Profiling and Role of Bioactive Molecules from Puntius sophore (Freshwater/Brackish Fish) Skin Mucus with Its Potent Antibacterial, Antiadhesion, and Antibiofilm Activities

 , ,
, ,  ,
,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Strains, Materials, and Growth Conditions
2.3. Collection and Maintenance of Fish
2.4. Collection of Fish Mucus
2.5. Antibacterial Activity
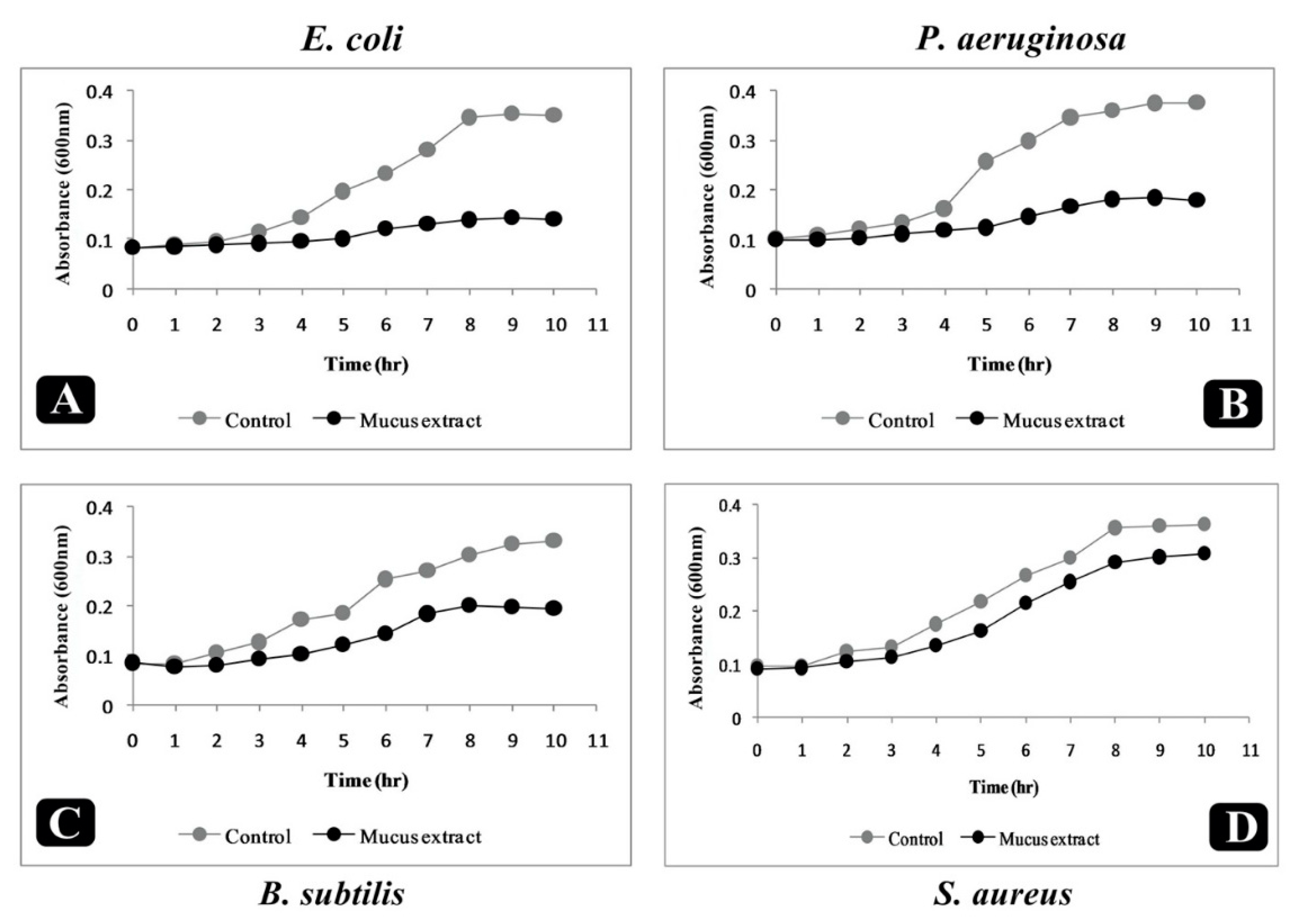
2.6. Effect of Puntius sophore Mucus Extract on Growth Kinetics of Bacteria
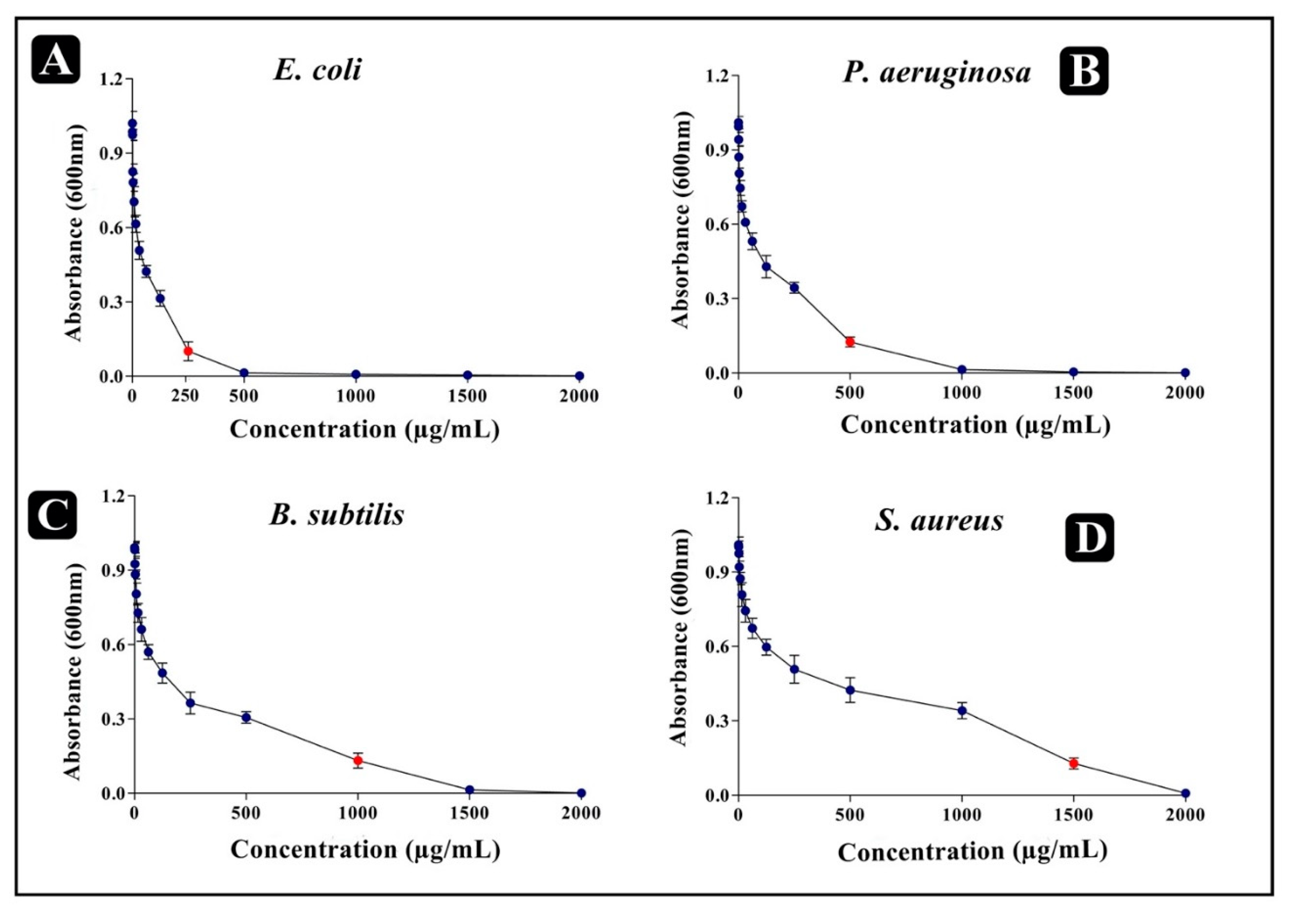
2.7. Determination of Minimum Inhibitory Concentration (MIC) by Serial Dilution Assay
2.8. Determination of Minimum Bactericidal Concentration (MBC)
2.9. Determination of Fractional Inhibitory Concentration Index (FICI)
2.10. Biofilm Assay
2.11. Assessment on Established Biofilms
2.12. Assessment on Adherence of Biofilms
2.13. Assessment of Antibiofilm Activity by Light Microscopy (LM)
2.14. Assessment of Antibiofilm Activity by Fluorescence Microscopy (FM)
2.15. Assessment of Antibiofilm Activity by Scanning Electron Microscopy (SEM)
2.16. Biofilm Metabolic Activity—XTT Reduction Assay
2.17. Cell Damage Assay
2.18. Extracellular Polysaccharide (EPS) Production Assay
2.19. Cytotoxicity Assay
2.20. Identification and Analysis of Bioactive Metabolites by High Resolution-Liquid Chromatography Mass Spectroscopy (HR-LCMS)
2.21. Statistical Analysis
3. Results
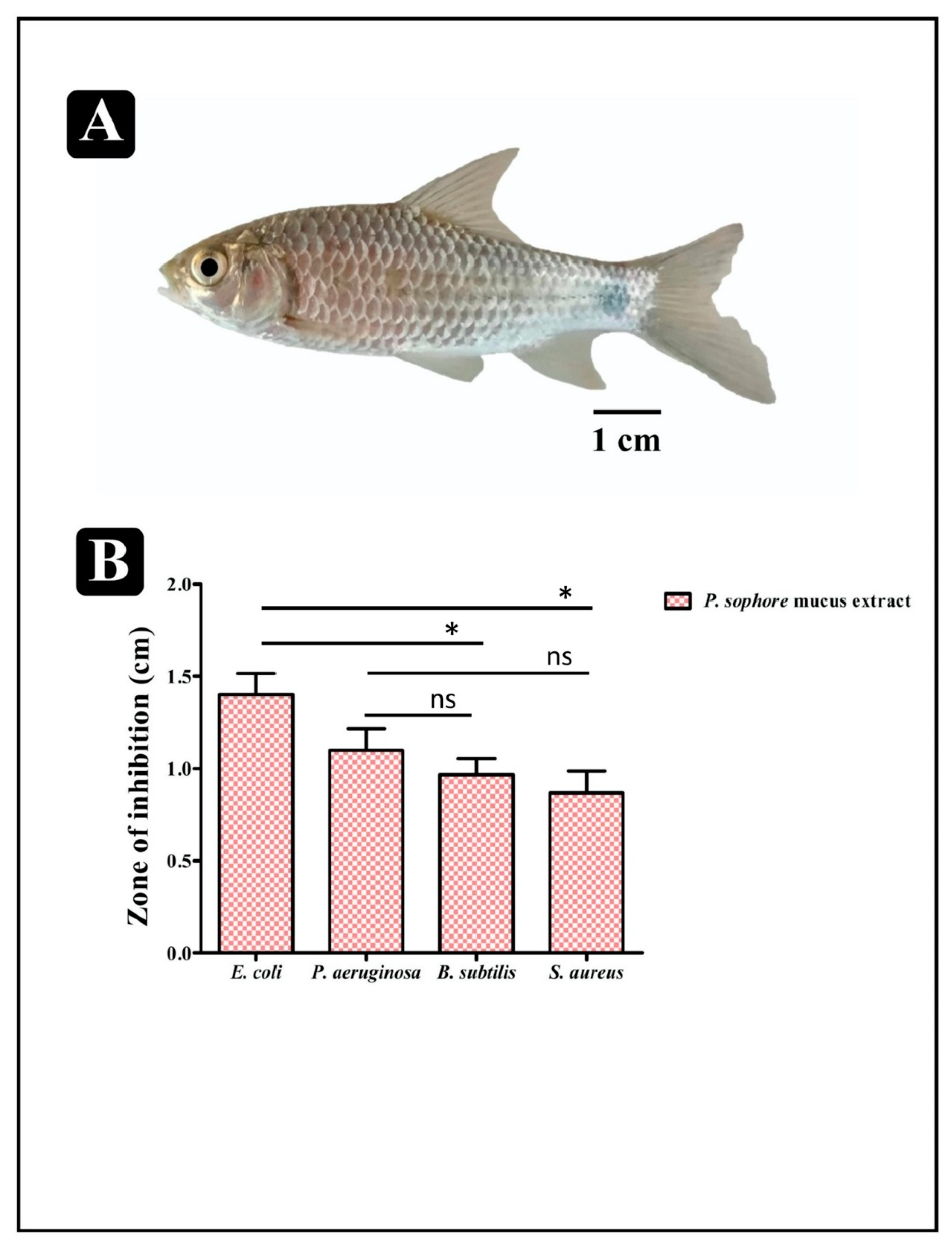
3.1. Antibacterial Susceptibility Profile of Puntius sophore Mucus Extract
3.2. Bacterial Killing Assay
3.3. Checkerboard Test
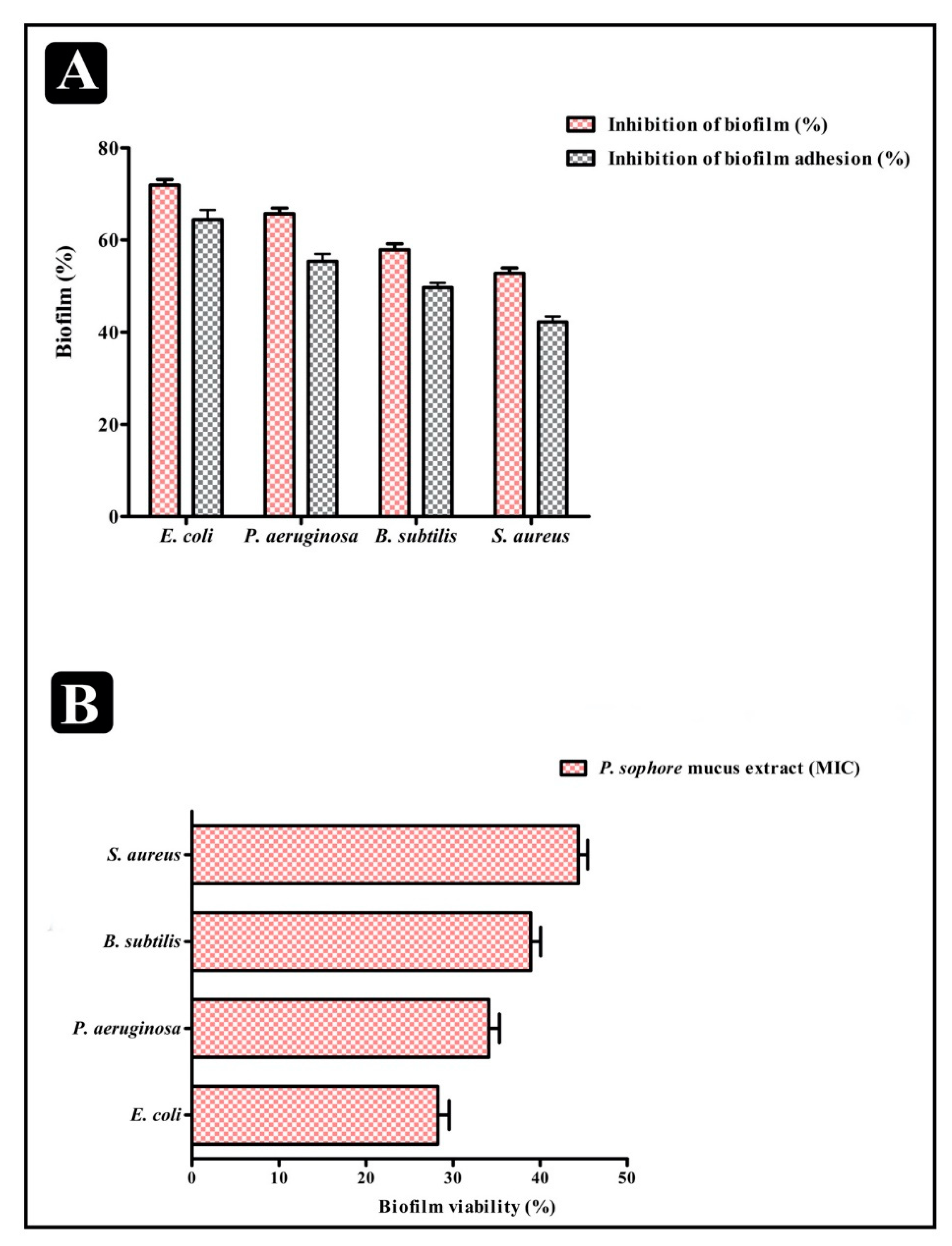
3.4. Antibiofilm Properties of P. sophore Mucus Extract
3.5. Effect of Puntius sophore Mucus Extract on Biofilms
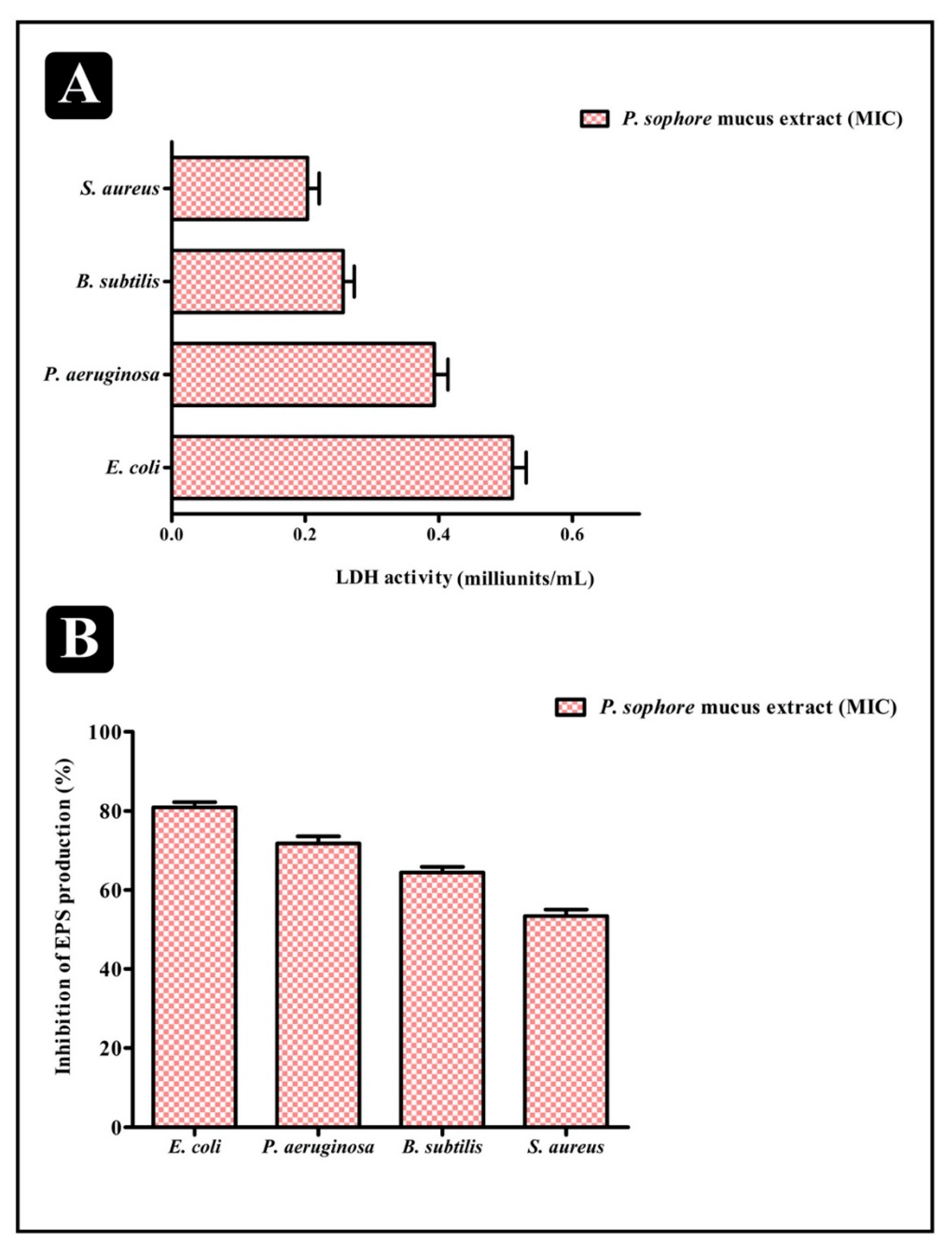
3.6. Effect of Puntius sophore Mucus Extract on Bacterial Cells Entrapped in Biofilms
3.7. Extracellular Polysaccharide (EPS) Production
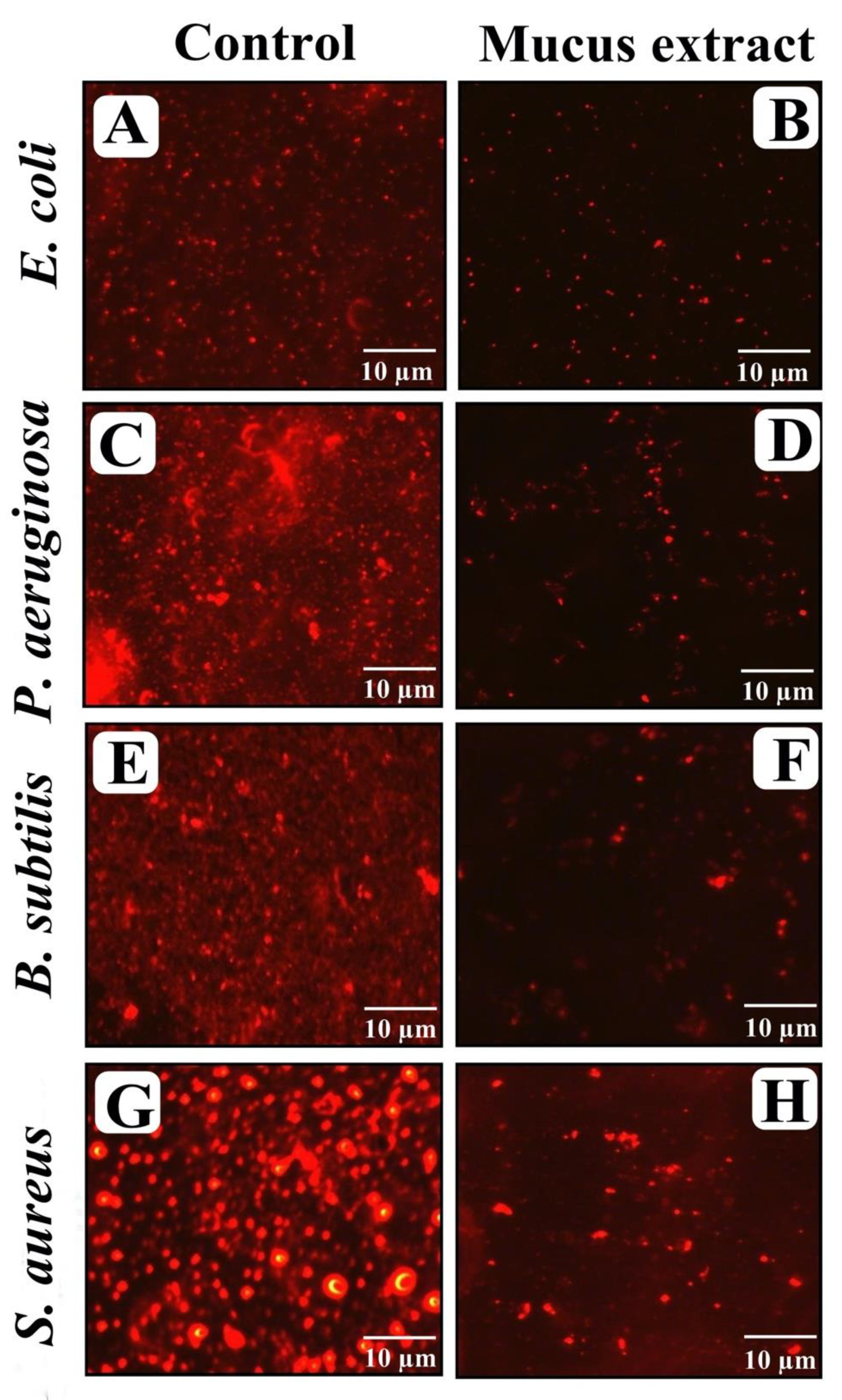
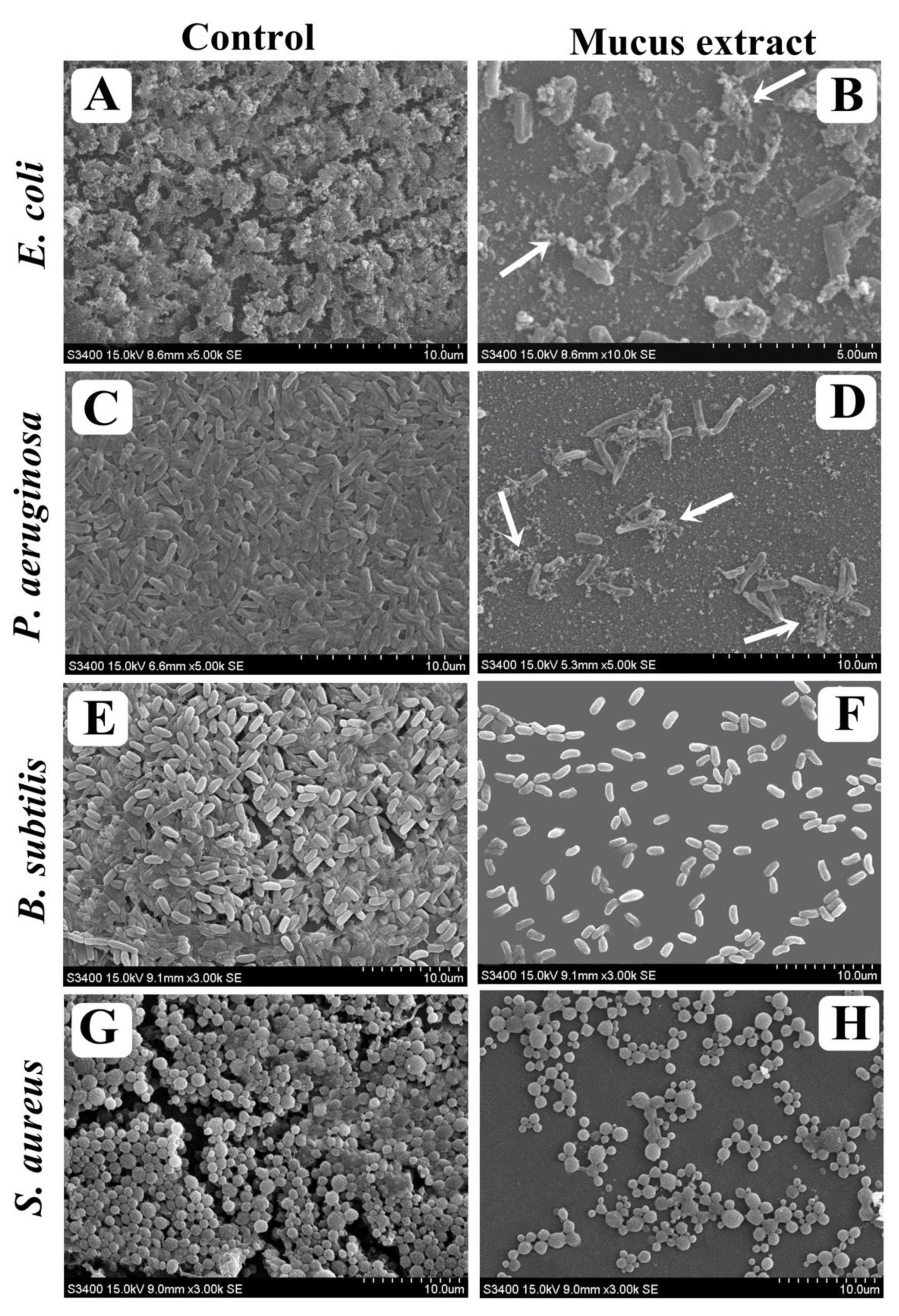
3.8. Visualization of Disrupted Biofilms by Microscopic Analysis (LM, FM, and SEM)
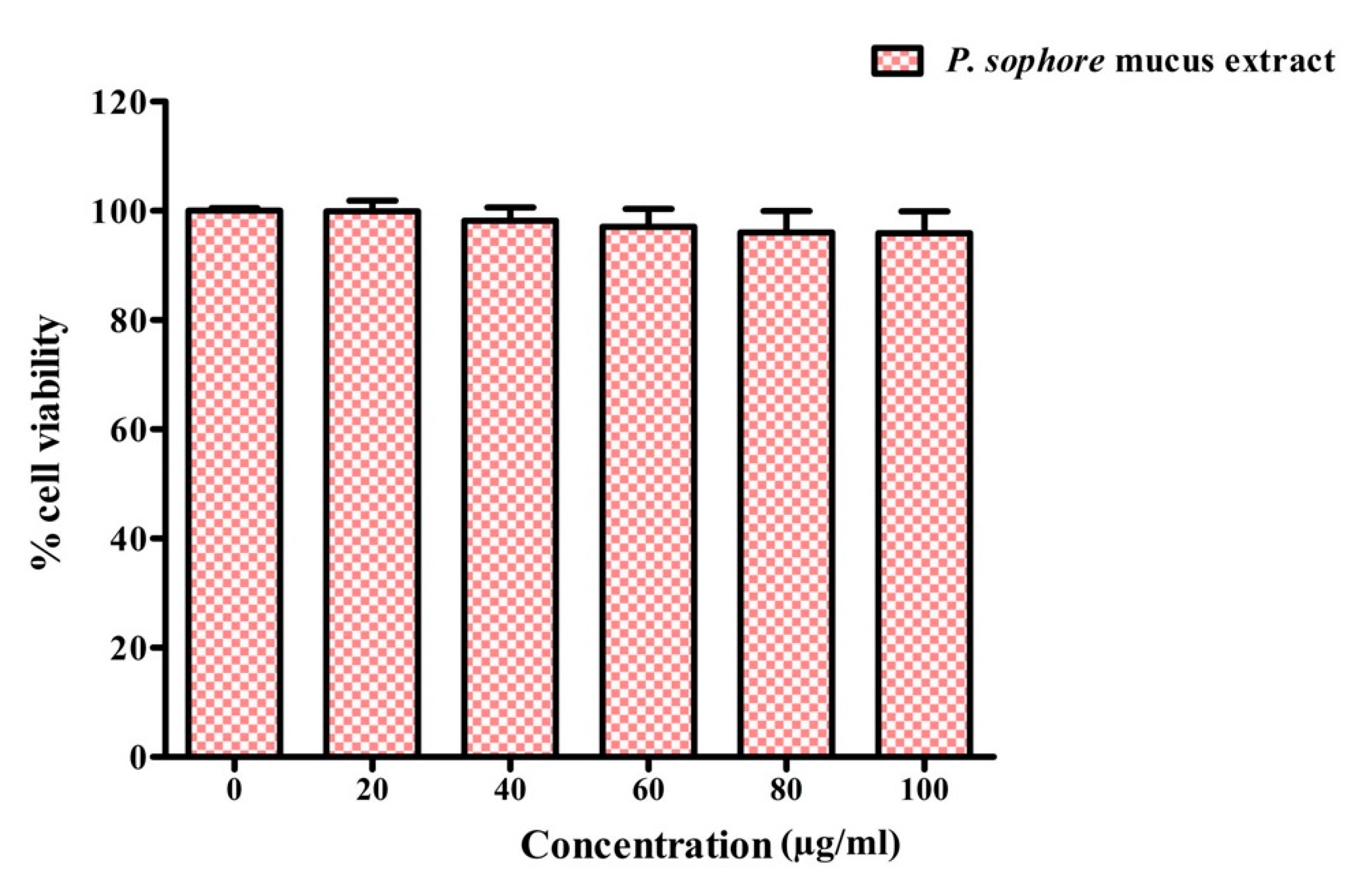
3.9. Cytotoxicity of Puntius sophore Mucus Extract to Normal Colon Cells
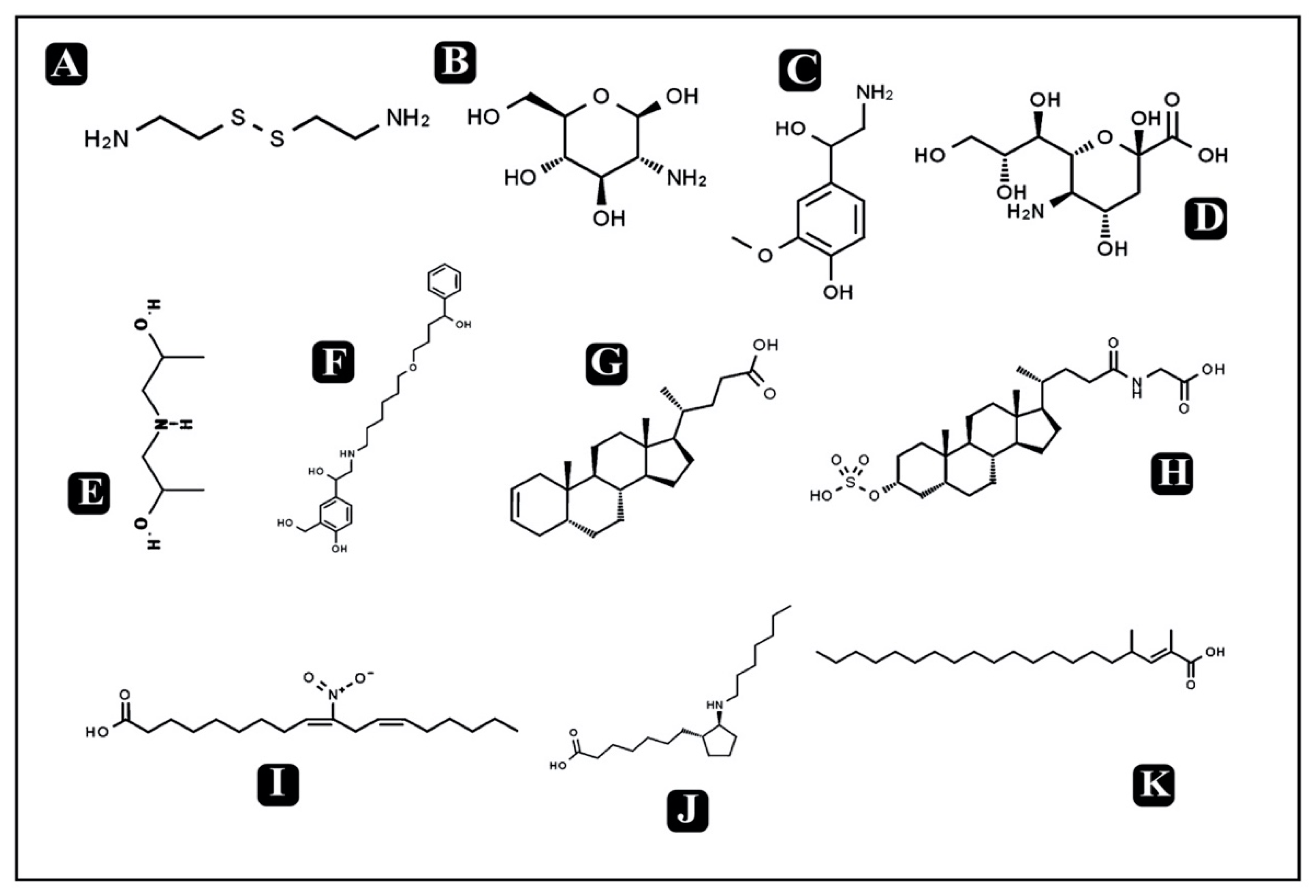
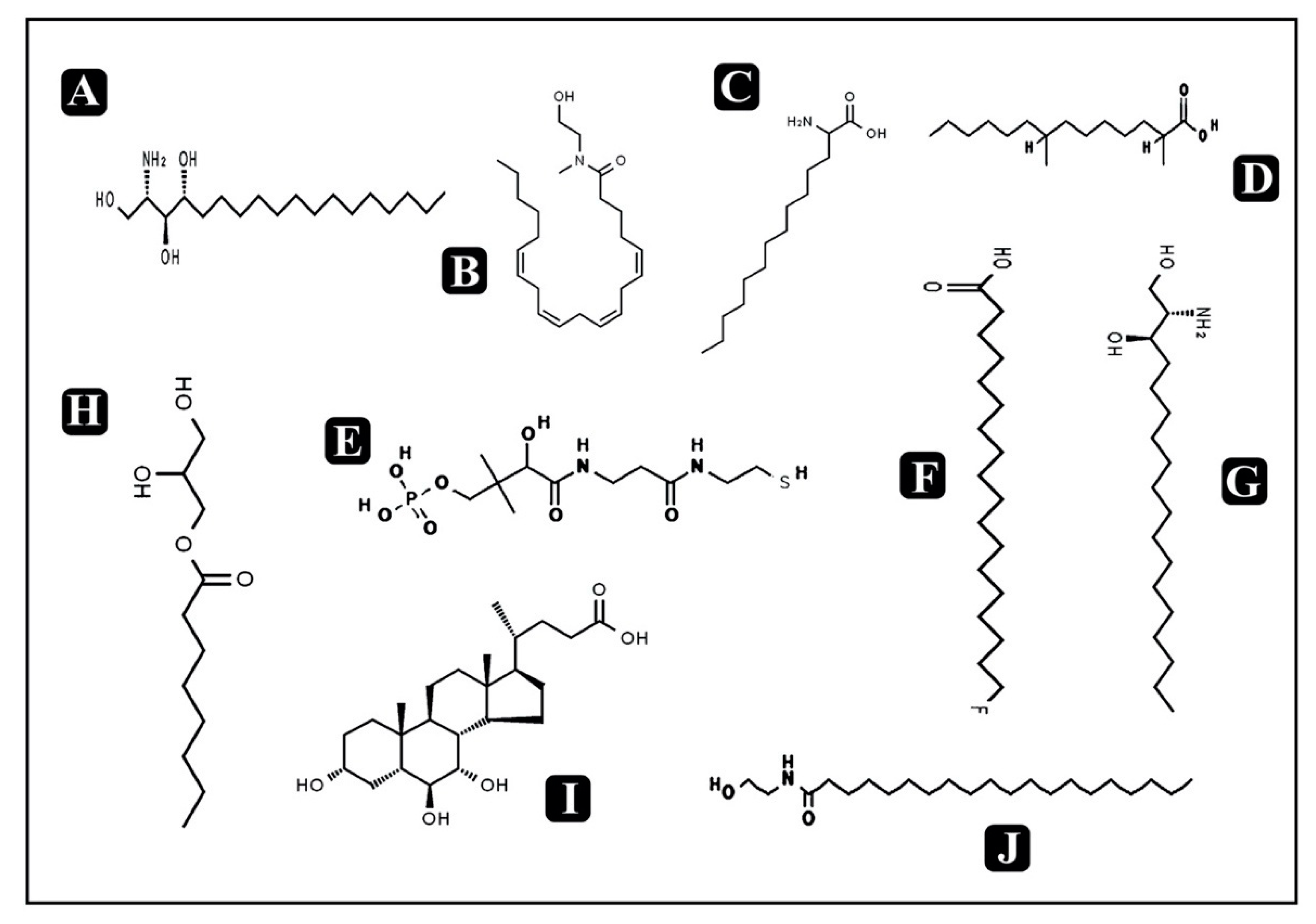
3.10. Bioactive Compounds Present in Puntius sophore Mucus Extract
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Donlan, R.M. Biofilms: Microbial Life on Surfaces. Emerg. Infect. Dis. 2002, 8, 881–890. [Google Scholar] [CrossRef] [PubMed]
- Limoli, D.; Jones, C.J.; Wozniak, D.J. Bacterial Extracellular Polysaccharides in Biofilm Formation and Function. Microbiol. Spectr. 2015, 3, 223–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karygianni, L.; Ren, Z.; Koo, H.; Thurnheer, T. Biofilm Matrixome: Extracellular Components in Structured Microbial Communities. Trends Microbiol. 2020. [Google Scholar] [CrossRef]
- Flemming, H.C. EPS-Then and Now. Microorganisms 2016, 4, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.; Singh, S.K.; Chowdhury, I.; Singh, R. Understanding the Mechanism of Bacterial Biofilms Resistance to Antimicrobial Agents. Open Microbiol. J. 2017, 11, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Ciofu, O.; Tolker-Nielsen, T. Tolerance and Resistance of Pseudomonas aeruginosa Biofilms to Antimicrobial Agents-How P. aeruginosa Can Escape Antibiotics. Front. Microbiol. 2019, 10, 913. [Google Scholar] [CrossRef] [Green Version]
- Sharma, D.; Misba, L.; Khan, A.U. Antibiotics versus biofilm: An emerging battleground in microbial communities. Antimicrob. Resist. Infect. Control. 2019, 8, 76. [Google Scholar] [CrossRef]
- Lebeaux, D.; Ghigo, J.-M.; Beloin, C. Biofilm-Related Infections: Bridging the Gap between Clinical Management and Fundamental Aspects of Recalcitrance toward Antibiotics. Microbiol. Mol. Biol. Rev. 2014, 78, 510–543. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Moser, C.; Wang, H.-Z.; Høiby, N.; Song, Z.-J. Strategies for combating bacterial biofilm infections. Int. J. Oral Sci. 2015, 7, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Jakobsen, T.H.; Tolker-Nielsen, T.; Givskov, M. Bacterial Biofilm Control by Perturbation of Bacterial Signaling Processes. Int. J. Mol. Sci. 2017, 18, 1970. [Google Scholar] [CrossRef] [Green Version]
- Adnan, M.; Alshammari, E.; Patel, M.; Ashraf, S.A.; Khan, S.; Hadi, S. Significance and potential of marine microbial natural bioactive compounds against biofilms/biofouling: Necessity for green chemistry. PeerJ 2018, 6, e5049. [Google Scholar] [CrossRef] [PubMed]
- Azad, C.S.; Saxena, M.; Siddiqui, A.J.; Bhardwaj, J.; Puri, S.K.; Dutta, G.P.; Anand, N.; Saxena, A.K.; Anand, N. Synthesis of primaquine glyco-conjugates as potential tissue schizontocidal antimalarial agents. Chem. Biol. Drug Des. 2017, 90, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Adnan, M. Bioactive potential of essential oil extracted from the leaves of Eucalyptus globulus (Myrtaceae). J. Pharmacogn. Phytochem. 2019, 8, 213–216. [Google Scholar]
- Adnan, M.; Patel, M.; Deshpande, S.; Alreshidi, M.; Siddiqui, A.J.; Reddy, M.N.; Emira, N.; De Feo, V. Effect of Adiantum philippense Extract on Biofilm Formation, Adhesion With Its Antibacterial Activities Against Foodborne Pathogens, and Characterization of Bioactive Metabolites: An in vitro-in silico Approach. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Alshammari, E.; Patel, M.; Sachidanandan, M.; Kumar, P.; Adnan, M. Potential Evaluation and Health Fostering Intrinsic Traits of Novel Probiotic Strain Enterococcus durans F3 Isolated from the Gut of Fresh Water Fish Catla catla. Food Sci. Anim. Resour. 2019, 39, 844–861. [Google Scholar] [CrossRef] [Green Version]
- Mahanty, A.; Ganguly, S.; Verma, A.; Sahoo, S.; Mitra, P.; Paria, P.; Sharma, A.P.; Singh, B.K.; Mohanty, B.P. Nutrient Profile of Small Indigenous Fish Puntius sophore: Proximate Composition, Amino Acid, Fatty Acid and Micronutrient Profiles. Natl. Acad. Sci. Lett. 2014, 37, 39–44. [Google Scholar] [CrossRef]
- Ahamed, F.; Ahmed, Z.F.; Hossain, M.Y.; Ohtomi, J. Growth study of the Pool Barb Puntius sophore (Cyprinidae: Barbinae) through multi model inferences. Zool. Stud. 2012, 51, 1077–1085. [Google Scholar]
- Sarjubala, W.M.S.; Hawaibam, R.; Chungkham, S. Nutritional properties of some freshwater fish species of Manipur, India. J. Coldwater Fish. 2018, 1, 48–53. [Google Scholar]
- Adnan, M.; Patel, M.; Reddy, M.N.; Alshammari, E. Formulation, evaluation and bioactive potential of Xylaria primorskensis terpenoid nanoparticles from its major compound xylaranic acid. Sci. Rep. 2018, 8, 1740. [Google Scholar] [CrossRef] [PubMed]
- Adnan, M.; Patel, M.; Hadi, S. Functional and health promoting inherent attributes of Enterococcus hirae F2 as a novel probiotic isolated from the digestive tract of the freshwater fish Catla catla. PeerJ 2017, 5, e3085. [Google Scholar] [CrossRef] [Green Version]
- Diamond, G.; Zasloff, M.; Eck, H.; Brasseur, M.; Maloy, W.L.; Bevins, C.L. Tracheal antimicrobial peptide, a cysteine-rich peptide from mammalian tracheal mucosa: Peptide isolation and cloning of a cDNA. Proc. Natl. Acad. Sci. USA 1991, 88, 3952–3956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing. Twenty Fourth Informational Supplement, M100-S24.; National Committee for Clinical Laborartory Standards: Wayne, PA, USA, 2014. [Google Scholar]
- Kuete, V. African Medicinal Spices and Vegetables and Their Potential in the Management of Metabolic Syndrome. In Medicinal Spices and Vegetables from Africa; Elsevier BV: Amsterdam, The Netherlands, 2017; pp. 315–327. [Google Scholar]
- Silveira, C.P.; Torres-Rodriguez, J.M.; Alvarado-Ramírez, E.; Murciano-Gonzalo, F.; Dolande, M.; Panizo, M.; Reviakina, V. MICs and minimum fungicidal concentrations of amphotericin B, itraconazole, posaconazole and terbinafine in Sporothrix schenckii. J. Med. Microbiol. 2009, 58, 1607–1610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pillai, S.K.; Moellering, R.C.; Eliopoulos, G.M. Antimicrobial combinations. In Antibiotics in Laboratory Medicine, 5th ed.; Lorian, V., Ed.; The Lippincott Williams & Wilkins Co.: Philadelphia, PA, USA, 2005; pp. 365–440. [Google Scholar]
- Božić, D.D.; Milenković, M.; Ivković, B.; Ćirković, I. Antibacterial activity of three newly-synthesized chalcones & synergism with antibiotics against clinical isolates of methicillin-resistant Staphylococcus aureus. Indian J. Med. Res. 2014, 140, 130–137. [Google Scholar]
- Lee, K.W.K.; Periasamy, S.; Mukherjee, M.; Xie, C.; Kjelleberg, S.; Rice, S.A. Biofilm development and enhanced stress resistance of a model, mixed-species community biofilm. ISME J. 2013, 8, 894–907. [Google Scholar] [CrossRef]
- Nostro, A.; Roccaro, A.S.; Bisignano, G.; Marino, A.; Cannatelli, M.A.; Pizzimenti, F.C.; Cioni, P.L.; Procopio, F.; Blanco, A.R. Effects of oregano, carvacrol and thymol on Staphylococcus aureus and Staphylococcus epidermidis biofilms. J. Med. Microbiol. 2007, 56, 519–523. [Google Scholar] [CrossRef]
- Plyuta, V.; Zaitseva, J.; Lobakova, E.; Zagoskina, N.; Kuznetsov, A.; Khmel, I.A. Effect of plant phenolic compounds on biofilm formation byPseudomonas aeruginosa. APMIS 2013, 121, 1073–1081. [Google Scholar] [CrossRef]
- Musthafa, K.S.; Ravi, A.V.; Annapoorani, A.; Packiavathy, S.V.; Pandian, S.K. Evaluation of Anti-Quorum-Sensing Activity of Edible Plants and Fruits through Inhibition of the N-Acyl-Homoserine Lactone System in Chromobacterium violaceum and Pseudomonas aeruginosa. Chemotherapy 2010, 56, 333–339. [Google Scholar] [CrossRef]
- Adnan, M.; Sousa, A.M.; Machado, I.; Pereira, M.O.; Khan, S.; Morton, G.; Hadi, S. Role of bolA and rpoS genes in biofilm formation and adherence pattern by Escherichia coli K-12 MG1655 on polypropylene, stainless steel, and silicone surfaces. Acta Microbiol. Immunol. Hung. 2017, 64, 179–189. [Google Scholar] [CrossRef] [Green Version]
- Patel, M.; Reddy, M.N. Discovery of the World’s Smallest Terrestrial Pteridophyte. Sci. Rep. 2018, 8, 5911. [Google Scholar] [CrossRef] [Green Version]
- Ramage, G.; Walle, K.V.; Wickes, B.L.; López-Ribot, J.L. Standardized Method for In Vitro Antifungal Susceptibility Testing of Candida albicansBiofilms. Antimicrob. Agents Chemother. 2001, 45, 2475–2479. [Google Scholar] [CrossRef] [Green Version]
- Nett, J.E.; Cain, M.T.; Crawford, K.; Andes, D.R. Optimizing a Candida Biofilm Microtiter Plate Model for Measurement of Antifungal Susceptibility by Tetrazolium Salt Assay. J. Clin. Microbiol. 2011, 49, 1426–1433. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, A.J.; Bhardwaj, J.; Goyal, M.; Prakash, K.; Adnan, M.; Alreshidi, M.M.; Patel, M.; Soni, A.; Redman, W. Immune responses in liver and spleen against Plasmodium yoelii pre-erythrocytic stages in Swiss mice model. J. Adv. Res. 2020, 24, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Borucki, M.K.; Krug, M.J.; Muraoka, W.T.; Call, D.R. Discrimination among Listeria monocytogenes isolates using a mixed genome DNA microarray. Veter. Microbiol. 2003, 92, 351–362. [Google Scholar] [CrossRef]
- Reddy, M.N.; Adnan, M.; Alreshidi, M.M.; Saeed, M.; Patel, M. Evaluation of Anticancer, Antibacterial and Antioxidant Properties of a Medicinally Treasured Fern Tectaria coadunata with its Phytoconstituents Analysis by HR-LCMS. Anti-Cancer Agents Med. Chem. 2020, 20, 1. [Google Scholar] [CrossRef] [PubMed]
- Fraser-Pitt, D.J.; Mercer, D.K.; Smith, D.; Kowalczuk, A.; Robertson, J.; Lovie, E.; Perenyi, P.; Cole, M.J.; Doumith, M.; Hill, R.L.R.; et al. Cysteamine, an Endogenous Aminothiol, and Cystamine, the Disulfide Product of Oxidation, IncreasePseudomonas aeruginosaSensitivity to Reactive Oxygen and Nitrogen Species and Potentiate Therapeutic Antibiotics against Bacterial Infection. Infect. Immun. 2018, 86, e00947-17. [Google Scholar] [CrossRef] [Green Version]
- Malik, S.; Singh, M.; Mathur, A. Antimicrobial Activity of Food Grade Glucosamine. Int. J. Biotechnol. Bioeng. Res. 2013, 4, 307–312. [Google Scholar]
- Sakko, M.; Moore, C.; Novak-Frazer, L.; Rautemaa, V.; Sorsa, T.; Hietala, P.; Järvinen, A.; Bowyer, P.; Tjäderhane, L.; Rautemaa, R. 2-hydroxyisocaproic acid is fungicidal for CandidaandAspergillusspecies. Mycoses 2013, 57, 214–221. [Google Scholar] [CrossRef]
- Bellini, A.M.; Mencini, E.; Quaglio, M.P.; Guameri, M.; Fini, A. Antimicrobial activity of basic cholane derivatives. X. Synthesis of 3α- and 3β-amino-5β-cholan-24-oic acids. Steroids 1991, 56, 395–398. [Google Scholar] [CrossRef]
- Choi, J.-S.; Park, N.-H.; Hwang, S.-Y.; Sohn, J.H.; Kwak, I.; Cho, K.K.; Choi, I.S. The antibacterial activity of various saturated and unsaturated fatty acids against several oral pathogens. J. Environ. Biol. 2013, 34, 673–676. [Google Scholar]
- Başpınar, Y.; Kotmakçı, M.; Öztürk, I. Antimicrobial Activity of Phytosphingosine Nanoemulsions against Bacteria and Yeasts. Celal Bayar Üniv. Fen Bilimleri Derg. 2018, 14, 223–228. [Google Scholar] [CrossRef]
- Fischer, C.L. Antimicrobial Activity of Host-Derived Lipids. Antibiotics 2020, 9, 75. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.-S.; Redondo-Solano, M.; Thippareddi, H. Inactivation of Escherichia coli O157:H7 and Salmonella spp. on alfalfa seeds by caprylic acid and monocaprylin. Int. J. Food Microbiol. 2010, 144, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Fukiya, S.; Yokota, A. Comprehensive evaluation of the bactericidal activities of free bile acids in the large intestine of humans and rodents. J. Lipid Res. 2017, 58, 1143–1152. [Google Scholar] [CrossRef] [Green Version]
- Adnan, M.; Morton, G.; Hadi, S. Analysis of rpoS and bolA gene expression under various stress-induced environments in planktonic and biofilm phase using 2−ΔΔCT method. Mol. Cell. Biochem. 2011, 357, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Adnan, M.; Morton, G.; Singh, J.; Hadi, S. Contribution of rpoS and bolA genes in biofilm formation in Escherichia coli K-12 MG1655. Mol. Cell. Biochem. 2010, 342, 207–213. [Google Scholar] [CrossRef]
- Jamal, M.; Ahmad, W.; Andleeb, S.; Jalil, F.; Imran, M.; Nawaz, M.A.; Hussain, T.; Ali, M.; Rafiq, M.; Kamil, M.A. Bacterial biofilm and associated infections. J. Chin. Med. Assoc. 2018, 81, 7–11. [Google Scholar] [CrossRef]
- Song, F.; Koo, H.; Ren, D. Effects of Material Properties on Bacterial Adhesion and Biofilm Formation. J. Dent. Res. 2015, 94, 1027–1034. [Google Scholar] [CrossRef]
- Adnan, M.; Alshammari, E.; Ashraf, S.A.; Patel, K.; Lad, K.; Patel, M. Physiological and Molecular Characterization of Biosurfactant Producing Endophytic Fungi Xylaria regalis from the Cones of Thuja plicata as a Potent Plant Growth Promoter with Its Potential Application. BioMed Res. Int. 2018, 2018, 7362148. [Google Scholar] [CrossRef] [Green Version]
- Adnan, M.; Ashraf, S.A.; Khan, S.; Alshammari, E.; AwadElkareem, A.M. Effect of pH, temperature and incubation time on cordycepin production from Cordyceps militaris using solid-state fermentation on various substrates. CyTA - J. Food 2017, 15, 617–621. [Google Scholar] [CrossRef] [Green Version]
- Raison, R.L.; Dos Remedios, N. The Hagfish Immune System. In The Biology of Hagfishes; Springer Science and Business Media LLC: Berlin, Germany, 1998; pp. 334–344. [Google Scholar]
- Rolff, J. Why did the acquired immune system of vertebrates evolve? Dev. Comp. Immunol. 2007, 31, 476–482. [Google Scholar] [CrossRef]
- Spitzer, R.H.; Koch, E.A. Hagfish Skin and Slime Glands. In The Biology of Hagfishes; Springer Science and Business Media LLC: Berlin, Germany, 1998; pp. 109–132. [Google Scholar]
- Subramanian, S.; MacKinnon, S.L.; Ross, N.W. A comparative study on innate immune parameters in the epidermal mucus of various fish species. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2007, 148, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Shephard, K.L. Mucus on the epidermis of fish and its influence on drug delivery. Adv. Drug Deliv. Rev. 1993, 11, 403–417. [Google Scholar] [CrossRef]
- Button, B.; Boucher, R.C.; University of North Carolina Virtual Lung Group. Role of mechanical stress in regulating airway surface hydration and mucus clearance rates. Respir. Physiol. Neurobiol. 2008, 163, 189–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Austin, B.; McIntosh, D. Natural antibacterial compounds on the surface of rainbow trout, Salmo gairdneri Richardson. J. Fish Dis. 1988, 11, 275–277. [Google Scholar] [CrossRef]
- Fouz, B.; Devesa, S.; Gravningen, K.; Barja, J.L.; Toranzo, A.E. Antibacterial action of the mucus of turbot. Bull. Eur. Assoc. Fish Pathol. 1990, 10, 56–59. [Google Scholar]
- Hjelmeland, K.; Christie, M.; Raa, J. Skin mucus protease from rainbow trout, Salmo gairdneri Richardson, and its biological significance. J. Fish Biol. 1983, 23, 13–22. [Google Scholar] [CrossRef]
- Grinde, B.; Jollès, J.; Jollès, P. Purification and characterization of two lysozymes from rainbow trout (Salmo gairdneri). JBIC J. Biol. Inorg. Chem. 1988, 173, 269–273. [Google Scholar] [CrossRef]
- Nagashima, Y.; Sendo, A.; Shimakura, K.; Shiomi, K.; Kobayashi, T.; Kimura, B.; Fujii, T. Antibacterial factors in skin mucus of rabbitfishes. J. Fish Biol. 2001, 58, 1761–1765. [Google Scholar] [CrossRef]
- Sarmaşik, A. Antimicrobial peptides: A potential therapeutic alternative for the treatment of fish diseases. Turk. J. Biol. 2002, 26, 201–207. [Google Scholar]
- Mai-Prochnow, A.; Clauson, M.; Hong, J.; Murphy, A.B. Gram positive and Gram negative bacteria differ in their sensitivity to cold plasma. Sci. Rep. 2016, 6, 38610. [Google Scholar] [CrossRef] [Green Version]
- Rogers, H.J.; Perkins, H.R.; Ward, J.B. Microbial Cell Walls and Membranes. Microb. Cell Walls Membr. 1980. [Google Scholar]
- Vollmer, W.; Blanot, D.; De Pedro, M.A. Peptidoglycan structure and architecture. FEMS Microbiol. Rev. 2008, 32, 149–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuppulakshmi, C.; Prakash, M.; Gunasekaran, G.; Manimegalai, G.; Sarojini, S. Antibacterial properties of fish mucus from Channa punctatus and Cirrhinus mrigala. Eur. Rev. Med. Pharmacol. Sci. 2008, 12, 149–153. [Google Scholar] [PubMed]
- Manivasagan, P.; Neelamegam, A.; Ashokkumar, S.; Palanisamy, S. Studies on the proteinaceous gel secretion from the skin of the catfish, Arius maculatus (Thunberg, 1792). Afr. J. Biotechnol. 2009, 8, 7125–7129. [Google Scholar]
- Subramanian, S.; Ross, N.W.; MacKinnon, S.L. Comparison of antimicrobial activity in the epidermal mucus extracts of fish. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2008, 150, 85–92. [Google Scholar] [CrossRef]
- Bragadeeswaran, S.; Thangaraj, S. Hemolytic and Antibacterial Studies on Skin Mucus of Eel Fish, Anguilla anguilla Linnaues, 1758. Asian J. Biol. Sci. 2011, 4, 272–276. [Google Scholar] [CrossRef] [Green Version]
- Ellis, A. Immunity to bacteria in fish. Fish Shellfish. Immunol. 1999, 9, 291–308. [Google Scholar] [CrossRef]
- Cole, A.M.; Weis, P.; Diamond, G. Isolation and Characterization of Pleurocidin, an Antimicrobial Peptide in the Skin Secretions of Winter Flounder. J. Biol. Chem. 1997, 272, 12008–12013. [Google Scholar] [CrossRef] [Green Version]
- Saritha, K.; Rajesh, A.; Manjulatha, K.; Setty, O.H.; Yenugu, S. Mechanism of antibacterial action of the alcoholic extracts of Hemidesmus indicus (L.) R. Br. ex Schult, Leucas aspera (Wild.), Plumbago zeylanica L., and Tridax procumbens (L.) R. Br. ex Schult. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Ocampo, P.S.; Lázár, V.; Papp, B.; Arnoldini, M.; Wiesch, P.A.Z.; Busa-Fekete, R.; Fekete, G.; Pál, C.; Ackermann, M.; Bonhoeffer, S. Antagonism between Bacteriostatic and Bactericidal Antibiotics Is Prevalent. Antimicrob. Agents Chemother. 2014, 58, 4573–4582. [Google Scholar] [CrossRef] [Green Version]
- Leme, A.P.; Koo, H.; Bellato, C.; Bedi, G.; Cury, J. The Role of Sucrose in Cariogenic Dental Biofilm Formation—New Insight. J. Dent. Res. 2006, 85, 878–887. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.; Hwang, G.; Liu, Y.; Wang, Y.; Singh, A.P.; Vorsa, N.; Koo, H. Cranberry Flavonoids Modulate Cariogenic Properties of Mixed-Species Biofilm through Exopolysaccharides-Matrix Disruption. PLoS ONE 2015, 10, e0145844. [Google Scholar] [CrossRef] [PubMed]
- Lentino, J.R. Prosthetic Joint Infections: Bane of Orthopedists, Challenge for Infectious Disease Specialists. Clin. Infect. Dis. 2003, 36, 1157–1161. [Google Scholar] [CrossRef] [PubMed]
- Desbois, A.; Smith, V.J. Antibacterial free fatty acids: Activities, mechanisms of action and biotechnological potential. Appl. Microbiol. Biotechnol. 2009, 85, 1629–1642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, R.W. Fish cutaneous mucus: A new source of skin surface lipid. Lipids 1970, 5, 947–949. [Google Scholar] [CrossRef]
- Ekman, D.R.; Skelton, D.M.; Davis, J.M.; Villeneuve, D.L.; Cavallin, J.E.; Schroeder, A.; Jensen, K.M.; Ankley, G.T.; Collette, T.W. Metabolite Profiling of Fish Skin Mucus: A Novel Approach for Minimally-Invasive Environmental Exposure Monitoring and Surveillance. Environ. Sci. Technol. 2015, 49, 3091–3100. [Google Scholar] [CrossRef]
- Jais, A.M.M.; Matori, M.; Kittakoop, P.; Sowanborirux, K. Fatty Acid Compositions in Mucus and Roe of Haruan, Channa Striatus, for Wound Healing. Gen. Pharmacol. Vasc. Syst. 1998, 30, 561–563. [Google Scholar] [CrossRef] [Green Version]
- Di Somma, A.; Moretta, A.; Canè, C.; Cirillo, A.; Duilio, A. Antimicrobial and Antibiofilm Peptides. Biomolecules 2020, 10, 652. [Google Scholar] [CrossRef] [Green Version]
- Galdiero, E.; Lombardi, L.; Falanga, A.; Libralato, G.; Guida, M.; Carotenuto, R. Biofilms: Novel Strategies Based on Antimicrobial Peptides. Pharmaceutics 2019, 11, 322. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.K.; Kang, H.K.; Ko, S.J.; Hong, M.J.; Bang, J.K.; Seo, C.H.; Park, Y. Mechanisms driving the antibacterial and antibiofilm properties of Hp1404 and its analogue peptides against multidrug-resistant Pseudomonas aeruginosa. Sci. Rep. 2018, 8, 1763. [Google Scholar] [CrossRef] [Green Version]
- Chung, P.Y.; Khanum, R. Antimicrobial peptides as potential anti-biofilm agents against multidrug-resistant bacteria. J. Microbiol. Immunol. Infect. 2017, 50, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Andrea, A.; Molchanova, N.; Jenssen, H. Antibiofilm Peptides and Peptidomimetics with Focus on Surface Immobilization. Biomolecules 2018, 8, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raheem, N.; Straus, S.K. Mechanisms of Action for Antimicrobial Peptides with Antibacterial and Antibiofilm Functions. Front. Microbiol. 2019, 10, 2866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dostert, M.; Belanger, C.R.; Hancock, R.E. Design and Assessment of Anti-Biofilm Peptides: Steps toward Clinical Application. J. Innate Immun. 2018, 11, 193–204. [Google Scholar] [CrossRef]
- Montanaro, L.; Poggi, A.; Visai, L.; Ravaioli, S.; Campoccia, D.; Speziale, P.; Arciola, C.R. Extracellular DNA in Biofilms. Int. J. Artif. Organs 2011, 34, 824–831. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strains | P. sophore Mucus Extract (µg/mL) | Gentamicin (µg/mL) | ||
|---|---|---|---|---|
| MIC | MBC | MIC | MBC | |
| E. coli | 250 | 500 | 7.8 | 15 |
| P. aeruginosa | 500 | 1000 | 15 | 31 |
| B. subtilis | 1000 | 1500 | 15 | 31 |
| S. aureus | 1500 | >1500 | 15 | 31 |
| Bacterial Strain | Mucus Extract | Gentamicin | FICI | Effect |
|---|---|---|---|---|
| MIC * | MIC * | |||
| E. coli | 31 | 1.9 | 0.375 | Synergy |
| P. aeruginosa | 83 | 4.4 | 0.452 | Synergy |
| B. subtilis | 200 | 44 | 0.485 | Synergy |
| S. aureus | 375 | 44 | 0.535 | Additive |
| Bioactive Metabolites | Formula | Class | m/z | RT (min) | Mass | Mode of Action | References |
|---|---|---|---|---|---|---|---|
| Cysteamine | C4H12N2S2 | Organic disulfide | 150.15 | 0.769 | 152.04 | Anti-infective activity against bacteria, viruses, and malarial parasites | [38] |
| Glucosamine | C6H13NO5 | Amino sugar | 180.19 | 1.004 | 179.07 | Antibacterial activity against different Gram- positive and Gram-negative bacteria and antifungal activity against few fungal strains | [39] |
| Lys Ser Phe | C18H28N4O5 | Small peptide | 387.46 | 1.026 | 380.20 | - | |
| Met Lys | C11H23N3O3S | Small peptide | 282.59 | 1.034 | 277.14 | - | |
| Normetanephrine | C9H13NO3 | Catecholamine | 181.07 | 1.046 | 183.08 | - | |
| Dodecaprenylphosphategalacturonic acid | C31H51O10P | Bactoprenol | 620.64 | 1.048 | 614.32 | - | |
| Bis (2-hydroxypropyl) amine | C6H15NO2 | Amino alcohol | 130.24 | 1.096 | 133.11 | - | |
| Lys Met Thr | C15H30N4O5S | Small peptide | 380.20 | 1.1 | 378.19 | - | |
| Neuraminic acid | C9H17NO8 | Amino sugar | 272.67 | 1.443 | 267.09 | Antibacterial activity against different Gram- positive and Gram-negative bacteria and antifungal activity against few fungal strains | [40] |
| Thr Ile Tyr | C19H29N3O6 | Small peptide | 403.27 | 3.217 | 395.20 | - | |
| Pro ArgGln | C16H29N7O5 | Small peptide | 398.26 | 4.654 | 399.22 | - | |
| Hydroxysalmeterol | C25H37NO5 | Amino alcohol | 433.29 | 4.836 | 431.27 | - | |
| 5beta-Chol-2-en-24-oic Acid | C24H38O2 | Alcohol | 356.46 | 6.578 | 358.28 | Antimicrobialactivity | [41] |
| Sulfolithocholylglycine | C26H43NO7S | Glycine conjugate (sterol lipid) | 512.29 | 7.034 | 513.27 | - | |
| 10-nitro,9Z,12Z-octadecadienoic acid | C18H31NO4 | Fatty acids | 325.20 | 7.59 | 325.22 | Antimicrobial activity against oral pathogens | [42] |
| 3-Ketosphingosine | C18H35NO2 | Sphingosine (sphingolipid) | 296.24 | 9.879 | 297.26 | - | |
| GlnGln Met | C15H27N5O6S | Small peptide | 404.19 | 10.039 | 405.17 | - | |
| 13-Azaprostanoic acid | C19H37NO2 | Fatty acids | 311.20 | 10.51 | 311.28 | Gastroprotective activity | |
| 2,4-Dimethyl-2-eicosenoic acid | C22H42O2 | Fatty acids | 337.32 | 10.841 | 338.31 | ||
| Phytosphingosine | C18H39NO3 | Sphingolipids | 315.31 | 11.247 | 317.29 | Anti-inflammatory and antimicrobial activity against different bacteria and yeast | [43] |
| N-methyl N-(2-hydroxy-ethyl) arachidonoyl amine | C23H39NO2 | Fatty amides | 360.16 | 11.691 | 361.29 | - | |
| ArgGlnPhe | C20H31N7O5 | Small peptide | 447.35 | 11.958 | 449.23 | - | |
| 2-Amino-tetradecanoic acid | C14H29NO2 | Fatty acids | 244.20 | 12.626 | 243.21 | ||
| 2,4-Dimethyl-tetradecanoic acid | C16H32O2 | Fatty acids | 256.23 | 13.833 | 256.24 | ||
| D-Pantetheine 4’-phosphate | C11H23N2O7PS | Organophosphorus compound | 356.09 | 14.121 | 358.09 | - | |
| 18-Fluoro-octadecanoic acid | C18H35FO2 | Fatty acids | 302.19 | 18.385 | 302.26 | ||
| Dihydrosphingosine | C18H39NO2 | Amino alcohol | 309.35 | 18.774 | 301.29 | Antimicrobial activity against a variety of opportunistic bacteria, viruses and fungi | [44] |
| 1-Octanoyl-rac-glycerol | C11H22O4 | Monoacylglyc-erol (glycerolipid) | 220.31 | 18.875 | 218.29 | Antibacterial activity against D. congolensis, Campylobacter spp., E. coli, Listeria spp., and Salmonella spp. | [45] |
| GlnArg Lys | C17H34N8O5 | Small peptide | 437.45 | 19.598 | 430.26 | - | |
| 3alpha,6beta,7alpha-Trihydroxy-5beta-cholan-24-oic acid | C24H40O5 | Sterol lipid | 410.29 | 19.622 | 408.28 | Antibacterial activity against Bifidobacterium breve, Blautiacoccoides, and Bacteroides thetaiotaomicron | [46] |
| PheGlnArg | C20H31N7O5 | Small peptide | 457.21 | 19.879 | 449.23 | - | |
| N-(2-hydroxyethyl) icosanamide | C22H45NO2 | Endocannabinoids | 360.37 | 20.094 | 355.34 | - | |
| Lys Gln Leu | C17H33N5O5 | Small peptide | 397.23 | 26.95 | 387.24 | - | |
| Arg Ser Ser | C12H24N6O6 | Small peptide | 352.20 | 26.995 | 348.17 | - | |
| ArgGlnArg | C17H34N10O5 | Small peptide | 462.11 | 26.997 | 458.26 | - | |
| Ala Lys Ile | C15H30N4O4 | Small peptide | 349.28 | 27.063 | 330.22 | - | |
| Ile Thr Pro | C15H27N3O5 | Small peptide | 355.39 | 27.072 | 329.19 | - | |
| Lys Gln Leu | C17H33N5O5 | Small peptide | 390.21 | 27.118 | 387.24 | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Patel, M.; Ashraf, M.S.; Siddiqui, A.J.; Ashraf, S.A.; Sachidanandan, M.; Snoussi, M.; Adnan, M.; Hadi, S. Profiling and Role of Bioactive Molecules from Puntius sophore (Freshwater/Brackish Fish) Skin Mucus with Its Potent Antibacterial, Antiadhesion, and Antibiofilm Activities. Biomolecules 2020, 10, 920. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10060920
Patel M, Ashraf MS, Siddiqui AJ, Ashraf SA, Sachidanandan M, Snoussi M, Adnan M, Hadi S. Profiling and Role of Bioactive Molecules from Puntius sophore (Freshwater/Brackish Fish) Skin Mucus with Its Potent Antibacterial, Antiadhesion, and Antibiofilm Activities. Biomolecules. 2020; 10(6):920. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10060920
Chicago/Turabian StylePatel, Mitesh, Mohammad Saquib Ashraf, Arif Jamal Siddiqui, Syed Amir Ashraf, Manojkumar Sachidanandan, Mejdi Snoussi, Mohd Adnan, and Sibte Hadi. 2020. "Profiling and Role of Bioactive Molecules from Puntius sophore (Freshwater/Brackish Fish) Skin Mucus with Its Potent Antibacterial, Antiadhesion, and Antibiofilm Activities" Biomolecules 10, no. 6: 920. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10060920