
Selective Biological Effects of Selenium-Enriched Polysaccharide (Se-Le-30) Isolated from Lentinula edodes Mycelium on Human Immune Cells

 , , , and
, , , and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Biosynthesis and Extraction of Se-Enriched Polysaccharide Fraction (Se-Le-30)
2.2. Se-Le-30 Fraction Structure Examination
2.3. The Analysis of the Biological Activity of Se-Le-30
2.3.1. The Effects of Se-Le-30 on Mitogen-Stimulated Human Peripheral Blood Mononuclear Cells (PBMCs) Proliferation
2.3.2. The Effects of Se-Le-30 on Alloantigen-Stimulated Human Peripheral Blood Mononuclear Cells (PBMCs) Proliferation in Mixed Lymphocyte Reaction (MLR)
2.3.3. The Effects of Se-Le-30 on Cytotoxicity of Natural Killer (NK) Cells
2.3.4. The Effects of Se-Le-30 on TNF-α, IFN-γ, TGF-β, and IL-10 Production by CD3+ T Cells
2.3.5. The Effects of Se-Le-30 on Superoxide Production by Granulocytes
2.4. Statistical Analysis
3. Results
3.1. Structural Analysis
3.2. Biological Activity of Se-Le-30
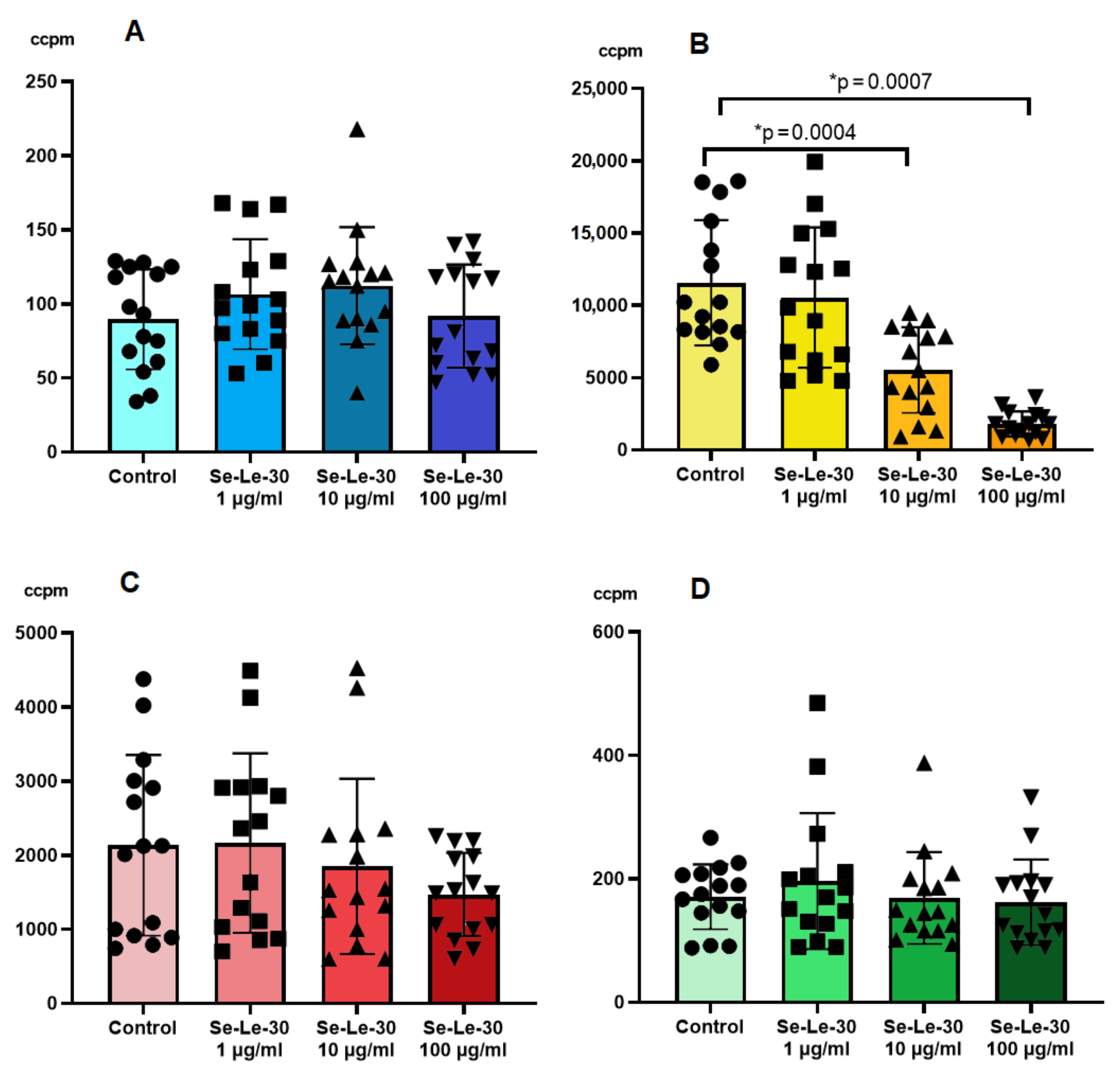
3.2.1. The Effect of Se-Le-30 on Proliferation of the Mitogen-Stimulated Human Peripheral Blood Mononuclear Cells (PBMCs)
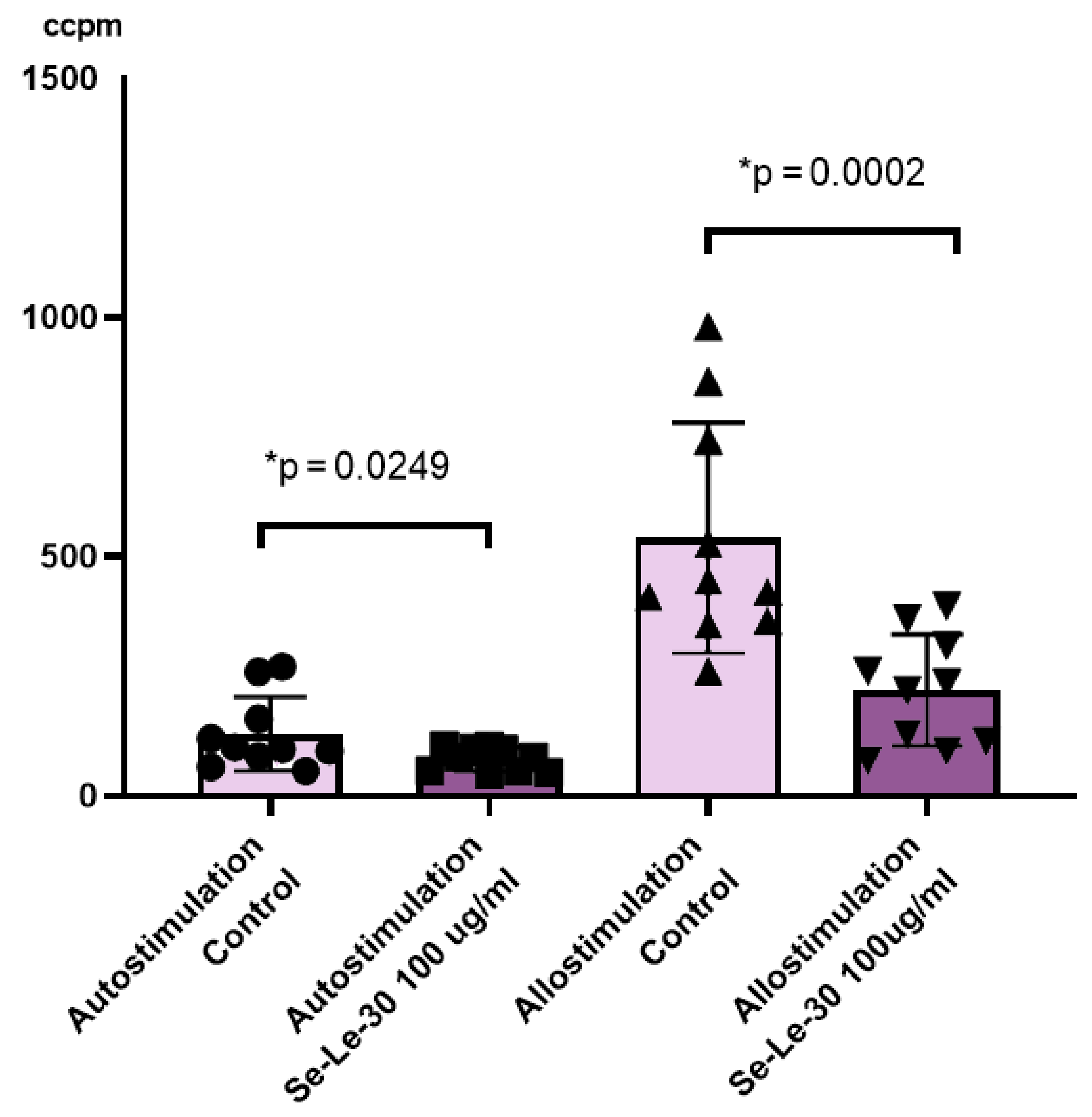
3.2.2. The Effect of Se-Le-30 on Alloantigen-Stimulated Human Peripheral Blood Mononuclear Cells (PBMCs) Proliferation in Mixed Lymphocyte Reaction (MLR)
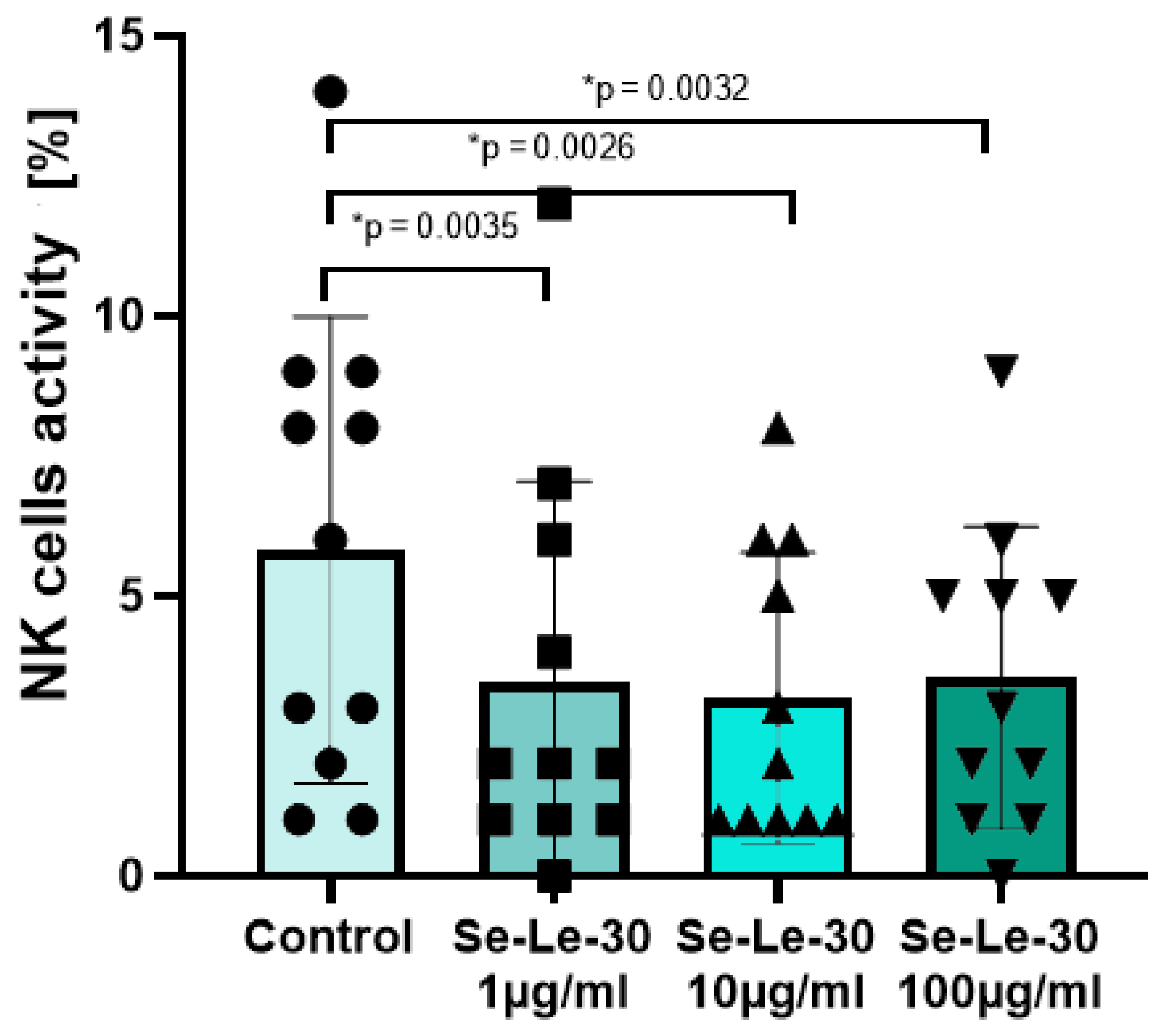
3.2.3. The Effect of Se-Le-30 on Cytotoxicity of Natural Killer (NK) Cells
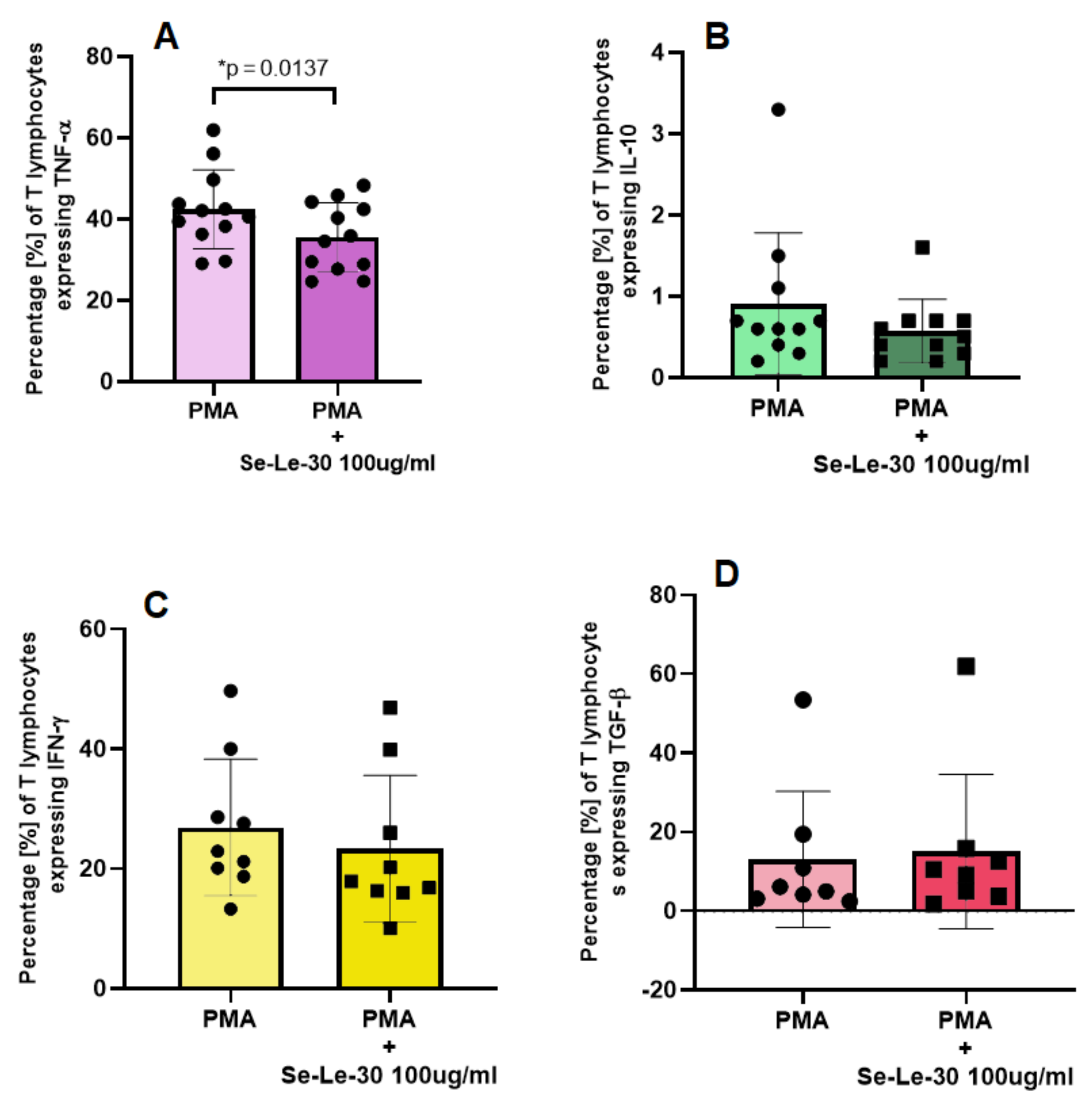
3.2.4. The Effect of Se-Le-30 on Tumor Necrosis Factor (TNF)-α, Interleukin (IL)-10, Interferon (INF)-γ and Transforming Growth Factor (TGF)-β Production by CD3+ T Cells
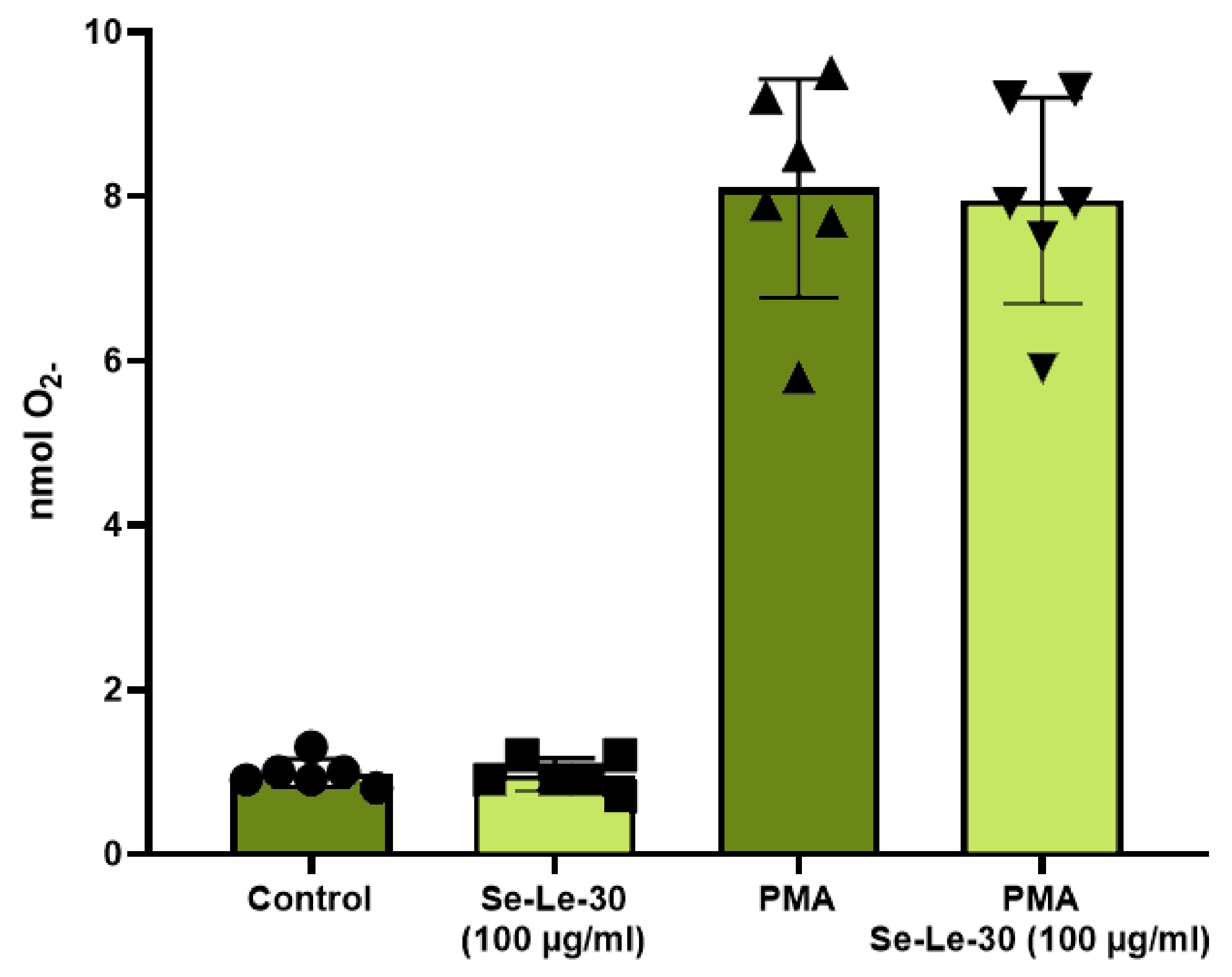
3.2.5. The Effect of Se-Le-30 on Superoxide Anion (O2-) Production by Granulocytes
4. Discussion
5. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Elsayed, E.A.; El Enshasy, H.; Wadaan, M.A.M.; Aziz, R. Mushrooms: A Potential Natural Source of Anti-Inflammatory Compounds for Medical Applications. Mediat. Inflamm. 2014, 2014, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Wasser, S.P. Medicinal mushrooms as a source of antitumor and immunomodulating polysaccharides. Appl. Microbiol. Biotechnol. 2002, 60, 258–274. [Google Scholar] [CrossRef]
- Borchers, A.T.; Keen, C.L.; Gershwin, M.E. Mushrooms, Tumors, and Immunity: An Update. Exp. Biol. Med. 2004, 229, 393–406. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Shen, M.; Song, Q.; Xie, J. Biological activities and pharmaceutical applications of polysaccharide from natural resources: A review. Carbohydr. Polym. 2018, 183, 91–101. [Google Scholar] [CrossRef]
- Zheng, R.; Jie, S.; Hanchuan, D.; Moucheng, W. Characterization and immunomodulating activities of polysaccharide from Lentinus edodes. Int. Immunopharmacol. 2005, 5, 811–820. [Google Scholar] [CrossRef]
- Ferreira, S.; Passos, C.; Madureira, P.; Vilanova, M.; Coimbra, M.A. Structure–function relationships of immunostimulatory polysaccharides: A review. Carbohydr. Polym. 2015, 132, 378–396. [Google Scholar] [CrossRef]
- Sullivan, R.; Smith, J.E.; Rowan, N.J. Medicinal Mushrooms and Cancer Therapy: Translating a traditional practice into Western medicine. Perspect. Biol. Med. 2006, 49, 159–170. [Google Scholar] [CrossRef]
- Mahajna, J.A.; Yassin, M.; Wasser, S.P. Mushrooms Extracts Having Anticancer Activity. US Patent 7 258 862 B2, 21 August 2007. [Google Scholar]
- Kataoka, H.; Shimura, T.; Mizoshita, T.; Kubota, E.; Mori, Y.; Mizushima, T.; Wada, T.; Ogasawara, N.; Tanida, S.; Sasaki, M.; et al. Lentinan with S-1 and paclitaxel for gastric cancer chemotherapy improve patient quality of life. Hepatogastroenterology 2009, 56, 547–550. [Google Scholar]
- Higashi, D.; Seki, K.; Ishibashi, Y.; Egawa, Y.; Koga, M.; Sasaki, T.; Hirano, K.; Mikami, K.; Futami, K.; Maekawa, T.; et al. The effect of lentinan combination therapy for unresectable advanced gastric cancer. Anticancer. Res. 2012, 32, 2365–2368. [Google Scholar]
- Spallholz, J.E.; Boylan, L.M.; Larsen, H.S. Advances in Understanding Selenium’s Role in the Immune System. Ann. N. Y. Acad. Sci. 1990, 587, 123–139. [Google Scholar] [CrossRef]
- Hawkes, W.C.; Kelley, D.S.; Taylor, P.C. The Effects of Dietary Selenium on the Immune System in Healthy Men. Biol. Trace Elem. Res. 2001, 81, 189–213. [Google Scholar] [CrossRef]
- Hoffmann, P.R.; Berry, M.J. The influence of selenium on immune responses. Mol. Nutr. Food Res. 2008, 52, 1273–1280. [Google Scholar] [CrossRef]
- Rayman, M.P. Selenium and human health. Lancet 2012, 379, 1256–1268. [Google Scholar] [CrossRef]
- Malinowska, E.; Klimaszewska, M.; Strączek, T.; Schneider, K.; Kapusta, C.; Podsadni, P.; Łapienis, G.; Dawidowski, M.; Kleps, J.; Górska, S.; et al. Selenized polysaccharides—Biosynthesis and structural analysis. Carbohydr. Polym. 2018, 198, 407–417. [Google Scholar] [CrossRef]
- Kaleta, B.; Górski, A.; Zagożdżon, R.; Cieślak, M.; Kaźmierczak-Barańska, J.; Nawrot, B.; Klimaszewska, M.; Malinowska, E.; Górska, S.; Turło, J. Selenium-containing polysaccharides from Lentinula edodes—Biological activity. Carbohydr. Polym. 2019, 223, 115078. [Google Scholar] [CrossRef]
- Chihara, G.; Hamuro, J.; Maeda, Y.; Arai, Y.; Fukuoka, F. Fractionation and purification of the polysaccharides with marked antitumor activity, especially lentinan, from Lentinus edodes (Berk.) Sing. (an edible mushroom). Cancer Res. 1970, 30, 2776–2781. [Google Scholar]
- Klimaszewska, M.; Górska, S.; Łapienis, G.; Kaleta, B.; Górska, S.; Kaszowska, M.; Dawidowski, M.; Gamian, A.; Zagożdżon, R.; Górski, A.; et al. Identification of the Primary Structure of Selenium-Containing Polysaccharides Selectively Inhibiting T-Cell Proliferation. Molecules 2021, 26, 5404. [Google Scholar] [CrossRef]
- Turło, J.; Gutkowska, B.; Herold, F. Effect of selenium enrichment on antioxidant activities and chemical composition of Lentinula edodes (Berk.) Pegl. mycelial extracts. Food Chem. Toxicol. 2010, 48, 1085–1091. [Google Scholar] [CrossRef]
- Turło, J.; Gutkowska, B.; Herold, F.; Gajzlerska, W.; Dawidowski, M.; Dorociak, A.; Zobel, A. Biological Availability and Preliminary Selenium Speciation in Selenium-Enriched Mycelium of Lentinula edodes (Berk.). Food Biotechnol. 2011, 25, 16–29. [Google Scholar] [CrossRef]
- Cheong, K.-L.; Wu, D.-T.; Zhao, J.; Li, S.-P. A rapid and accurate method for the quantitative estimation of natural polysaccharides and their fractions using high performance size exclusion chromatography coupled with multi-angle laser light scattering and refractive index detector. J. Chromatogr. A 2015, 1400, 98–106. [Google Scholar] [CrossRef]
- Yap, A.-T.; Ng, M.-L. An Improved Method for the Isolation of Lentinan from the Edible and Medicinal Shiitake Mushroom, Lentinus edodes (Berk.) Sing. (Agaricomycetideae). Int. J. Med. Mushrooms 2001, 3, 11. [Google Scholar] [CrossRef]
- Wang, S.-X.; Liu, Q.-Y.; Li, Y. Lentinan ameliorates burn sepsis by attenuating CD4 + CD25 + Tregs. Burns 2016, 42, 1513–1521. [Google Scholar] [CrossRef]
- Fruehauf, J.P.; Bonnard, G.D.; Herberman, R.B. The effect of lentinan on production of interleukin-1 by human monocytes. Immunopharmacology 1982, 5, 65–74. [Google Scholar] [CrossRef]
- Chihara, G. Immunopharmacology of Lentinan, a polysaccharide isolated from Lentinus edodes: Its application as a host defense potentiator. Int. J. Oriental Med. 1992, 17, 57–77. [Google Scholar]
- Izawa, M.; Ohno, K.; Amikura, K.; Hamuro, J. Lentinan augments the production of Interleukin 3 and colony stimulating factor(s). In Manipulation of Host Defence Mechanisms; Aoki, T., Ed.; Excerpta Medica: Amsterdam, The Netherlands, 1984; p. 59. [Google Scholar]
- Maeda, Y.Y.; Sakaizumi, M.; Moriwaki, K.; Chihara, G.; Yonekawa, H. Genetical control on lentinan-induced acute phase responses and vascular responses. Folia Histochem. Cytobiol. 1992, 30, 207–209. [Google Scholar]
- Wang, X.-E.; Wang, Y.-H.; Zhou, Q.; Peng, M.; Zhang, J.; Chen, M.; Ma, L.-J.; Xie, G.-M. Immunomodulatory Effect of Lentinan on Aberrant T Subsets and Cytokines Profile in Non-small Cell Lung Cancer Patients. Pathol. Oncol. Res. 2018, 26, 499–505. [Google Scholar] [CrossRef]
- Hobbs, C. Medicinal Value of Lentinus edodes (Berk.) Sing. (Agaricomycetideae). A Literature Review. Int. J. Med. Mushrooms 2000, 2, 287–302. [Google Scholar] [CrossRef]
- Mian, M.F.; Lauzon, N.M.; Andrews, D.; Lichty, B.D.; Ashkar, A.A. FimH Can Directly Activate Human and Murine Natural Killer Cells via TLR4. Mol. Ther. 2010, 18, 1379–1388. [Google Scholar] [CrossRef]
- Tani, M.; Tanimura, H.; Yamaue, H.; Iwahashi, M.; Tsunoda, T.; Tamai, M.; Noguchi, K.; Arii, K. In vitro generation of activated natural killer cells and cytotoxic macrophages with lentinan. Eur. J. Clin. Pharmacol. 1992, 42, 623–627. [Google Scholar] [CrossRef]
- Huyan, T.; Li, Q.; Yang, H.; Jin, M.-L.; Zhang, M.-J.; Ye, L.-J.; Li, J.; Huang, Q.-S.; Yin, D.-C. Protective effect of polysaccharides on simulated microgravity-induced functional inhibition of human NK cells. Carbohydr. Polym. 2014, 101, 819–827. [Google Scholar] [CrossRef]
- Amino, M.; Noguchi, R.; Yata, J.; Matsumura, J.; Hirayama, R.; Abe, O.; Enomoto, K.; Asato, Y. Studies on the effect of lentinan on human immune system. II. In vivo effect on NK activity, MLR induced killer activity and PHA induced blastic response of lymphocytes in cancer patients. Gan Kagaku Ryoho. Cancer Chemother. 1983, 10, 2000–2006. [Google Scholar]
- Péater, G.; Káaroly, V.; Imre, B.; Jáanos, F.; Kaneko, Y. Effects of Lentinan on Cytotoxic Functions of Human Lymphocytes. Immunopharmacol. Immunotoxicol. 1988, 10, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Taylor, P.; Brown, G.; Reid, D.M.; Willment, J.; Martinez-Pomares, L.; Gordon, S.; Wong, S.Y.C. The β-Glucan Receptor, Dectin-1, Is Predominantly Expressed on the Surface of Cells of the Monocyte/Macrophage and Neutrophil Lineages. J. Immunol. 2002, 169, 3876–3882. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaleta, B.; Roszczyk, A.; Zych, M.; Kniotek, M.; Zagożdżon, R.; Klimaszewska, M.; Malinowska, E.; Pac, M.; Turło, J. Selective Biological Effects of Selenium-Enriched Polysaccharide (Se-Le-30) Isolated from Lentinula edodes Mycelium on Human Immune Cells. Biomolecules 2021, 11, 1777. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11121777
Kaleta B, Roszczyk A, Zych M, Kniotek M, Zagożdżon R, Klimaszewska M, Malinowska E, Pac M, Turło J. Selective Biological Effects of Selenium-Enriched Polysaccharide (Se-Le-30) Isolated from Lentinula edodes Mycelium on Human Immune Cells. Biomolecules. 2021; 11(12):1777. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11121777
Chicago/Turabian StyleKaleta, Beata, Aleksander Roszczyk, Michał Zych, Monika Kniotek, Radosław Zagożdżon, Marzenna Klimaszewska, Eliza Malinowska, Michał Pac, and Jadwiga Turło. 2021. "Selective Biological Effects of Selenium-Enriched Polysaccharide (Se-Le-30) Isolated from Lentinula edodes Mycelium on Human Immune Cells" Biomolecules 11, no. 12: 1777. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11121777