
RadA, a MSCRAMM Adhesin of the Dominant Symbiote Ruminococcus gnavus E1, Binds Human Immunoglobulins and Intestinal Mucins

 ,
,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. radA Cluster Analysis
2.2. The radA Gene Is Only Expressed “In Situ”
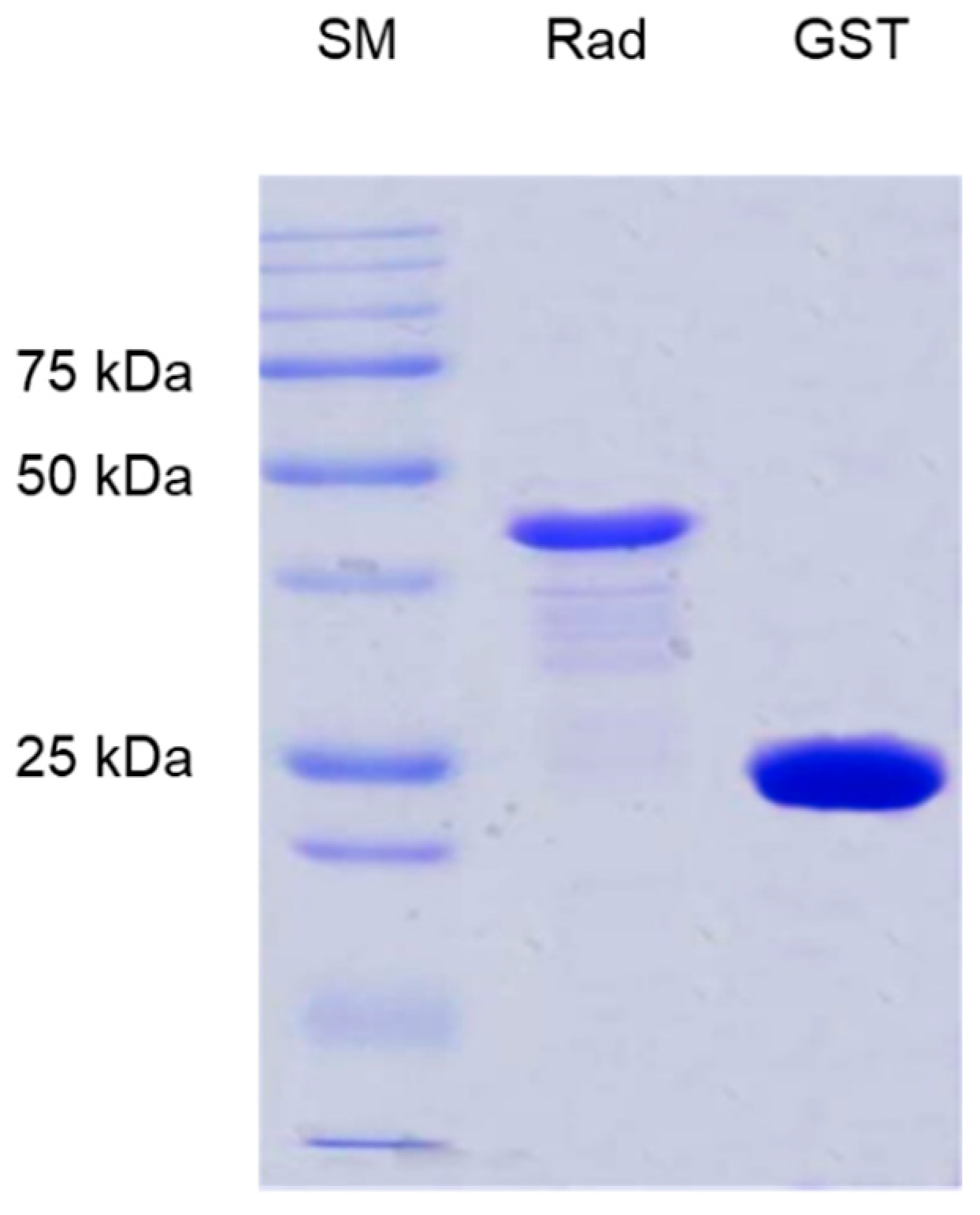
2.3. Heterologous Expression of GST-RadA35-252
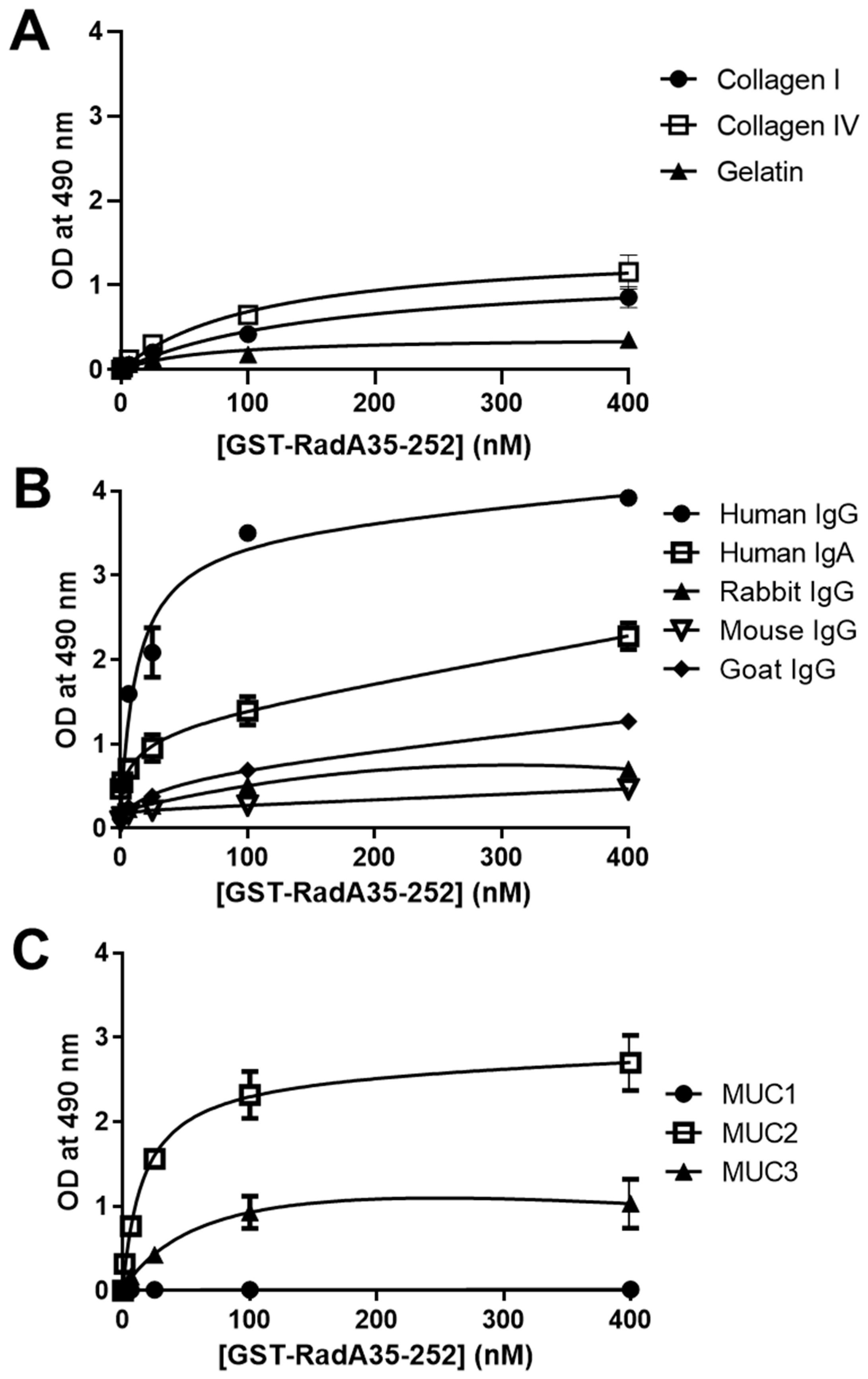
2.4. GST-RadA35-252 Binds to Immunoglobulins and Mucins
2.5. GST- RadA35-252 Recognizes a Carbohydrate and a Peptide Motif
2.6. GST-RadA35-252 Recognizes a Specific Carbohydrate Motif of Type II Mucin
2.7. GST-RadA35-252 Binds to the Surface of Human Intestinal Epithelial Cells
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Media
4.2. Animals, Ethics Statement and Experimental Design
4.3. Cloning
4.4. Expression and Purification of Recombinant GST-Tagged Fusion Rad Domains
4.5. radA Expression
4.6. Bioinformatics Analysis
4.7. Solid-Phase Binding Assays
4.8. Binding Assay Using Human Intestinal Epithelial Cells
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Bry, L.; Falk, P.G.; Midtvedt, T.; Gordon, J.I. A Model of Host-Microbial Interactions in an Open Mammalian Ecosystem. Science 1996, 273, 1380–1383. [Google Scholar] [CrossRef] [PubMed]
- Hooper, L.V.; Xu, J.; Falk, P.G.; Midtvedt, T.; Gordon, J.I. A Molecular Sensor That Allows a Gut Commensal to Control Its Nutrient Foundation in a Competitive Ecosystem. Proc. Natl. Acad. Sci. USA 1999, 96, 9833–9838. [Google Scholar] [CrossRef] [Green Version]
- Pizarro-Cerdá, J.; Cossart, P. Bacterial Adhesion and Entry into Host Cells. Cell 2006, 124, 715–727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stones, D.H.; Krachler, A.M. Against the Tide: The Role of Bacterial Adhesion in Host Colonization. Biochem. Soc. Trans. 2016, 44, 1571–1580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Susmitha, A.; Bajaj, H.; Madhavan Nampoothiri, K. The Divergent Roles of Sortase in the Biology of Gram-Positive Bacteria. Cell Surf. 2021, 7, 100055. [Google Scholar] [CrossRef]
- Madani, A.; Garakani, K.; Mofrad, M.R.K. Molecular Mechanics of Staphylococcus Aureus Adhesin, CNA, and the Inhibition of Bacterial Adhesion by Stretching Collagen. PLoS ONE 2017, 12, e0179601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rich, R.L.; Kreikemeyer, B.; Owens, R.T.; LaBrenz, S.; Narayana, S.V.; Weinstock, G.M.; Murray, B.E.; Höök, M. Ace Is a Collagen-Binding MSCRAMM from Enterococcus Faecalis. J. Biol. Chem. 1999, 274, 26939–26945. [Google Scholar] [CrossRef] [Green Version]
- Nallapareddy, S.R.; Weinstock, G.M.; Murray, B.E. Clinical Isolates of Enterococcus Faecium Exhibit Strain-Specific Collagen Binding Mediated by Acm, a New Member of the MSCRAMM Family. Mol. Microbiol. 2003, 47, 1733–1747. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.H.; Avilés-Reyes, A.; Scott-Anne, K.; Gregoire, S.; Watson, G.E.; Sampson, E.; Progulske-Fox, A.; Koo, H.; Bowen, W.H.; Lemos, J.A.; et al. The Collagen Binding Protein Cnm Contributes to Oral Colonization and Cariogenicity of Streptococcus Mutans OMZ175. Infect. Immun. 2015, 83, 2001–2010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Ponnuraj, K.; Xu, Y.; Ganesh, V.K.; Sillanpää, J.; Murray, B.E.; Narayana, S.V.L.; Höök, M. The Enterococcus Faecalis MSCRAMM ACE Binds Its Ligand by the Collagen Hug Model. J. Biol. Chem. 2007, 282, 19629–19637. [Google Scholar] [CrossRef] [Green Version]
- Johansson, M.E.V.; Ambort, D.; Pelaseyed, T.; Schütte, A.; Gustafsson, J.K.; Ermund, A.; Subramani, D.B.; Holmén-Larsson, J.M.; Thomsson, K.A.; Bergström, J.H.; et al. Composition and Functional Role of the Mucus Layers in the Intestine. Cell. Mol. Life Sci. 2011, 68, 3635–3641. [Google Scholar] [CrossRef]
- Cantarel, B.L.; Coutinho, P.M.; Rancurel, C.; Bernard, T.; Lombard, V.; Henrissat, B. The Carbohydrate-Active EnZymes Database (CAZy): An Expert Resource for Glycogenomics. Nucleic Acids Res. 2009, 37, D233–D238. [Google Scholar] [CrossRef]
- Flint, H.J.; Scott, K.P.; Duncan, S.H.; Louis, P.; Forano, E. Microbial Degradation of Complex Carbohydrates in the Gut. Gut Microbes 2012, 3, 289–306. [Google Scholar] [CrossRef] [Green Version]
- Marcobal, A.; Southwick, A.M.; Earle, K.A.; Sonnenburg, J.L. A Refined Palate: Bacterial Consumption of Host Glycans in the Gut. Glycobiology 2013, 23, 1038–1046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, P.H.; Kolarich, D.; Packer, N.H. Mucin-Type O-Glycosylation--Putting the Pieces Together. FEBS J. 2010, 277, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Moran, A.P.; Gupta, A.; Joshi, L. Sweet-Talk: Role of Host Glycosylation in Bacterial Pathogenesis of the Gastrointestinal Tract. Gut 2011, 60, 1412–1425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juge, N. Microbial Adhesins to Gastrointestinal Mucus. Trends Microbiol. 2012, 20, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Roos, S.; Jonsson, H. A High-Molecular-Mass Cell-Surface Protein from Lactobacillus Reuteri 1063 Adheres to Mucus Components. Microbiology 2002, 148, 433–442. [Google Scholar] [CrossRef] [Green Version]
- MacKenzie, D.A.; Tailford, L.E.; Hemmings, A.M.; Juge, N. Crystal Structure of a Mucus-Binding Protein Repeat Reveals an Unexpected Functional Immunoglobulin Binding Activity. J. Biol. Chem. 2009, 284, 32444–32453. [Google Scholar] [CrossRef] [Green Version]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A Human Gut Microbial Gene Catalogue Established by Metagenomic Sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramare, F.; Nicoli, J.; Dabard, J.; Corring, T.; Ladire, M.; Gueugneau, A.M.; Raibaud, P. Trypsin-Dependent Production of an Antibacterial Substance by a Human Peptostreptococcus Strain in Gnotobiotic Rats and in Vitro. Appl. Environ. Microbiol. 1993, 59, 2876–2883. [Google Scholar] [CrossRef] [Green Version]
- Gomez, A.; Ladiré, M.; Marcille, F.; Fons, M. Trypsin Mediates Growth Phase-Dependent Transcriptional Tegulation of Genes Involved in Biosynthesis of Ruminococcin A, a Lantibiotic Produced by a Ruminococcus Gnavus Strain from a Human Intestinal Microbiota. J. Bacteriol. 2002, 184, 18–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crost, E.H.; Ajandouz, E.H.; Villard, C.; Geraert, P.A.; Puigserver, A.; Fons, M. Ruminococcin C, a New Anti-Clostridium Perfringens Bacteriocin Produced in the Gut by the Commensal Bacterium Ruminococcus Gnavus E1. Biochimie 2011, 93, 1487–1494. [Google Scholar] [CrossRef]
- Midtvedt, T. Effects of Antimicrobial Agents upon the Functional Part of the Intestinal Flora. Scand. J. Infect. Dis. Suppl. 1986, 49, 85–88. [Google Scholar]
- Chiumento, S.; Roblin, C.; Kieffer-Jaquinod, S.; Tachon, S.; Leprètre, C.; Basset, C.; Aditiyarini, D.; Olleik, H.; Nicoletti, C.; Bornet, O.; et al. Ruminococcin C, a Promising Antibiotic Produced by a Human Gut Symbiont. Sci. Adv. 2019, 5, eaaw9969. [Google Scholar] [CrossRef] [Green Version]
- Roblin, C.; Chiumento, S.; Bornet, O.; Nouailler, M.; Müller, C.S.; Jeannot, K.; Basset, C.; Kieffer-Jaquinod, S.; Couté, Y.; Torelli, S.; et al. The Unusual Structure of Ruminococcin C1 Antimicrobial Peptide Confers Clinical Properties. Proc. Natl. Acad. Sci. USA 2020, 117, 19168–19177. [Google Scholar] [CrossRef]
- Graziani, F.; Pujol, A.; Nicoletti, C.; Dou, S.; Maresca, M.; Giardina, T.; Fons, M.; Perrier, J. Ruminococcus Gnavus E1 Modulates Mucin Expression and Intestinal Glycosylation. J. Appl. Microbiol. 2016, 120, 1403–1417. [Google Scholar] [CrossRef] [Green Version]
- Pujol, A.; Crost, E.H.; Simon, G.; Barbe, V.; Vallenet, D.; Gomez, A.; Fons, M. Characterization and Distribution of the Gene Cluster Encoding RumC, an Anti-Clostridium Perfringens Bacteriocin Produced in the Gut. FEMS Microbiol. Ecol. 2011, 78, 405–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zong, Y.; Xu, Y.; Liang, X.; Keene, D.R.; Höök, A.; Gurusiddappa, S.; Höök, M.; Narayana, S.V.L. A “Collagen Hug” Model for Staphylococcus Aureus CNA Binding to Collagen. EMBO J. 2005, 24, 4224–4236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chagnot, C.; Listrat, A.; Astruc, T.; Desvaux, M. Bacterial Adhesion to Animal Tissues: Protein Determinants for Recognition of Extracellular Matrix Components. Cell. Microbiol. 2012, 14, 1687–1696. [Google Scholar] [CrossRef]
- Mack, D.R.; Ahrne, S.; Hyde, L.; Wei, S.; Hollingsworth, M.A. Extracellular MUC3 Mucin Secretion Follows Adherence of Lactobacillus Strains to Intestinal Epithelial Cells in Vitro. Gut 2003, 52, 827–833. [Google Scholar] [CrossRef] [Green Version]
- Deivanayagam, C.C.; Rich, R.L.; Carson, M.; Owens, R.T.; Danthuluri, S.; Bice, T.; Höök, M.; Narayana, S.V. Novel Fold and Assembly of the Repetitive B Region of the Staphylococcus Aureus Collagen-Binding Surface Protein. Structure 2000, 8, 67–78. [Google Scholar] [CrossRef] [Green Version]
- Miljkovic, M.; Bertani, I.; Fira, D.; Jovcic, B.; Novovic, K.; Venturi, V.; Kojic, M. Shortening of the Paracasei Subsp. Paracasei BGNJ1-64 AggLb Protein Switches Its Activity from Auto-Aggregation to Biofilm Formation. Front. Microbiol. 2016, 7, 1422. [Google Scholar] [CrossRef] [Green Version]
- Crost, E.H.; Tailford, L.E.; Le Gall, G.; Fons, M.; Henrissat, B.; Juge, N. Utilisation of Mucin Glycans by the Human Gut Symbiont Ruminococcus Gnavus Is Strain-Dependent. PLoS ONE 2013, 8, e76341. [Google Scholar] [CrossRef] [Green Version]
- Bruel, L.; Sulzenbacher, G.; Cervera Tison, M.; Pujol, A.; Nicoletti, C.; Perrier, J.; Galinier, A.; Ropartz, D.; Fons, M.; Pompeo, F.; et al. α-Galactosidase/Sucrose Kinase (AgaSK), a Novel Bifunctional Enzyme from the Human Microbiome Coupling Galactosidase and Kinase Activities. J. Biol. Chem. 2011, 286, 40814–40823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguilera, M.; Rakotoarivonina, H.; Brutus, A.; Giardina, T.; Simon, G.; Fons, M. Aga1, the First Alpha-Galactosidase from the Human Bacteria Ruminococcus Gnavus E1, Efficiently Transcribed in Gut Conditions. Res. Microbiol. 2012, 163, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Cervera-Tison, M.; Tailford, L.E.; Fuell, C.; Bruel, L.; Sulzenbacher, G.; Henrissat, B.; Berrin, J.G.; Fons, M.; Giardina, T.; Juge, N. Functional Analysis of Family GH36 α-Galactosidases from Ruminococcus Gnavus E1: Insights into the Metabolism of a Plant Oligosaccharide by a Human Gut Symbiont. Appl. Environ. Microbiol. 2012, 78, 7720–7732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dabard, J.; Bridonneau, C.; Phillipe, C.; Anglade, P.; Molle, D.; Nardi, M.; Ladiré, M.; Girardin, H.; Marcille, F.; Gomez, A.; et al. Ruminococcin A, a New Lantibiotic Produced by a Ruminococcus Gnavus Strain Isolated from Human Feces. Appl. Environ. Microbiol. 2001, 67, 4111–4118. [Google Scholar] [CrossRef] [Green Version]
- Vallenet, D.; Calteau, A.; Dubois, M.; Amours, P.; Bazin, A.; Beuvin, M.; Burlot, L.; Bussell, X.; Fouteau, S.; Gautreau, G.; et al. MicroScope: An Integrated Platform for the Annotation and Exploration of Microbial Gene Functions through Genomic, Pangenomic and Metabolic Comparative Analysis. Nucleic Acids Res. 2020, 48, D579–D589. [Google Scholar] [CrossRef] [Green Version]
- Augeron, C.; Laboisse, C.L. Emergence of Permanently Differentiated Cell Clones in a Human Colonic Cancer Cell Line in Culture after Treatment with Sodium Butyrate. Cancer Res. 1984, 44, 3961–3969. [Google Scholar]
- Graziani, F.; Pinton, P.; Olleik, H.; Pujol, A.; Nicoletti, C.; Sicre, M.; Quinson, N.; Ajandouz, E.H.; Perrier, J.; Pasquale, E.D.; et al. Deoxynivalenol Inhibits the Expression of Trefoil Factors (TFF) by Intestinal Human and Porcine Goblet Cells. Arch. Toxicol. 2019, 93, 1039–1049. [Google Scholar] [CrossRef] [PubMed]
- Ajandouz, E.H.; Berdah, S.; Moutardier, V.; Bege, T.; Birnbaum, D.J.; Perrier, J.; Di Pasquale, E.; Maresca, M. Hydrolytic Fate of 3/15-Acetyldeoxynivalenol in Humans: Specific Deacetylation by the Small Intestine and Liver Revealed Using in Vitro and Ex Vivo Approaches. Toxins 2016, 8, 232. [Google Scholar] [CrossRef] [PubMed]
- Razafimanjato, H.; Benzaria, A.; Taïeb, N.; Guo, X.-J.; Vidal, N.; Di Scala, C.; Varini, K.; Maresca, M. The Ribotoxin Deoxynivalenol Affects the Viability and Functions of Glial Cells. Glia 2011, 59, 1672–1683. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Coated protein | Bmax | Km (nM) |
|---|---|---|
| Collagen I | 0.82 ± 0.11 | 161.9 ± 23.81 |
| Collagen IV | 1.16 ± 0.14 | 112.5 ± 21.86 |
| Human IgG | 3.92 ± 0.47 | 13.75 ± 0.34 |
| Human IgA | 2.14 ± 0.46 | 24.71 ± 1.86 |
| Goat IgG | 1.05 ± 0.06 | 58.89 ± 9.90 |
| Rabbit IgG | 0.69 ± 0.32 | 34.22 ± 5.61 |
| Mouse IgG | 0.38 ± 0.11 | 39.82 ± 7.21 |
| Mucin type I | 0.08 ± 0.02 | 110.1 ± 30.12 |
| Mucin type II | 2.56 ± 0.23 | 16.61 ± 0.46 |
| Mucin type III | 1.06 ± 0.38 | 78.52 ± 6.43 |
| Gelatin | 0.38 ± 0.13 | 66.18 ± 18.49 |
| Monosaccharide | IC50 (mM) | % of Inhibition at 500 mM |
|---|---|---|
| Frc | 23.0 ± 8.44 | 1.61 ± 2.18 |
| Fuc | 29.04 ± 8.36 | 2.73 ± 2.42 |
| Gal | 937.01 ± 173.38 | 34.68 ± 2.73 |
| GalNAc | 506.12 ± 139.19 | 51.61 ± 3.41 |
| Glc | 5.28 ± 1.06 | 10.75 ± 6.38 |
| GlcNAc | 25.98 ± 4.52 | 2.18 ± 4.36 |
| Man | 14.00 ± 2.89 | 2.82 ± 1.62 |
| Sialic acid | 1685.32 ± 576.84 | 17.94 ± 5.81 |
| Primer | Sequence |
|---|---|
| Ol35-Eco | 5′-CCGGAATTCTTAGAACAGTCAGAGAATAAAGCG-3′ |
| Ol252-Xho | 5′-CCGCTCGAGATTTAAATATTCTCCGGTAAGATCACCCGG-3′ |
| Ad1F | 5′-CGGCTTCTGATTTTAAAGGGATTAC-3′ |
| Ad2R | 5′-GTTTTGCACTTGGCTCTTCA-3′ |
| Ad2F | 5′-GAAGAGCCAAGTGCAAAAG-3′ |
| Ad3R | 5′-CTGAAAGGTGTGTGTAAAAGTG-3′ |
| Ad3F | 5′-CACTTTTACACACACCTTTCAG-3′ |
| Ad4R | 5′-GTCACCTCATTTAATGGAAG-3′ |
| Ad5F | 5′-CATGAGGAACAGGCTCCAAT-3′ |
| Ad6R | 5′-TCTTTGTGCGTCTGATTCTC-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maresca, M.; Alatou, R.; Pujol, A.; Nicoletti, C.; Perrier, J.; Giardina, T.; Simon, G.; Méjean, V.; Fons, M. RadA, a MSCRAMM Adhesin of the Dominant Symbiote Ruminococcus gnavus E1, Binds Human Immunoglobulins and Intestinal Mucins. Biomolecules 2021, 11, 1613. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11111613
Maresca M, Alatou R, Pujol A, Nicoletti C, Perrier J, Giardina T, Simon G, Méjean V, Fons M. RadA, a MSCRAMM Adhesin of the Dominant Symbiote Ruminococcus gnavus E1, Binds Human Immunoglobulins and Intestinal Mucins. Biomolecules. 2021; 11(11):1613. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11111613
Chicago/Turabian StyleMaresca, Marc, Radia Alatou, Ange Pujol, Cendrine Nicoletti, Josette Perrier, Thierry Giardina, Gwenola Simon, Vincent Méjean, and Michel Fons. 2021. "RadA, a MSCRAMM Adhesin of the Dominant Symbiote Ruminococcus gnavus E1, Binds Human Immunoglobulins and Intestinal Mucins" Biomolecules 11, no. 11: 1613. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11111613