrossRef] [PubMed]
- Harwood, J.L.; Guschina, I.A. The versatility of algae and their lipid metabolism. Biochimie 2009, 91, 679–684. [Google Scholar] [CrossRef]
- Murata, N.; Los, D.A. Membrane fluidity and temperature perception. Plant Physiol. 1997, 115, 875–879. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.C.; Sinha, R.P.; Häder, D. Role of lipids and fatty acids in stress tolerance in cyanobacteria. Acta Protozool. 2002, 41, 297–308. [Google Scholar]
- Lordan, S.; Ross, R.P.; Stanton, C. Marine bioactives as functional food ingredients: Potential to reduce the incidence of chronic diseases. Mar. Drugs 2011, 9, 1056–1100. [Google Scholar] [CrossRef] [Green Version]
- Plouguerné, E.; da Gama, B.A.P.; Pereira, R.C.; Barreto-Bergter, E. Glycolipids from seaweeds and their potential biotechnological applications. Front. Cell. Infect. Microbiol. 2014, 4, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Maciel, E.; Leal, M.C.; Lillebø, A.I.; Domingues, P.; Domingues, M.R.; Calado, R. Bioprospecting of marine macrophytes using MS-based lipidomics as a new approach. Mar. Drugs 2016, 14, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Costa, E.; Silva, J.; Mendonça, S.H.S.; Abreu, M.H.M.H.; Domingues, M.R.; Mendonça, S.H.; Abreu, M.H.M.H.; Domingues, M.R. Lipidomic approaches towards deciphering glycolipids from microalgae as a reservoir of bioactive lipids. Mar. Drugs 2016, 14, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinhagen, S.; Karez, R.; Weinberger, F. Cryptic, alien and lost species: Molecular diversity of Ulva sensu lato along the German coasts of the North and Baltic Seas. Eur. J. Phycol. 2019, 54, 466–483. [Google Scholar] [CrossRef] [Green Version]
- Da Costa, E.; Domingues, P.; Melo, T.; Coelho, E.; Pereira, R.; Calado, R.; Abreu, M.H.; Domingues, M.R. Lipidomic signatures reveal seasonal shifts on the relative abundance of high-valued lipids from the brown algae Fucus vesiculosus. Mar. Drugs 2019, 17, 335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef] [Green Version]
- R Core Team R. A language and environment for statistical computing, The R Project for Statistical. Computing. 2014. Available online: http://www.R-project.org/ (accessed on 13 November 2019).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2009. [Google Scholar]
- Guschina, I.A.; Harwood, J.L. Lipids and lipid metabolism in eukaryotic algae. Prog. Lipid Res. 2006, 45, 160–186. [Google Scholar] [CrossRef]
- Thompson, G.A. Lipids and membrane function in green algae. Biochim. Biophys. Acta. 1996, 1302, 17–45. [Google Scholar] [CrossRef]
- Niemelä, P.S.; Castillo, S.; Sysi-aho, M.; Orě, M. Bioinformatics and computational methods for lipidomics. J. Chromatogr. B 2009, 877, 2855–2862. [Google Scholar]
- Pradas, I.; Huynh, K.; Cabré, R.; Ayala, V.; Meikle, P.J.; Jové, M.; Pamplona, R. Lipidomics reveals a tissue-specific fingerprint. Front. Physiol. 2018, 9, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Kumari, P.; Bijo, A.J.; Mantri, V.A.; Reddy, C.R.K.; Jha, B. Fatty acid profiling of tropical marine macroalgae: An analysis from chemotaxonomic and nutritional perspectives. Phytochemistry 2013, 86, 44–56. [Google Scholar] [CrossRef] [PubMed]
- Nelson, M.M.; Phleger, C.F.; Nichols, P.D. Seasonal lipid composition in macroalgae of the Northeastern Pacific Ocean. Bot. Mar. 2002, 45, 58–65. [Google Scholar] [CrossRef]
- Li, Y.H.; Wang, D.; Xu, X.T.; Gao, X.X.; Sun, X.; Xu, N.J. Physiological responses of a green algae (Ulva prolifera) exposed to simulated acid rain and decreased salinity. Photosynthetica 2017, 55, 623–629. [Google Scholar] [CrossRef]
- Rozentsvet, O.A.; Nesterov, V.N. Lipids and fatty acids from Ulva intestinalis from estuaries of the Caspian Basin (Elton region). Chem. Nat. Compd. 2012, 48, 544–547. [Google Scholar] [CrossRef]
- Kumari, P.; Kumar, M.; Reddy, C.R.; Jha, B. Algal lipids, fatty acids and sterols. In Functional Ingredients from Algae for Foods and Nutraceuticals; Domínguez, H., Ed.; Woodhead Publishing Limited: Cambridge, UK, 2013; pp. 119–166. ISBN 978-0-85709-868-9. [Google Scholar]
- Kostetsky, E.; Chopenko, N.; Barkina, M.; Velansky, P.; Sanina, N. Fatty acid composition and thermotropic behavior of glycolipids and other membrane lipids of Ulva lactuca (Chlorophyta) inhabiting different climatic zones. Mar. Drugs 2018, 16, 494. [Google Scholar] [CrossRef] [Green Version]
- Van Ginneken, V.; Gittenberger, A.; Rensing, M.; De Vries, E.; Peeters, E.T.H.M.; Verheij, E. Seaweed competition: Ulva sp. has the potential to produce the betaine lipid diacylglyceryl-O-4’-(N, N, N, -Trimethyl) homoserine (DGTS) in order to replace phosphatidylcholine (PC) under phosphate-limiting conditions in the P-limited Dutch Wadden Sea and outcompete an aggressive non-indigenous Gracilaria vermiculophylla red drift algae out of this unique Unesco World Heritage coastal area. Fish. Oceanogr. 2017, 2, 555596. [Google Scholar]
- Gasulla, F.; Vom Dorp, K.; Dombrink, I.; Zähringer, U.; Gisch, N.; Dörmann, P.; Bartels, D. The role of lipid metabolism in the acquisition of desiccation tolerance in Craterostigma plantagineum: A comparative approach. Plant J. 2013, 75, 726–741. [Google Scholar] [CrossRef]
- Vu, H.S.; Shiva, S.; Roth, M.R.; Tamura, P.; Zheng, L.; Li, M.; Sarowar, S.; Honey, S.; McEllhiney, D.; Hinkes, P.; et al. Lipid changes after leaf wounding in Arabidopsis thaliana: Expanded lipidomic data form the basis for lipid co-occurrence analysis. Plant J. 2014, 80, 728–743. [Google Scholar] [CrossRef]
- Villares, R.; Carballeira, A. Seasonal variation in the concentrations of nutrients in two green macroalgae and nutrient levels in sediments in the Rías Baixas (NW Spain). Estuar. Coast. Shelf Sci. 2003, 58, 887–900. [Google Scholar] [CrossRef]
- Cañavate, J.P.; Armada, I.; R’ios, J.L.; Hachero-Cruzado, I.; Canãvate, J.P.; Armada, I.; Riós, J.L.; Hachero-Cruzado, I. Exploring occurrence and molecular diversity of betaine lipids across taxonomy of marine microalgae. Phytochemistry 2016, 124, 68–78. [Google Scholar] [CrossRef]
- Murakami, H.; Nobusawa, T.; Hori, K.; Shimojima, M.; Ohta, H. Betaine lipid is crucial for adapting to low temperature and phosphate deficiency in Nannochloropsis. Plant Physiol. 2018, 177, 181–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okazaki, Y.; Otsuki, H.; Narisawa, T.; Kobayashi, M.; Sawai, S.; Kamide, Y.; Kusano, M.; Aoki, T.; Hirai, M.Y.; Saito, K. A new class of plant lipid is essential for protection against phosphorus depletion. Nat. Commun. 2013, 4, 1510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, N. Betaine Lipids. Bot. Mag. Tokyo 1992, 1, 185–197. [Google Scholar] [CrossRef]
- Niell, F.X.; Fernández, C.; Figueroa, F.L.; Figueiras, F.G.; Fuentes, J.M.; Pérez-Llorens, J.L.; Garcia-Sánchez, M.J.; Hernández, I.; Fernández, J.A.; Espejo, M.; et al. Spanish Atlantic coast. In Marine Benthic Vegetation; Recent Changes and The Effects of Eutrophication; Schram, W., Nienhuis, P.H., Eds.; Springer: Berlin, Germany, 1996; pp. 265–282. [Google Scholar]
- Fort, A.; Mannion, C.; Fariñas-Franco, J.M.; Sulpice, R. Green tides select for fast expanding Ulva strains. Sci. Total Environ. 2020, 698, 134337. [Google Scholar] [CrossRef]
- Dalsgaard, J.; St. John, M.; Kattner, G.; Müller-Navarra, D.; Hagen, W. Fatty acid trophic markers in pelagic marine environment. Adv. Mar. Biol. 2003, 46, 225–340. [Google Scholar]
- Hurd, C.; Harrison, P.; Bischof, K.; Lobban, C. Physico-chemical factors as environmental stressors in seaweed biology. In Seaweed Ecology and Physiology; Cambridge University Press: Cambridge, UK, 2014; pp. 294–348. [Google Scholar]
- Cowan, A.K. Phospholipids as plant growth regulators. Plant Growth Regul. 2006, 48, 97–109. [Google Scholar] [CrossRef]
- Carvalho, S.; Ravara, A.; Quintino, V.; Rodrigues, A.M. Macrobenthic community characterisation of an estuary from the western coast of Portugal (Sado estuary) prior to dredging operations. Bol. Inst. Esp. Oceanogr. 2001, 17, 179–190. [Google Scholar]
- Lopes, C.B.; Lillebø, A.I.; Dias, J.M.; Pereira, E.; Vale, C.; Duarte, A.C. Nutrient dynamics and seasonal succession of phytoplankton assemblages in a Southern European Estuary: Ria de Aveiro, Portugal. Estuar. Coast. Shelf Sci. 2007, 71, 480–490. [Google Scholar] [CrossRef]
- Vidal, T.; Calado, A.J.; Moita, M.T.; Cunha, M.R. Phytoplankton dynamics in relation to seasonal variability and upwelling and relaxation patterns at the mouth of Ria de Aveiro (West Iberian Margin) over a four-year period. PLoS ONE 2017, 12, e0177237. [Google Scholar] [CrossRef]
- Guo, Q.; Liu, L.; Barkla, B.J. Membrane lipid remodeling in response to salinity. Int. J. Mol. Sci. 2019, 20, 4264. [Google Scholar] [CrossRef] [Green Version]
- Kumar, M.; Kumari, P.; Reddy, C.R.K.; Jha, B. Chapter Four-Salinity and desiccation induced oxidative stress acclimation in seaweeds. Adv. Bot. Res. 2014, 71, 91–123. [Google Scholar]
- Xu, X.Q.; Beardall, J. Effect of salinity on fatty acid composition of a green microalga from an antarctic hypersaline lake. Phytochemistry 1997, 45, 655–658. [Google Scholar] [CrossRef]
- Stigter, T.; Dill, A.; Malta, E.; Santos, R. Nutrient sources for green macroalgae in the Ria Formosa lagoon—Assessing the role of groundwater. In Hydrogeology—Groundwater and Ecosystems; Taylor & Francis: Abingdon, UK, 2013; pp. 153–167. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Al | Pe | RAr | RAv | RF | RP | RV | SE | VC | |
|---|---|---|---|---|---|---|---|---|---|
| DGMG (16:1) | D | ND | D | D | D | ND | D | D | ND |
| DGMG (16:2) | ND | ND | ND | ND | ND | ND | D | ND | ND |
| DGMG (16:3) | ND | ND | ND | ND | ND | ND | D | ND | ND |
| DGMG (16:4) | ND | ND | ND | D | ND | ND | ND | ND | ND |
| DGDG (30:1) | ND | ND | D | ND | D | ND | ND | D | D |
| DGDG (36:3) | ND | D | D | D | D | D | D | D | D |
| DGDG (38:9) | D | ND | D | D | D | ND | D | D | D |
| SQDG (28:0) | D | ND | D | D | D | ND | D | ND | D |
| SQDG (30:1) | D | D | D | D | D | ND | D | D | D |
| DGTS (28:0) | D | ND | D | D | D | ND | D | D | D |
| DGTS (40:5) | D | D | D | D | D | D | D | ND | D |
| DGTS (42:11) | D | D | D | ND | D | D | D | D | D |
| MGTS (20:0) | D | D | D | D | D | D | ND | D | D |
| PC (30:0) | D | D | D | D | D | ND | D | D | ND |
| PC (34:4) | D | D | D | D | D | ND | D | D | D |
| PC (38:8) | D | D | D | D | D | ND | D | D | D |
| PC (40:10) | D | D | D | ND | D | ND | D | D | D |
| PC (40:7) | D | D | D | D | D | ND | D | D | D |
| LPE (22:5) | D | D | D | ND | D | D | D | ND | D |
| PE (34:5) | D | D | ND | D | D | D | D | D | ND |
| PE (36:6) | D | D | ND | ND | D | ND | D | D | ND |
| PE (40:9) | D | D | D | ND | D | ND | D | ND | D |
| LPG (16:1) | D | D | D | D | D | D | D | D | ND |
| m/z | Lipid Species (C:N) | Fatty Acyl Chain | Formula |
|---|---|---|---|
| [M + NH4]+ | |||
| 732.4677 | MGDG (32:8) | 16:4-16:4 | C41H66NO10 |
| 742.5443 | MGDG (32:3) | 16:3-16:0 and 18:3-14:0 | C41H76NO10 |
| 748.5919 | MGDG (32:0) | 16.0-16:0 | C41H82NO10 |
| 762.5133 | MGDG (34:7) | 18:3-16:4 | C43H72NO10 |
| 786.5145 | MGDG (36:9) | 20:5-16:4 | C45H72NO10 |
| 960.6582 | DGDG (36:3) * | 18:2-18:1 | C51H94O15N |
| [M + H]+ | |||
| 656.5451 | DGTS (28:0) * | 14:0-14:0 and 16:0-12:0 | C38H74O7N |
| 808.6058 | DGTS (40:8) | 22:5-18:3 | C50H82O7N |
| 810.624 | DGTS (40:7) * | 22:5-18:2 | C50H84O7N |
| 812.6424 | DGTS (40:6) | 22:5-18:1 | C50H86O7N |
| 814.6572 | DGTS (40:5) * | 22:1-18:4 | C50H88O7N |
| 830.5900 | DGTS (42:11) * | b) | C52H80O7N |
| 834.6232 | DGTS (42:9) | 22:5-20:4 | C52H84O7N |
| 860.6368 | DGTS (44:10) | 22:5-22:5 | C54H86O7N |
| 502.4104 | MGTS (18:0) | 18:0 | C28H56O6N |
| 558.4716 | MGTS (22:0) | 22:0 | C32H64O6N |
| 706.5372 | PC (30:0) * | a) | C38H77NO8P |
| 802.53668 | PC (38:8) | b) | C46H77NO8P |
| 826.5352 | PC (40:10) * | b) | C48H77NO8P |
| 832.5826 | PC (40:7) | a) | C48H83NO8P |
| 528.3084 | LPE (22:5) * | 22:5 | C27H47NO7P |
| 786.5055 | PE (40:9) * | a) | C45H73O8NP |
| [M − H]− | |||
| 737.4507 | SQDG (28:0) * | 14:0-14:0 and 12:0-16:0 | C37H69O12S |
| 763.466 | SQDG (30:1) * | 16:1-14:0 | C39H71O12S |
| 481.2569 | LPG (16:1) * | 16:1 | C22H42O9P |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
da Costa, E.; Ricardo, F.; Melo, T.; Mamede, R.; Abreu, M.H.; Domingues, P.; Domingues, M.R.; Calado, R. Site-Specific Lipidomic Signatures of Sea Lettuce (Ulva spp., Chlorophyta) Hold the Potential to Trace Their Geographic Origin. Biomolecules 2020, 10, 489. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10030489
da Costa E, Ricardo F, Melo T, Mamede R, Abreu MH, Domingues P, Domingues MR, Calado R. Site-Specific Lipidomic Signatures of Sea Lettuce (Ulva spp., Chlorophyta) Hold the Potential to Trace Their Geographic Origin. Biomolecules. 2020; 10(3):489. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10030489
Chicago/Turabian Styleda Costa, Elisabete, Fernando Ricardo, Tânia Melo, Renato Mamede, Maria H. Abreu, Pedro Domingues, M. Rosário Domingues, and Ricardo Calado. 2020. "Site-Specific Lipidomic Signatures of Sea Lettuce (Ulva spp., Chlorophyta) Hold the Potential to Trace Their Geographic Origin" Biomolecules 10, no. 3: 489. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10030489