
The Protein Kinase Inhibitor Midostaurin Improves Functional Neurological Recovery and Attenuates Inflammatory Changes Following Traumatic Cervical Spinal Cord Injury

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Care and SCI
2.2. Spleen Mass
2.3. RNA and Protein Extraction
2.4. RT-qPCR
2.5. Western Blotting
2.6. Multiplex Luminex Assay
2.7. Neurobehavioral Assessments
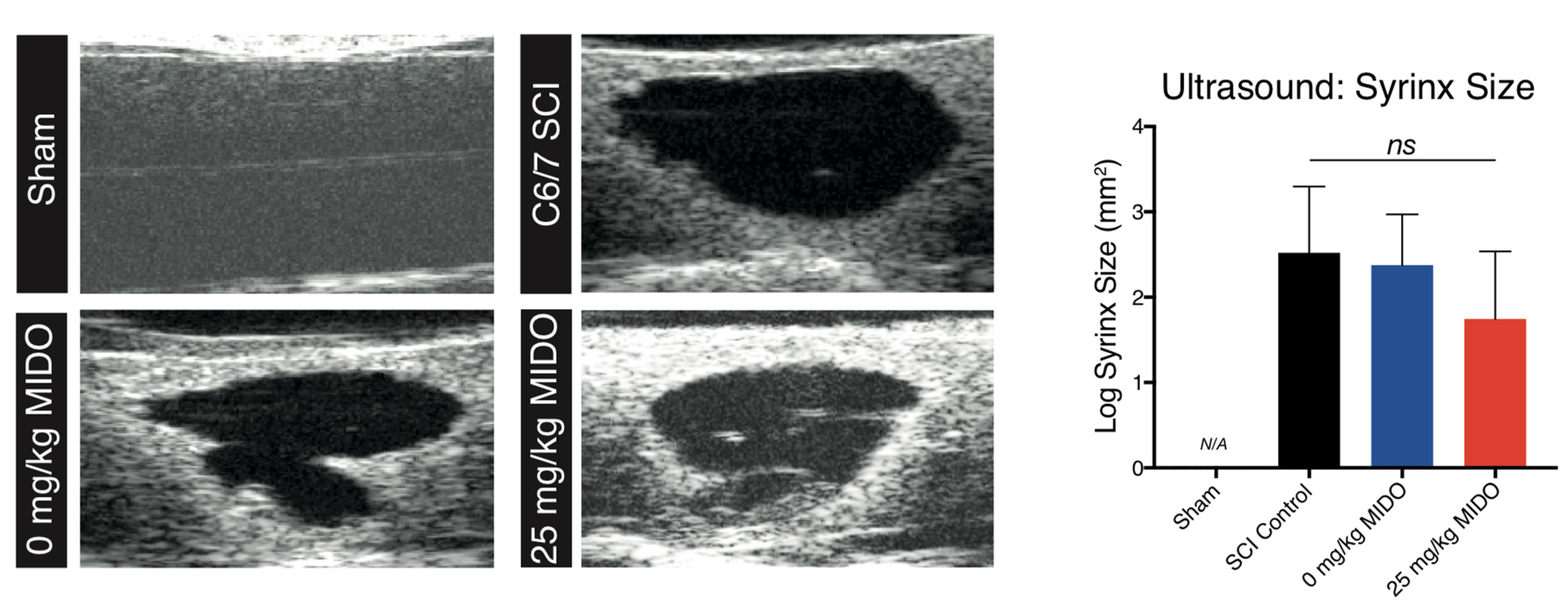
2.8. Ultrasonography
2.9. Histomorphometric Analysis
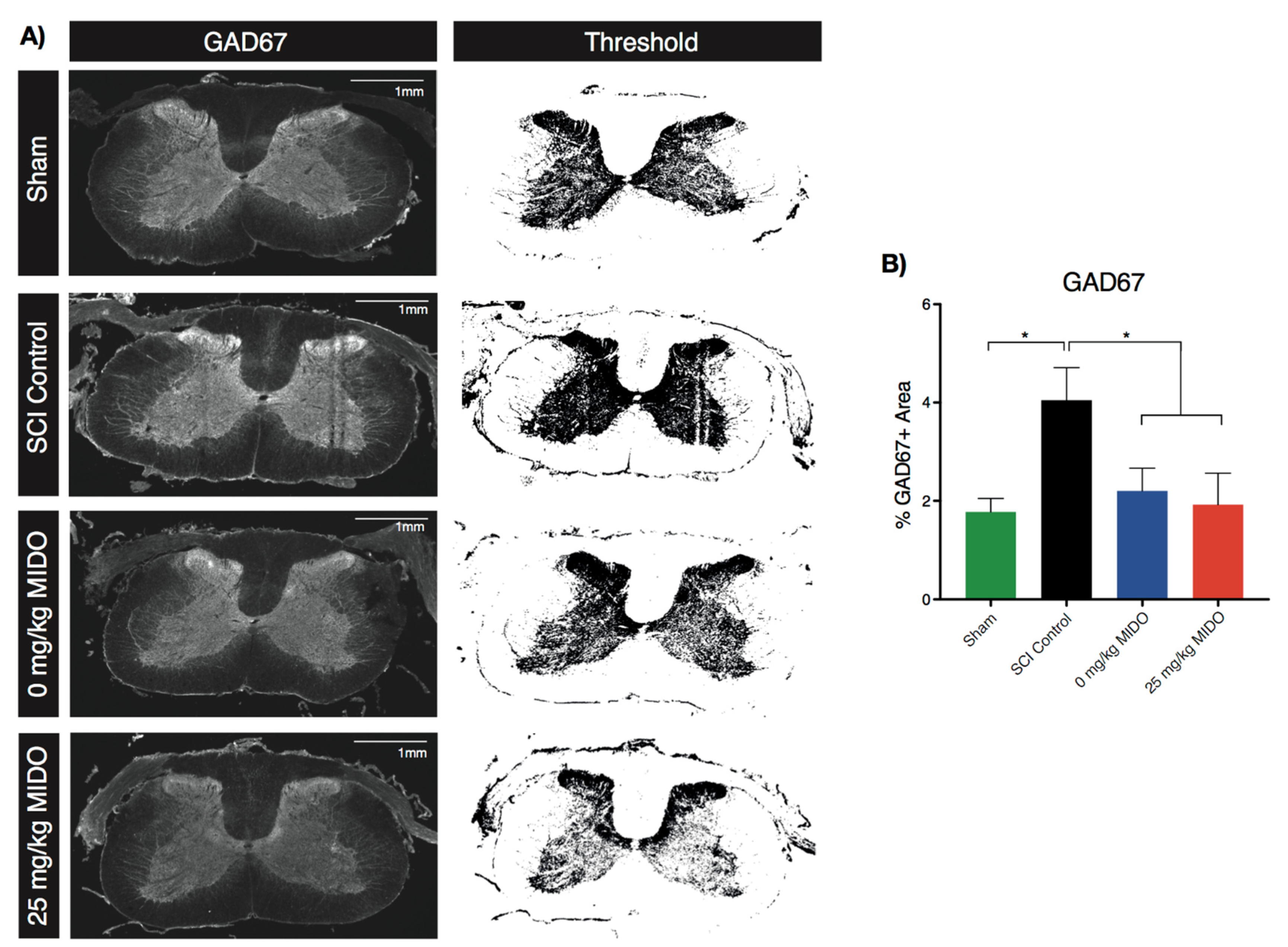
2.10. Immunohistochemistry
2.11. Statistical and Bioinformatics Analysis
3. Results
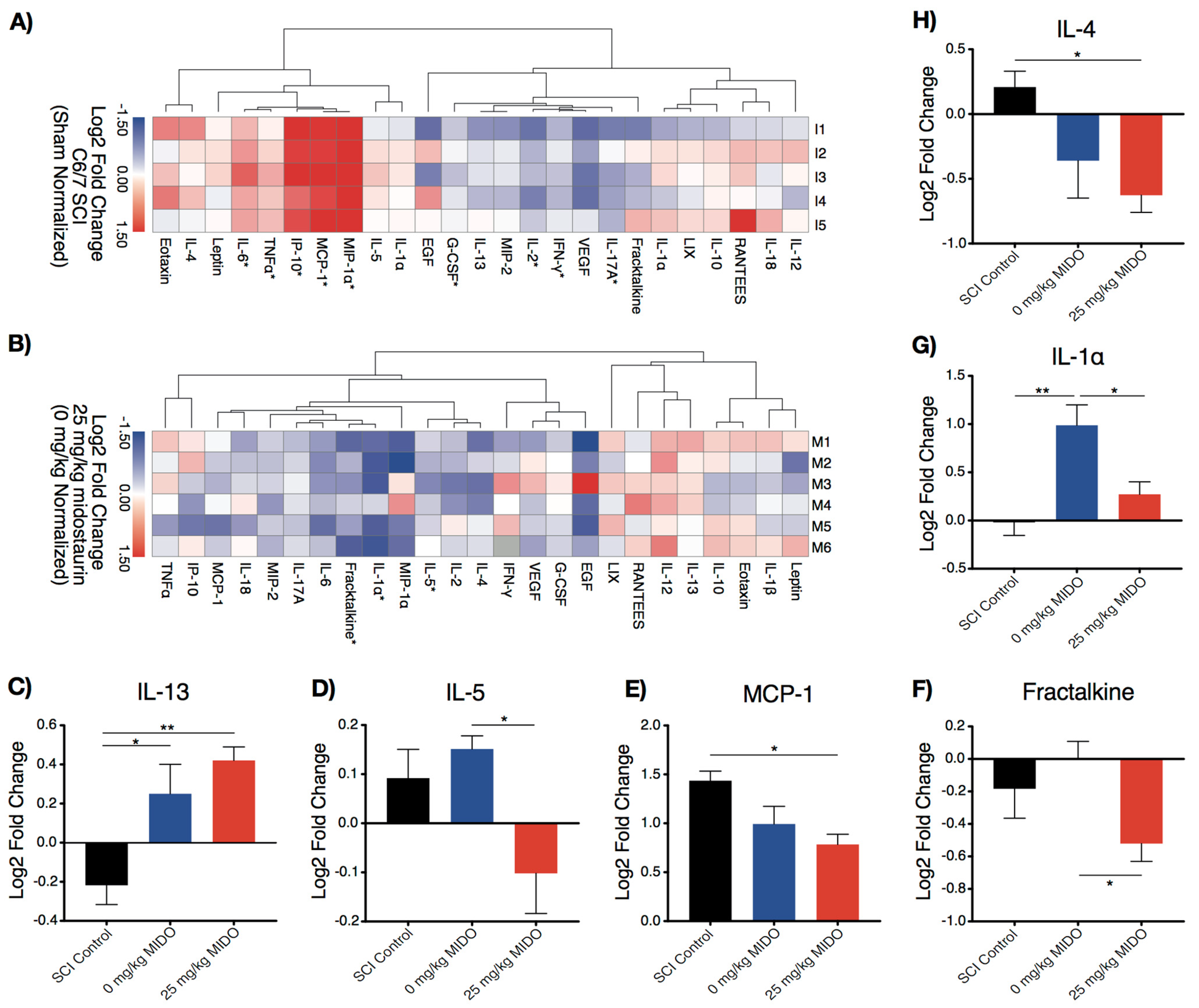
3.1. Midostaurin Alters the Local Inflammatory Response
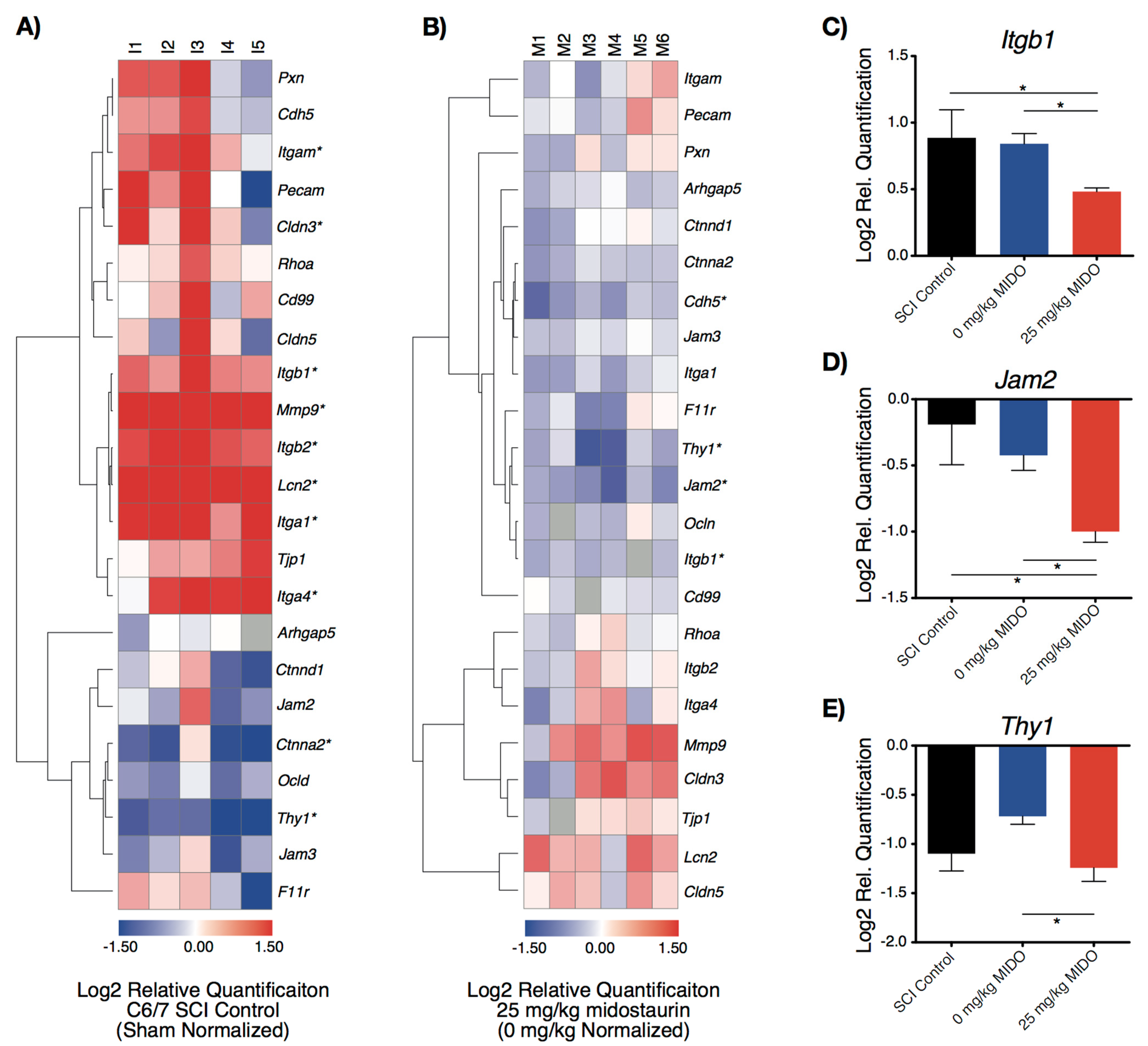
3.2. Midostaurin Reduces the Expression of Trans-Endothelial Migratory Genes
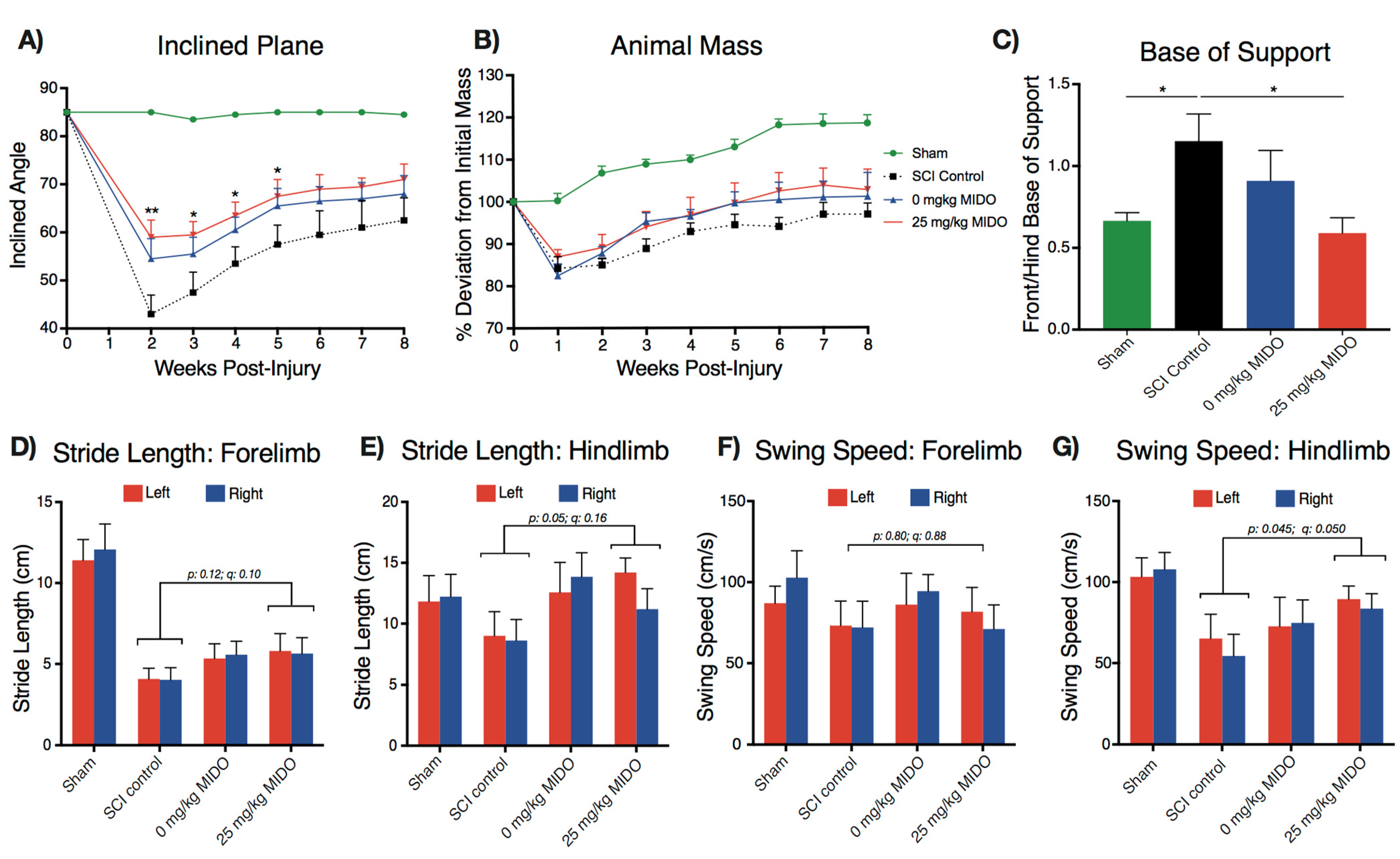
3.3. Midostaurin and DMSO Act Synergistically to Improve Functional Recovery
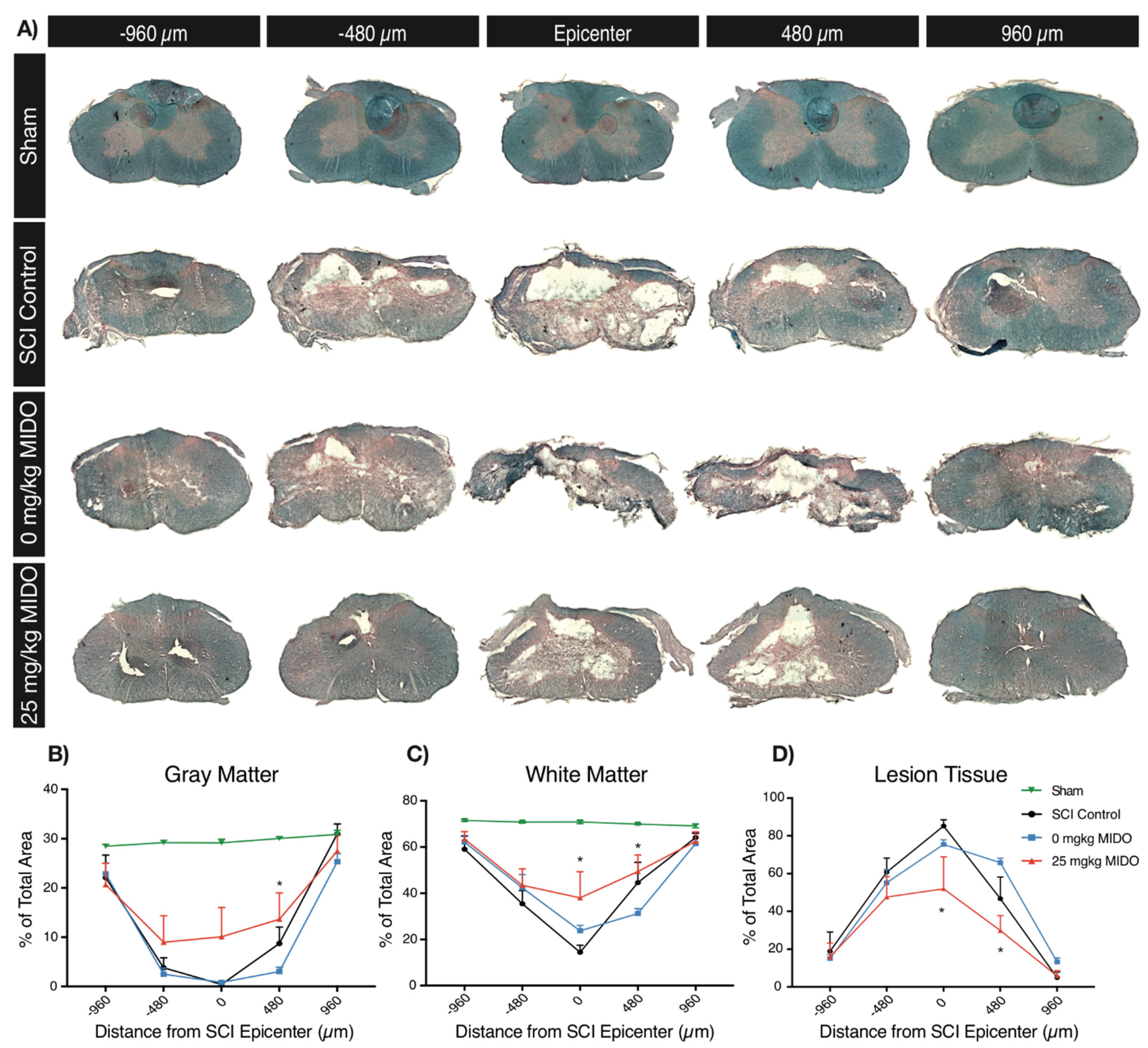
3.4. Histomorphometric Analysis Reveals Improved Tissue Preservation
3.5. Immunohistological Examinations Reveal Restored GAD67 Level
4. Discussion
4.1. Early Molecular Response to Midostaurin
4.2. Neuroprotective Effects of DMSO
4.3. Modulatory Impact of Neuroprotective Treatments
4.4. Staurosporine-Derived Inhibitors as Therapeutics for Traumatic SCI
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ahuja, C.S.; Wilson, J.R.; Nori, S.; Kotter, M.R.N.; Druschel, C.; Curt, A.; Fehlings, M.G. Traumatic Spinal Cord Injury. Nat. Rev. Dis. Primers 2017, 3, 17018. [Google Scholar] [CrossRef]
- James, S.L.; Theadom, A.; Ellenbogen, R.G.; Bannick, M.S.; Montjoy-Venning, W.; Lucchesi, L.R.; Abbasi, N.; Abdulkader, R.; Abraha, H.N.; Adsuar, J.C.; et al. Global, Regional, and National Burden of Traumatic Brain Injury and Spinal Cord Injury, 1990–2016: A Systematic Analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 2019, 18, 56–87. [Google Scholar] [CrossRef] [Green Version]
- Badhiwala, J.H.; Wilson, J.R.; Fehlings, M.G. Global Burden of Traumatic Brain and Spinal Cord Injury. Lancet Neurol. 2019, 18, 24–25. [Google Scholar] [CrossRef] [Green Version]
- Zavvarian, M.-M.; Toossi, A.; Khazaei, M.; Hong, J.; Fehlings, M. Novel Innovations in Cell and Gene Therapies for Spinal Cord Injury. F1000Res 2020, 9, 279. [Google Scholar] [CrossRef]
- James, N.D.; McMahon, S.B.; Field-Fote, E.C.; Bradbury, E.J. Neuromodulation in the Restoration of Function after Spinal Cord Injury. Lancet Neurol. 2018, 17, 905–917. [Google Scholar] [CrossRef] [Green Version]
- Fleming, J.C.; Norenberg, M.D.; Ramsay, D.A.; Dekaban, G.A.; Marcillo, A.E.; Saenz, A.D.; Pasquale-Styles, M.; Dietrich, W.D.; Weaver, L.C. The Cellular Inflammatory Response in Human Spinal Cords after Injury. Brain 2006, 129, 3249–3269. [Google Scholar] [CrossRef]
- Tator, C.; Fehlings, M.G. Review of the Secondary Injury Theory of Acute Spinal Cord Trauma with Emphasis on Vascular Mechanisms. J. Neurosurg. 1991, 75, 15–26. [Google Scholar] [CrossRef]
- Sofroniew, M.V. Dissecting Spinal Cord Regeneration. Nature 2018, 557, 343–350. [Google Scholar] [CrossRef] [PubMed]
- O’Shea, T.M.; Burda, J.E.; Sofroniew, M.V. Cell Biology of Spinal Cord Injury and Repair. J. Clin. Investig. 2017, 127, 3259–3270. [Google Scholar] [CrossRef] [PubMed]
- Zavvarian, M.-M.; Hong, J.; Fehlings, M.G. The Functional Role of Spinal Interneurons Following Traumatic Spinal Cord Injury. Front. Cell. Neurosci. 2020, 14, 127. [Google Scholar] [CrossRef] [PubMed]
- Chio, J.C.T.; Wang, J.; Badner, A.; Hong, J.; Surendran, V.; Fehlings, M.G. The Effects of Human Immunoglobulin G on Enhancing Tissue Protection and Neurobehavioral Recovery after Traumatic Cervical Spinal Cord Injury Are Mediated through the Neurovascular Unit. J. Neuroinflamm. 2019, 16, 141. [Google Scholar] [CrossRef] [PubMed]
- Kwon, B.K.; Okon, E.; Hillyer, J.; Mann, C.; Baptiste, D.; Weaver, L.C.; Fehlings, M.G.; Tetzlaff, W. A Systematic Review of Non-Invasive Pharmacologic Neuroprotective Treatments for Acute Spinal Cord Injury. J. Neurotrauma 2011, 28, 1545–1588. [Google Scholar] [CrossRef]
- Fehlings, M.G.; Wilson, J.R.; Harrop, J.S.; Kwon, B.K.; Tetreault, L.A.; Arnold, P.M.; Singh, J.M.; Hawryluk, G.; Dettori, J.R. Efficacy and Safety of Methylprednisolone Sodium Succinate in Acute Spinal Cord Injury: A Systematic Review. Glob. Spine J. 2017, 7, 116S–137S. [Google Scholar] [CrossRef]
- Hurlbert, R.J.; Hadley, M.N.; Walters, B.C.; Aarabi, B.; Dhall, S.S.; Gelb, D.E.; Rozzelle, C.J.; Ryken, T.C.; Theodore, N. Pharmacological Therapy for Acute Spinal Cord Injury. Neurosurgery 2013, 72, 93–105. [Google Scholar] [CrossRef] [Green Version]
- Fehlings, M.G.; Wilson, J.R.; Cho, N. Methylprednisolone for the Treatment of Acute Spinal Cord Injury: Counterpoint. Neurosurgery 2014, 61, 36–42. [Google Scholar] [CrossRef] [PubMed]
- McClendon, C.L.; Kornev, A.P.; Gilson, M.K.; Taylor, S.S. Dynamic Architecture of a Protein Kinase. Proc. Natl. Acad. Sci. USA 2014, 111, E4623–E4631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manning, G.; Whyte, D.B.; Martinez, R.; Hunter, T.; Sudarsanam, S. The Protein Kinase Complement of the Human Genome. Science 2002, 298, 1912–1934. [Google Scholar] [CrossRef] [Green Version]
- Zarrin, A.A.; Bao, K.; Lupardus, P.; Vucic, D. Kinase Inhibition in Autoimmunity and Inflammation. Nat. Rev. Drug Discov. 2020. [Google Scholar] [CrossRef]
- Schmidlin, T.; Debets, D.O.; van Gelder, C.A.G.H.; Stecker, K.E.; Rontogianni, S.; van den Eshof, B.L.; Kemper, K.; Lips, E.H.; van den Biggelaar, M.; Peeper, D.S.; et al. High-Throughput Assessment of Kinome-Wide Activation States. Cell Syst. 2019, 9, 366–374.e5. [Google Scholar] [CrossRef] [Green Version]
- Kasuya, Y.; Umezawa, H.; Hatano, M. Stress-Activated Protein Kinases in Spinal Cord Injury: Focus on Roles of P38. Int. J. Mol. Sci. 2018, 19, 867. [Google Scholar] [CrossRef] [Green Version]
- Hara, M.; Takayasu, M.; Watanabe, K.; Noda, A.; Takagi, T.; Suzuki, Y.; Yoshida, J. Protein Kinase Inhibition by Fasudil Hydrochloride Promotes Neurological Recovery after Spinal Cord Injury in Rats. J. Neurosurg. Spine 2000, 93, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Sivasankaran, R.; Pei, J.; Wang, K.C.; Zhang, Y.P.; Shields, C.B.; Xu, X.-M.; He, Z. PKC Mediates Inhibitory Effects of Myelin and Chondroitin Sulfate Proteoglycans on Axonal Regeneration. Nat. Neurosci. 2004, 7, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.-S.; Ryu, J.-H.; Zuo, Z.; Yang, S.-M.; Chang, H.-W.; Do, S.-H. Riluzole Attenuates Excitatory Amino Acid Transporter Type 3 Activity in Xenopus Oocytes via Protein Kinase C Inhibition. Eur. J. Pharmacol. 2013, 713, 39–43. [Google Scholar] [CrossRef]
- Noh, K.-M.; Hwang, J.-Y.; Shin, H.-C.; Koh, J.-Y. A Novel Neuroprotective Mechanism of Riluzole: Direct Inhibition of Protein Kinase, C. Neurobiol. Dis. 2000, 7, 375–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagoshi, N.; Nakashima, H.; Fehlings, M. Riluzole as a Neuroprotective Drug for Spinal Cord Injury: From Bench to Bedside. Molecules 2015, 20, 7775–7789. [Google Scholar] [CrossRef]
- Tetreault, L.A.; Zhu, M.P.; Wilson, J.R.; Karadimas, S.K.; Fehlings, M.G. The Impact of Riluzole on Neurobehavioral Outcomes in Preclinical Models of Traumatic and Nontraumatic Spinal Cord Injury: Results From a Systematic Review of the Literature. Glob. Spine J. 2020, 10, 216–229. [Google Scholar] [CrossRef] [PubMed]
- Lazarus, H.; Miller, M. Midostaurin: An Emerging Treatment for Acute Myeloid Leukemia Patients. J. Blood Med. 2016, 73. [Google Scholar] [CrossRef] [Green Version]
- Stone, R.M.; Mandrekar, S.J.; Sanford, B.L.; Laumann, K.; Geyer, S.; Bloomfield, C.D.; Thiede, C.; Prior, T.W.; Döhner, K.; Marcucci, G.; et al. Midostaurin plus Chemotherapy for Acute Myeloid Leukemia with a FLT3 Mutation. N. Engl. J. Med. 2017, 377, 454–464. [Google Scholar] [CrossRef]
- Gotlib, J.; Kluin-Nelemans, H.C.; George, T.I.; Akin, C.; Sotlar, K.; Hermine, O.; Awan, F.T.; Hexner, E.; Mauro, M.J.; Sternberg, D.W.; et al. Efficacy and Safety of Midostaurin in Advanced Systemic Mastocytosis. N. Engl. J. Med. 2016, 374, 2530–2541. [Google Scholar] [CrossRef]
- Kasamon, Y.L.; Ko, C.; Subramaniam, S.; Ma, L.; Yang, Y.; Nie, L.; Shord, S.; Przepiorka, D.; Farrell, A.T.; McKee, A.E.; et al. FDA Approval Summary: Midostaurin for the Treatment of Advanced Systemic Mastocytosis. Oncologist 2018, 23, 1511–1519. [Google Scholar] [CrossRef] [Green Version]
- Levis, M. Midostaurin Approved for FLT3-Mutated AML. Blood 2017, 129, 3403–3406. [Google Scholar] [CrossRef]
- Stansfield, L.C.; Pollyea, D.A. Midostaurin: A New Oral Agent Targeting FMS-Like Tyrosine Kinase 3-Mutant Acute Myeloid Leukemia. Pharmacotherapy 2017, 37, 1586–1599. [Google Scholar] [CrossRef]
- Luskin, M.R.; DeAngelo, D.J. Midostaurin/PKC412 for the Treatment of Newly Diagnosed FLT3 Mutation-Positive Acute Myeloid Leukemia. Expert Rev. Hematol. 2017, 10, 1033–1045. [Google Scholar] [CrossRef]
- Manley, P.W.; Caravatti, G.; Furet, P.; Roesel, J.; Tran, P.; Wagner, T.; Wartmann, M. Comparison of the Kinase Profile of Midostaurin (Rydapt) with That of Its Predominant Metabolites and the Potential Relevance of Some Newly Identified Targets to Leukemia Therapy. Biochemistry 2018, 57, 5576–5590. [Google Scholar] [CrossRef]
- Dutreix, C.; Lorenzo, S.; Wang, Y. Comparison of Two Endogenous Biomarkers of CYP3A4 Activity in a Drug–Drug Interaction Study between Midostaurin and Rifampicin. Eur. J. Clin. Pharm. 2014, 70, 915–920. [Google Scholar] [CrossRef] [Green Version]
- Dutreix, C.; Munarini, F.; Lorenzo, S.; Roesel, J.; Wang, Y. Investigation into CYP3A4-Mediated Drug–Drug Interactions on Midostaurin in Healthy Volunteers. Cancer Chemother. Pharm. 2013, 72, 1223–1234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Growney, J.D.; Clark, J.J.; Adelsperger, J.; Stone, R.; Fabbro, D.; Griffin, J.D.; Gilliland, D.G. Activation Mutations of Human C-KIT Resistant to Imatinib Mesylate Are Sensitive to the Tyrosine Kinase Inhibitor PKC412. Blood 2005, 106, 721–724. [Google Scholar] [CrossRef]
- Peter, B.; Bibi, S.; Eisenwort, G.; Wingelhofer, B.; Berger, D.; Stefanzl, G.; Blatt, K.; Herrmann, H.; Hadzijusufovic, E.; Hoermann, G.; et al. Drug-Induced Inhibition of Phosphorylation of STAT5 Overrides Drug Resistance in Neoplastic Mast Cells. Leukemia 2018, 32, 1016–1022. [Google Scholar] [CrossRef] [PubMed]
- Gani, O.A.B.S.M.; Engh, R.A. Protein Kinase Inhibition of Clinically Important Staurosporine Analogues. Nat. Prod. Rep. 2010, 27, 489. [Google Scholar] [CrossRef] [PubMed]
- Bourhill, T.; Narendran, A.; Johnston, R.N. Enzastaurin: A Lesson in Drug Development. Crit. Rev. Oncol. Hematol. 2017, 112, 72–79. [Google Scholar] [CrossRef]
- Orchard, R.C.; Wilen, C.B.; Virgin, H.W. Sphingolipid Biosynthesis Induces a Conformational Change in the Murine Norovirus Receptor and Facilitates Viral Infection. Nat. Microbiol. 2018, 3, 1109–1114. [Google Scholar] [CrossRef]
- Lorenz, L.; Axnick, J.; Buschmann, T.; Henning, C.; Urner, S.; Fang, S.; Nurmi, H.; Eichhorst, N.; Holtmeier, R.; Bódis, K.; et al. Mechanosensing by Β1 Integrin Induces Angiocrine Signals for Liver Growth and Survival. Nature 2018, 562, 128–132. [Google Scholar] [CrossRef]
- Tin, A.; Li, Y.; Brody, J.A.; Nutile, T.; Chu, A.Y.; Huffman, J.E.; Yang, Q.; Chen, M.-H.; Robinson-Cohen, C.; Macé, A.; et al. Large-Scale Whole-Exome Sequencing Association Studies Identify Rare Functional Variants Influencing Serum Urate Levels. Nat. Commun. 2018, 9, 4228. [Google Scholar] [CrossRef] [Green Version]
- Lessard, S.J.; MacDonald, T.L.; Pathak, P.; Han, M.S.; Coffey, V.G.; Edge, J.; Rivas, D.A.; Hirshman, M.F.; Davis, R.J.; Goodyear, L.J. JNK Regulates Muscle Remodeling via Myostatin/SMAD Inhibition. Nat. Commun. 2018, 9, 3030. [Google Scholar] [CrossRef]
- Bass, J.J.; Wilkinson, D.J.; Rankin, D.; Phillips, B.E.; Szewczyk, N.J.; Smith, K.; Atherton, P.J. An Overview of Technical Considerations for Western Blotting Applications to Physiological Research. Scand. J. Med. Sci. Sports 2017, 27, 4–25. [Google Scholar] [CrossRef]
- Rivlin, A.S.; Tator, C.H. Objective Clinical Assessment of Motor Function after Experimental Spinal Cord Injury in the Rat. J. Neurosurg. 1977, 47, 577–581. [Google Scholar] [CrossRef] [Green Version]
- Basso, D.M.; Beattie, M.S.; Bresnahan, J.C. A Sensitive and Reliable Locomotor Rating Scale for Open Field Testing in Rats. J. Neurotrauma 1995, 12, 1–21. [Google Scholar] [CrossRef]
- Soubeyrand, M.; Badner, A.; Vawda, R.; Chung, Y.S.; Fehlings, M.G. Very High Resolution Ultrasound Imaging for Real-Time Quantitative Visualization of Vascular Disruption after Spinal Cord Injury. J. Neurotrauma 2014, 31, 1767–1775. [Google Scholar] [CrossRef] [Green Version]
- Chio, J.C.T.; Wang, J.; Surendran, V.; Li, L.; Zavvarian, M.-M.; Pieczonka, K.; Fehlings, M.G. Delayed Administration of High Dose Human Immunoglobulin G Enhances Recovery after Traumatic Cervical Spinal Cord Injury by Modulation of Neuroinflammation and Protection of the Blood Spinal Cord Barrier. Neurobiol. Dis. 2021, 148, 105187. [Google Scholar] [CrossRef]
- Gawriluk, T.R.; Simkin, J.; Thompson, K.L.; Biswas, S.K.; Clare-Salzler, Z.; Kimani, J.M.; Kiama, S.G.; Smith, J.J.; Ezenwa, V.O.; Seifert, A.W. Comparative Analysis of Ear-Hole Closure Identifies Epimorphic Regeneration as a Discrete Trait in Mammals. Nat. Commun. 2016, 7, 11164. [Google Scholar] [CrossRef]
- Wilcox, J.T.; Satkunendrarajah, K.; Nasirzadeh, Y.; Laliberte, A.M.; Lip, A.; Cadotte, D.W.; Foltz, W.D.; Fehlings, M.G. Generating Level-Dependent Models of Cervical and Thoracic Spinal Cord Injury: Exploring the Interplay of Neuroanatomy, Physiology, and Function. Neurobiol. Dis. 2017, 105, 194–212. [Google Scholar] [CrossRef] [PubMed]
- Badner, A.; Hacker, J.; Hong, J.; Mikhail, M.; Vawda, R.; Fehlings, M.G. Splenic Involvement in Umbilical Cord Matrix-Derived Mesenchymal Stromal Cell-Mediated Effects Following Traumatic Spinal Cord Injury. J. Neuroinflamm. 2018, 15, 219. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.-C.; Shieh, H.-R.; Chen, Y.-J. Midostaurin (PKC412) Modulates Differentiation and Maturation of Human Myeloid Dendritic Cells. Toxicol. Vitr. 2010, 24, 1705–1710. [Google Scholar] [CrossRef]
- Gutierrez, L.; Jang, M.; Zhang, T.; Akhtari, M.; Alachkar, H. Midostaurin Reduces Regulatory T Cells Markers in Acute Myeloid Leukemia. Sci. Rep. 2018, 8, 17544. [Google Scholar] [CrossRef] [PubMed]
- Badner, A.; Vawda, R.; Laliberte, A.; Hong, J.; Mikhail, M.; Jose, A.; Dragas, R.; Fehlings, M. Early Intravenous Delivery of Human Brain Stromal Cells Modulates Systemic Inflammation and Leads to Vasoprotection in Traumatic Spinal Cord Injury: Intravenous Human Brain Stromal Cells in SCI. STEM CELLS Transl. Med. 2016, 5, 991–1003. [Google Scholar] [CrossRef]
- Figley, S.A.; Khosravi, R.; Legasto, J.M.; Tseng, Y.-F.; Fehlings, M.G. Characterization of Vascular Disruption and Blood–Spinal Cord Barrier Permeability Following Traumatic Spinal Cord Injury. J. Neurotrauma 2014, 31, 541–552. [Google Scholar] [CrossRef] [Green Version]
- Cormier, K.W.; Woodgett, J.R. Recent Advances in Understanding the Cellular Roles of GSK-3. F1000Res 2017, 6, 167. [Google Scholar] [CrossRef] [Green Version]
- Jaworski, T.; Banach-Kasper, E.; Gralec, K. GSK3 at the Intersection of Neuronal Plasticity and Neurodegeneration. Neural Plast. 2019, 2019, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Hur, E.-M.; Zhou, F.-Q. GSK3 Signalling in Neural Development. Nat. Rev. Neurosci. 2010, 11, 539–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eldar-Finkelman, H.; Martinez, A. GSK-3 Inhibitors: Preclinical and Clinical Focus on CNS. Front. Mol. Neurosci. 2011, 4. [Google Scholar] [CrossRef] [Green Version]
- Mantovani, A.; Sica, A.; Sozzani, S.; Allavena, P.; Vecchi, A.; Locati, M. The Chemokine System in Diverse Forms of Macrophage Activation and Polarization. Trends Immunol. 2004, 25, 677–686. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, R.; Tanaka, M.; Fukunaga, M.; Tateishi, T.; Kikuchi, H.; Motomura, K.; Matsushita, T.; Ohyagi, Y.; Kira, J. Restoration of Microglial Function by Granulocyte-Colony Stimulating Factor in ALS Model Mice. J. Neuroimmunol. 2010, 229, 51–62. [Google Scholar] [CrossRef]
- Wang, S.; Smith, G.M.; Selzer, M.E.; Li, S. Emerging Molecular Therapeutic Targets for Spinal Cord Injury. Expert Opin. Ther. Targets 2019, 23, 787–803. [Google Scholar] [CrossRef] [PubMed]
- Wanner, I.B.; Anderson, M.A.; Song, B.; Levine, J.; Fernandez, A.; Gray-Thompson, Z.; Ao, Y.; Sofroniew, M.V. Glial Scar Borders Are Formed by Newly Proliferated, Elongated Astrocytes That Interact to Corral Inflammatory and Fibrotic Cells via STAT3-Dependent Mechanisms after Spinal Cord Injury. J. Neurosci. 2013, 33, 12870–12886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ley, K.; Laudanna, C.; Cybulsky, M.I.; Nourshargh, S. Getting to the Site of Inflammation: The Leukocyte Adhesion Cascade Updated. Nat. Rev. Immunol. 2007, 7, 678–689. [Google Scholar] [CrossRef]
- Vestweber, D. How Leukocytes Cross the Vascular Endothelium. Nat. Rev. Immunol. 2015, 15, 692–704. [Google Scholar] [CrossRef]
- Weber, C.; Fraemohs, L.; Dejana, E. The Role of Junctional Adhesion Molecules in Vascular Inflammation. Nat. Rev. Immunol 2007, 7, 467–477. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, H. Integrin Signalling and Function in Immune Cells. Immunology 2012, 135, 268–275. [Google Scholar] [CrossRef] [Green Version]
- Leyton, L.; Díaz, J.; Martínez, S.; Palacios, E.; Pérez, L.A.; Pérez, R.D. Thy-1/CD90 a Bidirectional and Lateral Signaling Scaffold. Front. Cell Dev. Biol. 2019, 7, 132. [Google Scholar] [CrossRef]
- Vanlandewijck, M.; He, L.; Mäe, M.A.; Andrae, J.; Ando, K.; Del Gaudio, F.; Nahar, K.; Lebouvier, T.; Laviña, B.; Gouveia, L.; et al. A Molecular Atlas of Cell Types and Zonation in the Brain Vasculature. Nature 2018, 554, 475–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, J.; Chang, A.; Liu, Y.; Wang, J.; Fehlings, M.G. Incomplete Spinal Cord Injury Reverses the Level-Dependence of Spinal Cord Injury Immune Deficiency Syndrome. Int. J. Mol. Sci. 2019, 20, 3762. [Google Scholar] [CrossRef] [Green Version]
- Hong, J.; Chang, A.; Zavvarian, M.-M.; Wang, J.; Liu, Y.; Fehlings, M. Level-Specific Differences in Systemic Expression of Pro- and Anti-Inflammatory Cytokines and Chemokines after Spinal Cord Injury. Int. J. Mol. Sci. 2018, 19, 2167. [Google Scholar] [CrossRef] [Green Version]
- Poniatowski, Ł.A.; Wojdasiewicz, P.; Krawczyk, M.; Szukiewicz, D.; Gasik, R.; Kubaszewski, Ł.; Kurkowska-Jastrzębska, I. Analysis of the Role of CX3CL1 (Fractalkine) and Its Receptor CX3CR1 in Traumatic Brain and Spinal Cord Injury: Insight into Recent Advances in Actions of Neurochemokine Agents. Mol. Neurobiol. 2017, 54, 2167–2188. [Google Scholar] [CrossRef] [Green Version]
- Freria, C.M.; Hall, J.C.E.; Wei, P.; Guan, Z.; McTigue, D.M.; Popovich, P.G. Deletion of the Fractalkine Receptor, CX3CR1, Improves Endogenous Repair, Axon Sprouting, and Synaptogenesis after Spinal Cord Injury in Mice. J. Neurosci. 2017, 37, 3568–3587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatori, K.; Nagai, A.; Heisel, R.; Ryu, J.K.; Kim, S.U. Fractalkine and Fractalkine Receptors in Human Neurons and Glial Cells. J. Neurosci. Res. 2002, 69, 418–426. [Google Scholar] [CrossRef] [PubMed]
- Sato, A.; Ohtaki, H.; Tsumuraya, T.; Song, D.; Ohara, K.; Asano, M.; Iwakura, Y.; Atsumi, T.; Shioda, S. Interleukin-1 Participates in the Classical and Alternative Activation of Microglia/Macrophages after Spinal Cord Injury. J. Neuroinflamm. 2012, 9, 553. [Google Scholar] [CrossRef] [Green Version]
- Bastien, D.; Bellver Landete, V.; Lessard, M.; Vallieres, N.; Champagne, M.; Takashima, A.; Tremblay, M.-E.; Doyon, Y.; Lacroix, S. IL-1 Gene Deletion Protects Oligodendrocytes after Spinal Cord Injury through Upregulation of the Survival Factor Tox3. J. Neurosci. 2015, 35, 10715–10730. [Google Scholar] [CrossRef] [Green Version]
- Yates, A.G.; Jogia, T.; Gillespie, E.R.; Couch, Y.; Ruitenberg, M.J.; Anthony, D.C. Acute IL-1RA Treatment Suppresses the Peripheral and Central Inflammatory Response to Spinal Cord Injury. J. NeuroInflamm. 2021, 18, 15. [Google Scholar] [CrossRef]
- Greenfeder, S.; Umland, S.P.; Cuss, F.M.; Chapman, R.W.; Egan, R.W. Th2 Cytokines and Asthma The Role of Interleukin-5 in Allergic Eosinophilic Disease. Respir. Res. 2001, 2, 1–9. [Google Scholar] [CrossRef]
- Roufosse, F. Targeting the Interleukin-5 Pathway for Treatment of Eosinophilic Conditions Other than Asthma. Front. Med. 2018, 5, 49. [Google Scholar] [CrossRef] [Green Version]
- Rouleau, P.; Ung, R.-V.; Lapointe, N.P.; Guertin, P.A. Hormonal And Immunological Changes in Mice after Spinal Cord Injury. J. Neurotrauma 2007, 24, 367–378. [Google Scholar] [CrossRef]
- Ma, M.; Wei, T.; Boring, L.; Charo, I.F.; Ransohoff, R.M.; Jakeman, L.B. Monocyte Recruitment and Myelin Removal Are Delayed Following Spinal Cord Injury in Mice with CCR2 Chemokine Receptor Deletion. J. Neurosci. Res. 2002, 68, 691–702. [Google Scholar] [CrossRef]
- Chomarat, P.; Banchereau, J. Interleukin-4 and Lnterleukin-13: Their Similarities and Discrepancies. Int. Rev. Immunol. 1998, 17, 1–52. [Google Scholar] [CrossRef]
- Dooley, D.; Lemmens, E.; Vangansewinkel, T.; Le Blon, D.; Hoornaert, C.; Ponsaerts, P.; Hendrix, S. Cell-Based Delivery of Interleukin-13 Directs Alternative Activation of Macrophages Resulting in Improved Functional Outcome after Spinal Cord Injury. Stem Cell Rep. 2016, 7, 1099–1115. [Google Scholar] [CrossRef]
- Lima, R.; Monteiro, S.; Lopes, J.; Barradas, P.; Vasconcelos, N.; Gomes, E.; Assunção-Silva, R.; Teixeira, F.; Morais, M.; Sousa, N.; et al. Systemic Interleukin-4 Administration after Spinal Cord Injury Modulates Inflammation and Promotes Neuroprotection. Pharmaceuticals 2017, 10, 83. [Google Scholar] [CrossRef] [Green Version]
- Broadwell, R.; Salcman, M.; Kaplan, R. Morphologic Effect of Dimethyl Sulfoxide on the Blood-Brain Barrier. Science 1982, 217, 164–166. [Google Scholar] [CrossRef] [PubMed]
- Turan, N.N.; Akar, F.; Budak, B.; Seren, M.; Parlar, A.İ.; Sürücü, S.; Ulus, A.T. How DMSO, a Widely Used Solvent, Affects Spinal Cord Injury. Ann. Vasc. Surg. 2008, 22, 98–105. [Google Scholar] [CrossRef]
- Jacob, S.W.; de la Torre, J.C. Pharmacology of Dimethyl Sulfoxide in Cardiac and CNS Damage. Pharmacol. Rep. 2009, 61, 225–235. [Google Scholar] [CrossRef]
- Tillakaratne, N.J.K.; Mouria, M.; Ziv, N.B.; Roy, R.R.; Edgerton, V.R.; Tobin, A.J. Increased Expression of Glutamate Decarboxylase (GAD67) in Feline Lumbar Spinal Cord After Complete Thoracic Spinal Cord Transection. J. Neurosci. Res. 2000, 60, 219–230. [Google Scholar] [CrossRef]
- Neumann, M.; Wang, Y.; Kim, S.; Hong, S.M.; Jeng, L.; Bilgen, M.; Liu, J. Assessing Gait Impairment Following Experimental Traumatic Brain Injury in Mice. J. Neurosci. Methods 2009, 176, 34–44. [Google Scholar] [CrossRef] [Green Version]
- Flynn, J.R.; Conn, V.L.; Boyle, K.A.; Hughes, D.I.; Watanabe, M.; Velasquez, T.; Goulding, M.D.; Callister, R.J.; Graham, B.A. Anatomical and Molecular Properties of Long Descending Propriospinal Neurons in Mice. Front. Neuroanat. 2017, 11. [Google Scholar] [CrossRef] [Green Version]
- Crump, M.; Leppä, S.; Fayad, L.; Lee, J.J.; Di Rocco, A.; Ogura, M.; Hagberg, H.; Schnell, F.; Rifkin, R.; Mackensen, A.; et al. Randomized, Double-Blind, Phase III Trial of Enzastaurin Versus Placebo in Patients Achieving Remission After First-Line Therapy for High-Risk Diffuse Large B-Cell Lymphoma. J. Clin. Oncol. 2016, 34, 2484–2492. [Google Scholar] [CrossRef]
- Wick, W.; Puduvalli, V.K.; Chamberlain, M.C.; van den Bent, M.J.; Carpentier, A.F.; Cher, L.M.; Mason, W.; Weller, M.; Hong, S.; Musib, L.; et al. Phase III Study of Enzastaurin Compared With Lomustine in the Treatment of Recurrent Intracranial Glioblastoma. J. Clin. Oncol. 2010, 28, 1168–1174. [Google Scholar] [CrossRef] [Green Version]
- Lanz, T.V.; Becker, S.; Osswald, M.; Bittner, S.; Schuhmann, M.K.; Opitz, C.A.; Gaikwad, S.; Wiestler, B.; Litzenburger, U.M.; Sahm, F.; et al. Protein Kinase Cβ as a Therapeutic Target Stabilizing Blood–Brain Barrier Disruption in Experimental Autoimmune Encephalomyelitis. Proc. Natl. Acad. Sci. USA 2013, 110, 14735–14740. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zavvarian, M.-M.; Hong, J.; Khazaei, M.; Chio, J.C.T.; Wang, J.; Badner, A.; Fehlings, M.G. The Protein Kinase Inhibitor Midostaurin Improves Functional Neurological Recovery and Attenuates Inflammatory Changes Following Traumatic Cervical Spinal Cord Injury. Biomolecules 2021, 11, 972. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11070972
Zavvarian M-M, Hong J, Khazaei M, Chio JCT, Wang J, Badner A, Fehlings MG. The Protein Kinase Inhibitor Midostaurin Improves Functional Neurological Recovery and Attenuates Inflammatory Changes Following Traumatic Cervical Spinal Cord Injury. Biomolecules. 2021; 11(7):972. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11070972
Chicago/Turabian StyleZavvarian, Mohammad-Masoud, James Hong, Mohamad Khazaei, Jonathon Chon Teng Chio, Jian Wang, Anna Badner, and Michael G. Fehlings. 2021. "The Protein Kinase Inhibitor Midostaurin Improves Functional Neurological Recovery and Attenuates Inflammatory Changes Following Traumatic Cervical Spinal Cord Injury" Biomolecules 11, no. 7: 972. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11070972