
A High Throughput Lipidomics Method Using Scheduled Multiple Reaction Monitoring

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Lipid Extraction from Human Plasma
2.3. Liquid Chromatography-Mass Spectrometry
2.4. Method Development
2.5. Variable Retention Time Window (Variable-RTW) and Relative Dwell Time Weightage (Relative DTW)
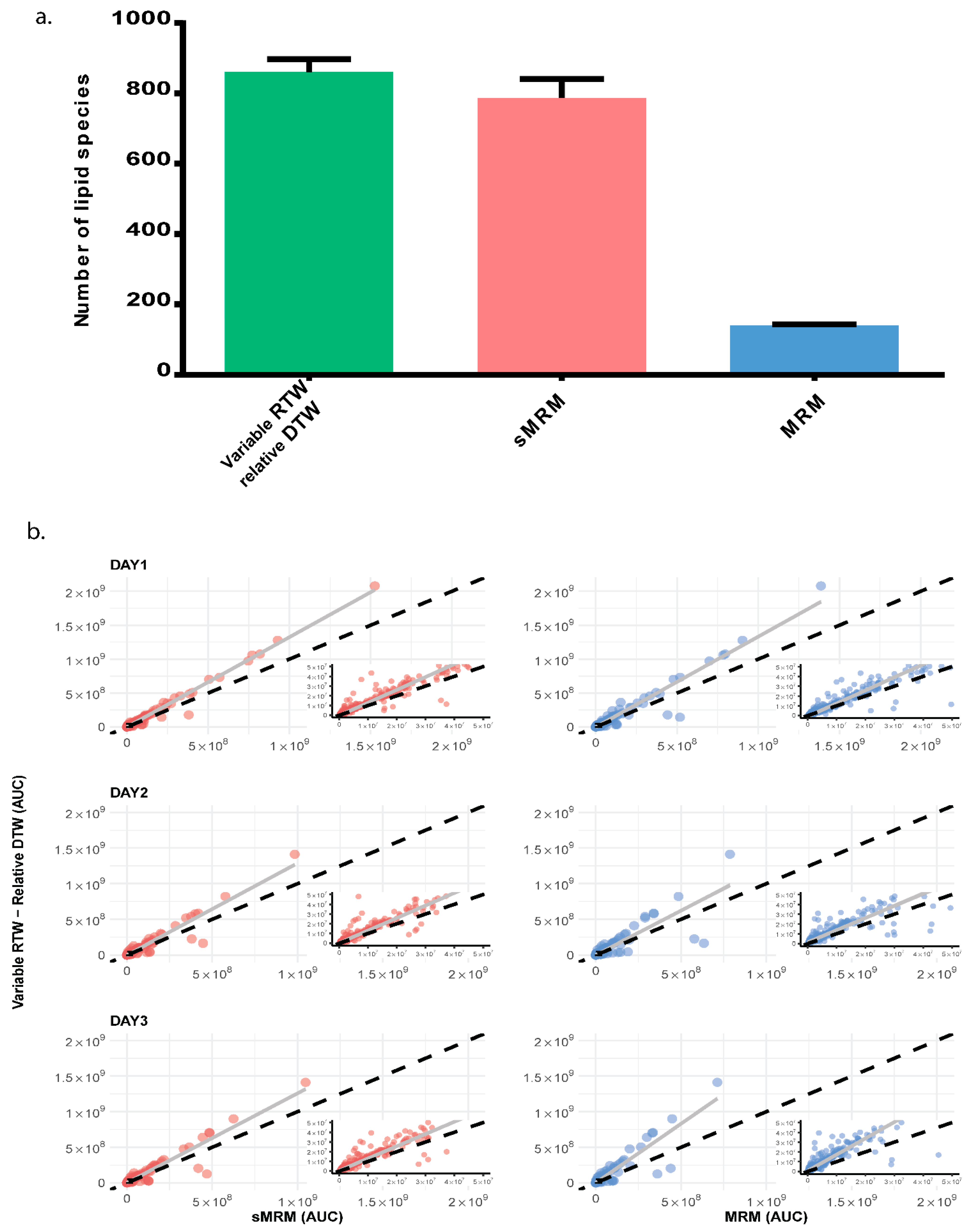
2.6. Comparison of MRM, Scheduled MRM and Scheduled MRM with Variable Retention Time Window and Relative Dwell Time Weightage
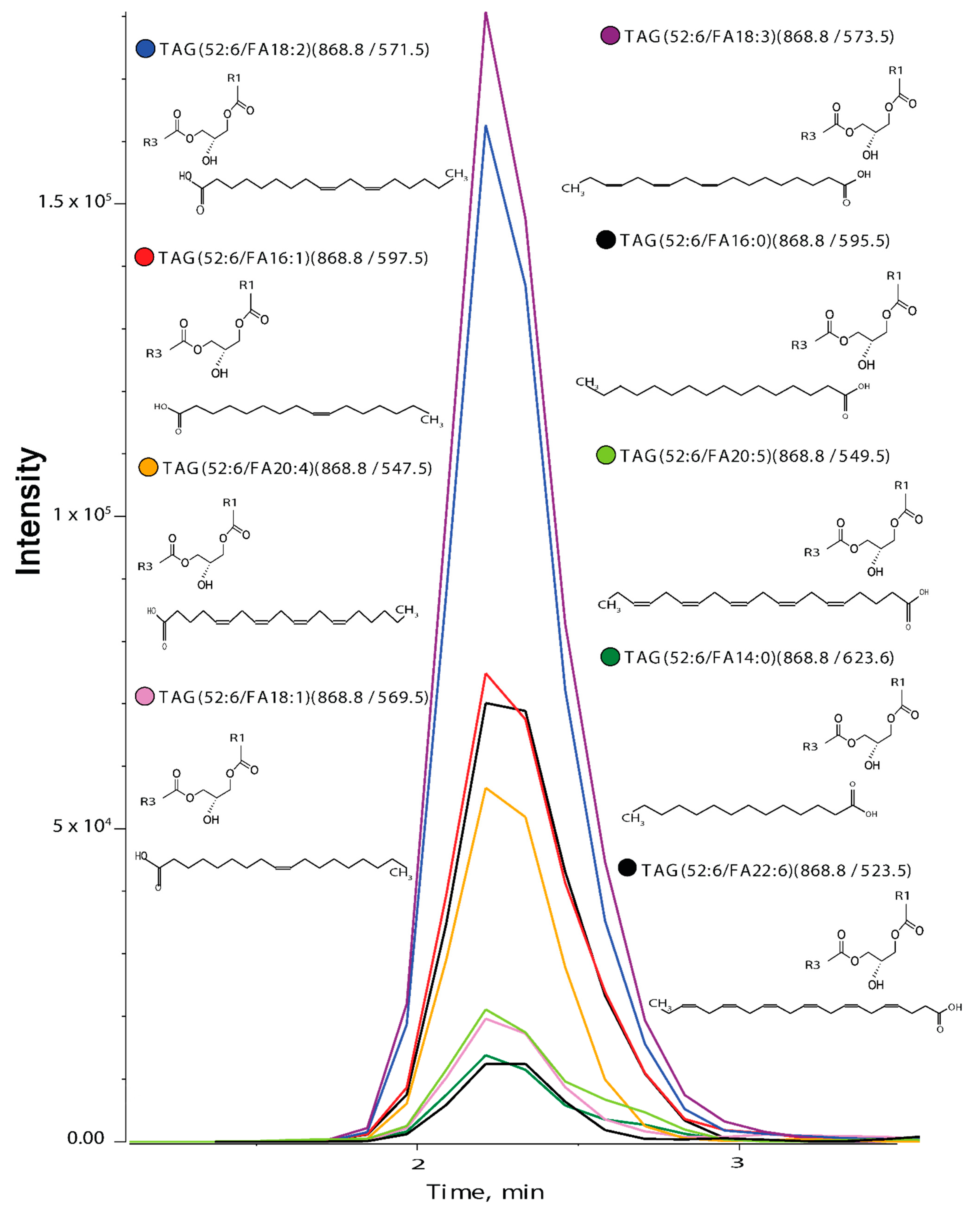
2.7. Identification of Isomers within Lipid Classes
- There was less than 10% missing values and these were imputed with the median value of the particular lipid species across the samples.
- Based on the sum of the peak areas of all TAG/PL species, the species abundance and the isomer abundance within that species were calculated (Supplementary Tables S3 and S4).
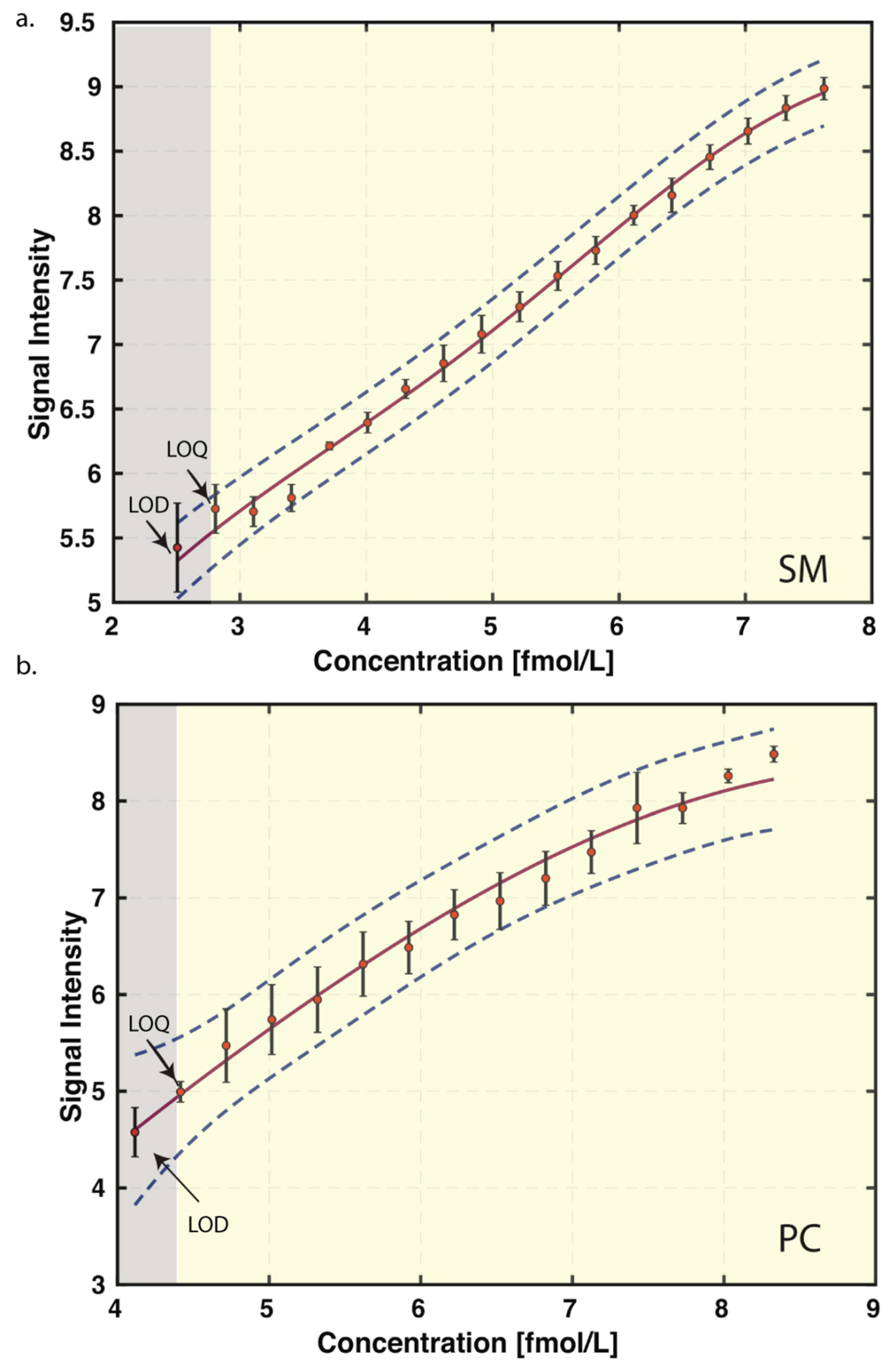
2.8. Limit of Detection and Quantitation
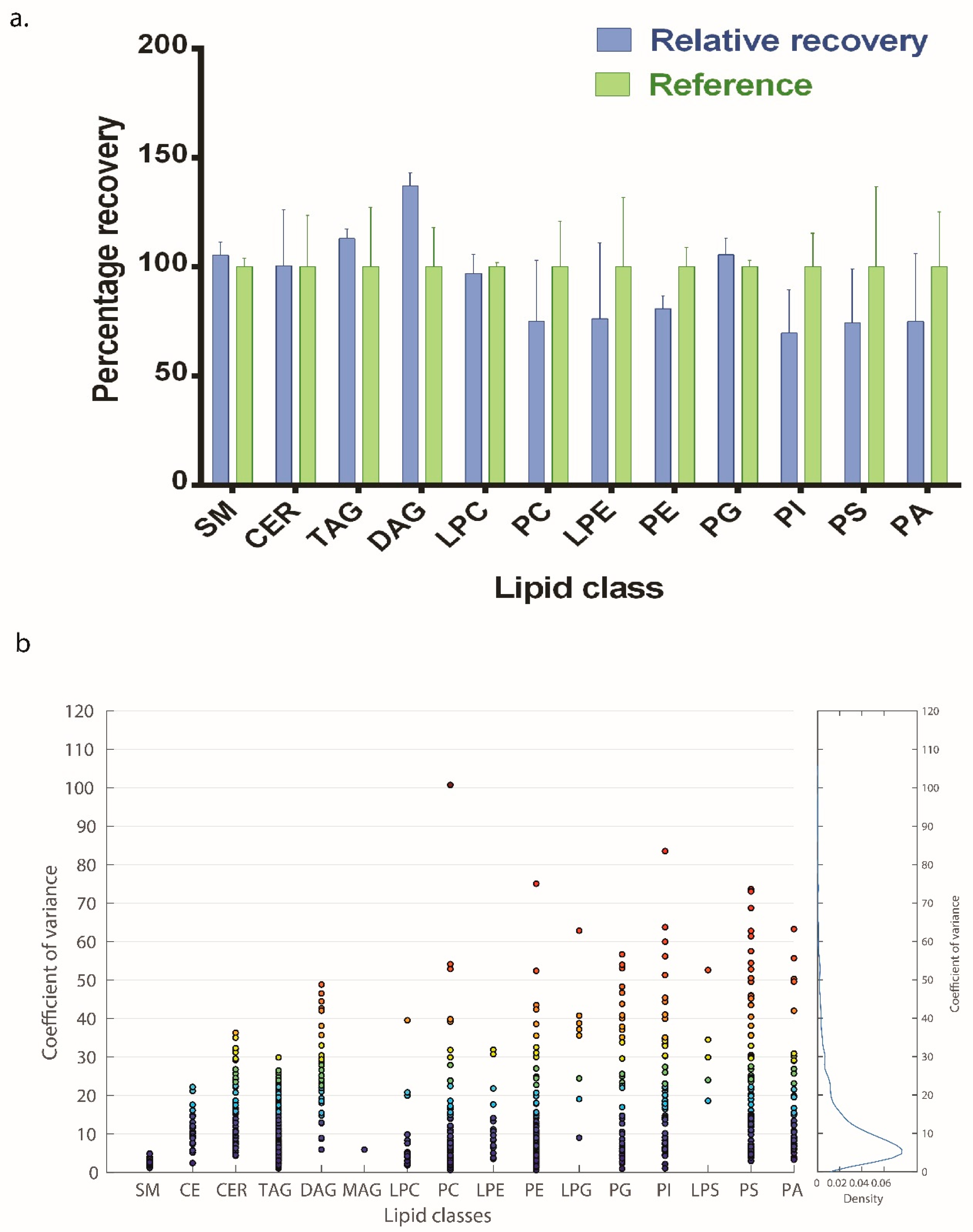
2.9. Spike and Recovery and Coefficient of Variance
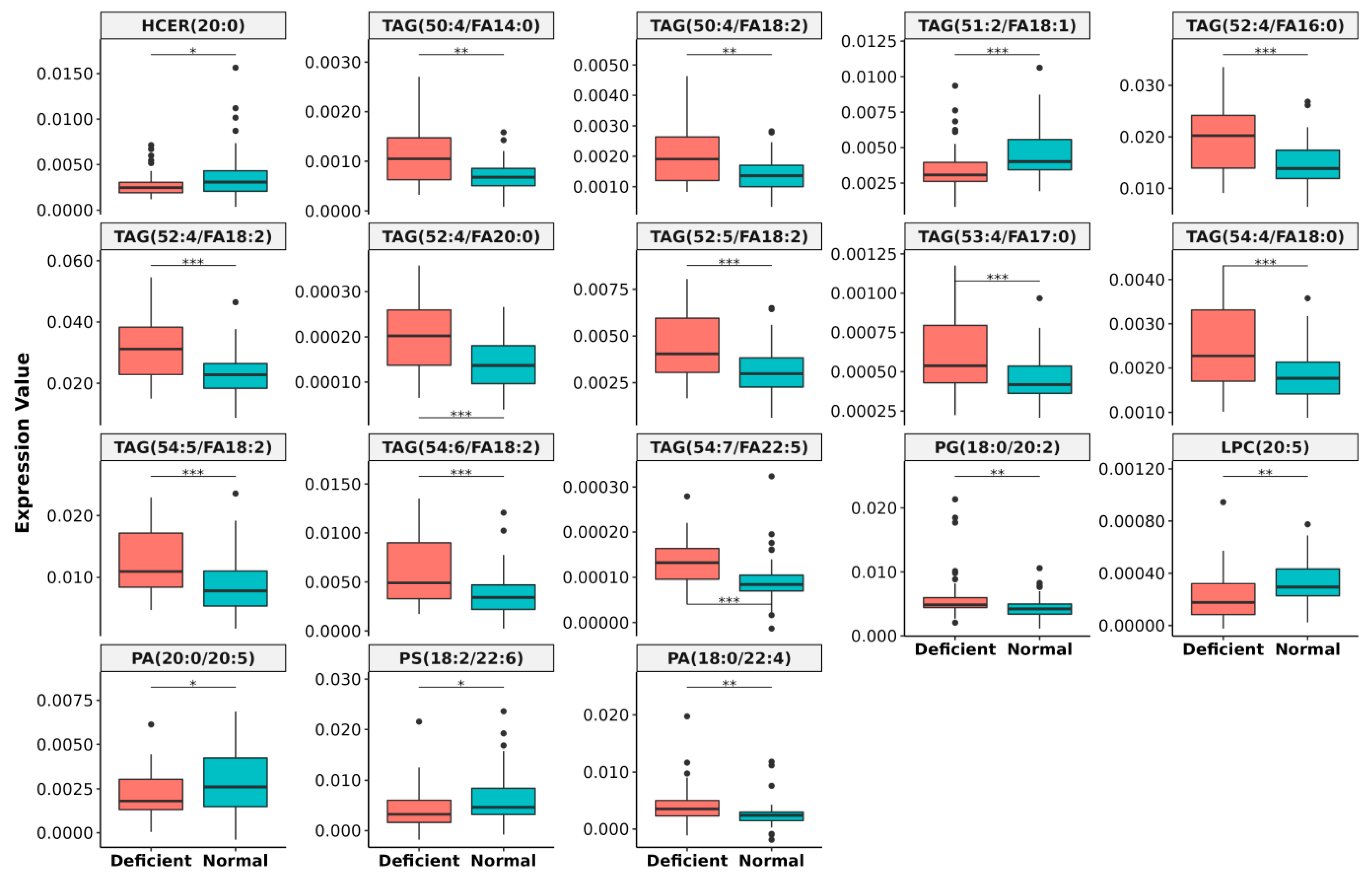
2.10. Alteration of Plasma Lipids Due to Vitamin B12 Deficiency
- Imputation was carried out for less than 1% missing data in the dataset with median value based on lipid species specific value across the two groups.
- Batch effect correction for 759 lipid species was carried out to reduce technical variance and remove sex bias using the ComBat function from the SVA package [42].
- Differential expression (DE) analysis was performed by two-sided Wilcoxon rank-sum test, where 55 lipid species were found to be altered by more than 1.3 folds with p < 0.05 between the two groups (Supplementary Table S7).
- For important feature selection, Boruta algorithm using random forest was applied to identify possible lipid markers and iteratively removes lipid species that were statistically less relevant than a random probe between the two groups [42]. The ‘Boruta’ function from the R library package was applied on 93 samples with 55 lipids as features, where 18 lipids were finally selected as putative markers.
- A two-sample Wilcoxon test with p-value adjustment (BH method) was further applied to show the significant relationship between the categories of identified lipid markers.
2.11. Data Analysis
3. Results
3.1. Identification of Isomers within Lipid Classes
3.2. Limit of Blank (LoB), Limit of Detection (LoD), Limit of Quantitation (LoQ), and Linear Range
3.3. Spike and Recovery and Coefficient of Variation
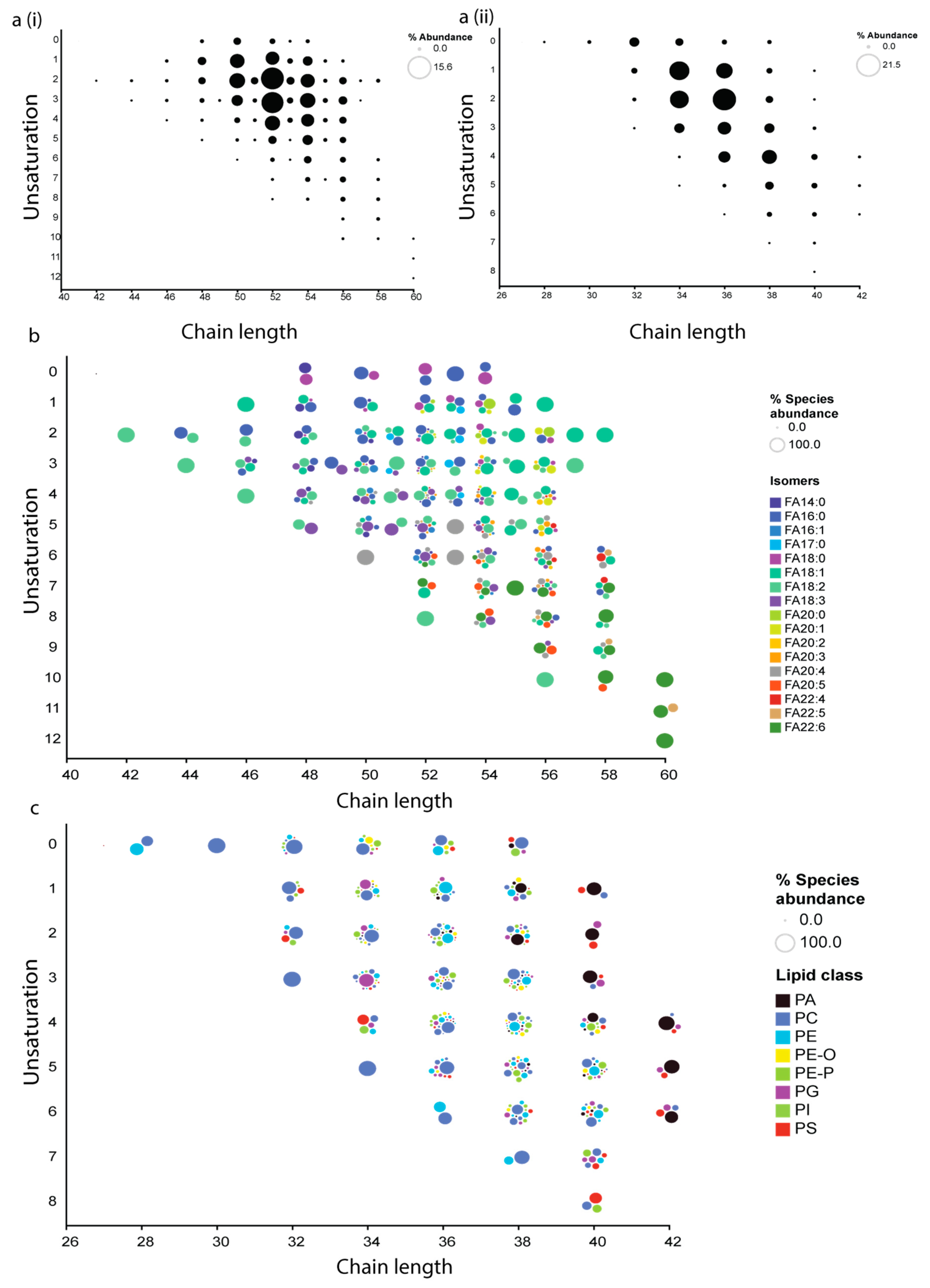
3.4. Lipidomics Study in Normal and Vitamin B12 Deficient Human Plasma
4. Discussion
Lipidomics Study in Normal and Vitamin B12 Deficient Human Plasma
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smilowitz, J.T.; Zivkovic, A.M.; Wan, Y.-J.Y.; Watkins, S.M.; Nording, M.L.; Hammock, B.D.; German, J.B. Nutritional Lipidomics: Molecular Metabolism, Analytics, and Diagnostics. Mol. Nutr. Food Res. 2013, 57, 1319–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muro, E.; Atilla-Gokcumen, G.E.; Eggert, U.S. Lipids in Cell Biology: How Can We Understand Them Better? MBoC 2014, 25, 1819–1823. [Google Scholar] [CrossRef] [PubMed]
- Yáñez-Mó, M.; Siljander, P.R.-M.; Andreu, Z.; Zavec, A.B.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological Properties of Extracellular Vesicles and Their Physiological Functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Meer, G.; Voelker, D.R.; Feigenson, G.W. Membrane Lipids: Where They Are and How They Behave. Nat. Rev. Mol. Cell Biol. 2008, 9, 112–124. [Google Scholar] [CrossRef]
- Glomset, J.A. Protein-Lipid Interactions on the Surfaces of Cell Membranes. Curr. Opin. Struct. Biol. 1999, 9, 425–427. [Google Scholar] [CrossRef]
- Ye, R.; Onodera, T.; Scherer, P.E. Lipotoxicity and β Cell Maintenance in Obesity and Type 2 Diabetes. J. Endocr. Soc. 2019, 3, 617–631. [Google Scholar] [CrossRef] [Green Version]
- Fu, S.; Yang, L.; Li, P.; Hofmann, O.; Dicker, L.; Hide, W.; Lin, X.; Watkins, S.M.; Ivanov, A.R.; Hotamisligil, G.S. Aberrant Lipid Metabolism Disrupts Calcium Homeostasis Causing Liver Endoplasmic Reticulum Stress in Obesity. Nature 2011, 473, 528–531. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Zhang, Y.; Ren, J. Autophagic Regulation of Lipid Homeostasis in Cardiometabolic Syndrome. Front. Cardiovasc. Med. 2018, 5, 38. [Google Scholar] [CrossRef] [Green Version]
- Beloribi-Djefaflia, S.; Vasseur, S.; Guillaumond, F. Lipid Metabolic Reprogramming in Cancer Cells. Oncogenesis 2016, 5, e189. [Google Scholar] [CrossRef]
- Wymann, M.P.; Schneiter, R. Lipid Signalling in Disease. Nat. Rev. Mol. Cell Biol. 2008, 9, 162–176. [Google Scholar] [CrossRef]
- Quehenberger, O.; Dennis, E.A. The Human Plasma Lipidome. N. Engl. J. Med. 2011, 365, 1812–1823. [Google Scholar] [CrossRef] [PubMed]
- Shevchenko, A.; Simons, K. Lipidomics: Coming to Grips with Lipid Diversity. Nat. Rev. Mol. Cell Biol. 2010, 11, 593–598. [Google Scholar] [CrossRef] [PubMed]
- Sud, M.; Fahy, E.; Cotter, D.; Brown, A.; Dennis, E.A.; Glass, C.K.; Merrill, A.H., Jr.; Murphy, R.C.; Raetz, C.R.H.; Russell, D.W.; et al. LMSD: LIPID MAPS Structure Database. Nucleic Acids Res. 2007, 35, D527–D532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pradas, I.; Huynh, K.; Cabré, R.; Ayala, V.; Meikle, P.J.; Jové, M.; Pamplona, R. Lipidomics Reveals a Tissue-Specific Fingerprint. Front. Physiol. 2018, 9, 1165. [Google Scholar] [CrossRef] [Green Version]
- van Meer, G. Cellular Lipidomics. EMBO J. 2005, 24, 3159–3165. [Google Scholar] [CrossRef] [Green Version]
- Brügger, B.; Erben, G.; Sandhoff, R.; Wieland, F.T.; Lehmann, W.D. Quantitative Analysis of Biological Membrane Lipids at the Low Picomole Level by Nano-Electrospray Ionization Tandem Mass Spectrometry. Proc. Natl. Acad. Sci. USA 1997, 94, 2339–2344. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Shon, J.C.; Liu, K.-H. Mass Spectrometry-Based Lipidomics and Its Application to Biomedical Research. J. Lifestyle Med. 2014, 4, 17–33. [Google Scholar] [CrossRef] [Green Version]
- Wenk, M.R. The Emerging Field of Lipidomics. Nat. Rev. Drug Discov. 2005, 4, 594–610. [Google Scholar] [CrossRef]
- Han, X.; Gross, R.W. Global Analyses of Cellular Lipidomes Directly from Crude Extracts of Biological Samples by ESI Mass Spectrometry: A Bridge to Lipidomics. J. Lipid Res. 2003, 44, 1071–1079. [Google Scholar] [CrossRef] [Green Version]
- Kirkwood, J.S.; Maier, C.; Stevens, J.F. Simultaneous, Untargeted Metabolic Profiling of Polar and Nonpolar Metabolites by LC-Q-TOF Mass Spectrometry. Curr. Protoc. Toxicol. 2013, 56, 4–39. [Google Scholar] [CrossRef] [Green Version]
- Takeda, H.; Izumi, Y.; Takahashi, M.; Paxton, T.; Tamura, S.; Koike, T.; Yu, Y.; Kato, N.; Nagase, K.; Shiomi, M.; et al. Widely-Targeted Quantitative Lipidomics Method by Supercritical Fluid Chromatography Triple Quadrupole Mass Spectrometry. J. Lipid Res. 2018, 59, 1283–1293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Contrepois, K.; Mahmoudi, S.; Ubhi, B.K.; Papsdorf, K.; Hornburg, D.; Brunet, A.; Snyder, M. Cross-Platform Comparison of Untargeted and Targeted Lipidomics Approaches on Aging Mouse Plasma. Sci. Rep. 2018, 8, 17747. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.J.; Codreanu, S.G.; Goyal, S.; Wages, P.A.; Gorti, S.K.K.; Pearson, M.J.; Uribe, I.; Sherrod, S.D.; McLean, J.A.; Porter, N.A.; et al. Evaluating a Targeted Multiple Reaction Monitoring Approach to Global Untargeted Lipidomic Analyses of Human Plasma. Rapid Commun. Mass Spectrom. 2020, 34, e8911. [Google Scholar] [CrossRef] [PubMed]
- Dekker, B. Reduce Complexity by Choosing Your Reactions. Nat. Methods 2015, 12, 16. [Google Scholar] [CrossRef]
- Mao, C.; Xu, R.; Szulc, Z.M.; Bielawski, J.; Becker, K.P. Cloning and Characterization of a Mouse Endoplasmic Reticulum Alkaline Ceramidase: An Enzyme That Preferentially Regulates Metabolism of Very Long Chain. J. Biol. Chem. 2003, 278, 31184–31191. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Liu, X.; Wu, J.; Meehan, M.J.; Blevitt, J.M.; Dorrestein, P.C.; Milla, M.E. A Highly Efficient, High-Throughput Lipidomics Platform for the Quantitative Detection of Eicosanoids in Human Whole Blood. Anal. Biochem. 2013, 433, 181–188. [Google Scholar] [CrossRef]
- Weir, J.M.; Wong, G.; Barlow, C.K.; Greeve, M.A.; Kowalczyk, A.; Almasy, L.; Comuzzie, A.G.; Mahaney, M.C.; Jowett, J.B.M.; Shaw, J.; et al. Plasma Lipid Profiling in a Large Population-Based Cohort. J. Lipid Res. 2013, 54, 2898–2908. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Hu, C.; Zhao, X.; Dai, W.; Chen, S.; Lu, X.; Xu, G. Large-Scaled Human Serum Sphingolipid Profiling by Using Reversed-Phase Liquid Chromatography Coupled with Dynamic Multiple Reaction Monitoring of Mass Spectrometry: Method Development and Application in Hepatocellular Carcinoma. J. Chromatogr. A 2013, 1320, 103–110. [Google Scholar] [CrossRef]
- Liang, J.; Wu, W.-Y.; Sun, G.-X.; Wang, D.-D.; Hou, J.-J.; Yang, W.-Z.; Jiang, B.-H.; Liu, X.; Guo, D.-A. A Dynamic Multiple Reaction Monitoring Method for the Multiple Components Quantification of Complex Traditional Chinese Medicine Preparations: Niuhuang Shangqing Pill as an Example. J. Chromatogr. A 2013, 1294, 58–69. [Google Scholar] [CrossRef]
- Rao, Z.; Qin, H.; Wei, Y.; Zhou, Y.; Zhang, G.; Zhang, F.; Shao, Y.; Huang, J.; Wu, X.A. Development of a Dynamic Multiple Reaction Monitoring Method for Determination of Digoxin and Six Active Components of Ginkgo Biloba Leaf Extract in Rat Plasma. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2014, 959, 27–35. [Google Scholar] [CrossRef]
- Andrade, G.C.R.M.; Monteiro, S.H.; Francisco, J.G.; Figueiredo, L.A.; Botelho, R.G.; Tornisielo, V.L. Liquid Chromatography-Electrospray Ionization Tandem Mass Spectrometry and Dynamic Multiple Reaction Monitoring Method for Determining Multiple Pesticide Residues in Tomato. Food Chem. 2015, 175, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Jia, Z.-X.; Zhang, J.-L.; Shen, C.-P.; Ma, L. Profile and Quantification of Human Stratum Corneum Ceramides by Normal-Phase Liquid Chromatography Coupled with Dynamic Multiple Reaction Monitoring of Mass Spectrometry: Development of Targeted Lipidomic Method and Application to Human Stratum Corneum of Different Age Groups. Anal. Bioanal. Chem. 2016, 408, 6623–6636. [Google Scholar] [PubMed]
- Shah, I.; Petroczi, A.; Uvacsek, M.; Ránky, M.; Naughton, D.P. Hair-Based Rapid Analyses for Multiple Drugs in Forensics and Doping: Application of Dynamic Multiple Reaction Monitoring with LC-MS/MS. Chem. Cent. J. 2014, 8, 73. [Google Scholar] [CrossRef] [Green Version]
- Xu, G.; Amicucci, M.J.; Cheng, Z.; Galermo, A.G.; Lebrilla, C.B. Revisiting Monosaccharide Analysis-Quantitation of a Comprehensive Set of Monosaccharides Using Dynamic Multiple Reaction Monitoring. Analyst 2017, 143, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Achieve Broad Lipid Quantitation Using a High-Throughput Targeted Lipidomics Method. Available online: https://sciex.com (accessed on 15 March 2022).
- Armbruster, D.A.; Pry, T. Limit of Blank, Limit of Detection and Limit of Quantitation. Clin. Biochem. Rev. 2008, 29 (Suppl. 1), S49–S52. [Google Scholar] [PubMed]
- Armbruster, D.A.; Tillman, M.D.; Hubbs, L.M. Limit of Detection (LQD)/limit of Quantitation (LOQ): Comparison of the Empirical and the Statistical Methods Exemplified with GC-MS Assays of Abused Drugs. Clin. Chem. 1994, 40, 1233–1238. [Google Scholar] [CrossRef]
- Rower, J.E.; Bushman, L.R.; Hammond, K.P.; Kadam, R.S.; Aquilante, C.L. Validation of an LC/MS Method for the Determination of Gemfibrozil in Human Plasma and Its Application to a Pharmacokinetic Study. Biomed. Chromatogr. 2010, 24, 1300–1308. [Google Scholar] [CrossRef]
- van Amsterdam, P.; Companjen, A.; Brudny-Kloeppel, M.; Golob, M.; Luedtke, S.; Timmerman, P. The European Bioanalysis Forum Community’s Evaluation, Interpretation and Implementation of the European Medicines Agency Guideline on Bioanalytical Method Validation. Bioanalysis 2013, 5, 645–659. [Google Scholar] [CrossRef]
- Khan, A.; Shafiq, I.; Hassan Shah, M. Prevalence of Vitamin B12 Deficiency in Patients with Type II Diabetes Mellitus on Metformin: A Study from Khyber Pakhtunkhwa. Cureus 2017, 9, e1577. [Google Scholar] [CrossRef] [Green Version]
- Herrmann, W.; Obeid, R. Causes and Early Diagnosis of Vitamin B12 Deficiency. Dtsch. Arztebl. Int. 2008, 105, 680–685. [Google Scholar] [CrossRef]
- Johnson, W.E.; Li, C.; Rabinovic, A. Adjusting Batch Effects in Microarray Expression Data Using Empirical Bayes Methods. Biostatistics 2007, 8, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.A.; Lalitha, A.; Pavithra, D.; Padmavathi, I.J.N.; Ganeshan, M.; Rao, K.R.; Venu, L.; Balakrishna, N.; Shanker, N.H.; Reddy, S.U.; et al. Maternal Dietary Folate And/or Vitamin B12 Restrictions Alter Body Composition (adiposity) and Lipid Metabolism in Wistar Rat Offspring. J. Nutr. Biochem. 2013, 24, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Quehenberger, O.; Armando, A.M.; Brown, A.H.; Milne, S.B.; Myers, D.S.; Merrill, A.H.; Bandyopadhyay, S.; Jones, K.N.; Kelly, S.; Shaner, R.L.; et al. Lipidomics Reveals a Remarkable Diversity of Lipids in Human Plasma. J. Lipid Res. 2010, 51, 3299–3305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallego, S.F.; Højlund, K.; Ejsing, C.S. Easy, Fast, and Reproducible Quantification of Cholesterol and Other Lipids in Human Plasma by Combined High Resolution MSX and FTMS Analysis. J. Am. Soc. Mass Spectrom. 2018, 29, 34–41. [Google Scholar] [CrossRef]
- Marshall, D.L.; Criscuolo, A.; Young, R.S.E.; Poad, B.L.J.; Zeller, M.; Reid, G.E.; Mitchell, T.W.; Blanksby, S.J. Mapping Unsaturation in Human Plasma Lipids by Data-Independent Ozone-Induced Dissociation. J. Am. Soc. Mass Spectrom. 2019, 30, 1621–1630. [Google Scholar] [CrossRef] [PubMed]
- Medina, J.; van der Velpen, V.; Teav, T.; Guitton, Y.; Gallart-Ayala, H.; Ivanisevic, J. Single-Step Extraction Coupled with Targeted HILIC-MS/MS Approach for Comprehensive Analysis of Human Plasma Lipidome and Polar Metabolome. Metabolites 2020, 10, 495. [Google Scholar] [CrossRef] [PubMed]
- Rampler, E.; Schoeny, H.; Mitic, B.M.; El Abiead, Y.; Schwaiger, M.; Koellensperger, G. Simultaneous Non-Polar and Polar Lipid Analysis by on-Line Combination of HILIC, RP and High Resolution MS. Analyst 2018, 143, 1250–1258. [Google Scholar] [CrossRef] [Green Version]
- Schoeny, H.; Rampler, E.; Hermann, G.; Grienke, U.; Rollinger, J.M.; Koellensperger, G. Preparative Supercritical Fluid Chromatography for Lipid Class Fractionation-a Novel Strategy in High-Resolution Mass Spectrometry Based Lipidomics. Anal. Bioanal. Chem. 2020, 412, 2365–2374. [Google Scholar] [CrossRef] [Green Version]
- Buszewski, B.; Noga, S. Hydrophilic Interaction Liquid Chromatography (HILIC)—A Powerful Separation Technique. Anal. Bioanal. Chem. 2012, 402, 231–247. [Google Scholar] [CrossRef] [Green Version]
- Lange, M.; Fedorova, M. Evaluation of Lipid Quantification Accuracy Using HILIC and RPLC MS on the Example of NIST® SRM® 1950 Metabolites in Human Plasma. Anal. Bioanal. Chem. 2020, 412, 3573–3584. [Google Scholar] [CrossRef] [Green Version]
- Hines, K.M.; Herron, J.; Xu, L. Assessment of Altered Lipid Homeostasis by HILIC-Ion Mobility-Mass Spectrometry-Based Lipidomics. J. Lipid Res. 2017, 58, 809–819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berdeaux, O.; Juaneda, P.; Martine, L.; Cabaret, S.; Bretillon, L.; Acar, N. Identification and Quantification of Phosphatidylcholines Containing Very-Long-Chain Polyunsaturated Fatty Acid in Bovine and Human Retina Using Liquid Chromatography/tandem Mass Spectrometry. J. Chromatogr. A 2010, 1217, 7738–7748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, B.; Song, C.; Li, L.; Wang, M.; Jia, S.; Li, S.; Du, Z.; Ding, X.; Jiang, H. Qualitative Distribution of Endogenous Phosphatidylcholine and Sphingomyelin in Serum Using LC-MS/MS Based Profiling. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2020, 1155, 122289. [Google Scholar] [CrossRef] [PubMed]
- Baba, T.; Campbell, J.L.; Le Blanc, J.C.Y.; Baker, P.R.S. In-Depth Sphingomyelin Characterization Using Electron Impact Excitation of Ions from Organics and Mass Spectrometry. J. Lipid Res. 2016, 57, 858–867. [Google Scholar] [CrossRef] [Green Version]
- Tsugawa, H.; Ikeda, K.; Tanaka, W.; Senoo, Y.; Arita, M.; Arita, M. Comprehensive Identification of Sphingolipid Species by in Silico Retention Time and Tandem Mass Spectral Library. J. Cheminform. 2017, 9, 19. [Google Scholar] [CrossRef] [Green Version]
- Home-Lipidomics-Standards-Initiative (LSI). Available online: https://lipidomics-standards-initiative.org (accessed on 15 March 2022).
- Baba, T.; Campbell, J.L.; Le Blanc, J.C.Y.; Baker, P.R.S. Structural Identification of Triacylglycerol Isomers Using Electron Impact Excitation of Ions from Organics (EIEIO). J. Lipid Res. 2016, 57, 2015–2027. [Google Scholar] [CrossRef] [Green Version]
- Cao, W.; Cheng, S.; Yang, J.; Feng, J.; Zhang, W.; Li, Z.; Chen, Q.; Xia, Y.; Ouyang, Z.; Ma, X. Large-Scale Lipid Analysis with C=C Location and Sn-Position Isomer Resolving Power. Nat. Commun. 2020, 11, 375. [Google Scholar] [CrossRef] [Green Version]
- Tabassum, R.; Rämö, J.T.; Ripatti, P.; Koskela, J.T.; Kurki, M.; Karjalainen, J.; Palta, P.; Hassan, S.; Nunez-Fontarnau, J.; Kiiskinen, T.T.J.; et al. Genetic Architecture of Human Plasma Lipidome and Its Link to Cardiovascular Disease. Nat. Commun. 2019, 10, 4329. [Google Scholar] [CrossRef] [Green Version]
- Triebl, A.; Burla, B.; Selvalatchmanan, J.; Oh, J.; Tan, S.H.; Chan, M.Y.; Mellet, N.A.; Meikle, P.J.; Torta, F.; Wenk, M.R. Shared Reference Materials Harmonize Lipidomics across MS-Based Detection Platforms and Laboratories. J. Lipid Res. 2020, 61, 105–115. [Google Scholar] [CrossRef] [Green Version]
- Wolrab, D.; Chocholoušková, M.; Jirásko, R.; Peterka, O.; Holčapek, M. Validation of Lipidomic Analysis of Human Plasma and Serum by Supercritical Fluid Chromatography-Mass Spectrometry and Hydrophilic Interaction Liquid Chromatography-Mass Spectrometry. Anal. Bioanal. Chem. 2020, 412, 2375–2388. [Google Scholar] [CrossRef]
- Saraswathy, K.N.; Joshi, S.; Yadav, S.; Garg, P.R. Metabolic Distress in Lipid & One Carbon Metabolic Pathway through Low Vitamin B-12: A Population Based Study from North India. Lipids Health Dis. 2018, 17, 96. [Google Scholar] [PubMed] [Green Version]
- Khaire, A.; Rathod, R.; Kale, A.; Joshi, S. Vitamin B12 and Omega-3 Fatty Acids Together Regulate Lipid Metabolism in Wistar Rats. Prostaglandins Leukot. Essent. Fat. Acids 2015, 99, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, A.; Dangat, K.; Kale, A.; Sable, P.; Chavan-Gautam, P.; Joshi, S. Effects of Altered Maternal Folic Acid, Vitamin B12 and Docosahexaenoic Acid on Placental Global DNA Methylation Patterns in Wistar Rats. PLoS ONE 2011, 6, e17706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, S.; Kale, A.; Dangat, K.; Sable, P.; Kulkarni, A.; Joshi, S. Maternal Micronutrients (folic Acid and Vitamin B(12)) and Omega 3 Fatty Acids: Implications for Neurodevelopmental Risk in the Rat Offspring. Brain Dev. 2012, 34, 64–71. [Google Scholar] [CrossRef]
- Adaikalakoteswari, A.; Jayashri, R.; Sukumar, N.; Venkataraman, H.; Pradeepa, R.; Gokulakrishnan, K.; Anjana, R.M.; McTernan, P.G.; Tripathi, G.; Patel, V.; et al. Vitamin B12 Deficiency Is Associated with Adverse Lipid Profile in Europeans and Indians with Type 2 Diabetes. Cardiovasc. Diabetol. 2014, 13, 129. [Google Scholar] [CrossRef] [Green Version]
- Kumar, J.; Garg, G.; Sundaramoorthy, E.; Prasad, P.V.; Karthikeyan, G.; Ramakrishnan, L.; Ghosh, S.; Sengupta, S. Vitamin B12 Deficiency Is Associated with Coronary Artery Disease in an Indian Population. Clin. Chem. Lab. Med. 2009, 47, 334–338. [Google Scholar] [CrossRef]
- Mahalle, N.; Kulkarni, M.V.; Garg, M.K.; Naik, S.S. Vitamin B12 Deficiency and Hyperhomocysteinemia as Correlates of Cardiovascular Risk Factors in Indian Subjects with Coronary Artery Disease. J. Cardiol. 2013, 61, 289–294. [Google Scholar] [CrossRef] [Green Version]
- Toufektsian, M.-C.; Salen, P.; Laporte, F.; Tonelli, C.; de Lorgeril, M. Dietary Flavonoids Increase Plasma Very Long-Chain (n-3) Fatty Acids in Rats. J. Nutr. 2011, 141, 37–41. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lipid Class | Ion Mode | Number of Lipid Species in MRM Library | Internal Standard | Q1 | Q3 | Retention Time | DP | EP | CE | CxP | LoD Conc. (pmol/L) | LoQ Conc. (pmol/L) | Coefficient of Determination (R2) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SM | ESI+ | 12 | SM (d18:1–18:1(d9)) | 738.7 | 184.2 | 11.89 | 80 | 10 | 43 | 15 | 0.319 | 0.639 | 0.99 |
| CE | ESI+ | 21 | Ceramide (17:0) | 552.5 | 264.3 | 2.58 | 80 | 10 | 43 | 15 | 6.082 | 12.164 | 0.99 |
| Cer | ESI+ | 62 | |||||||||||
| TAG | ESI+ | 445 | TAG (15:0–18:1(d7)–15:0) | 829.4 | 570.5 | 2.39 | 80 | 10 | 38 | 15 | 17.233 | 34.466 | 0.99 |
| DAG | ESI+ | 50 | DAG (15:0–18:1(d7)) | 605.5 | 346.3 | 2.46 | 80 | 10 | 25 | 15 | 999.184 | 1998.367 | 0.99 |
| MAG | ESI+ | 17 | |||||||||||
| LPC | ESI− | 16 | LPC (18:1(d7)) | 587.4 | 288.3 | 12.63 | −80 | −10 | −50 | −15 | 0.368 | 5.887 | 0.99 |
| PC | ESI− | 79 | PC (15:0–18:1(d7)) | 811.6 | 288.3 | 9.73 | −80 | −10 | −50 | −15 | 13.024 | 26.048 | 0.98 |
| LPE | ESI− | 16 | LPE (18:1(d7)) | 485.3 | 288.3 | 13.09 | −80 | −10 | −50 | −15 | 1.329 | 5.318 | 0.99 |
| PE | ESI− | 142 | PE (15:0–18:1(d7)) | 709.6 | 288.3 | 10.61 | −80 | −10 | −50 | −15 | 0.245 | 0.979 | 0.99 |
| LPG | ESI− | 16 | PG (15:0–18:1(d7)) | 740.5 | 288.3 | 6.59 | −80 | −10 | −50 | −15 | 0.291 | 0.291 | 0.99 |
| PG | ESI− | 78 | |||||||||||
| LPI | ESI− | 16 | PI (15:0–18:1(d7)) | 828.6 | 288.3 | 13.09 | −80 | −10 | −50 | −15 | 2.639 | 10.557 | 0.98 |
| PI | ESI− | 77 | |||||||||||
| LPS | ESI− | 16 | PS (15:0–18:1(d7)) | 753.5 | 288.3 | 9.78 | −80 | −10 | −50 | −15 | 41.961 | 167.846 | 0.99 |
| PS | ESI− | 78 | |||||||||||
| PA | ESI− | 77 | PA (15:0–18:1(d7)) | 666.5 | 288.3 | 11.85 | −80 | −10 | −50 | −15 | 41.897 | 167.587 | 0.97 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhaskar, A.K.; Naushin, S.; Ray, A.; Singh, P.; Raj, A.; Pradhan, S.; Adlakha, K.; Siddiqua, T.J.; Malakar, D.; Dash, D.; et al. A High Throughput Lipidomics Method Using Scheduled Multiple Reaction Monitoring. Biomolecules 2022, 12, 709. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12050709
Bhaskar AK, Naushin S, Ray A, Singh P, Raj A, Pradhan S, Adlakha K, Siddiqua TJ, Malakar D, Dash D, et al. A High Throughput Lipidomics Method Using Scheduled Multiple Reaction Monitoring. Biomolecules. 2022; 12(5):709. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12050709
Chicago/Turabian StyleBhaskar, Akash Kumar, Salwa Naushin, Arjun Ray, Praveen Singh, Anurag Raj, Shalini Pradhan, Khushboo Adlakha, Towfida Jahan Siddiqua, Dipankar Malakar, Debasis Dash, and et al. 2022. "A High Throughput Lipidomics Method Using Scheduled Multiple Reaction Monitoring" Biomolecules 12, no. 5: 709. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12050709