
Neutrophil in the Pancreatic Tumor Microenvironment


Department of Pathology & Clinical Labs, Rogel Cancer Center and Center for RNA Biomedicine, University of Michigan, Ann Arbor, MI 48109, USA
*
Author to whom correspondence should be addressed.
Biomolecules 2021, 11(8), 1170; https://0-doi-org.brum.beds.ac.uk/10.3390/biom11081170
Submission received: 27 June 2021
/
Revised: 30 July 2021
/
Accepted: 5 August 2021
/
Published: 7 August 2021
(This article belongs to the Collection Recent Advances in Pancreatic Cancer)
Abstract
:Pancreatic ductal adenocarcinoma (PDAC) is a malignancy with a poor prognosis and low survival rates. PDAC is characterized by a fibroinflammatory tumor microenvironment enriched by abundant fibroblasts and a variety of immune cells, contributing to its aggressiveness. Neutrophils are essential infiltrating immune cells in the PDAC microenvironment. Recent studies have identified several cellular mechanisms by which neutrophils are recruited to tumor lesion and promote tumorigenesis. This review summarizes the current understanding of the interplay between neutrophils, tumor cells, and other components in the PDAC tumor microenvironment. The prognosis and therapeutic implications of neutrophils in PDAC are also discussed.
1. Introduction
Pancreatic ductal adenocarcinoma (PDAC) is one of the most lethal cancers with a five-year survival rate of 10% and is predicted to be the second leading cause of cancer-related death by 2030 [1,2]. The poor prognosis is associated with delayed diagnosis and the failure to achieve a durable response to available treatment modalities. Recent studies have highlighted the importance of the tumor microenvironment in PDAC progression and immune evasion. The dense desmoplastic stroma in PDAC contains abundant cancer-associated fibroblasts (CAFs). Another critical feature of the PDAC immune environment is the relative void of cytotoxic lymphocytes (CTLs) in the tumor core [3]. In order to overcome the tumor escaping host immune surveillance, it is crucial to understand the interaction between tumor cells and their surroundings.
Neutrophils are the most abundant immune cells in circulation and form an essential part of the innate immune system to respond against infection and inflammatory insults. Recent studies discovered an early homogeneous neutrophil progenitor subset in human bone marrow defined by surface markers CD71 and CD117 [4]. These CD71+ neutrophils proliferate and expand in the blood and tumors from cancer patients. It has become increasingly clear that neutrophils make up a substantial proportion of the immune infiltrate in a wide variety of cancers, including PDAC [5], breast cancer [6], colorectal cancer [7], melanoma [8], renal cell carcinoma [9], hepatocellular carcinoma [10], and others [11,12,13]. Intratumor neutrophils and high IL-8 levels are associated with poor outcomes to immune checkpoint inhibitors and worse survival in patients with advanced cancers. However, the role of neutrophils in tumor development is still unclear. Some studies have proposed classifying tumor-associated neutrophils (TANs) into two polarization states, tumor-suppressing N1 neutrophils and tumor-promoting N2 neutrophils [14]. In the lung cancer model, TGF-β inhibition leads to the recruitment and activation of N1-type neutrophils, which can kill tumor cells and inhibit tumor development [14]. Interferon-β signaling has been shown to polarize neutrophils to N1 phenotype [15]. Additionally, pro-inflammatory or immunostimulatory cytokines, such as interleukin-12 (IL-12), CXCL9, CXCL10, and CCL3, are released from N1 neutrophils and facilitate recruitment and activation of CD8+ T cells [16]. On the other hand, exposure to TGF-β transforms neutrophils to the N2 phenotype [14]. N2 neutrophils have been reported to have strong immunosuppressive and tumor-promoting functions, including the promotion of tumor metastases and angiogenesis [17,18]. N1 neutrophils are characterized as CD101+, CD177+, CD170low, CD54+, HLA-DR+, CD86+, and CD15high, whereas N2 neutrophils are CD170high, CD117+, Lectin-like oxidized LDL receptor 1 (LOX1)+, CD84+, junctional adhesion molecule-like (JAML)+, and PD-L1+ [15]. However, this dichotomized classification of TANs may be overly simplified because, in some tumors, such as 3-MCA-induced primary sarcomas, TANs have a mixed phenotype between N1 and N2 [14,15]. Furthermore, neutrophils can be trained to reprogram their transcriptome and epigenetics and take on the anti-tumor effect [19]. In the context of PDAC, TANs are associated with poor prognoses [20].
One way the tumor evades the host immune system is through a group of heterogeneous immature myeloid cells, called myeloid-derived suppressor cells (MDSCs). MDSCs suppress immune responses by multiple signaling pathways. MDSCs can induce antigen-specific CD8+ T cell tolerance via generating reactive oxygen species (ROS) and peroxynitrite [21]. In addition, MDSCs suppress natural killer (NK) cell cytotoxicity by inhibiting the activation of Stat5 [22]. Markers for MDSCs include CD33 and CD11b in humans and Gr-1 and Cd11b in mice [23]. Based on phenotypic features, MDSCs can be characterized into two major types, monocytic MDSCs (M-MDSCs) and granulocytic MDSCs (G-MDSCs) [24]. Mouse M-MDSCs are Ly6G−Ly6 Chigh, and G-MDSCs are Ly6G+ Ly6 Clow [25]. In humans, CD66b and CD15 are markers for M-MDSCs and G-MDSCs [25]. However, it is believed that the MDSCs described in most studies are, in fact, a subset of neutrophils [26,27] due to a significant overlap in the expression of functional molecules or surface molecules between neutrophils and G-MDSC [28].
In this review, we will focus on recent discoveries in the interaction between tumor cells, neutrophils, and other stromal cells in the TME of PDAC. We will also discuss the clinical implications of neutrophils in PDAC.
2. Interactions between Neutrophil and Tumor
2.1. Tumor Cells Attract Neutrophils
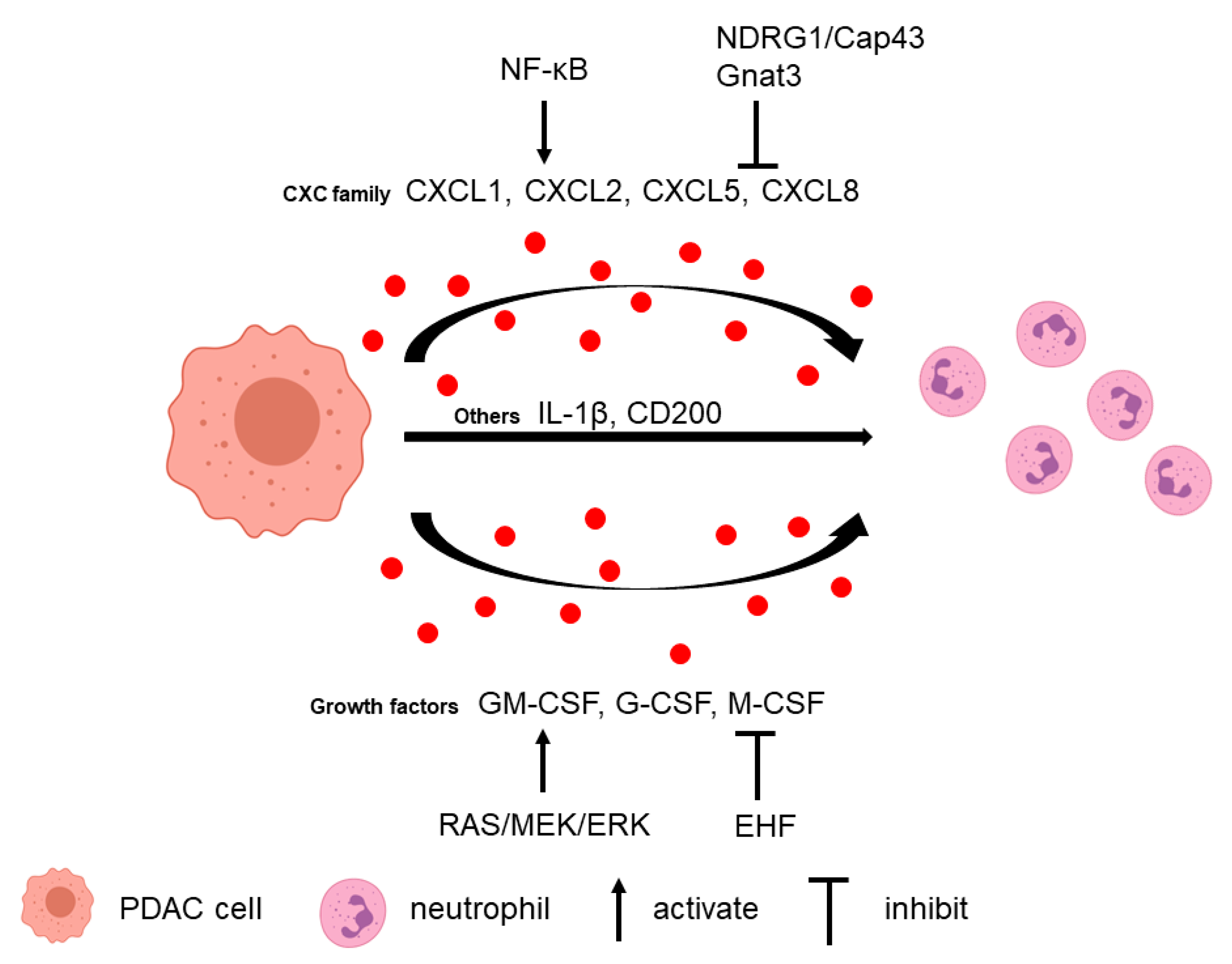
Neutrophils can be recruited to the PDAC microenvironment via multiple tumor-secreted factors (Figure 1). Several CXC families of chemokines have been attributed a major role in neutrophil recruitment, including CXCL1, CXCL2, CXCL5, and CXCL8 [29,30,31]. Neutrophils express CXC receptors CXCR1 and CXCR2 that can respond to the tumor-derived CXC family chemokines. CXCR1 binds to chemokines CXCL6 and CXCL8, and CXCR2 binds to chemokines CXCL1–3 and CXCL5–8 [32]. CXCR2 expression was upregulated and associated with tumor size in PDAC [33]. In Cxcr2 knockout PKF (LSL-KrasG12D/+; Tgfbr2flox/flox, Ptf1a-Cre) mice, infiltration of myeloperoxidase+ (MPO+) neutrophils and CD11b+Ly6G+ MDSCs to tumor is decreased compared to control animals [34]. In a KPC (LSL-KrasG12D/+; LSL-Trp53R172H/+; Pdx1-Cre) mouse model, the secretion of Cxcl1, Cxcl2, and Cxcl5 from tumor cells is increased and related to high numbers of MPO+ neutrophils compared to the normal pancreas [29]. Another study demonstrated that CXCL5 has the greatest fold increase in human PDAC and correlated with both tumor-infiltrating CD15+ granulocytes and neutrophil elastase+ (NE+) granulocytes [30]. Although there is limited literature on the specific role of the CXCL5/CXCR2 axis in regulating G-MDSC in PDAC, G-MDSC was shown to be positively correlated with CXCL5 level in human renal cell cancer, and blockage of CXCR2 reduced tumor weight with increased effector T cell infiltration in a renal cell cancer mouse model [35]. In a mouse melanoma model, CXCL5 is the primary chemokine to attract G-MDSCs to the tumor, which induced epithelial-mesenchymal transition, tumor cell migration, and metastasis [36]. In another gastric cancer model, MDSCs migration was partially attenuated with CXCR2 inhibitor, which suggested that the CXCL5/CXCR2 axis might play a role in MDSCs’ recruitment [37]. Furthermore, CXCL5 can recruit MDSCs to the tumor via CXCR2 and promote breast cancer progression [38].
In tumor cells, neutrophil-recruiting chemokine secretions can be regulated by several signaling pathways. For example, N-myc downstream regulated gene 1 (NDRG1)/Cap43 downregulates the expression of CXCL1, CXCL5, CXCL8, and VEGF in tumor cells, leading to the suppression of neutrophil infiltration [31]. In addition, ablation of α-gustducin (Gnat3) in KrasG12D-expressing pancreas increases the levels of different chemokines, including CXCL1 and CXCL2, which lead to G-MDSC recruitment and altered MDSC gene expression in early neoplasia [39]. NF-ĸB is another critical signaling pathway in which these chemokines are regulated. Knockdown of Vasohibin-2 (VASH2), an endothelium-derived angiogenesis inhibitor, in murine pancreatic cancer cells downregulates NF-ĸB signaling, which results in decreased infiltration of CD11b+ Ly6G+ G-MDSC mediated by low Cxcl2 and Cxcl5 [40]. Another study found that in response to KRAS/MEK inhibition, NF-ĸB activation induces CXCL5 secretion in pancreatic cancer cells, which elevated CD11b+Ly6G+ neutrophil infiltration [41].
Growth factors secreted by tumor cells also play a role in neutrophil recruitment. Granulocyte macrophage colony-stimulating factor (GM-CSF), granulocyte colony-stimulating factor (G-CSF), and monocyte colony-stimulating factor (M-CSF) are upregulated in PDAC cells, promoting the survival and recruitment of G-MDSC and CD11b+ Ly6G+ neutrophils in vivo [42]. G-CSF expression is upregulated by activating the RAS/MEK/ERK pathway in PDAC cells through the ETS transcription factor [43]. In addition, ETS homologous factor (EHF) directly suppresses the GM-CSF level by binding to its promoters. EHF loss in tumor cells relieves the transcriptional suppression of GM-CSF, which induces MDSCs conversion, expansion, and function [44].
Other cytokines and proteins derived from pancreatic cancer cells also influence neutrophil infiltration. Tumor cell-derived IL-1β contributes to neutrophil infiltration since depletion of tumor cell-derived IL-1β significantly decreases the stromal accumulation of CD11b+ Gr1+ MDSC and CD11b+ Ly6G+ TANs [45]. CD200 (OX-2; OX-90) from the epithelial PDAC cells and α-SMA+ stromal cells enhances CD200R+ MDSC expansion and increases their immunosuppressive activity on T-cell proliferation [46]. Tumor cell-intrinsic ubiquitin-specific protease 22 (USP22) regulates immune cell infiltration in implanted PDAC tumors [47]. USP22 ablation results in decreased total myeloid cells and G-MDSCs. Reg3g can promote pancreatic carcinogenesis, and its overexpression in tumor cells enhances CD11b+Gr1+ MDSC recruitment [48].
2.2. Neutrophils Promote Tumor Cell Survival and Metastasis
Multiple studies have demonstrated that tumor-infiltrating neutrophils have a pro-tumor role in PDAC. PDACs can be subtyped into squamous, aberrantly differentiated endocrine exocrine, pancreatic progenitor, and immunogenic subsets based on transcriptomic data [49]. PDACs can also be classified based on whether they have normal or activated stroma [50]. A study used gene set variation analysis (GSVA) to compare the enrichment of these PDAC subset gene signatures between TAN-high and TAN-med/low PDACs [41]. TAN-high PDACs were found to significantly enrich genes in the squamous and normal stroma subtypes, which had the poorest prognosis. Several studies in PDAC have found neutrophil depletion could inhibit tumor growth and metastasis. Depleting G-MDSC in the KPC mouse model using anti-Ly6G antibody, 1A8, resulted in an increase in cleaved caspase-3 (CC3)-positive tumor cells, which suggested that depletion of G-MDSC increased tumor epithelial apoptosis [42]. Another study used anti-Ly6G antibody 1A8 in KPC mice, which reduced the number of MPO+ cells infiltrating tumors. Although there was no difference in overall or tumor-free survival, depletion of neutrophils abrogated tumor metastasis [29]. In KPC cell-derived orthotopic tumors, anti-Ly6G treatment delayed tumor growth, and a combination of anti-Ly6G and anti-PD-1 treatment showed a synergistic anti-tumor effect [51]. Importantly, PDAC patient-derived neutrophils significantly promoted the migration and invasion of pancreatic cancer cells in wound healing assay and transwell assay in vitro, whereas neutrophils derived from normal individuals did not [52].
3. Interactions between Neutrophil and TME
3.1. T Cell
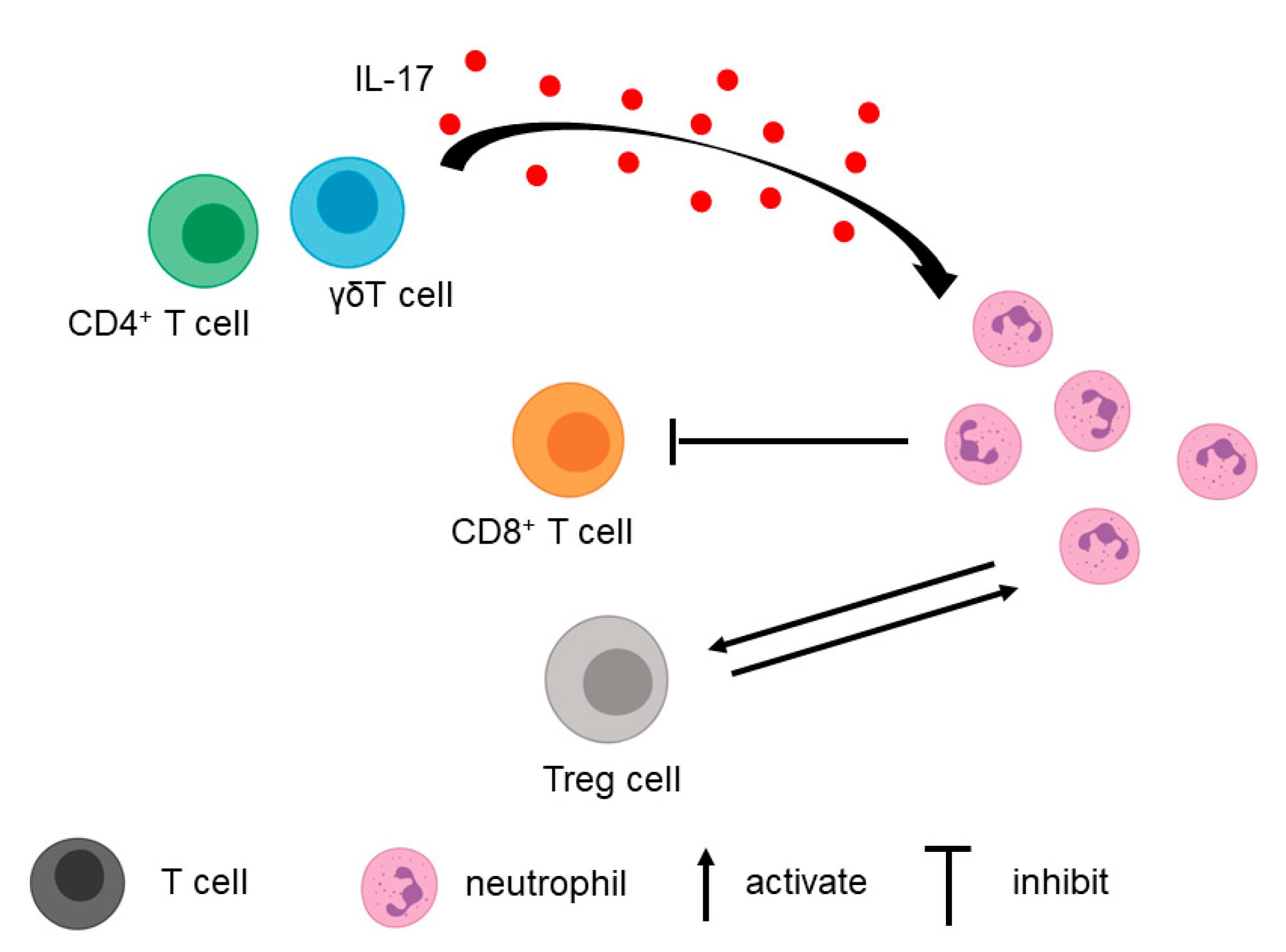
In addition to tumor cells, T cells in the TME contribute to the recruitment of neutrophils (Figure 2). A study by Zhang et al. found that IL-17, a cytokine secreted primarily by CD4+ and γδT cells, played a role in neutrophil recruitment [51]. When the IL-17 signaling pathway was blocked using anti-IL17/anti-IL17R neutralizing antibodies in the orthotopic pancreatic cancer mouse model, myeloid cell recruitment decreased, whereas the total number of CD8+ T cells and activated CD8+ T cells increased [51,53]. This finding shows that neutrophils and T cells can influence each other.
Different subsets of T cells can interact with TANs. For example, MDSCs can directly interact with Treg [54]. When MDSCs are depleted using anti-Gr-1 RB6–8C5 antibody, Foxp3+ Treg recruitment is inhibited in PDAC [54]. Furthermore, MDSCs are able to induce Treg cell proliferation in a cell–cell-dependent manner. Treg cells, in turn, can affect the survival and/or the proliferation of MDSCs [54].
Organoid/immune cell co-culture models demonstrated that G-MDSCs could inhibit CTL proliferation. Depletion of G-MDSCs using cabozantinib, a tyrosine kinase inhibitor known to deplete MDSCs, increased CTL proliferation and effector function, resulting in cancer cells’ sensitization to anti-PD-1/PD-L1 treatment [55]. G-MDSC (CD11b+ Gr-1high Ly6Cint) and M-MDSC (CD11b+ Gr-1int Ly6Chigh) isolated from KPC mice suppressed T cell proliferation and induced apoptosis of activated T cells in vitro. Depleting G-MDSC using anti-Ly6G antibody significantly increased the percentage and the absolute number of CD8+ T cells in KPC mouse tumors [42].
A different study found that MDSC isolated from bone marrow or KPC tumors inhibited proliferation and induced apoptosis of CD8+ T cells in the presence of dendritic cells (DC) presenting a high-affinity cognate peptide, even following T cell initial activation by DCs [56]. In addition, MDSCs could induce DNA damage and p53 pathway activation in CD8+ T cells through an iNOS-dependent pathway [56].
3.2. Fibroblasts, Macrophages, and Extracellular Matrix
Pancreatic stellate cells (PSCs) and CAFs are essential components in the pancreatic TME. These stromal cells also interact with immune cells, including neutrophils. One study demonstrated that PSCs induced MDSC expansion in the orthotopic PDAC model [57]. In the same study, co-transplantation of PSCs and tumor cells significantly increased G-MDSCs infiltration in the tumor [57]. In turn, neutrophil DNA could activate PSC and increase PSC proliferation, enhancing the production of the matrix metallopeptidase 2 (MMP2) and matrix metallopeptidase 9 (MMP9) secreted by PSCs through interaction with the receptor for advanced glycation end productions (RAGE) [58]. Another study showed infiltrated neutrophils in PDAC reprogramed the functional status of stellate cells, which included reduced collagen production and increased protease synthesis of PSCs [59].
Tumor-associated macrophages (TAMs) are also significant components of the microenvironment and have been demonstrated to be correlated with worse patient prognosis in PDAC [60]. In a preclinical model, targeting both neutrophils and macrophages enhanced chemotherapeutic efficacy and improved the survival of mice with orthotopic PDAC tumors [30]. In breast cancer, IL-10 from MDSCs reduced IL-12 production by macrophages, resulting in the polarization towards an M2 phenotype [61]. Further studies on the interaction between neutrophils and macrophages in PDAC would be beneficial.
Additionally, neutrophils can reprogram the extracellular matrix of PDAC. Targeted depletion of G-MDSC in the KPC mouse model decreases ECM deposition and the appearance of patent blood vessels, with increased vessel diameters [42]. Neutrophil-derived MMP9 has a direct and robust proangiogenic effect independent and additive to PDAC-derived VEGF [62].
4. Neutrophil Extracellular Trap in PDAC
The formation of neutrophil extracellular trap (NET) is a cellular function of neutrophils. Release of NET, called NETosis, is considered a defense mechanism to trap and kill bacteria and other pathogens. NETs consist of extracellular DNA and several proteins, including MPO and neutrophil elastase (NE) [63]. Studies across different cancers have suggested that NETs play an essential role in tumor progression and metastasis [64,65,66]. NETs induce the awakening of dormant cancer cells to form metastasis in lungs in mouse models of breast and prostate cancer [67]. The role of NETs in PDAC drew much attention as our understanding of neutrophils in tumors improved.
Many studies have demonstrated that pancreatic cancer cells induce NET formation. For example, the conditioned media from PDAC cell line AsPC-1 was able to induce NET formation, and NETs could promote AsPC-1 cell migration and invasion as well as angiogenesis in vitro [68]. Another group found that conditioned media from KPC cells previously exposed to IL-17 induced NETosis in mouse neutrophils in vitro [51]. Moreover, serum from PDAC patients significantly increased NET formation and reduced NET degradation [51]. Tumor-derived protein tissue inhibitor of metalloproteinases-1 (TIMP-1), which correlated with poor prognosis in PDAC, directly triggered the formation of NETs. This effect depended on the interaction of TIMP-1 with its receptor CD63 and subsequent ERK signaling [69]. In addition to tumor cells, CAF-conditioned media can also induce ROS-dependent NETosis by secreting Amyloid β A4 protein. In turn, NET formation promotes CAF expansion, contractility, and deposition of matrix components supportive of tumor growth.
Recent studies suggested that NETs contribute to PDAC development. Electron microscope images of co-cultured pancreatic cancer cells with neutrophils showed that NETs could capture pancreatic cancer cells by their spider web-like structures [70]. The same study further demonstrated that NET-derived HMGB1, which could be degraded by thrombomodulin, potentiated tumor aggressiveness by inducing epithelial-mesenchymal transition (EMT) [70]. In another study, when NET supernatant was introduced, pancreatic cancer cell proliferation increased in a dose-dependent manner [52]. This increase in proliferation was reversed when DNase was present. Similarly, conditioned media from activated neutrophils promoted migration, invasion, and EMT in PDAC cells via the IL-1β/EGFR/ERK pathway [52]. Protein Arginine Deiminase 4 (PAD4), which citrullinates histones, is critical in the NET release [71]. PAD4 inhibitor, GSK484, treatment reduced NET formation and completely suppressed pancreatic tumor growth in the xenograft mouse model [72].
5. Clinical Implications of Neutrophil and NET in PDAC
5.1. Prognosis in PDAC
The neutrophil-to-lymphocyte ratio (NLR) offers critical prognostic information in PDAC as well as breast cancer, lung cancer, and other types of cancer [73,74]. Studies have demonstrated that increased NLR correlates with a poor prognosis in patients with resectable and unresectable pancreatic cancer [30,75,76,77]. The combination of NLR and platelet-lymphocyte ratio significantly improves the prognostic stratification of metastatic PDAC patients [78]. For PDAC patients undergoing chemotherapy, both baseline NLR and post-chemotherapy NLR change (the post-chemotherapy NLR divided by the baseline NLR) are independent prognostic factors in overall survival [79]. Relative changes of NLR after two doses of immune checkpoint blockade are significantly correlated with an increased risk of death for PDAC patients [80]. Increased CXCR2+ neutrophils in the blood and bone marrow are correlated with worse clinical outcomes [30]. Baseline neutrophilia and increased absolute neutrophil count are associated with worse overall survival in patients with locally advanced pancreatic cancer [81,82].
Intratumoral neutrophils are also correlated with PDAC patient outcomes. CD177, a marker for neutrophils, is negatively correlated with the overall survival of PDAC patients [83]. CD66b is another marker for neutrophils. Higher levels of CD66b+ tumor-infiltrating neutrophils are significantly associated with shorter survival in PDAC patients [5]. Another group found that a high CD15+ TAN to CD8+ lymphocyte ratio in PDAC was associated with worse overall survival [30].
A recent study showed that plasma NET levels using SYTOX-positive areas could also predict the survival of PDAC patients [69]. High plasma levels of NET markers were indicators of increased death risk of PDAC patients. The combination of plasma TIMP-1 and NETs markedly enhanced the prognostic power. TIMP-1high NEThigh-patients had an almost sixteen-fold increase in mortality rate compared to TIMP-1low NETlow-patients. Notably, a combination of plasma levels of TIMP-1 and NETs with the clinically established marker CA19–9 exhibited a superior prognostic value as compared to CA19–9 alone. Tumor-Infiltrating CitH3-positive NETs were associated with poor overall survival and recurrence-free survival in PDAC patients. In addition, the combination of NETs with the 8th edition TNM staging system generated a novel model that improved the predictive accuracy for survival [84].
5.2. Neutrophil as a Potential Therapeutic Target in PDAC
The existing treatment of PDAC patients includes surgical resection, chemoradiation therapy, and immunotherapy, but only a small proportion of patients benefit from available therapies. Based on an increasing understanding of the role of TME in PDAC, neutrophils have emerged as a potential therapeutic target. Several preclinical studies with different mouse models have shown encouraging results of targeting neutrophils in pancreatic cancer (Table 1). The majority of neutrophil targeting preclinical studies use CXCR2 inhibitors or Ly6G antibodies [30]. This approach is in line with the main classes of current and past drug targets being receptors, among others [85]. In addition, the preference of CXCR2 as a target might lie in the fact that CXCR2 blockage not only inhibits the CXCL5/CXCR2 axis but also blocks the effect of other ligands of CXCR2, including CXCL1–3 and CXCL6–8. In breast cancer, CXCR2 inhibitors were demonstrated to be safe and tolerable, and relevant clinical trials are currently underway (NCT02370238) [86]. However, ligands, including cytokines, have become more important drug targets in the past two decades, largely due to the increased antibody-based therapies. Although no study was found in PDAC, CXCL5 inhibition emerged as a new strategy to attenuate tumor angiogenesis and thus slow tumor progression [87]. CXCL5 neutralizing antibodies treatment decreased the metastatic rate of breast cancer cells [88]. In lung cancer, CXCL5 neutralizing antibody improved the efficacy of tyrosine kinase inhibitor gefitinib without increased side effects [89]. More studies need to be conducted to investigate the therapeutic potential of CXCL5 blockage in PDAC.
6. Conclusions and Perspectives
More and more studies show that neutrophils play crucial roles in influencing the PDAC microenvironment and PDAC progression. The interactions among neutrophils, PDAC cells, and other components in TME are complex and are regulated by several signaling pathways. In addition, several parameters related to neutrophils are used as potential biomarkers for predicting clinical prognosis in PDAC patients. Moreover, unraveling the exact mechanisms of neutrophil function in PDAC development and new findings in preclinical and clinical studies will undoubtedly further our understanding of PDAC pathophysiology and provide a breakthrough in the treatment.
Author Contributions
Conceptualization, L.J. and J.S.; writing—original draft preparation, L.J.; writing—review and editing, H.S.K. and J.S.; visualization, L.J.; supervision, J.S.; funding acquisition, J.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Cancer Institute of the National Institutes of Health under award number K08CA234222.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Society, A.C. Cancer Facts & Figures 2020. Available online: https://www.cancer.org/content/dam/cancer-org/research/cancer-facts-and-statistics/annual-cancer-facts-and-figures/2020/cancer-facts-and-figures-2020.pdf (accessed on 6 August 2021).
- Rahib, L.; Smith, B.; Aizenberg, R.; Rosenzweig, A.; Fleshman, J.; Matrisian, L. Projecting cancer incidence and deaths to 2030: The unexpected burden of thyroid, liver, and pancreas cancers in the United States. Cancer Res. 2014, 74, 2913–2921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binnewies, M.; Roberts, E.; Kersten, K.; Chan, V.; Fearon, D.; Merad, M.; Coussens, L.; Gabrilovich, D.; Ostrand-Rosenberg, S.; Hedrick, C.; et al. Understanding the tumor immune microenvironment (TIME) for effective therapy. Nat. Med. 2018, 24, 541–550. [Google Scholar] [CrossRef]
- Dinh, H.Q.; Eggert, T.; Meyer, M.A.; Zhu, Y.P.; Olingy, C.E.; Llewellyn, R.; Wu, R.; Hedrick, C.C. Coexpression of CD71 and CD117 Identifies an Early Unipotent Neutrophil Progenitor Population in Human Bone Marrow. Immunity 2020, 53, 319–334. [Google Scholar] [CrossRef] [PubMed]
- Ino, Y.; Yamazaki-Itoh, R.; Shimada, K.; Iwasaki, M.; Kosuge, T.; Kanai, Y.; Hiraoka, N. Immune cell infiltration as an indicator of the immune microenvironment of pancreatic cancer. Br. J. Cancer 2013, 108, 914–923. [Google Scholar] [CrossRef] [PubMed]
- Soto-Perez-de-Celis, E.; Chavarri-Guerra, Y.; Leon-Rodriguez, E.; Gamboa-Dominguez, A. Tumor-Associated Neutrophils in Breast Cancer Subtypes. Asian Pac. J. Cancer Prev. APJCP 2017, 18, 2689–2693. [Google Scholar] [CrossRef]
- Galdiero, M.; Bianchi, P.; Grizzi, F.; Di Caro, G.; Basso, G.; Ponzetta, A.; Bonavita, E.; Barbagallo, M.; Tartari, S.; Polentarutti, N.; et al. Occurrence and significance of tumor-associated neutrophils in patients with colorectal cancer. Int. J. Cancer 2016, 139, 446–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, T.O.; Schmidt, H.; Møller, H.J.; Donskov, F.; Høyer, M.; Sjoegren, P.; Christensen, I.J.; Steiniche, T. Intratumoral neutrophils and plasmacytoid dendritic cells indicate poor prognosis and are associated with pSTAT3 expression in AJCC stage I/II melanoma. Cancer 2012, 118, 2476–2485. [Google Scholar] [CrossRef]
- Yuen, K.C.; Liu, L.-F.; Gupta, V.; Madireddi, S.; Keerthivasan, S.; Li, C.; Rishipathak, D.; Williams, P.; Kadel, E.E.; Koeppen, H.; et al. High systemic and tumor-associated IL-8 correlates with reduced clinical benefit of PD-L1 blockade. Nat. Med. 2020, 26, 693–698. [Google Scholar] [CrossRef]
- Arvanitakis, K.; Mitroulis, I.; Germanidis, G. Tumor-Associated Neutrophils in Hepatocellular Carcinoma Pathogenesis, Prognosis, and Therapy. Cancers 2021, 13, 2899. [Google Scholar] [CrossRef]
- Caruso, R.A.; Bellocco, R.; Pagano, M.; Bertoli, G.; Rigoli, L.; Inferrera, C. Prognostic value of intratumoral neutrophils in advanced gastric carcinoma in a high-risk area in northern Italy. Mod. Pathol. 2002, 15, 831–837. [Google Scholar] [CrossRef]
- Fossati, G.; Ricevuti, G.; Edwards, S.W.; Walker, C.; Dalton, A.; Rossi, M.L. Neutrophil infiltration into human gliomas. Acta Neuropathol. 1999, 98, 349–354. [Google Scholar] [CrossRef]
- Schalper, K.A.; Carleton, M.; Zhou, M.; Chen, T.; Feng, Y.; Huang, S.-P.; Walsh, A.M.; Baxi, V.; Pandya, D.; Baradet, T.; et al. Elevated serum interleukin-8 is associated with enhanced intratumor neutrophils and reduced clinical benefit of immune-checkpoint inhibitors. Nat. Med. 2020, 26, 688–692. [Google Scholar] [CrossRef]
- Fridlender, Z.; Sun, J.; Kim, S.; Kapoor, V.; Cheng, G.; Ling, L.; Worthen, G.; Albelda, S. Polarization of tumor-associated neutrophil phenotype by TGF-beta: “N1” versus “N2” TAN. Cancer Cell 2009, 16, 183–194. [Google Scholar] [CrossRef] [Green Version]
- Jaillon, S.; Ponzetta, A.; Di Mitri, D.; Santoni, A.; Bonecchi, R.; Mantovani, A. Neutrophil diversity and plasticity in tumour progression and therapy. Nat. Rev. Cancer 2020, 20, 485–503. [Google Scholar] [CrossRef]
- Wang, X.; Qiu, L.; Li, Z.; Wang, X.Y.; Yi, H. Understanding the Multifaceted Role of Neutrophils in Cancer and Autoimmune Diseases. Front. Immunol. 2018, 9, 2456. [Google Scholar] [CrossRef] [PubMed]
- Shojaei, F.; Wu, X.; Zhong, C.; Yu, L.; Liang, X.; Yao, J.; Blanchard, D.; Bais, C.; Peale, F.; van Bruggen, N.; et al. Bv8 regulates myeloid-cell-dependent tumour angiogenesis. Nature 2007, 450, 825–831. [Google Scholar] [CrossRef] [PubMed]
- Schmielau, J.; Finn, O. Activated granulocytes and granulocyte-derived hydrogen peroxide are the underlying mechanism of suppression of t-cell function in advanced cancer patients. Cancer Res. 2001, 61, 4756–4760. [Google Scholar]
- Kalafati, L.; Kourtzelis, I.; Schulte-Schrepping, J.; Li, X.; Hatzioannou, A.; Grinenko, T.; Hagag, E.; Sinha, A.; Has, C.; Dietz, S.; et al. Innate Immune Training of Granulopoiesis Promotes Anti-tumor Activity. Cell 2020, 183, 771–785. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Liu, L.; Xu, H.; Wu, C.; Xiang, J.; Xu, J.; Liu, C.; Long, J.; Ni, Q.; Yu, X. Infiltrating immune cells and gene mutations in pancreatic ductal adenocarcinoma. Br. J. Surg. 2016, 103, 1189–1199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagaraj, S.; Gupta, K.; Pisarev, V.; Kinarsky, L.; Sherman, S.; Kang, L.; Herber, D.L.; Schneck, J.; Gabrilovich, D.I. Altered recognition of antigen is a mechanism of CD8+ T cell tolerance in cancer. Nat. Med. 2007, 13, 828–835. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Yu, S.; Kappes, J.; Wang, J.; Grizzle, W.E.; Zinn, K.R.; Zhang, H.G. Expansion of spleen myeloid suppressor cells represses NK cell cytotoxicity in tumor-bearing host. Blood 2007, 109, 4336–4342. [Google Scholar] [CrossRef]
- Marvel, D.; Gabrilovich, D. Myeloid-derived suppressor cells in the tumor microenvironment: Expect the unexpected. J. Clin. Investig. 2015, 125, 3356–3364. [Google Scholar] [CrossRef] [PubMed]
- Youn, J.I.; Nagaraj, S.; Collazo, M.; Gabrilovich, D.I. Subsets of myeloid-derived suppressor cells in tumor-bearing mice. J. Immunol. 2008, 181, 5791–5802. [Google Scholar] [CrossRef]
- Youn, J.I.; Gabrilovich, D.I. The biology of myeloid-derived suppressor cells: The blessing and the curse of morphological and functional heterogeneity. Eur. J. Immunol. 2010, 40, 2969–2975. [Google Scholar] [CrossRef] [PubMed]
- Coffelt, S.; Wellenstein, M.; de Visser, K. Neutrophils in cancer: Neutral no more. Nat. Rev. Cancer 2016, 16, 431–446. [Google Scholar] [CrossRef] [Green Version]
- Shaul, M.E.; Fridlender, Z.G. Tumour-associated neutrophils in patients with cancer. Nat. Rev. Clin. Oncol. 2019, 16, 601–620. [Google Scholar] [CrossRef]
- Fridlender, Z.G.; Sun, J.; Mishalian, I.; Singhal, S.; Cheng, G.; Kapoor, V.; Horng, W.; Fridlender, G.; Bayuh, R.; Worthen, G.S.; et al. Transcriptomic analysis comparing tumor-associated neutrophils with granulocytic myeloid-derived suppressor cells and normal neutrophils. PLoS ONE 2012, 7, e31524. [Google Scholar] [CrossRef] [PubMed]
- Steele, C.; Karim, S.; Leach, J.; Bailey, P.; Upstill-Goddard, R.; Rishi, L.; Foth, M.; Bryson, S.; McDaid, K.; Wilson, Z.; et al. CXCR2 Inhibition Profoundly Suppresses Metastases and Augments Immunotherapy in Pancreatic Ductal Adenocarcinoma. Cancer Cell 2016, 29, 832–845. [Google Scholar] [CrossRef] [Green Version]
- Nywening, T.M.; Belt, B.A.; Cullinan, D.R.; Panni, R.Z.; Han, B.J.; Sanford, D.E.; Jacobs, R.C.; Ye, J.; Patel, A.A.; Gillanders, W.E.; et al. Targeting both tumour-associated CXCR2(+) neutrophils and CCR2(+) macrophages disrupts myeloid recruitment and improves chemotherapeutic responses in pancreatic ductal adenocarcinoma. Gut 2018, 67, 1112–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosoi, F.; Izumi, H.; Kawahara, A.; Murakami, Y.; Kinoshita, H.; Kage, M.; Nishio, K.; Kohno, K.; Kuwano, M.; Ono, M. N-myc downstream regulated gene 1/Cap43 suppresses tumor growth and angiogenesis of pancreatic cancer through attenuation of inhibitor of kappaB kinase beta expression. Cancer Res. 2009, 69, 4983–4991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baggiolini, M.; Dewald, B.; Moser, B. Human chemokines: An update. Annu. Rev. Immunol. 1997, 15, 675–705. [Google Scholar] [CrossRef] [PubMed]
- Qiao, B.; Luo, W.; Liu, Y.; Wang, J.; Liu, C.; Liu, Z.; Chen, S.; Gu, J.; Qi, X.; Wu, T. The prognostic value of CXC chemokine receptor 2 (CXCR2) in cancers: A meta-analysis. Oncotarget 2018, 9, 15068–15076. [Google Scholar] [CrossRef] [Green Version]
- Sano, M.; Ijichi, H.; Takahashi, R.; Miyabayashi, K.; Fujiwara, H.; Yamada, T.; Kato, H.; Nakatsuka, T.; Tanaka, Y.; Tateishi, K.; et al. Blocking CXCLs-CXCR2 axis in tumor-stromal interactions contributes to survival in a mouse model of pancreatic ductal adenocarcinoma through reduced cell invasion/migration and a shift of immune-inflammatory microenvironment. Oncogenesis 2019, 8, 8. [Google Scholar] [CrossRef]
- Najjar, Y.G.; Rayman, P.; Jia, X.; Pavicic, P.G., Jr.; Rini, B.I.; Tannenbaum, C.; Ko, J.; Haywood, S.; Cohen, P.; Hamilton, T.; et al. Myeloid-Derived Suppressor Cell Subset Accumulation in Renal Cell Carcinoma Parenchyma Is Associated with Intratumoral Expression of IL1β, IL8, CXCL5, and Mip-1α. Clin. Cancer Res. 2017, 23, 2346–2355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toh, B.; Wang, X.; Keeble, J.; Sim, W.J.; Khoo, K.; Wong, W.C.; Kato, M.; Prevost-Blondel, A.; Thiery, J.P.; Abastado, J.P. Mesenchymal transition and dissemination of cancer cells is driven by myeloid-derived suppressor cells infiltrating the primary tumor. PLoS Biol. 2011, 9, e1001162. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Guo, F. A selective sphingosine-1-phosphate receptor 1 agonist SEW-2871 aggravates gastric cancer by recruiting myeloid-derived suppressor cells. J. Biochem. 2018, 163, 77–83. [Google Scholar] [CrossRef]
- Yu, F.; Shi, Y.; Wang, J.; Li, J.; Fan, D.; Ai, W. Deficiency of Kruppel-like factor KLF4 in mammary tumor cells inhibits tumor growth and pulmonary metastasis and is accompanied by compromised recruitment of myeloid-derived suppressor cells. Int. J. Cancer 2013, 133, 2872–2883. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, M.T.; Kemp, S.B.; Salas-Escabillas, D.J.; Zhang, Y.; Steele, N.G.; The, S.; Long, D.; Benitz, S.; Yan, W.; Margolskee, R.F.; et al. The Gustatory Sensory G-Protein GNAT3 Suppresses Pancreatic Cancer Progression in Mice. Cell. Mol. Gastroenterol. Hepatol. 2021, 11, 349–369. [Google Scholar] [CrossRef] [PubMed]
- Iida-Norita, R.; Kawamura, M.; Suzuki, Y.; Hamada, S.; Masamune, A.; Furukawa, T.; Sato, Y. Vasohibin-2 plays an essential role in metastasis of pancreatic ductal adenocarcinoma. Cancer Sci. 2019, 110, 2296–2308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chao, T.; Furth, E.; Vonderheide, R. CXCR2-Dependent Accumulation of Tumor-Associated Neutrophils Regulates T-cell Immunity in Pancreatic Ductal Adenocarcinoma. Cancer Immunol. Res. 2016, 4, 968–982. [Google Scholar] [CrossRef] [Green Version]
- Stromnes, I.; Brockenbrough, J.; Izeradjene, K.; Carlson, M.; Cuevas, C.; Simmons, R.; Greenberg, P.; Hingorani, S. Targeted depletion of an MDSC subset unmasks pancreatic ductal adenocarcinoma to adaptive immunity. Gut 2014, 63, 1769–1781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phan, V.T.; Wu, X.; Cheng, J.H.; Sheng, R.X.; Chung, A.S.; Zhuang, G.; Tran, C.; Song, Q.; Kowanetz, M.; Sambrone, A.; et al. Oncogenic RAS pathway activation promotes resistance to anti-VEGF therapy through G-CSF-induced neutrophil recruitment. Proc. Natl. Acad. Sci. USA 2013, 110, 6079–6084. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Jiang, W.; Zhao, K.; Wang, H.; Zhou, T.; Bai, W.; Wang, X.; Zhao, T.; Huang, C.; Gao, S.; et al. Tumoral EHF predicts the efficacy of anti-PD1 therapy in pancreatic ductal adenocarcinoma. J. Exp. Med. 2019, 216, 656–673. [Google Scholar] [CrossRef] [Green Version]
- Das, S.; Shapiro, B.; Vucic, E.A.; Vogt, S.; Bar-Sagi, D. Tumor Cell-Derived IL1β Promotes Desmoplasia and Immune Suppression in Pancreatic Cancer. Cancer Res. 2020, 80, 1088–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choueiry, F.; Torok, M.; Shakya, R.; Agrawal, K.; Deems, A.; Benner, B.; Hinton, A.; Shaffer, J.; Blaser, B.W.; Noonan, A.M.; et al. CD200 promotes immunosuppression in the pancreatic tumor microenvironment. J. Immunother. Cancer 2020, 8. [Google Scholar] [CrossRef]
- Li, J.; Yuan, S.; Norgard, R.J.; Yan, F.; Yamazoe, T.; Blanco, A.; Stanger, B.Z. Tumor Cell-Intrinsic USP22 Suppresses Antitumor Immunity in Pancreatic Cancer. Cancer Immunol. Res. 2020, 8, 282–291. [Google Scholar] [CrossRef]
- Liu, X.; Zhou, Z.; Cheng, Q.; Wang, H.; Cao, H.; Xu, Q.; Tuo, Y.; Jiang, L.; Zou, Y.; Ren, H.; et al. Acceleration of pancreatic tumorigenesis under immunosuppressive microenvironment induced by Reg3g overexpression. Cell Death Dis. 2017, 8, e3033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, P.; Chang, D.K.; Nones, K.; Johns, A.L.; Patch, A.M.; Gingras, M.C.; Miller, D.K.; Christ, A.N.; Bruxner, T.J.; Quinn, M.C.; et al. Genomic analyses identify molecular subtypes of pancreatic cancer. Nature 2016, 531, 47–52. [Google Scholar] [CrossRef]
- Moffitt, R.A.; Marayati, R.; Flate, E.L.; Volmar, K.E.; Loeza, S.G.; Hoadley, K.A.; Rashid, N.U.; Williams, L.A.; Eaton, S.C.; Chung, A.H.; et al. Virtual microdissection identifies distinct tumor- and stroma-specific subtypes of pancreatic ductal adenocarcinoma. Nat. Genet. 2015, 47, 1168–1178. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chandra, V.; Riquelme Sanchez, E.; Dutta, P.; Quesada, P.; Rakoski, A.; Zoltan, M.; Arora, N.; Baydogan, S.; Horne, W.; et al. Interleukin-17-induced neutrophil extracellular traps mediate resistance to checkpoint blockade in pancreatic cancer. J. Exp. Med. 2020, 217, e20190354. [Google Scholar] [CrossRef]
- Jin, W.; Yin, H.; Li, H.; Yu, X.J.; Xu, H.X.; Liu, L. Neutrophil extracellular DNA traps promote pancreatic cancer cells migration and invasion by activating EGFR/ERK pathway. J. Cell. Mol. Med. 2021. [Google Scholar] [CrossRef] [PubMed]
- McAllister, F.; Bailey, J.M.; Alsina, J.; Nirschl, C.J.; Sharma, R.; Fan, H.; Rattigan, Y.; Roeser, J.C.; Lankapalli, R.H.; Zhang, H.; et al. Oncogenic Kras activates a hematopoietic-to-epithelial IL-17 signaling axis in preinvasive pancreatic neoplasia. Cancer Cell 2014, 25, 621–637. [Google Scholar] [CrossRef] [Green Version]
- Siret, C.; Collignon, A.; Silvy, F.; Robert, S.; Cheyrol, T.; André, P.; Rigot, V.; Iovanna, J.; van de Pavert, S.; Lombardo, D.; et al. Deciphering the Crosstalk Between Myeloid-Derived Suppressor Cells and Regulatory T Cells in Pancreatic Ductal Adenocarcinoma. Front. Immunol. 2019, 10, 3070. [Google Scholar] [CrossRef] [PubMed]
- Holokai, L.; Chakrabarti, J.; Lundy, J.; Croagh, D.; Adhikary, P.; Richards, S.S.; Woodson, C.; Steele, N.; Kuester, R.; Scott, A.; et al. Murine- and Human-Derived Autologous Organoid/Immune Cell Co-Cultures as Pre-Clinical Models of Pancreatic Ductal Adenocarcinoma. Cancers 2020, 12, 3816. [Google Scholar] [CrossRef]
- Cartwright, A.N.R.; Suo, S.; Badrinath, S.; Kumar, S.; Melms, J.; Luoma, A.; Bagati, A.; Saadatpour, A.; Izar, B.; Yuan, G.C.; et al. Immunosuppressive Myeloid Cells Induce Nitric Oxide-Dependent DNA Damage and p53 Pathway Activation in CD8(+) T Cells. Cancer Immunol. Res. 2021. [Google Scholar] [CrossRef]
- Li, C.; Cui, L.; Yang, L.; Wang, B.; Zhuo, Y.; Zhang, L.; Wang, X.; Zhang, Q.; Zhang, S. Pancreatic Stellate Cells Promote Tumor Progression by Promoting an Immunosuppressive Microenvironment in Murine Models of Pancreatic Cancer. Pancreas 2020, 49, 120–127. [Google Scholar] [CrossRef]
- Miller-Ocuin, J.; Liang, X.; Boone, B.; Doerfler, W.; Singhi, A.; Tang, D.; Kang, R.; Lotze, M.; Zeh, H. DNA released from neutrophil extracellular traps (NETs) activates pancreatic stellate cells and enhances pancreatic tumor growth. Oncoimmunology 2019, 8, e1605822. [Google Scholar] [CrossRef]
- Mayer, P.; Dinkic, C.; Jesenofsky, R.; Klauss, M.; Schirmacher, P.; Dapunt, U.; Hackert, T.; Uhle, F.; Hänsch, G.M.; Gaida, M.M. Changes in the microarchitecture of the pancreatic cancer stroma are linked to neutrophil-dependent reprogramming of stellate cells and reflected by diffusion-weighted magnetic resonance imaging. Theranostics 2018, 8, 13–30. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Hang, J.J.; Han, T.; Zhuo, M.; Jiao, F.; Wang, L.W. The M2 phenotype of tumor-associated macrophages in the stroma confers a poor prognosis in pancreatic cancer. Tumour Biol. J. Int. Soc. Oncodevelopmental Biol. Med. 2016, 37, 8657–8664. [Google Scholar] [CrossRef]
- Sinha, P.; Clements, V.K.; Bunt, S.K.; Albelda, S.M.; Ostrand-Rosenberg, S. Cross-talk between myeloid-derived suppressor cells and macrophages subverts tumor immunity toward a type 2 response. J. Immunol. 2007, 179, 977–983. [Google Scholar] [CrossRef] [PubMed]
- Bausch, D.; Pausch, T.; Krauss, T.; Hopt, U.T.; Fernandez-del-Castillo, C.; Warshaw, A.L.; Thayer, S.P.; Keck, T. Neutrophil granulocyte derived MMP-9 is a VEGF independent functional component of the angiogenic switch in pancreatic ductal adenocarcinoma. Angiogenesis 2011, 14, 235–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef] [PubMed]
- Berger-Achituv, S.; Brinkmann, V.; Abed, U.A.; Kühn, L.I.; Ben-Ezra, J.; Elhasid, R.; Zychlinsky, A. A proposed role for neutrophil extracellular traps in cancer immunoediting. Front. Immunol. 2013, 4, 48. [Google Scholar] [CrossRef] [Green Version]
- Demers, M.; Wong, S.L.; Martinod, K.; Gallant, M.; Cabral, J.E.; Wang, Y.; Wagner, D.D. Priming of neutrophils toward NETosis promotes tumor growth. Oncoimmunology 2016, 5, e1134073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanamaru, R.; Ohzawa, H.; Miyato, H.; Matsumoto, S.; Haruta, H.; Kurashina, K.; Saito, S.; Hosoya, Y.; Yamaguchi, H.; Yamashita, H.; et al. Low density neutrophils (LDN) in postoperative abdominal cavity assist the peritoneal recurrence through the production of neutrophil extracellular traps (NETs). Sci. Rep. 2018, 8, 632. [Google Scholar] [CrossRef]
- Albrengues, J.; Shields, M.A.; Ng, D.; Park, C.G.; Ambrico, A.; Poindexter, M.E.; Upadhyay, P.; Uyeminami, D.L.; Pommier, A.; Küttner, V.; et al. Neutrophil extracellular traps produced during inflammation awaken dormant cancer cells in mice. Science 2018, 361, eaao4227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, H.S.; Gu, J.; Kim, J.E.; Nam, Y.; Song, J.W.; Kim, H.K. Cancer cell-induced neutrophil extracellular traps promote both hypercoagulability and cancer progression. PLoS ONE 2019, 14, e0216055. [Google Scholar] [CrossRef] [Green Version]
- Schoeps, B.; Eckfeld, C.; Prokopchuk, O.; Böttcher, J.; Häußler, D.; Steiger, K.; Demir, I.; Knolle, P.; Soehnlein, O.; Jenne, D.; et al. TIMP-1 triggers neutrophil extracellular trap formation in pancreatic cancer. Cancer Res. 2021. [Google Scholar] [CrossRef]
- Kajioka, H.; Kagawa, S.; Ito, A.; Yoshimoto, M.; Sakamoto, S.; Kikuchi, S.; Kuroda, S.; Yoshida, R.; Umeda, Y.; Noma, K.; et al. Targeting neutrophil extracellular traps with thrombomodulin prevents pancreatic cancer metastasis. Cancer Lett. 2021, 497, 1–13. [Google Scholar] [CrossRef]
- Cools-Lartigue, J.; Spicer, J.; Najmeh, S.; Ferri, L. Neutrophil extracellular traps in cancer progression. Cell. Mol. Life Sci. CMLS 2014, 71, 4179–4194. [Google Scholar] [CrossRef]
- Munir, H.; Jones, J.O.; Janowitz, T.; Hoffmann, M.; Euler, M.; Martins, C.P.; Welsh, S.J.; Shields, J.D. Stromal-driven and Amyloid β-dependent induction of neutrophil extracellular traps modulates tumor growth. Nat. Commun. 2021, 12, 683. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Yang, H.; Cai, D.; Xiang, L.; Fang, W.; Wang, R. Preoperative peripheral blood neutrophil-to-lymphocyte ratios (NLR) and platelet-to-lymphocyte ratio (PLR) related nomograms predict the survival of patients with limited-stage small-cell lung cancer. Transl. Lung Cancer Res. 2021, 10, 866–877. [Google Scholar] [CrossRef]
- Orditura, M.; Galizia, G.; Diana, A.; Saccone, C.; Cobellis, L.; Ventriglia, J.; Iovino, F.; Romano, C.; Morgillo, F.; Mosca, L.; et al. Neutrophil to lymphocyte ratio (NLR) for prediction of distant metastasis-free survival (DMFS) in early breast cancer: A propensity score-matched analysis. ESMO Open 2016, 1, e000038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcea, G.; Ladwa, N.; Neal, C.P.; Metcalfe, M.S.; Dennison, A.R.; Berry, D.P. Preoperative neutrophil-to-lymphocyte ratio (NLR) is associated with reduced disease-free survival following curative resection of pancreatic adenocarcinoma. World J. Surg. 2011, 35, 868–872. [Google Scholar] [CrossRef]
- Stotz, M.; Gerger, A.; Eisner, F.; Szkandera, J.; Loibner, H.; Ress, A.L.; Kornprat, P.; AlZoughbi, W.; Seggewies, F.S.; Lackner, C.; et al. Increased neutrophil-lymphocyte ratio is a poor prognostic factor in patients with primary operable and inoperable pancreatic cancer. Br. J. Cancer 2013, 109, 416–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, R.; Takagi, T.; Hikichi, T.; Konno, N.; Sugimoto, M.; Watanabe, K.O.; Nakamura, J.; Waragai, Y.; Kikuchi, H.; Takasumi, M.; et al. Derived neutrophil/lymphocyte ratio predicts gemcitabine therapy outcome in unresectable pancreatic cancer. Oncol. Lett. 2016, 11, 3441–3445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toledano-Fonseca, M.; Cano, M.; Inga, E.; Gómez-España, A.; Guil-Luna, S.; García-Ortiz, M.; Mena-Osuna, R.; De la Haba-Rodriguez, J.; Rodríguez-Ariza, A.; Aranda, E. The Combination of Neutrophil-Lymphocyte Ratio and Platelet-Lymphocyte Ratio with Liquid Biopsy Biomarkers Improves Prognosis Prediction in Metastatic Pancreatic Cancer. Cancers 2021, 13, 1210. [Google Scholar] [CrossRef] [PubMed]
- Luo, G.; Guo, M.; Liu, Z.; Xiao, Z.; Jin, K.; Long, J.; Liu, L.; Liu, C.; Xu, J.; Ni, Q.; et al. Blood Neutrophil–Lymphocyte Ratio Predicts Survival in Patients with Advanced Pancreatic Cancer Treated with Chemotherapy. Ann. Surg. Oncol. 2015, 22, 670–676. [Google Scholar] [CrossRef] [PubMed]
- Shang, J.; Han, X.; Zha, H.; Tao, H.; Li, X.; Yuan, F.; Chen, G.; Wang, L.; Ma, J.; Hu, Y. Systemic Immune-Inflammation Index and Changes of Neutrophil-Lymphocyte Ratio as Prognostic Biomarkers for Patients With Pancreatic Cancer Treated With Immune Checkpoint Blockade. Front. Oncol. 2021, 11, 585271. [Google Scholar] [CrossRef]
- Schernberg, A.; Vernerey, D.; Goldstein, D.; Van Laethem, J.; Glimelius, B.; van Houtte, P.; Bonnetain, F.; Louvet, C.; Hammel, P.; Huguet, F. Predictive Value of Neutrophils Count for Local Tumor Control After Chemoradiotherapy in Patients With Locally Advanced Pancreatic Carcinoma. Int. J. Radiat. Oncol. Biol. Phys. 2021. [Google Scholar] [CrossRef]
- Deng, G.; Yan, H.; Guo, Z.; Dai, G. Correlation between Baseline Serum Tumor Markers and Clinical Characteristic Factors in Patients with Advanced Pancreatic Cancer. OncoTargets Ther. 2020, 13, 11151–11163. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Fang, T.; Huang, L.; Wang, H.; Zhang, L.; Wang, Z.; Cui, Y. Neutrophils infiltrating pancreatic ductal adenocarcinoma indicate higher malignancy and worse prognosis. Biochem. Biophys. Res. Commun. 2018, 501, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.; Xu, H.X.; Zhang, S.R.; Li, H.; Wang, W.Q.; Gao, H.L.; Wu, C.T.; Xu, J.Z.; Qi, Z.H.; Li, S.; et al. Tumor-Infiltrating NETs Predict Postsurgical Survival in Patients with Pancreatic Ductal Adenocarcinoma. Ann. Surg. Oncol. 2019, 26, 635–643. [Google Scholar] [CrossRef]
- Attwood, M.M.; Jonsson, J.; Rask-Andersen, M.; Schiöth, H.B. Soluble ligands as drug targets. Nat. Rev. Drug Discov. 2020, 19, 695–710. [Google Scholar] [CrossRef] [PubMed]
- Schott, A.F.; Goldstein, L.J.; Cristofanilli, M.; Ruffini, P.A.; McCanna, S.; Reuben, J.M.; Perez, R.P.; Kato, G.; Wicha, M. Phase Ib Pilot Study to Evaluate Reparixin in Combination with Weekly Paclitaxel in Patients with HER-2-Negative Metastatic Breast Cancer. Clin. Cancer Res. 2017, 23, 5358–5365. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Wang, H.; Sun, M.; Deng, X.; Wu, X.; Ma, Y.; Li, M.; Shuoa, S.M.; You, Q.; Miao, L. CXCL5/CXCR2 axis in tumor microenvironment as potential diagnostic biomarker and therapeutic target. Cancer Commun. 2020, 40, 69–80. [Google Scholar] [CrossRef] [Green Version]
- Hsu, Y.L.; Hou, M.F.; Kuo, P.L.; Huang, Y.F.; Tsai, E.M. Breast tumor-associated osteoblast-derived CXCL5 increases cancer progression by ERK/MSK1/Elk-1/snail signaling pathway. Oncogene 2013, 32, 4436–4447. [Google Scholar] [CrossRef] [Green Version]
- Kuo, P.L.; Huang, M.S.; Hung, J.Y.; Chou, S.H.; Chiang, S.Y.; Huang, Y.F.; Yang, C.J.; Tsai, M.J.; Chang, W.A.; Hsu, Y.L. Synergistic effect of lung tumor-associated dendritic cell-derived HB-EGF and CXCL5 on cancer progression. Int. J. Cancer 2014, 135, 96–108. [Google Scholar] [CrossRef]
- Wang, X.; Hu, L.-P.; Qin, W.-T.; Yang, Q.; Chen, D.-Y.; Li, Q.; Zhou, K.-X.; Huang, P.-Q.; Xu, C.-J.; Li, J.; et al. Identification of a subset of immunosuppressive P2RX1-negative neutrophils in pancreatic cancer liver metastasis. Nat. Commun. 2021, 12, 174. [Google Scholar] [CrossRef]
Figure 1.
Tumor cell-secreted factors attract neutrophils. Tumor cells can release several factors to recruit neutrophils, such as the CXC family (CXCL1, CXCL2, CXCL5, and CXCL8), growth factors (GM-CSF, G-CSF, and M-CSF), IL-1β, and CD200. Different pathways regulate these factor secretions.
Figure 1.
Tumor cell-secreted factors attract neutrophils. Tumor cells can release several factors to recruit neutrophils, such as the CXC family (CXCL1, CXCL2, CXCL5, and CXCL8), growth factors (GM-CSF, G-CSF, and M-CSF), IL-1β, and CD200. Different pathways regulate these factor secretions.

Figure 2.
Interactions between neutrophils and T cells. IL-17 secreted from CD4+ T cells and γδT cells attracts neutrophils. Neutrophils can inhibit CD8+ T cell proliferation, activation, and recruitment. Neutrophils and Treg cells can activate each other.
Figure 2.
Interactions between neutrophils and T cells. IL-17 secreted from CD4+ T cells and γδT cells attracts neutrophils. Neutrophils can inhibit CD8+ T cell proliferation, activation, and recruitment. Neutrophils and Treg cells can activate each other.


{kind=link}
{kind=link}
Table 1.
Preclinical studies of targeting neutrophils in pancreatic cancer.
| Mouse Model | Cell Line | Treatment | Results | Reference |
|---|---|---|---|---|
| LSL-KrasG12D/+; Tgfbr2flox/flox; Ptf1a-cre (PKF) Cxcr2+/+ and Cxcr2+/− | - | - | Cxcr2 knockout significantly extends the survival of PDAC mice. | [34] |
| LSL-KrasG12D/+; LSL-Trp53R172H/+; Pdx1-Cre (KPC) Cxcr2+/+ and Cxcr2-/− | - | anti-Ly6G antibody 1A8 CXCR2 signaling inhibitor pepducin CXCR2 inhibitor AZ13381758 | Cxcr2 deletion completely abrogated metastasis. Depletion of Ly6G+ cells resulted in strong suppression of metastasis. Pepducin treatment resulted in a significant increase in survival. CXCR2 inhibitor treatment prolonged the survival of KPC mice and significantly protected from metastasis. | [29] |
| KPC | - | anti-Ly6G antibody 1A8 | Depletion of Ly6G+ cells induced tumor cell apoptosis. | [42] |
| C57BL/6 | KPC cells | anti-Ly6G antibody | Depletion of Ly6G+ cells significantly delayed tumor growth in the orthotopic model. | [51] |
| C57BL/6 | KPC cells | CXCR2 inhibitor SB225002 anti-Ly6G antibody 1A8 | CXCR2 inhibitor or depletion of Ly6G+ cells reduced tumor burden in subcutaneous and orthotopic models. | [30] |
| Cxcr2+/+ and Cxcr2−/− | KPC cells | - | CXCR2 ablation inhibited tumor growth in the subcutaneous model. | [41] |
| Padi4+/+ and Padi4−/− | Panc02 cells | - | NET-deficiency decreased pancreatic tumor burden and increased survival. | [58] |
| C57BL/6 | KPC cells | PAD4 inhibitor GSK484 | GSK484 treatment completely suppressed tumor growth in the subcutaneous model. | [72] |
| C57BL/6 | KPC cells | anti-Ly6G antibody 1A8 | Depletion of Ly6G+ cells reduced the formation of liver metastases in the intrasplenic model. | [90] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Jin, L.; Kim, H.S.; Shi, J. Neutrophil in the Pancreatic Tumor Microenvironment. Biomolecules 2021, 11, 1170. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11081170
AMA Style
Jin L, Kim HS, Shi J. Neutrophil in the Pancreatic Tumor Microenvironment. Biomolecules. 2021; 11(8):1170. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11081170
Chicago/Turabian StyleJin, Lin, Hong Sun Kim, and Jiaqi Shi. 2021. "Neutrophil in the Pancreatic Tumor Microenvironment" Biomolecules 11, no. 8: 1170. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11081170
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.